Emergence of artemisinin-based combination therapy resistance markers in Plasmodium falciparum from the Brazilian tri-border region of the Guiana Shield
Yanka E. A. R. Salazar, Antonio M. Rezende, Maria C. S. B. Puça, Jaime Louzada, Maria E. P. Mascarenhas, Sonja Lagström, Danielle Fletcher, Joseli Oliveira-Ferreira, José P. Gil, Tais N. de Sousa

TL;DR
The study finds emerging drug resistance markers in malaria parasites in northern Brazil, suggesting the need for genomic surveillance to prevent spread.
Contribution
First report of the pfpm2/3 hybrid and pfmdr1-NFD haplotype in Brazil, highlighting new resistance markers.
Findings
The pfmdr1-NFD haplotype was found in 6% of isolates, likely from Venezuela.
The pfpm2/3 hybrid was detected in 4% of isolates, a first in Brazil.
pfcrt-C350R was present in 7% of isolates, indicating reduced drug susceptibility.
Abstract
Plasmodium falciparum malaria remains a public health concern in the Brazilian Amazon, particularly among mobile and remote populations. Antimalarial drug resistance threatens elimination efforts, particularly in the Guiana Shield, which has historically been linked to the spread of resistant parasites. We analyzed 99 P. falciparum isolates collected in Roraima State, northern Brazil (2016–2020), using targeted deep sequencing. Variants were assessed in pfcrt, pfmdr1, and pfk13, as well as copy number variation in pfpm2 and pfpm3, including detection of the pfpm2/3 hybrid. High coverage enabled the detection of both dominant and minority alleles. Among the 99 isolates, the pfmdr1-NFD haplotype was found in 6% (6/99, all likely originating from Venezuela), pfcrt-C350R in 7% (7/99), and pfpm2 and pfpm3 amplifications in 5% (5/94) and 6% (6/97), respectively, and the pfpm2/3 hybrid in 4%…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
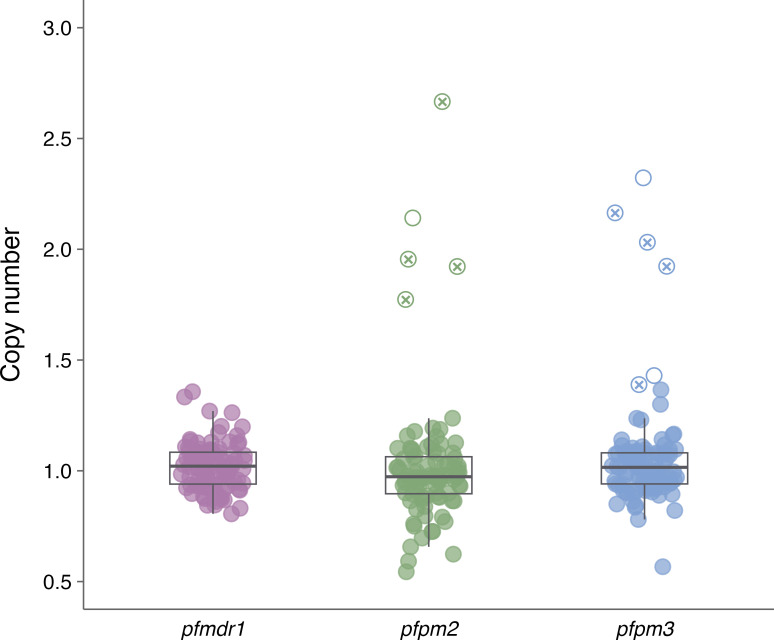 Fig 1
Fig 1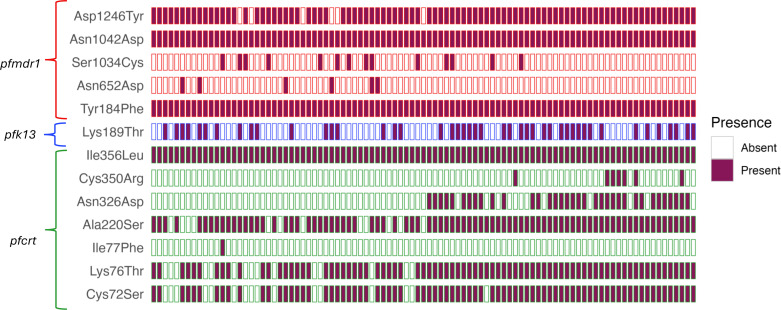 Fig 2
Fig 2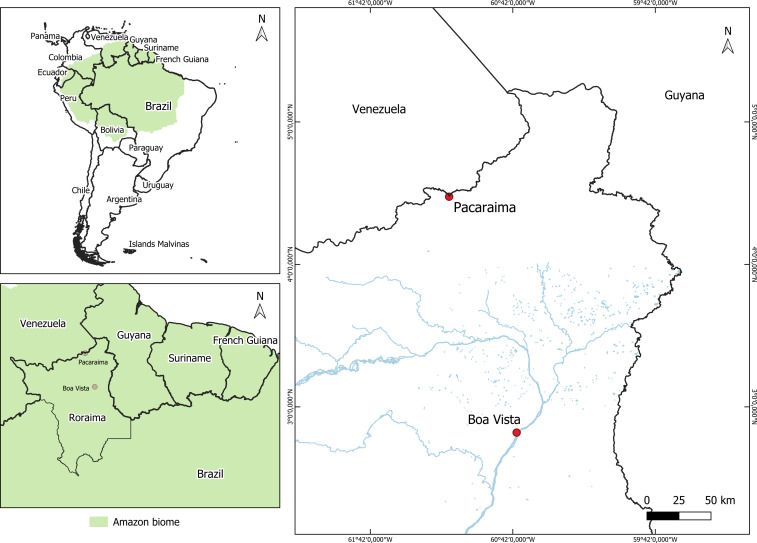 Fig 3
Fig 3| Characteristics | Total population |
|---|---|
| Age (years), median (interquartile range) | 31 (25–40) |
| Gender, | |
| Male | 68 (68.7) |
| Female | 31 (31.3) |
| Occupation, | |
| Gold mining | 86 (86.9) |
| Others | 13 (13.1) |
| Country of infection, | |
| Brazil | 13 (13.1) |
| French Guiana | 1 (1.0) |
| Guyana | 8 (8.0) |
| Suriname | 1 (1.0) |
| Venezuela | 76 (76.7) |
| Malaria recurrence episodes, | |
| No recurrence | 93 (94) |
| Recurrence | 6 (6) |
- —Swedish Research Councilhttp://dx.doi.org/10.13039/501100004359
- —Fundação de Amparo à Pesquisa do Estado de Minas Geraishttp://dx.doi.org/10.13039/501100004901
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Pharmaceutical Quality and Counterfeiting · Trypanosoma species research and implications
INTRODUCTION
In Brazil, the incidence of Plasmodium falciparum malaria has significantly decreased since the introduction of artemisinin-based combination therapies (ACTs), particularly artemether-lumefantrine (AL) as the first-line treatment (1). However, imported P. falciparum cases have risen in recent years, particularly among mobile and vulnerable populations in the Brazilian Amazon, such as Indigenous people and gold miners (2). In the Yanomami territory, one of the largest Indigenous groups in the Amazon, malaria has sharply increased in association with the expansion of illegal gold mining, aggravating health inequities and creating conditions for uncontrolled transmission (3). From 2011 to 2023, 418,185 malaria cases were reported in Indigenous villages (25.4% P. falciparum + mixed infections) and 152,884 in gold mining areas (4). These trends represent a setback for Brazil’s elimination efforts, especially given the intensified cross-border mobility that facilitates the introduction and spread of drug-resistant parasites (5).
Antimalarial drug resistance, including partial resistance to artemisinin (ART) and resistance to partner drugs, is an emerging concern that demands strengthened surveillance and timely interventions. While AL remains the first-line treatment in Brazil and other South American countries, dihydroartemisinin–piperaquine (DHA–PPQ) was occasionally used in French Guiana and is widely self-administered in illegal mining sites through products such as Artecom (dihydroartemisinin + piperaquine [PPQ] + trimethoprim) (6, 7). These drugs are often taken without supervision or correct dosing, contributing to selective pressure and the risk of resistance parasite emergence (6). Mutations in the propeller domain of the pfkelch13 gene are the current primary markers of partial resistance to ART, although additional markers or combinations may also contribute (8). Although these mutations have not been detected in Brazil, they have been reported in neighboring countries within the Guiana Shield, raising concerns about cross-border spread in areas of intense mobility (9, 10).
Reduced susceptibility to ACT partner drugs has been associated with treatment failures, including artesunate–mefloquine and DHA–PPQ in Southeast Asia (11). More recently, higher tolerance to AL has been documented in parts of Africa (12, 13). Despite this, lumefantrine (LUM) remains a key component of first-line therapies. LUM response is influenced by polymorphisms in the pfmdr1 gene, especially the NFD haplotype (N86, 184F, and D1246), which is selected under AL pressure and modulates drug transport activity (14). This effect is amplified by increased pfmdr1 copy number (pfmdr1 × N) (13). This phenomenon is partly explained by the function of the pfmdr1 product, P-glycoprotein homolog, which is inserted in the membrane of the digestive vacuole (DV) in an inside-out position. There, it exports substrates, including antimalarial drugs, into the DV, thereby reducing their cytoplasmic concentration and contributing to resistance phenotypes, particularly against non-DV centric drugs (15).
By contrast, PPQ acts similarly to chloroquine (CQ), targeting the digestive vacuole and inhibiting heme detoxification, though its mechanism of action is not fully understood (16, 17). Resistance to PPQ is primarily driven by pfcrt variants, with modulation by increased copy number of the plasmepsin II and III genes (pfpm2 and pfpm3), which encode digestive vacuole proteases. Among these, the South America-specific pfcrt-C350R mutation has been particularly implicated (6, 18). Moreover, recent findings in Africa have shown that the I356T mutation in pfcrt undergoes positive selection, co-occurring with pfpm3 amplifications after repeated treatment cycles, which reinforces the contribution of this SNP in reducing parasite susceptibility to PPQ (19).
To address the threat of emerging multidrug resistance, we investigated the genetic profile of P. falciparum isolates from the Guiana Shield, a region historically recognized as a hotspot for the emergence of antimalarial drug resistance in the Americas (6, 20). This highly dynamic border area experiences intense malaria transmission, mainly driven by a high-risk, mobile population engaged in gold mining activities. At the same time, malaria transmission in the Amazon is markedly heterogeneous, with intense outbreaks occurring alongside areas of lower endemicity and limited acquired immunity. In such contexts, resistant genotypes face little competition and can rapidly expand and become fixed through clonal sweeps, particularly under sustained drug pressure (21). Our findings provide a baseline for molecular surveillance and inform cross-border strategies for malaria control. A major strength of this study is the use of targeted deep sequencing, a high-resolution approach, to detect low-frequency resistant variants that are often missed by conventional methods, thereby reinforcing its value in complex transmission settings.
RESULTS
Characteristics of participants and epidemiological context
A total of 99 participants were included in this study, with a median age of 31 years (interquartile range: 25–40). Most individuals were male (68.7%), and the majority of infections were likely acquired in Venezuela (76.7%), particularly in mining areas, according to travel histories. Sample collection sites included Pacaraima (n = 36, 36.3%) and Boa Vista (n = 63, 63.6%). Although most isolates originated from Venezuela, additional samples likely represent infections from Brazil, French Guiana, Guyana, and Suriname (Table 1).
Prevalence of pfpm2/3 amplification and absence of pfmdr1 CNV
Copy number variation (CNV) analysis was successful for 97 (98%) samples for pfmdr1 and pfpm3 and 94 (95%) samples for pfpm2. No pfmdr1 amplifications were detected when applying the conservative >1.4 threshold, corresponding to an estimated minimum of 25% of parasites carrying the duplication within an infection (13). Although no isolates reached this cutoff, a few showed values between 1.3 and 1.4, potentially reflecting low-frequency multicopy variants in polyclonal infections. Thus, we cannot rule out the presence of minor pfmdr1 × N subpopulations below the adopted cutoff.
In contrast, pfpm2/3 amplifications were detected in 5% of samples (n = 5) for pfpm2 and 6% (n = 6) for pfpm3 (Fig. 1). Notably, 4% of the samples (n = 4) showed simultaneous amplification of both genes. Additionally, a breakpoint assay targeting the hybrid pfpm2/3 gene structure detected this variant in the same 4% (n = 4) of isolates, indicating the presence of this structural rearrangement.
Copy number variation of pfmdr1, pfpm2, and pfpm3 in Plasmodium falciparum isolates. Only samples with relative quantification above 1.4 were considered amplified. Each point represents an individual isolate, with color indicating the corresponding gene. Filled circles represent isolates without copy number amplification, while empty circles indicate amplified copies, and circles marked with x represent a hybrid sequence of pfpm2/3. Boxplots show the median (line), interquartile range (box), and values within 1.5× the interquartile range (whiskers).
Next-generation sequencing performance and data reliability for key resistance genes
Given the complexity of polyclonal infections, we employed a high-sensitivity targeted sequencing approach to detect minor parasite subpopulations and assessed the quality of next-generation sequencing (NGS) data across resistance-associated genes to ensure reliable variant analysis. Sequencing performance was robust, with a mean coverage of 94.7% for the panel of target genes. We herein focused on key genes of clinical and epidemiological importance, pfcrt, pfk13, and pfmdr1 genes, which consistently showed high and uniform coverage, supporting the reliability of variant detection. A minimum depth threshold of 100× was established to ensure confident identification of low-frequency variants. The overall sequencing largely exceeded this threshold, averaging more than 1,000×. Notably, pfk13 reached a mean depth of over 2,000×, pfmdr1 945×, and pfcrt approximately 438×. A moderate positive correlation was observed between gene GC content and sequencing depth (r = 0.503, P < 0.001, 95% CI: 0.259–0.687), indicating that GC-rich regions tend to yield higher read counts. A weaker but still significant correlation was found between GC content and gene coverage (r = 0.375, P = 0.008, 95% CI: 0.104–0.593), suggesting a more limited influence on the breadth of coverage. These results confirm the suitability of the data set for high-confidence molecular surveillance of antimalarial resistance in P. falciparum isolates.
C350R variant and predominance of pfcrt SVMNT 72–76 haplotype
The pfcrt-C350R variant was detected in 7% (n = 7) of isolates, with a mean sequencing depth of 262× at this position. Three of these infections were acquired in Brazil, two in Venezuela, one in Guyana, and one in French Guiana. This mutation, previously associated with PPQ resistance, even in the absence of pfpm2/3 duplications, was not detected in our initial Sanger sequencing but was identified through targeted deep sequencing, which enabled the detection of minority variants.
Classical pfcrt polymorphisms at codons 72–76 confirmed the predominance of the SVMNT haplotype, present in most isolates (C72S in 83% [n = 79] and K76T in 84% [n = 80]), whereas the wild-type CVMNK haplotype was detected in 15 isolates (16%). In addition, we identified the 7G8 haplotype in 33 isolates (35%), composed of SVMNT, A220S, N326D, and I356L. This haplotype is widely distributed across South America and confers high resistance to CQ and amodiaquine while imposing minimal fitness cost to the parasite (22).
Regarding the pfk13 gene, no validated or candidate mutations linked to ART partial resistance were detected. The K189T polymorphism was observed in 47% of our data set (n = 45); although not considered a validated or candidate marker, it has been reported in other studies (23) and is noted here for completeness.
Detection of pfmdr1 NFD haplotype associated with reduced lumefantrine susceptibility
Analysis of pfmdr1 revealed that the most prevalent variants were Y184F (100%, n = 95), N1042D (100%, n = 95), and D1246Y (94%, n = 89), with additional mutations observed at S1034C (15%, n = 14) and N652D (6%, n = 6). The wild-type N86 allele, combined with 184F and D1246, defined the NFD haplotype, which was identified in 6% (n = 6) of isolates (Fig. 2). This haplotype has been associated with reduced lumefantrine susceptibility and selection under AL treatment pressure (14). NGS detected five additional isolates carrying 184F that were not identified by Sanger sequencing, consistent with the increased sensitivity of deep sequencing for mixed infections.
Presence/absence matrix of variants identified in the pfcrt, pfk13, and pfmdr1 genes through NGS. The y-axis shows the variants detected in each gene of interest, indicated by color: red, pfmdr1; blue, pfk13; and green, pfcrt. The x-axis represents the analyzed isolates (n = 95). Only variants with a frequency ≥10% of the reads analyzed were considered.
Recurrence analysis
No associations were found between drug resistance markers and recurrence episodes. However, this analysis was limited by the small number of P. falciparum-related recurrence cases (n = 6). Recurrence was defined based on cases reported in the SIVEP-Malaria system up to 1 year before and 1 year after the date of inclusion in the study. The high proportion of imported infections, mainly from Venezuela, may also have affected recurrence estimates, as some participants likely returned to their country of origin and were not captured by the SIVEP-Malaria system.
DISCUSSION
The molecular characterization of P. falciparum isolates from the Guiana Shield region revealed highly relevant genetic variants with direct implications for antimalarial treatment efficacy in the Amazon. Although many infections were likely imported from neighboring countries, these markers were identified in patients treated in Brazil, underscoring the importance of surveillance in cross-border settings where malaria transmission is shaped by migration, informal drug use, and limited healthcare access. A central concern is the potential introduction and spread of resistant parasites into vulnerable groups, particularly Indigenous populations in remote areas. The Brazil–Venezuela–Guyana tri-border is a hotspot of mobility, driven by gold mining and migration, and overlaps with Indigenous territories. In Roraima, the Yanomami territory has been disproportionately affected, with a sharp increase in malaria linked to illegal mining and poor access to health services (3). In such contexts of heterogeneous transmission and limited immunity, resistant parasites may spread silently and hinder elimination strategies (24).
We identified gene amplifications of pfpm2 (5%) and pfpm3 (6%), with 4% of the isolates carrying both. These markers are confirmed determinants of PPQ resistance and have been associated with treatment failures (25). We applied a conservative 1.4 threshold to define gene amplification, based on recent studies suggesting that values above 1.3 may already indicate multicopy subpopulations in polyclonal infections (13, 19). This approach enhances our ability to detect emerging parasite variants that may still be clinically relevant. Although PPQ is not officially used in Brazil, its informal use among gold miners in remote Amazonian areas (2, 6) could impose selective pressure and facilitate the spread of parasites with reduced susceptibility.
We also detected the pfpm2/3 hybrid sequence in 4% of the isolates, marking the first report of this structural variant in Brazil. Previously described in Southeast Asia, this hybrid arises from a conserved genomic breakpoint between the two plasmepsin genes and has been linked to full PPQ resistance, independent of pfcrt mutations (26). While pfpm2 amplification alone has been associated with treatment failure and in vitro resistance (26, 27), the hybrid sequence provides an additional, more specific marker for molecular surveillance (28). The detection of this variant in Brazilian isolates raises concerns about cross-continental introduction through migratory flows, especially given its widespread circulation in Southeast Asia and the increasing movement of individuals between malaria-endemic regions (26, 29–31). Prior studies have already documented similar patterns in the spread of antimalarial resistance, underscoring the risk of global dissemination of resistant genotypes (32–34).
The pfcrt-C350R variant, a validated PPQ resistance marker, was found in 7% of isolates, a higher prevalence than previously reported in Brazil (6), suggesting a possible ongoing expansion of this variant. Of these, four were likely imported and three were acquired in Brazil, indicating that local circulation cannot be excluded. Importantly, the variant was identified only through targeted deep sequencing, reinforcing the value of high-resolution tools to capture minority variants. Given that C350R has been associated with reduced PPQ susceptibility even in the absence of plasmepsin duplications (6), its presence serves as a strong early indicator of shifting drug response profiles. The combination of multiple PPQ-associated markers increases concern that parasites could, over time, accumulate sets of resistance variants.
No validated or candidate pfk13 mutations associated with partial resistance to ART were detected, consistent with previous studies in Brazil (35). However, the recent detection of C580Y and G718S in neighboring countries (9, 10) highlights the risk of future introduction into Brazil, given high cross-border mobility.
No confirmed pfmdr1 amplifications were found above the >1.4 threshold, though some isolates showed intermediate values (1.3–1.4) suggestive of low-frequency multicopy subpopulations. In addition, we detected the *pfmdr1-*NFD haplotype in six isolates, all from patients with infections acquired in Venezuela. This haplotype has been increasingly reported in other endemic regions, particularly in parts of West Africa where AL is widely used as first-line therapy (15, 27, 36, 37), but its role as a validated resistance marker remains uncertain. The detection of both CNV signals and the NFD haplotype highlights the importance of continued monitoring of LUM efficacy in the region. Supporting this, experimental evidence suggests that parasites with the NFD haplotype in a background of increased pfmdr1 copy number may exhibit enhanced transporter activity, potentially leading to increased drug efflux and reduced drug efficacy (15). A comparable scenario has been reported in Angola, where isolates harboring both variants were associated with declining treatment efficacy of AL (13). These observations raise the possibility that South America could follow a similar trajectory, particularly if AL continues to be used extensively without adequate surveillance to track and contain resistant strains.
This study provides the first report in Brazil of the pfpm2/3 hybrid sequence and the pfmdr1-NFD haplotype, variants linked to reduced susceptibility to PPQ and LUM, respectively. These findings expand our understanding of the diversity and geographic distribution of resistance-associated markers in P. falciparum across South America, pointing to the advantages of high-resolution sequencing in detecting minority variants. Beyond their molecular significance, the detection of these variants in a highly mobile tri-border setting, overlapping with Indigenous territories heavily impacted by malaria, reinforces the urgency of coordinated regional surveillance. Overall, our results support genomic surveillance as an essential tool to anticipate emerging resistance, inform locally adapted interventions, and safeguard the efficacy of current antimalarial therapies.
MATERIALS AND METHODS
Study site and sample collection
This study was conducted in Roraima state, northern Brazil, with sample collection in the municipalities of Boa Vista and Pacaraima, between March 2016 and March 2020. These sites, located near the Venezuela border, are strategic for malaria surveillance due to high cross-border mobility driven by migration and informal mining (Fig. 3) (38). A total of 99 P. falciparum-positive samples were included from a previously conducted cross-sectional study in the region (38). Samples with higher parasite densities were prioritized to ensure sufficient DNA quality and quantity for downstream molecular analyses. Participants were symptomatic, aged ≥16 years, non-pregnant, and without signs of severe malaria. The initial diagnosis was based on microscopy, with species confirmation by quantitative PCR (qPCR). Additional methodological details are provided in the Supplementary Material.
Location of study sites in the Amazon border region of Roraima, Brazil. The map highlights the municipalities of Boa Vista and Pacaraima in Roraima State, northern Brazil. These sites are located near international borders with Venezuela and Guyana, in a region marked by intense cross-border mobility, including migratory flows associated with informal gold mining. The Amazon biome is shaded in green, and red dots identify study sites. This border region plays a critical role in the importation and transmission dynamics of Plasmodium falciparum in the Brazilian Amazon.
Detection of pfmdr1 and pfcrt SNPs
Polymorphisms in pfcrt (K76T, N326S, I356T, and C350R) and pfmdr1 (N86Y, Y184F, and D1246Y) were initially screened using PCR-RFLP and Sanger sequencing, following established protocols (6, 13). These conventional assays served as preliminary analyses to validate marker presence and provided reference points for subsequent NGS. The pfmdr1-NFD haplotype was defined by the combination of N86, 184F, and D1246. All preliminary laboratory experiments were performed at Karolinska Institutet (Solna, Sweden). Additional methodological details are provided in the Supplemental material.
Molecular analysis of pfpm2, pfpm3, and pfmdr1 genes
The CNV of pfpm2 (PF3D7_1408000), pfpm3 (PF3D7_1408100), and pfmdr1 (PF3D7_0523000) was assessed by qPCR, using β-tubulin as an internal reference gene. A multiplex qPCR approach was used for pfpm3 and pfmdr1 (13), while pfpm2 CNV followed the method by Pernaute-Lau and colleagues (19). Reference controls included 3D7 (single copy), Dd2 (pfmdr1 duplication), and NHP1034 (pfpm2/3 duplication). The pfpm2/3 hybrid was detected using a validated PCR-based breakpoint assay (28). Relative gene copy number was calculated using the ΔΔCt method, and a conservative threshold of >1.4 was applied to define gene amplification, acknowledging that values between 1.3 and 1.4 may represent minor multicopy subpopulations (13).
Selective whole genome amplification
All samples underwent selective whole genome amplification to enrich P. falciparum DNA before library preparation and NGS, following the MalariaGEN protocol v.2.0 and employed 10 previously validated primer pairs specific to the P. falciparum genome (39). Amplified products were purified using AMPure XP magnetic beads (Beckman Coulter), with 20 μL of input sample per reaction. Post-purification, DNA concentrations were reassessed using a fluorometric method (Qubit, Thermo Fisher Scientific) to verify the quantity, integrity, and overall suitability of the DNA for sequencing.
Targeted deep sequencing workflow and bioinformatic analysis
Genomic libraries were prepared using a custom hybrid-capture panel and sequenced on the Illumina NextSeq 2000 platform at the National Genomics Infrastructure, SciLifeLab (Stockholm, Sweden). Library construction included pre- and post-capture amplification steps. Sequencing generated ultra-deep coverage (mean >1,000 × across target genes), enabling reliable detection of both dominant and minor variants.
Raw reads underwent quality control, adapter trimming, and UMI processing using standard tools, including FastP and FastQC. High-quality reads were aligned to the P. falciparum 3D7 reference genome using BWA-MEM, and variant calling was performed with Mutect2 (GATK4), followed by rigorous filtering to retain only high-confidence variants. Identified variants were functionally annotated using SnpEff, and coverage metrics for target genes were calculated using BEDTools. While the overall proportion of polyclonal infections was not formally estimated, the ultra-deep coverage and allelic depth distributions allowed the confident identification of minor alleles. Alleles observed at lower frequencies were retained when supported by high read depth, providing strong evidence that they represented true minority variants within infections. A custom Python script was used to generate summary statistics and visual representations of read depth. Detailed protocols and parameter settings are provided in the Supplemental material.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ministério da Saúde. 2021. Guia de tratamento da malária no Brasil, 2nd ed. Brasília. Available from: www.bvsms.saude.gov.br
- 2Murta FLG, Marques LLG, Santos APC, Batista TSB, Mendes MO, Silva ED, Neto AVS, Fabiano M, Rodovalho SR, Monteiro WM, Lacerda MVG. 2021. Perceptions about malaria among Brazilian gold miners in an Amazonian border area: perspectives for malaria elimination strategies. Malar J 20:1–14. doi:10.1186/s 12936-021-03820-034174880 PMC 8236171 · doi ↗ · pubmed ↗
- 3Martins-Filho PR, Araújo FWC, Santos-Júnior LC, Santiago BM, Santos FHA, Araújo APD, Machado CEP, Lima S. 2024. The increase in cases and deaths from malaria in the Brazilian Yanomami territory is associated with the spread of illegal gold mining in the region: A 20-year ecological study. Travel Med Infect Dis 57:102686. doi:10.1016/j.tmaid.2023.10268638159876 · doi ↗ · pubmed ↗
- 4Garcia KKS, Soremekun S, Abrahão AA, Marchesini PB, Drakeley C, Ramalho WM, Siqueira AM. 2024. Is Brazil reaching malaria elimination? A time series analysis of malaria cases from 2011 to 2023. P Lo S Glob Public Health 4:e 0002845. doi:10.1371/journal.pgph.000284538295141 PMC 10830034 · doi ↗ · pubmed ↗
- 5Douine M, Lambert Y, Galindo MS, Mutricy L, Sanna A, Peterka C, Marchesini P, Hiwat H, Nacher M, Adenis A, Demar M, Musset L, Lazrek Y, Cairo H, Bordalo Miller J, Vreden S, Suarez-Mutis M. 2021. Self-diagnosis and self-treatment of malaria in hard-to-reach and mobile populations of the Amazon: results of Malakit, an international multicentric intervention research project. The Lancet Regional Health - Americas 4:100047. doi:10.1016/j.lana.2021.10004736776708 PMC 9903903 · doi ↗ · pubmed ↗
- 6Florimond C, de Laval F, Early AM, Sauthier S, Lazrek Y, Pelleau S, Monteiro WM, Agranier M, Taudon N, Morin F, Magris M, Lacerda MVG, Viana GMR, Herrera S, Adhin MR, Ferreira MU, Woodrow CJ, Awab GR, Cox H, Ade M-P, Mosnier E, Djossou F, Neafsey DE, Ringwald P, Musset L. 2024. Impact of piperaquine resistance in Plasmodium falciparum on malaria treatment effectiveness in The Guianas: a descriptive epidemiological study. Lancet Infect Dis 24:161–171. doi:10.1016/S 1473-3099(23)00502-937858325 PMC · doi ↗ · pubmed ↗
- 7Douine M, Lazrek Y, Blanchet D, Pelleau S, Chanlin R, Corlin F, Hureau L, Volney B, Hiwat H, Vreden S, Djossou F, Demar M, Nacher M, Musset L. 2018. Predictors of antimalarial self-medication in illegal gold miners in French Guiana: a pathway towards artemisinin resistance. J Antimicrob Chemother 73:231–239. doi:10.1093/jac/dkx 34329045645 · doi ↗ · pubmed ↗
- 8Ariey F, Witkowski B, Amaratunga C, Beghain J, Langlois AC, Khim N, Kim S, Duru V, Bouchier C, Ma L, et al.. 2014. A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature 505:50–55. doi:10.1038/nature 1287624352242 PMC 5007947 · doi ↗ · pubmed ↗