Taxonomy and phylogeny reveal new species of Volvariella (Volvariellaceae, Agaricales) and Volvopluteus (Pluteaceae, Agaricales) from eastern India

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
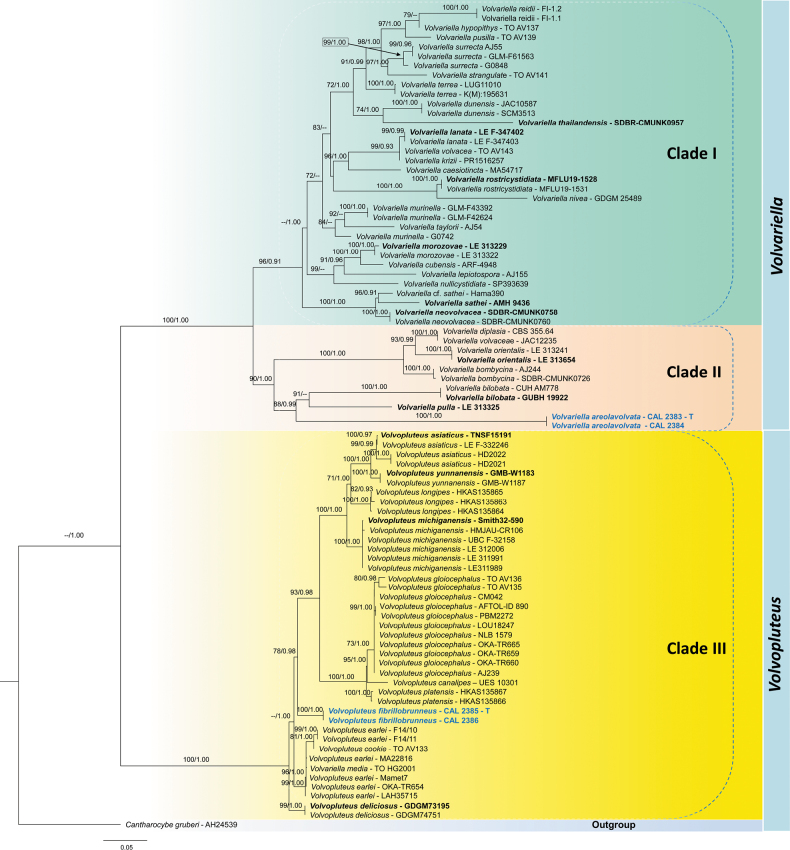 Figure 1
Figure 1 Figure 2
Figure 2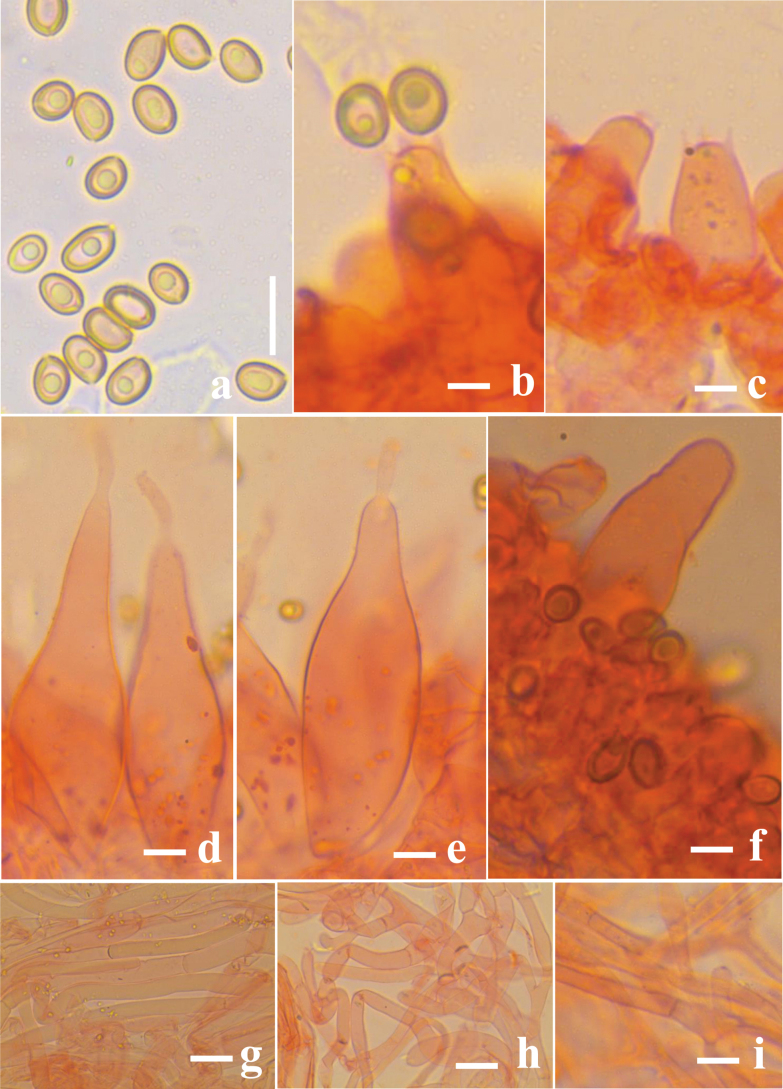 Figure 3
Figure 3 Figure 4
Figure 4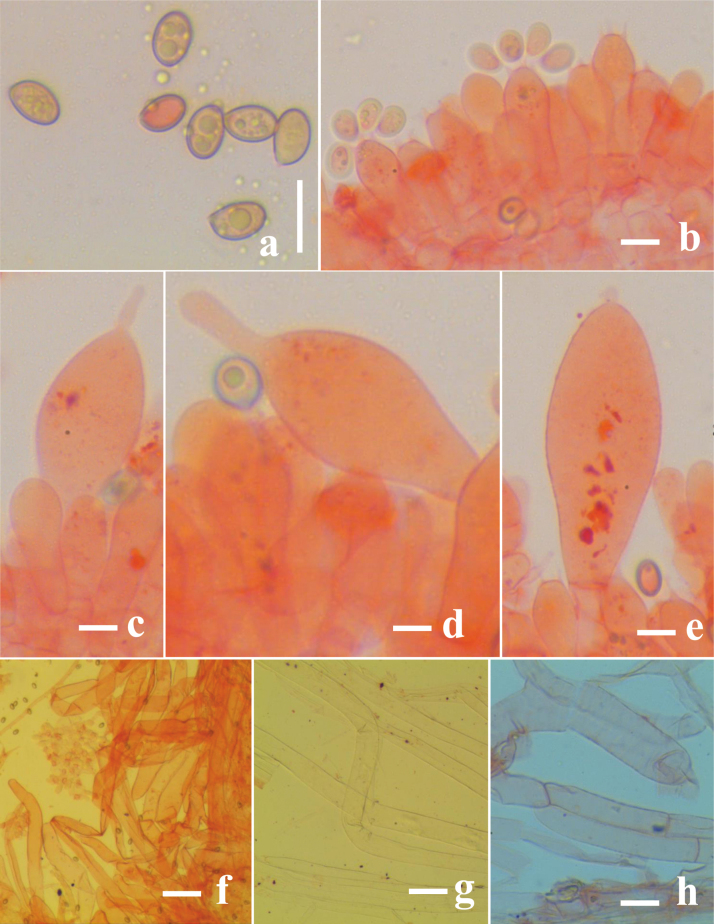 Figure 5
Figure 5| Species | State | Reference |
|---|---|---|
|
| Kerala |
|
|
| Kerala, Rajasthan | |
|
| West Bengal |
|
|
| West Bengal, Uttar Pradesh, | |
|
| Kerala |
|
|
| West Bengal, Uttar Pradesh | |
|
| West Bengal, Uttar Pradesh | |
|
| Rajasthan |
|
|
| Kerala | |
|
| Kerala, Rajasthan | |
|
| Punjab |
|
|
| Tripura |
|
|
| Assam |
|
|
| Kerala | |
|
| Kerala | |
|
| Kerala, West Bengal, Rajasthan | |
|
| Maharashtra |
|
|
| Kerala, Mizoram | |
|
| Kerala, West Bengal, Uttar Pradesh | |
|
| West Bengal, Uttar Pradesh | |
|
| Kerala, West Bengal, Assam, Tripura | |
|
| Maharashtra, Uttar Pradesh | |
| Punjab |
| |
| Punjab |
|
| Loci | PCR primers (forward/reverse) | Primer sequences | References |
|---|---|---|---|
| ITS | ITS1 | TCCGTAGGTGAACCTGCGG |
|
| ITS4 | TCCTCCGCTTATTGATATGC | ||
| LSU | LR0R | ACCCGCTGAACTTAAGC |
|
| LR3 | CCGTGTTTCAAGACGGG |
| Taxon name | Specimen voucher | Country of origin | GenBank accession no. | References | |
|---|---|---|---|---|---|
|
| nrLSU | ||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| CUH AM778 | India |
|
|
|
|
| GUBH 19922 | India |
|
|
|
|
| AJ244 | Spain |
|
|
|
|
| SDBR-CMUNK0726 | Thailand |
|
|
|
|
| MA54717 | Spain |
|
|
|
|
| Hama390 | Niger |
|
|
|
|
| ARF-4948 | USA |
|
| GenBank |
|
| CBS 355.64 | India |
|
|
|
|
| New Zealand |
|
| Unpublished | |
|
| SCM3513 | Spain |
|
|
|
|
| TO AV137 | Italy |
|
| Unpublished |
|
| PR1516257 | Czech Republic |
|
|
|
|
| LE F-347402 | Russia |
|
|
|
|
| LE F-347403 | Russia |
|
|
|
|
| AJ155 | USA |
|
|
|
|
| TO HG2001 | Italy |
|
|
|
|
| LE 313229 | Vietnam |
|
|
|
|
| LE 313322 | Vietnam |
|
|
|
|
| GLM-F43392 | Germany |
|
| Unpublished |
|
| GLM-F42624 | Germany |
|
| Unpublished |
|
| G0742 | Hungary |
|
| Varga et al. (2019) |
|
| SDBR-CMUNK0758 | Thailand |
|
|
|
|
| SDBR-CMUNK0760 | Thailand |
|
|
|
|
| GDGM 25489 | China |
|
|
|
|
| SP393639 | Brazil |
|
|
|
|
| LE 313241 | Vietnam |
|
|
|
|
| LE 313654 | Vietnam |
|
|
|
|
| LE 313325 | Vietnam |
|
|
|
|
| TO AV139 | Italy |
|
| Unpublished |
|
| FI-1.2 | Finland |
|
|
|
|
| FI-1.1 | Finland |
|
|
|
|
| MFLU19-1528 | Thailand |
|
|
|
|
| MFLU19-1531 | Thailand |
|
|
|
|
| AMH 9436 | India |
|
|
|
|
| TO AV141 | Italy |
|
| Unpublished |
|
| AJ55 | Spain |
|
|
|
|
| GLM-F61563 | Germany |
|
| Unpublished |
|
| G0848 | Hungary |
|
| Unpublished |
|
| AJ54 | Portugal |
|
|
|
|
| LUG11010 | France |
|
|
|
|
| K(M):195631 | United Kingdom |
|
| Unpublished |
|
| SDBR-CMUNK0957 | Thailand |
|
|
|
|
| New Zealand |
|
| Unpublished | |
|
| TO AV143 | Slovenia |
|
| Unpublished |
|
| TNSF15191 | Japan |
|
|
|
|
| LE F-332246 | Russia |
|
|
|
|
| HD2022 | China |
|
| Unpublished |
|
| HD2021 | China |
|
| Unpublished |
|
| UES 10301 | Mexico |
|
|
|
|
| TO AV133 | Italy |
|
|
|
|
| GDGM73195 | China |
|
|
|
|
| GDGM74751 | China |
|
|
|
|
| Pakistan |
|
| Unpublished | |
|
| OKA TR654 | Turkey |
|
| Unpublished |
|
| Mamet7 | Democratic Republic of Congo |
|
|
|
|
| MA22816 | Spain |
|
|
|
|
| F14/10 | Australia |
|
| Unpublished |
|
| F86/11 | Australia |
|
| Unpublished |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| TO AV136 | Italy |
|
|
|
|
| TO AV135 | Italy |
|
|
|
|
| AFTOL-ID 890 | USA |
|
|
|
|
| OKA-TR659 | Turkey |
|
| Unpublished |
|
| OKA-TR660 | Turkey |
|
| Unpublished |
|
| OKA TR665 | Turkey |
|
| Unpublished |
|
| AJ239 | Spain |
|
|
|
|
| CM042 | Algeria |
|
| Unpublished |
|
| LOU18247 | Spain |
|
|
|
|
| NLB 1579 | Australia |
|
| Unpublished |
|
| PBM2272 | USA |
|
|
|
|
| HKAS135864 | China |
|
|
|
|
| HKAS135865 | China |
|
|
|
|
| HKAS135863 | China |
|
|
|
|
| Smith32-590 | USA |
|
|
|
|
| LE 312006 | Russia |
|
|
|
|
| LE 311991 | Russia |
|
|
|
|
| HMJAU-CR106 | China |
|
|
|
|
| UBC F-32158 | Canada |
|
|
|
|
| Russia |
|
|
| |
|
| HKAS135867 | China |
|
|
|
|
| HKAS135866 | China |
|
|
|
|
| GMB-W1183 | China |
|
|
|
|
| GMB-W1187 | China |
|
|
|
|
| AH24539 | Spain |
|
|
|
| Name of the taxon | Pileus color | Pileus diameter (mm) | Stipe (mm) | Basidiospore (µm) | Basidia (µm) | Pleurocystidia (µm) | Cheilocystidia (µm) | Reference |
|---|---|---|---|---|---|---|---|---|
|
|
|
|
|
|
|
|
|
|
|
| Whitish to pale ochraceous | 70–190 | 70–190 × 7–20 | 6.5–9.5 × 4.0–6.5; Q = 1.20–1.70 | 20–30 × 8–11 | 40–100 × 10–20 | 50–110 × 10–25 |
|
|
| Entirely grayish brown pileus | 45–70 | 45–60 × 4–10 | 4.8–5.5 × 2.7–3.5; Q = 1.57–1.60 | 20–23 × 5–7 | 18.5–27.5 × 7–10 | 32–83 × 13–30 |
|
|
| Pileus grayish brown or ash brown | 40–75 | 55–70 × 3–5 | 5.5–7.0 × 4.0–5.0; Q = 1.20–1.59 | 15–22 × 7–8 | 52–83 × 14–34 | 45–75 × 14–23 |
|
| Name of the taxon | Pileus color | Pileus diameter (mm) | Stipe (mm) | Basidiospore (µm) | Basidia (µm) | Pleurocystidia (µm) | Cheilocystidia (µm) | Reference |
|---|---|---|---|---|---|---|---|---|
|
|
|
|
|
|
|
|
|
|
|
| White or ochraceous buff, light ochraceous at center | 25–45 | 35–65 × 3–7 | 11.0–16.0 × 7–9.5 | 30–47 × 10–18 | 47–98 × 14–20 | 25–50 × 10–35 |
|
|
| Surface usually grayish | 50–110 | 80–140 × 8–27 | 12.5–16.5 × 7.5–10 | 42–65 × 13–16 | 62–88 × 12–33 | 50–65 × 18–32 |
|
|
| Surface orange white | 41–59 | 36–85 × 4–6 | 11.2–16.0 × 7.2–14.4 | 25.6–46.4 × 9.6–17.6 | 20.8–86.4 × 8.8–34.0 | 32.0–80.0 × 9.6–40.0 |
|
|
| Slightly darker or pale yellowish biscuit in center | 20–50 | 40–70 × 5–10 | 12–15.5 × 7.3–9.5 | 35–51 × 10–14 | 31–85 × 15–38 | 30–75 × 11–24 |
|
|
| Uniformly grayish beige except darker brownish at the umbo | 30–110 | 35–120 × 5–22 | 12–17.5 × 7.5–10 | 42–56 × 12–16 | 45–60 × 20–40 | 32–70 × 10–25 |
|
|
| Slightly brownish at center, grayish white to dirty white | 48–70 | 130–150 × 4–10 | 14–20 × 8–11 | 35–60 × 16–23 | NR | 40–75 × 16–29 |
|
|
| Gray to brownish gray | 70–90 | 80–110 × 10–15 | 10.5–13.5 × 6.5–8 | 35–55 × 10–15 | 70–110 × 25–45 | 60–75 × 15–27 |
|
|
| Grayish white to dirty white, sometimes with a cream tinge | 30–45 | 50–80 × 4–10 | 15.5–20 × 9–12 | 35–68 × 15–20 | 65–95 × 18–40 | NR |
|
|
| Surface grayish orange, orange grey at center, with pinkish tinge towards margin | 10–112 | 70–165 × 18–25 | 11.2–16 × 7.2–12.8 | 17.6–34.0 × 8.9–16.0 | NR | NR |
|
|
| Pale grey when young, transitioning to white with grayish tints at maturity | 48–80 | 110–160 × 5–23 | 15.5–18.8 × 9.6–11.5 | 40.8–55 × 15.1–19.9 | 62–113.8 × 24.5–48.5 | 50.1–81.5 × 24.1–34.9 |
|
|
| Dark blonde center and birch grey sides | 59–85 | 120–137 × 11–13 | 11.3–12.9 × 6.0–6.9 | 43–54 × 10–15 | 60–82 × 16–25 | 60–86 × 14–27 |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Plant Pathogens and Fungal Diseases · Slime Mold and Myxomycetes Research
Introduction
The genus Volvariella Speg. was established by Spegazzini (1898) with Volvariella argentina Speg. as the type species. Members of the genus are widely distributed across temperate, subtropical, and tropical regions (Mehrotra and Aneja 1990). They are mostly found as saprobes on soil, in grasslands, on forest litter, and occasionally on decaying wood. Species of Volvariella are characterized by their pluteoid basidiomata, free lamellae, the presence of a membranous volva at the stipe base, a pinkish spore print, smooth and inamyloid basidiospores, and an inverse hymenophoral trama (Singer 1986; Orton 1986; Boekhout 1990; Kaygusuz et al. 2020; Kalichman et al. 2020; Kumla et al. 2022). However, in some studies, Volvariella was placed within the family Pluteaceae based on morphological similarities with the genus Pluteus (Justo et al. 2011a). Later, molecular phylogenetic studies showed that Volvariella s.l. is polyphyletic (Matheny et al. 2006).
Subsequent multi-gene phylogenetic analyses demonstrated that the Volvariella-like species with basidiospore lengths shorter than 11 μm and possessing a gelatinous ixocutis-type pileipellis form a distinct clade from Volvariella s.str. and are more closely related to the genus Pluteus (Justo et al. 2011a). This led to the segregation of the genus Volvopluteus Vizzini, Contu & Justo to accommodate these taxa, which are distinct from Volvariella s.str. and Pluteus s.str. (Justo et al. 2011a, 2011b). More recently, based on six-loci phylogenetic analyses, Volvariellaceae Vizzini, Consiglio & P. Alvarado was introduced to accommodate Volvariella s.str. within the order Agaricales (Vizzini et al. 2024). More than 147 species of Volvariella have been described around the world (Chattopadhyay et al. 2022; Vizzini et al. 2024; Hussain et al. 2025). To date, 22 species of Volvariella have been reported in India (Table 1), of which only eight have been reported from West Bengal, eastern India (Dutta et al. 2011, 2013; Chattopadhyay et al. 2022).
The genus Volvopluteus is typified by Volvopluteus gloiocephalus (DC.) Vizzini, Contu & Justo as the type species. Species of Volvopluteus are characterized by medium- to large-sized basidiomata, free lamellae, a pinkish spore print, the presence of a volva at the stipe base, inamyloid, subglobose to broadly ellipsoid basidiospores, and a gelatinous ixocutis-type pileipellis (Justo et al. 2011a; Kaygusuz et al. 2021). To date, only 12 species have been listed in Species Fungorum (www.speciesfungorum.org; accessed on 12 December 2025), viz. Vp. asiaticus Justo & Minnis, 2011, Vp. canalipes (Murrill) Montoya, Bandala & Esqueda, 2021, Vp. delicisus Chuan H. Li & T.H. Li, 2025, Vp. diversisporus M. Kaur & Yadw. Singh, 2014, Vp. earlei (Murrill) Vizzini, Contu & Justo, 2011, Vp. gloiocephalus (DC.) Vizzini, Contu & Justo, 2011, Vp. longipes Xue T. Zhu & L. J. Liu, 2025, Vp. michiganensis (A.H. Sm.) Justo & Minnis, 2011, Vp. platensis Xue T. Zhu & L. J. Liu, 2025, Vp. shafferii M. Kaur & Yadw. Singh, 2014, and Vp. yunnanensis D.G. Zheng & Karun., 2025. Among these, only two species have been reported from India, viz., Vp. diversisporus and Vp. shafferii (Table 1). However, both records are currently considered invalid in MycoBank (https://www.mycobank.org; Zheng et al. 2025). Importantly, no species of Volvopluteus has been reported from West Bengal in eastern India.
During repeated field surveys conducted from 2024 to 2025 in Barasat, West Bengal, India, several specimens belonging to the families Volvariellaceae and Pluteaceae were collected. Among the new collections, two new species belonging to Volvariella and Volvopluteus are introduced based on morphological characteristics and combined (nrITS and nrLSU) phylogenetic analyses. Detailed descriptions, illustrations, and phylogenetic analysis results of the two new species are presented.
Materials and methods
Sample collection and morphological observation
Fresh basidiomata of Volvariella and Volvopluteus were photographed in situ at Talikhola and Kazibari, Barasat, West Bengal, India. Collection details were recorded (Shaffer 1957; Boekhout 1990; Rathnayaka et al. 2024), and the specimens were taken to the laboratory at West Bengal State University in plastic collection boxes. Morphological descriptions of the specimens were made following Largent et al. (1977). For color terminology, Kornerup and Wanscher’s (1978) color code was strictly followed. Collected specimens were dried at 45–50 °C with an electric food dryer (Hu et al. 2022).
For microscopic characterization, thin sections from multiple basidiomata were revived in 5% KOH and stained with a 1% Congo red solution (Tarafder et al. 2023; Lu et al. 2024). The notation [30, 2, 1] indicates that measurements were conducted on a total of 30 basidiospores obtained from a multiple-sample collection at a specific time point. Additionally, the abbreviations X_m_, Q, and Q_m_ represent the following: X_mr_, the range of basidiospore arithmetic means of the spore length by spore width (± standard deviation), and X_mm_, the mean of basidiospore means (± SD); Q, the quotient of basidiospore length/width, indicated as a range of variation in n spores measured; Q_mr_, the range of Q_m_ values, and Q_mm_, the mean of Q_m_ values; n, the number of basidiospores measured; and s, the number of specimens involved, with the standard deviation specified. Dried voucher specimens were deposited in the Central National Herbarium (CAL).
DNA extraction, PCR amplification, and sequencing
Genomic DNA was extracted from the herbarium specimens using the E.Z.N.A.® Fungal DNA Mini Kit (Omega Bio-Tek, Inc., Norcross, USA), with slight modifications to the manufacturer’s protocol and to the protocol described in earlier studies (Tarafder et al. 2022, 2023). After DNA extraction, the primer pairs ITS1 and ITS4 (White et al. 1990; Gardes and Bruns 1993) were used to amplify the internal transcribed spacer (nrITS) gene regions of ribosomal DNA, and LR0R and LR3 (Vilgalys and Hester 1990) were used to amplify nrLSU. The DNA fragments were amplified on an Applied Biosystems 2720 automated thermal cycler, following the methods described by Dutta et al. (2017) and Tarafder et al. (2022). PCR amplification was carried out in a 25 µL reaction volume, which consisted of 12.5 µL of 2× Bench Top™ Taq Master Mix, 9.5 µL of ddH2O, 1 µL of each primer (10 µM), and 1 µL of genomic DNA. The PCR thermal cycling programs for nrITS and nrLSU amplification were as follows: an initial denaturation at 94 °C for 4 minutes, followed by 35 cycles of denaturation at 94 °C for 30 seconds, annealing at 56 °C for 45 seconds, elongation at 72 °C for 1 minute, then a final extension at 72 °C for 7 minutes, and storage at 4 °C. Following amplification, the PCR products were purified with the QIAquick® Gel Extraction Kit (QIAGEN, Germany) and sequenced with the primers listed in Table 2 using the commercial sequencing provider GeneSpec Pvt. Ltd. (Karnataka, India). All newly generated sequences in this study were deposited in GenBank (https://www.ncbi.nlm.nih.gov/genbank/) and listed in Table 3.
Dataset representation
The amplified sequences were checked for quality, assembled using the BioEdit Sequence Alignment Editor version 7.2.5 (Hall 1999), and used to perform BLASTn searches against the NCBI GenBank database (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Subsequently, closely related sequences of the taxa with zero E-values were retrieved from the database to generate the dataset. Additionally, the sequences used in earlier studies on Volvariella and Volvopluteus (Justo et al. 2011a; Malysheva et al. 2019; Chattopadhyay et al. 2022; Zheng et al. 2025) were also obtained from the database to prepare the final dataset (Table 3). The sequence of Cantharocybe gruberi has been used as an outgroup taxon as per the study by Malysheva et al. (2019).
Sequence alignment and phylogenetic analyses
The combined nrITS and nrLSU datasets were aligned in the CIPRES XSEDE resource using the online program MAFFT v7.427 (Katoh and Standley 2016; Katoh et al. 2019). The aligned sequences were then imported into Aliview v1.17.1 (Larsson 2014) for manual improvement and trimming of both ends.
In the CIPRES web portal (http://www.phylo.org/portal2/) (Miller et al. 2012), jModelTest 2.1.10 v20160303 (Darriba et al. 2012) was used to identify a statistically suitable model for the given dataset to perform Maximum likelihood (ML) analysis. The GTR+I+G model was chosen for the dataset based on the lowest BIC value of 14452.082637.
Maximum likelihood analysis was performed with RAxML-HPC2 v8.2.12 (Stamatakis 2014) on the CIPRES XSEDE resource using the selected model from jModelTest 2.1.10 v20160303, with 1,000 bootstrap replicates. Bayesian analyses (BA) were conducted with MrBayes v3.2.2 (Ronquist et al. 2012) using MCMC methods (Geyer 1991) under a GTR+I+G model. Markov chains were run for 1 × 10^6^ generations, saving a tree every 1000^th^ generation, with all remaining parameters set to default. The average standard deviation of split frequencies reached 0.013 at the end of the run. Convergence was further assessed by examining effective sample size (ESS) values and potential scale reduction factor (PSRF) values. ESS values for all parameters exceeded 200, including the log-likelihood (lnL) with an ESS of 215, indicating adequate sampling of the posterior distribution. PSRF values for all parameters were close to 1.00, confirming convergence among independent runs. The initial 25% of recovered trees were excluded as burn-in, and the remaining trees were then used to estimate the posterior probabilities of the group. ML bootstrap values (MLBS) ≥ 50% and Bayesian posterior probability (PP) values ≥ 0.90 are reported in the resulting tree (Fig. 1). The phylogenetic trees were visualized and edited using FigTree v1.4.0 software (Rambaut and Drummond 2012).
Phylogenetic tree for Volvariella and Volvopluteus generated from the combined nrITS and nrLSU rDNA sequences using maximum likelihood (−InL = 14452.082637) analysis and Bayesian inference (BI) analyses. Values to the left of ‘/’ are MLBS support, and those to the right of ‘/’ indicate Bayesian posterior probability (PP) support. The MLBS values ≥ 70% and PP values ≥ 0.90 are shown above or below the nodes. The newly generated sequences are shown in bold blue. Voucher numbers for the sequences are indicated in the tree after the taxon name, and other type species are indicated in bold black. ‘T’ denotes the holotype of the newly sequenced taxa.
Results
Phylogenetic analyses
The phylogenetic analyses were conducted using combined nrITS and nrLSU rDNA datasets. A total of 87 sequences were aligned, and the ends of each were subsequently trimmed. This resulted in a dataset consisting of 1,547 nucleotides; after trimming the ends of the individual alignments, the aligned datasets were 702 bp for nrITS and 845 bp for nrLSU, including gaps. The combined alignment comprised 649 parsimony-informative characters, 142 singleton sites, and 961 constant characters. The estimated base frequencies were as follows: A = 0.251, C = 0.215, G = 0.316, T = 0.218; substitution rates AC = 2.22735, AG = 5.33961, AT = 2.22735, CG = 1.00000, CT = 16.55070, GT = 1.00000. The phylogenetic tree derived from ML and MrBayes analyses exhibited nearly identical topology. Therefore, the tree obtained from ML analysis (−InL = 14452.082637) has been displayed in the present manuscript (Fig. 1), and the support values recovered from ML (MLBS ≥ 50%) and BA (PP ≥ 0.90) are presented.
In the phylogenetic tree (Fig. 1), taxa belonging to the genera Volvariella and Volvopluteus are separated from each other, and each genus appears monophyletic for the included taxa.
Volvariella s.str.
Members of the genus Volvariella are subdivided into two distinct clades (Clade I and Clade II), agreeing with Chattopadhyay et al. (2022). Clade I is well supported by the maximum likelihood bootstrap values (96% MLBS) but shows only moderate support in Bayesian posterior probability (0.91 PP) and includes all other Volvariella species. Clade II comprises taxa that include V. diplasia, V. volvacea, V. orientalis, and V. bombycina; in contrast, the remaining species of V. bilobata, originally reported from India, and V. pulla, recorded from Vietnam, together with the newly proposed species Volvariella areolavolvata (CAL 2383), are in this same clade, confirming its systematic position within the genus Volvariella (Fig. 1).
Volvopluteus s.str.
Phylogenetic analyses of the genus Volvopluteus (Clade III) are also statistically well supported (100% MLBS and 1.00 PP), and this clade includes all representative sequences of Volvopluteus, along with those of the newly described Indian taxon Vp. fibrillobrunneus (CAL 2385), confirming its systematic position within the genus Volvopluteus (Fig. 1). Moreover, Vp. fibrillobrunneus is further placed between Vp. platensis and Vp. earlei in the phylogenetic analysis, with moderate MLBS and strong PP support (78% MLBS/0.98 PP).
Taxonomy
Volvoriella
areolavolvata
Taxon classificationFungi
E. Tarafder, Enjam & A.R. Sherpa. sp. nov.
4024FB20-B0BB-549C-8CEF-ED5CB0B650BE
Index Fungorum: IF904792
Etymology.
The specific epithet “areolavolvata” is derived from the Latin aeriolatus (areolate cracking on the outer surface) and refers to the characteristic of the volva.
Diagnosis.
Volvariella areolavolvata differs from Volvariella bombycina by a rigid volva with a distinctive externally areolate, cracked surface that forms pale to dark brown patches, slightly smaller basidiospores (6.8–9.0 × 4.4–5.5 μm), and larger cheilocystidia (63–131 × 21–37 μm).
Holotype.
India • West Bengal: North-24-Parganas District, Barasat, Talikhola, 22°44'50.9"N, 88°26'54.7"E, elev. 13.0 m, solitary on dead wood of Ficus sp., 07 August 2025, A.R. Sherpa, E. Tarafder & E. Hoque, Sherpa 01/2025 (CAL 2383, holotype).
Description.
Basidiomata medium to large. Pileus 100–150 mm, fleshy, ovoid to subglobose when young, hemispherical to broadly campanulate, expanding obtusely umbonate; surface dry, sometimes viscid, covered with conspicuous silky fibrils or silky hairs; color white to whitish (10A1), often becoming slightly yellowish (4A5) in the center with age; margin entire. Context 10 mm thick at the disc, cream white (2A2). Lamellae free, 15–20 mm broad, crowded with lamellulae of different lengths; white when young, becoming pinkish brown (8A2–4) at maturity; edge concolorous, entire. Stipe 80–120 × 8–10 mm, central, cylindrical, curved at the base, solid; surface slightly pruinose, white (1A1) creamy, yellowish white (3B3), weakly hairy. Volva 25–30 mm broad, erect, rigid, thick, and fleshy, irregularly 5–6 lobbed; internally white (1A1), externally cracked areolate pale to dark brown (6F6–8) patches. Odor and taste indistinct.
Field photographs of the basidiomata of (a–d) Volvariella areolavolvata (CAL 2383, holotype). Scale bars: 10 mm (a–c); 15 mm (d).
Basidiospores [30/2/1] (6.8–)7.1–8.7(–9.0) × (4.4–)4.6–5.3(–5.5) µm, [X_mr_ = 7.61–7.92 × 4.85–4.95, X_mm_ = 7.75 ± 0.17 × 4.91 ± 0.33 μm, Q_mr_ = 1.23–1.85, Q_mm_ = 1.56 ± 0.18, n = 30 basidiospores per 2 specimens], oval to ellipsoid, hyaline to light yellowish in 5% KOH, smooth, dextrinoid, thin-walled, and apiculate. Basidia 17.4–26 × 7.4–10 µm, clavate, hyaline in 5% KOH, smooth, with 4 sterigmata, no clamp present at the base. Cheilocystidia 63–131 × 21–37 µm, hyaline in 5% KOH, thin-walled, mostly lageniform, with a long, narrow neck, forming a sterile layer at the lamellae edge. Pleurocystidia 44–56 × 13–17 µm, hyaline, thin-walled, broadly clavate with rounded apex or utriform, abundant in lamellae edge. Pileipellis a cutis made up of 10–37 µm broad, hyaline, thin-walled hyphae, clamp connections absent. Stipitipellis as compactly arranged cutis, 31–52 × 6.7–11.9 µm in diam., thin-walled, septate, clamp connections absent. Volva composed of septate, thin-walled 5.2–8.3 µm broad hyphae, with clamp connections absent.
Volvariella areolavolvata (CAL 2383, holotype). a. Basidiospores; b, c. Basidia; d, e. Cheilocystidia; f. Pleurocystidia; g. Pileipellis hyphae; h. Stipe hyphae; i. Hyphal cells in the volva. Scale bars: 10 μm (a); 20 μm (b–f); 15 μm (g–i).
Habitat and known distribution.
Solitary, on dead wood of Ficus sp. So far, it is known only from India.
Additional specimens examined.
India • West Bengal: North-24-Parganas district, Barasat, Talikhola, 22°44'50.9"N, 88°26'54.7"E, elev. 13.0 m, 12 August 2025, A.R. Sherpa, E. Tarafder & E. Hoque, Sherpa 02/2025 (CAL 2384, paratype).
GenBank numbers.
CAL 2383 (holotype): nrITS = PX776811, nrLSU = PX764990. CAL 2384: nrITS = PX776812, nrLSU = PX764991.
Notes.
Volvariella areolavolvata is morphologically close to Volvariella bombycina but can be distinguished by a combination of macroscopic and microscopic characters. Volvariella areolavolvata is characterized by comparatively large basidiomata, a white to creamy pileus lacking squamules, a pure white stipe, and a white volva with brown patches, in combination with medium-sized basidiospores (mean = 7.75 × 4.9 µm). In contrast, V. bombycina differs by its robust and stout basidiocarps, a light-colored to pale brownish pileus that is entirely squamulose, a thick stipe, and a large, brown volva (Fig. 2). Moreover, V. bombycina has distinctive microscopic characteristics, including relatively small cheilocystidia (50–110 × 10–25 μm; Table 4) with frequently elongated apices.
Table 4.: Comparison of Volvariella areolavolvata (CAL 2383) with its closely related taxa.
In our phylogenetic analyses, our collections (CAL 2383 and CAL 2384) clustered closely with Volvariella pulla and Volvariella bilobata but formed a distinct, well-supported clade (100% MLBS/1.00 PP; Fig. 1). Volvariella pulla, originally described from Vietnam (Malysheva et al. 2019), has an ochraceous or grayish brown stipe, an irregularly lobed volva colored dirty grey-brown with rusty brown spots, smaller basidiospores (5.5–8.0 × 4–5 μm vs. 6.8–9.0 × 4.4–5.5 μm), and much larger pleurocystidia (52–83 × 14–34 μm). The Indian collection of Volvariella bilobata differs from V. areolavolvata by its smaller pileus (45–70 mm diam.), colored grayish to dark brown, bilobed saccate volva, and much smaller basidiospores (4.8–5.5 × 2.7–3.5 μm; Fig. 3) and pleurocystidia (18.5–27.5 × 7–10 μm) (Chattopadhyay et al. 2022). Here, we introduce our collections as a novel species of V. areolavolvata based on morpho-molecular analyses.
Volvopluteus
fibrillobrunneus
Taxon classificationFungiAgaricalesPluteaceae
Enjam, E. Tarafder & A.R. Sherpa. sp. nov.
4795E1CE-F5EA-54C1-B191-12D362BE9128
Index Fungorum: IF904793
Etymology.
The specific epithet “fibrillobrunneus” refers to the brown color with a fibrillose pileus surface.
Diagnosis.
Volvopluteus fibrillobrunneus differs from Volvopluteus earlei by its fibrillose brown pileus surface with larger basidiocarps (Pileus < 50 mm in diameter Vp. earlei) and smaller basidiospores (8.3–11.5 × 5.1–7.1 μm).
Holotype.
India • West Bengal: North-24-Parganas district, Barasat, Near Kazibari Bus Stand, 22°44'45.5"N, 88°26'41.9"E, elev. 13.0 m, Scattered on the soil around with rich humus in broad-leaved forest, 18 September 2024, E. Hoque, AE-37/2024 (CAL 2385, holotype).
Description.
Basidiomata medium to large. Pileus 80–97 mm diam., plano-convex to plane with age, low umbo at center, not viscid, dry, glabrous, innately radially fibrillose, striate, surface light brown (6D4), grayish brown (6D3) with grey (5B2) when young, dark blonde (5D4) center and birch grey (5C2) sides after maturity, becoming lighter grayish brown (5F3–5E4) towards the margin and yellowish brown (5B2–5C4) after dried; margin striate, radially fibrillose, slightly cracked with maturity, cream whitish squamulose present. Context 4 mm broad, white to grayish white (2B1). Lamellae free, approximately 10 mm broad, close to crowded, pinkish white (8A3–8B3), brownish gray (11C2) at maturity. Stipe 120–140 × 10–14 mm, central, cylindrical, solid, white, glabrous, not viscid, when young visibly broadening towards the base and in maturity with bulbous base enclosed in sac-like volva. Volva 15–19 × 8–10 mm, free from the stipe, saccate, membranous, 2–3 lobed, with a white to light brownish outer surface and whitish inner surface. Odor and taste indistinct.
Field photographs of the basidiomata of (a–c) Volvopluteus fibrillobrunneus (CAL 2385, holotype) in natural habitat. Scale bars: 10 mm (a–c).
Basidiospores (8.3–)8.5–9.6(–11.5) × (5.1–)5.4–6.3(–7.1) µm, [X_mr_ = 8.65–9.43 × 5.88–6.25, X_mm_ = 9.85 ± 0.92 × 6.30 ± 0.49 μm, Q_mr_ = 1.36–1.75, Q_mm_ = 1.57 ± 0.19, n = 30 basidiospores per 2 specimens], ellipsoid to elongate, smooth, hyaline to light yellowish in 5% KOH, thin-walled, and apiculate. Basidia 25.7–45.6 × 9.8–11.9 µm, clavate, hyaline in 5% KOH, smooth, with 2–4 prominent sterigmata, no clamp at base. Cheilocystidia 34–72 × 15–29 µm, hyaline in 5% KOH, thick-walled, mostly clavate, forming a sterile layer at the lamellae edge. Pleurocystidia 53–75 × 14–21 µm, hyaline, thin-walled, lageniform rostrate with apical appendages. Pileipellis ixocutis, made up of hyphae 25–32 µm broad, hyaline, thin-walled, clamp connections absent. Stipitipellis as compactly arranged hyphae, 23–38 µm broad, thin-walled, clamp connections absent. Volva composed of septate interwoven cylindrical hyphae, thin-walled 9–13 µm broad, clamp connection absent.
Volvopluteus fibrillobrunneus (CAL 2385, holotype). a. Basidiospores; b. Basidia; c, d. Cheilocystidia; e. Pleurocystidia; f. Pileipellis hyphae; g. Stipe hyphae; h. Hyphal cells in the volva. Scale bars: 10 μm (a); 20 μm (b–e); 15 μm (f–i).
Habitat and known distribution.
Scattered on the soil around with rich humus in a broad-leaved forest. So far, it is only known from India.
Additional specimens examined.
India • West Bengal: North-24-Parganas district, Barasat, near Kazibari Bus Stand, 22°44'45.5"N, 88°26'41.9"E, elev. 13.0 m, scattered on the soil around with rich humus in broad-leaved forest, 21 September 2024, E. Hoque, AE-41/2024 (CAL 2386, paratype).
GenBank numbers.
CAL 2385 (holotype): nrITS = PX764988, nrLSU = PX764992; CAL 2386: nrITS = PX764989, nrLSU = PX764993.
Notes.
Volvopluteus fibrillobrunneus is characterized by a medium- to moderately large-sized (80–97 mm) pileus, entirely light brown to grayish brown, with a radially fibrillose surface; close to crowded lamellae colored pinkish white to brownish gray; a multi-lobed saccate volva with a white to light brownish outer surface and a whitish inner surface (Fig. 4); larger basidiospores measuring 8.3–11.5 × 5.1–7.1 μm; clavate to ventricose cheilocystidia measuring 34–72 × 15–29 μm; lageniform rostrate pleurocystidia with apical appendages measuring 53–75 × 14–21 μm; and habitat on rich humus soil (Fig. 5). Among macro-morphologically similar taxa (Table 5), Vp. earlei can be distinguished by its smaller basidiospores (11–16 × 8–11 μm), the presence of longer pleurocystidia (up to 110 μm), and the presence of cheilocystidia (Shaffer 1962; Justo and Castro 2010; Giannoni et al. 2018). Moreover, Volvopluteus earlei was originally described from Cuba (Murrill 1911) and later reported from the USA (Coker 1947), Mexico (Vázquez et al. 1989), Africa (Heinemann 1975), Argentina (Niveiro and Albertó 2012), Italy (Contu 2006; Giannoni et al. 2018), Spain (Justo and Castro 2010), Turkey (Kaygusuz et al. 2021), and India (Chouhan and Panwar 2021).
Table 5.: Comparison of Volvopluteus fibrillobrunneus (CAL 2385) and its closely related taxa. Abbreviations: NR = not reported.
In the phylogenetic analyses (Fig. 1), Vp. fibrillobrunneus is placed as a sister group to Vp. platensis, originally described from China (Zhu et al. 2025), and can be distinguished from Vp. fibrillobrunneus by the smaller basidiomata (48–70 mm broad in Vp. platensis), a slightly brownish to grayish white pileus, and clavate to fusoid cheilocystidia (Zhu et al. 2025). Hence, here we introduce our collections as a novel species of Volvopluteus s.str., i.e., Vp. fibrillobrunneus, based on morpho-molecular analyses.
Discussion
Geographically, West Bengal is located in eastern India and is bordered by the high Himalayan peaks in the north, coastal ecosystems in the south, and the Gangetic delta and plateaus in between (Tarafder et al. 2023). Subtropical broadleaved to subalpine forests dominate in the Himalayan region; in contrast, littoral and swamp forests cover the coastal regions, and dry deciduous forests prevail on the plateaus and in the Gangetic plains (Dutta et al. 2015). This wide range of phyto-topographical features, together with altitudinal and climatic variations, creates ample opportunities for the luxuriant growth of macrofungi. Although several species of Volvariella have been reported from West Bengal, India, most earlier studies were based solely on morphological characterization, viz., Volvariella castanea (Massee) G.C. Rath (Calcutta, Massee 1912), V. delicatula (Massee) Manjula (Calcutta, Massee 1912), V. diplasia (Berk. & Broome) Singer (Hooghly and Bose 1920a), V. pusilla (Pers.) Singer (South 24-Parganas ‘fide’ Dutta et al. 2011, 2013), V. terastia (Berk. & Broome) Singer (Berk. & Broome) Singer (Calcutta, Bose 1920b, 1921), V. thwaitesii (Hook. f. ex Berk.) G.C. Rath (Darjeeling, Berkeley 1850), and V. volvacea (Bull.) Singer (South 24-Parganas ‘fide’ Dutta et al. 2011, 2013).
Volvariella areolavolvata was characterized by a large-sized (100–150 mm), entirely white pileus with a slightly silky hairy surface, often becoming slightly yellowish in the center with age; crowded lamellae colored pinkish white to grayish brown; a volva that is rigid, thick, and fleshy, irregularly lobed, internally white, and externally cracked, with areolate pale to dark brown patches; smaller basidiospores (6.8–9.0 × 4.4–5.5 μm); mostly lageniform cheilocystidia (63–131 × 21–37 μm); broadly clavate with a rounded apex or utriform pleurocystidia (44–56 × 13–17 μm; Table 4); and habitat on dead wood. In our phylogenetic analyses, V. areolavolvata clustered closely with V. pulla and V. bilobata, forming a distinct and well-supported clade (100% MLBS/1.00 PP; Fig. 1).
Additionally, Vp. fibrillobrunneus falls between Vp. platensis and Vp. earlei in the phylogenetic analysis, with moderate MLBS and strong PP support (78% MLBS/0.98 PP). However, Volvopluteus fibrillobrunneus was characterized by a medium- to moderately large-sized (80–97 mm), entirely light brown to grayish brown pileus with a radially fibrillose surface; close to crowded lamellae colored pinkish white to brownish gray; a multi-lobed saccate volva with a white to light brownish outer surface and a whitish inner surface; larger basidiospores (8.3–11.5 × 5.1–7.1 μm); clavate to ventricose cheilocystidia (34–72 × 15–29 μm); and lageniform pleurocystidia (55–60 × 29–47 μm). Among macro-morphologically similar taxa (Table 5), Vp. earlei can be distinguished by its smaller basidiospores (11–16 × 8–11 μm) and the presence of longer pleurocystidia (up to 110 μm). In the phylogenetic tree for the combined dataset (Fig. 1), our newly described taxon is also close to Vp. platensis, which was initially described by relatively small basidiomata, with pileus diameters not exceeding 45 mm, a slightly brownish to grayish white pileus, and clavate to fusoid cheilocystidia (Zhu et al. 2025).
The discovery of V. areolavolvata and Vp. fibrillobrunneus increases our understanding of the diversity and distribution of these genera in eastern India. In conclusion, V. areolavolvata and Vp. fibrillobrunneus are supported by both morphological and phylogenetic analyses. Moreover, their discovery significantly advances the understanding of agaric diversity in eastern India and underscores the need for continued exploration and integrative taxonomic studies, given modern phylogenetic analyses in the region.
Supplementary Material
XML Treatment for Volvoriella areolavolvata
XML Treatment for Volvopluteus fibrillobrunneus
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Boekhout T (1990) Volvariella. In: Bas C, Kuyper Th W, Noordeloos ME, Vellinga EC (Eds) Flora Agaricina Neerlandica. AA Balkema, Rotterdam, The Netherlands, 56–64.
- 2Bose SR (1920 a) Records of Agaricaceae from Bengal. Journal of the Asiatic Society of Bengal 16: 347–354.
- 3Bougher NL, Barrett MD (2023) Volvopluteus earlei and Volvariella taylorii: new fungi for Western Australia (Basidiomycota: Agaricales: Pluteaceae). Nuytsia 34: 187–201. 10.58828/nuy 01054 · doi ↗
- 4Chattopadhyay P, Talukdar M, Beypih J, Tayung K, Dutta AK (2022) A new species of Volvariella (Agaricales, Basidiomycota) from West Bengal, India. Phytotaxa 567(1): 36–48. 10.11646/phytotaxa.567.1.3 · doi ↗
- 5Chouhan R, Panwar C (2021) The genus Volvariella from Mount Abu, Rajasthan, India. Journal of Mycopathological Research 59: 63–66.
- 6Contu M (2006) Volvariella earlei (Basidiomycota, Pluteaceae) nuova per l’Europa, e note sulla tassonomia di Volvariella media sensu J. Lange. Micologia e Vegetazione Mediterranea 21(2): 101–106. [in Italian]
- 7Crous PW, Wingfield MJ, Burgess TI, Carnegie AJ, Hardy G, Smith D, Summerell BA, Cano-Lira JF, Guarro J, Houbraken J, Lombard L, Martin MP, Sandoval-Denis M, Alexandrova AV, Barnes CW, Baseia IG, Bezerra JDP, Guarnaccia V, May TW, Hernandez-Restrepo M, Stchigel AM, Miller AN, Ordonez ME, Abreu VP, Accioly T, Agnello C, Agustin Colman A, Albuquerque CC, Alfredo DS, Alvarado P, Araujo-Magalhaes GR, Arauzo S, Atkinson T, Barili A, Barreto RW, Bezerra JL, Cabral TS, Camello Rodriguez F, Cruz R, Dani · doi ↗ · pubmed ↗
- 8Daniëls PP, Hama O, Fernández A, García-Pantaleón FI, Baragé M, Ibrahim D, Alcántara MR (2015) First records of some Asian macromycetes in Africa. Mycotaxon 130(2): 337–359. 10.5248/130.337 · doi ↗