Morpho-molecular characterization reveals three Cortinarius subgenus Dermocybe species from southwest China (Agaricales, Cortinariaceae)

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
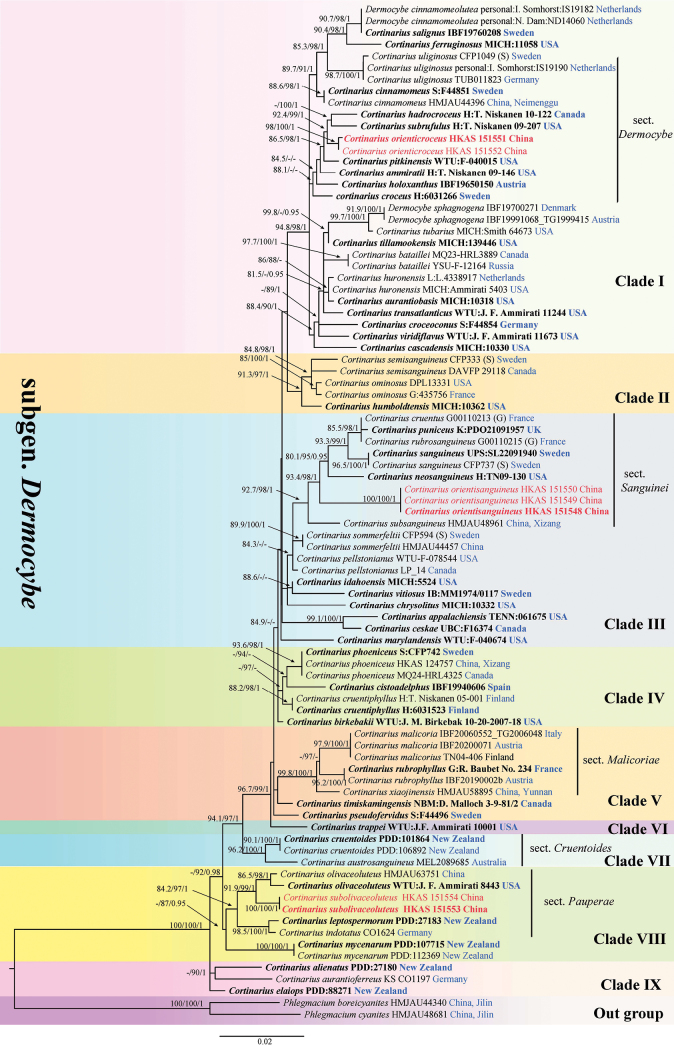 Figure 1
Figure 1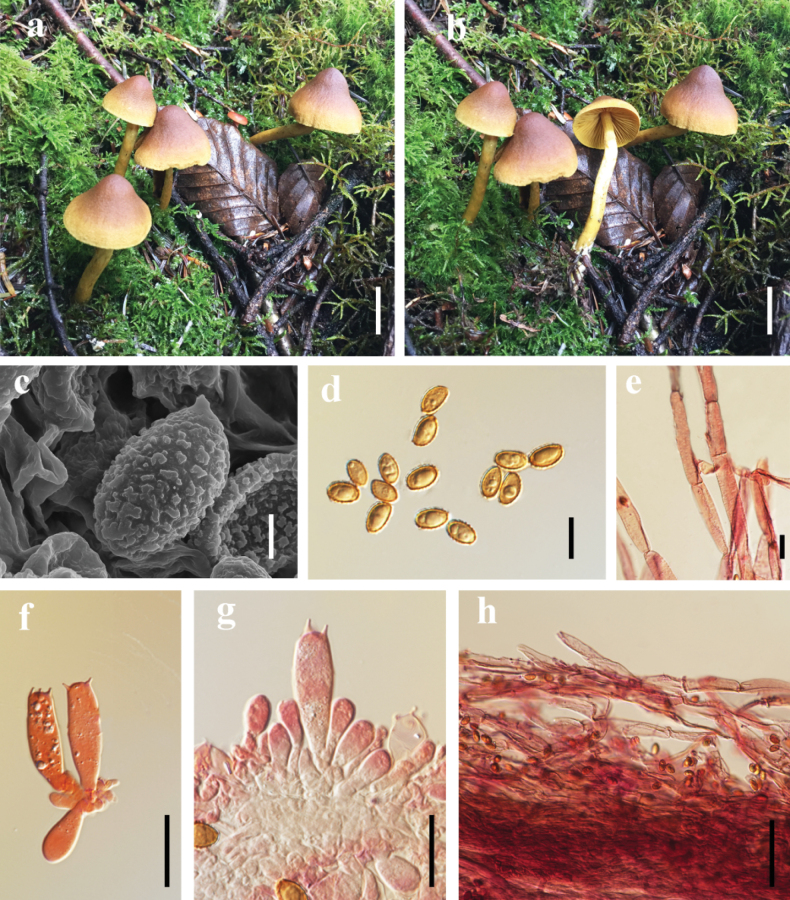 Figure 2
Figure 2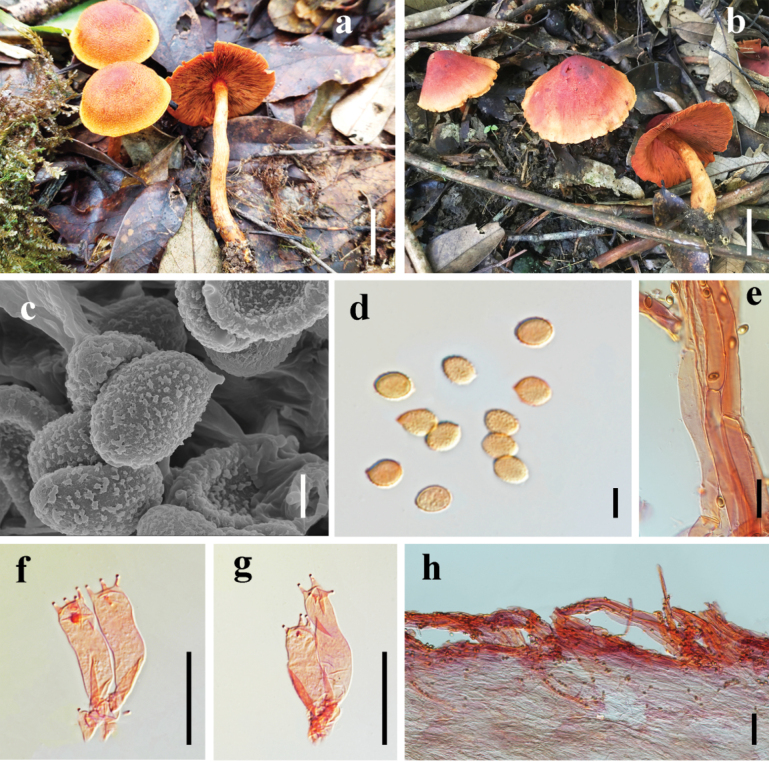 Figure 3
Figure 3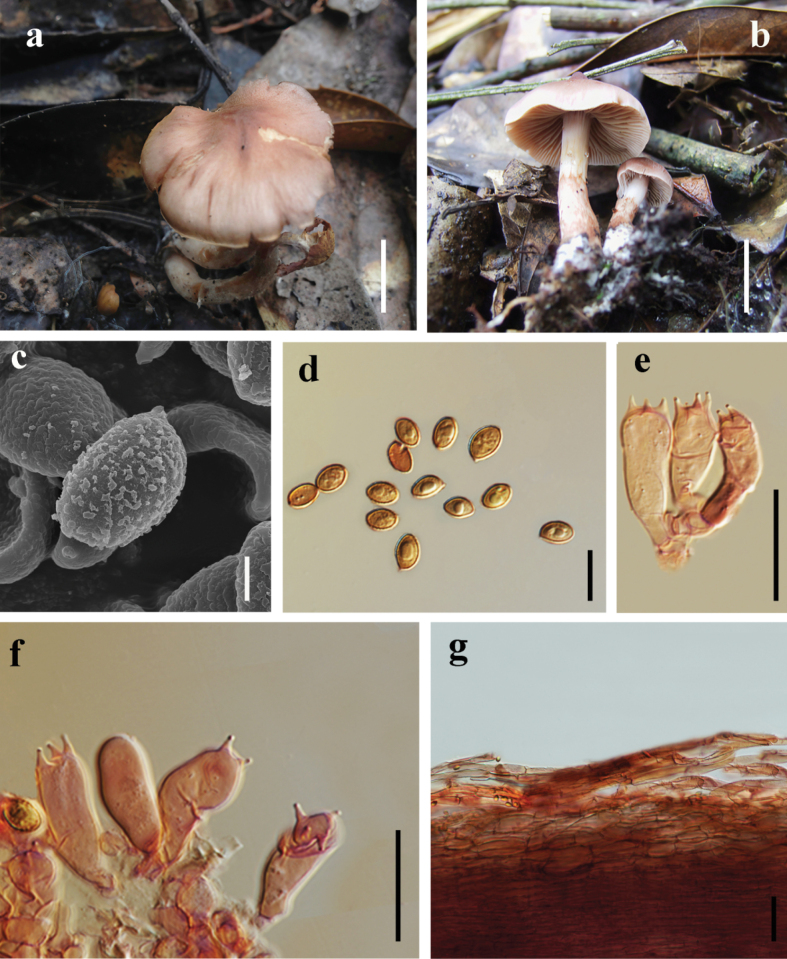 Figure 4
Figure 4| Species | Voucher/Isolate no. | GenBank accession no. | Distribution | References | |
|---|---|---|---|---|---|
|
| LSU | ||||
| H:TN09-130 |
|
| USA |
| |
|
| G00110215 (G) |
| – | France |
|
|
| CFP737 (S) |
|
| Sweden |
|
|
| HMJAU48961 |
|
| China | unpublish |
|
| H:6031266 |
| – | Sweden | Niskanen et al. (2014) |
|
| HMJAU44396 |
|
| China |
|
|
| TUB011823 |
|
| Germany |
|
|
| IBF20190002b |
| – | Austria | unpublish |
|
| HMJAU58895 |
|
| China |
|
|
| MEL2089685 |
|
| Australia |
|
|
| HMJAU63751 |
| – | China | unpublish |
|
| WTU:J. F. Ammirati 8443 |
| – | USA | Niskanen et al. (2014) |
|
| CO1624 |
|
| Germany | unpublish |
|
| PDD:27183 |
|
| New Zealand | unpublish |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| KS CO1197 |
| – | Germany |
|
|
| PDD:112369 |
| – | New Zealand |
|
|
| HMJAU44457 |
|
| China |
|
|
| IBF19760208 |
| – | Sweden |
|
|
| personal:N. Dam:ND14060 |
| – | Netherlands | unpublish |
|
| personal:I. Somhorst:IS19182 |
| – | Netherlands |
|
|
| MICH:11058 |
| – | USA |
|
|
| personal:I. Somhorst:IS19190 |
| – | Netherlands | unpublish |
|
| S:F44851 |
| – | Sweden |
|
|
| IBF19650150 |
| – | Austria |
|
|
| H:T. Niskanen 09-146 |
| – | USA | Niskanen et al. (2014) |
|
| WTU:F-040015 |
| – | USA | Niskanen et al. (2014) |
|
| H:T. Niskanen 09-207 |
| – | USA | Niskanen et al. (2014) |
|
| H:T. Niskanen 10-122 |
| – | Canada | Niskanen et al. (2014) |
|
| MICH:10330 |
| – | USA |
|
|
| S:F44854 |
| – | Germany | Niskanen et al. (2014) |
|
| WTU:J. F. Ammirati 11673 |
| – | USA | Niskanen et al. (2014) |
|
| MQ23-HRL3889 |
| – | Canada | unpublish |
|
| YSU-F-12164 |
| – | Russia |
|
|
| MICH:139446 |
| – | USA | Niskanen et al. (2014) |
|
| WTU:J. F. Ammirati 11244 |
| – | USA | Niskanen et al. (2014) |
|
| MICH:10318 |
| – | USA |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| MICH:Ammirati 5403 |
| – | USA |
|
|
| L:L.4338917 |
| – | Netherlands |
|
|
| MICH:Smith 64673 |
| – | USA |
|
|
| IBF19991068 _TG1999415 |
| – | Austria | unpublish |
|
| IBF19700271 |
| – | Denmark | unpublish |
|
| WTU:J.F. Ammirati 10001 |
| – | USA | Liu Y. (1995) |
|
| CFP333 (S) |
| – | Sweden |
|
|
| MICH:10362 |
| – | USA |
|
|
| DAVFP 29118 |
| – | Canada |
|
|
| G:435756 |
| – | France |
|
|
| DPL13331 |
| – | USA | unpublish |
|
| WTU:F-040674 |
| – | USA |
|
|
| MICH:5524 |
| – | USA |
|
|
| MICH:10332 |
| – | USA |
|
|
| WTU-F-078544 |
| – | USA | unpublish |
|
| LP_14 |
| – | Canada |
|
|
| CFP594 (S) |
| – | Sweden |
|
|
| TENN:061675 |
| – | USA | Niskanen et al. (2014) |
|
| UBC: |
| – | Canada |
|
|
| H:T. Niskanen 05-001 |
| – | Finland | Niskanen et al. (2014) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| H:6031523 |
| – | Finland | Niskanen et al. (2014) |
|
| IBF19940606 |
| – | Spain |
|
|
| S:CFP742 |
| – | Sweden |
|
|
|
| – | China | unpublish | |
|
| G00110213 (G) |
| – | France |
|
|
| K:PDO21091957 |
| – | UK |
|
|
| UPS:SL22091940 |
| – | Sweden |
|
|
| WTU:J. M. Birkebak 10-20-2007-18 |
| – | USA | Niskanen et al. (2014) |
|
| NBM:D. Malloch 3-9-81/2 |
| – | Canada | Niskanen et al. (2014) |
|
| S:F44496 |
| – | Sweden |
|
|
| IB:MM1974/0117 |
| – | Sweden |
|
|
| IBF20060552_TG2006048 |
| – | Italy | unpublish |
|
| IBF20200071 |
| – | Austria |
|
|
| TN04-406 |
| – | Finland |
|
|
| G:R. Baubet No. 234 |
| – | France | Niskanen et al. (2014) |
|
| PDD:101864 |
| – | New Zealand | unpublish |
|
| PDD:106892 |
| – | New Zealand |
|
|
| PDD:107715 |
| – | New Zealand |
|
|
| PDD:88271 |
| – | New Zealand |
|
|
| PDD:27180 |
| – | New Zealand |
|
|
| CFP1049 (S) |
| – | Sweden |
|
|
| MQ24-HRL4325 |
| – | Canada |
|
|
| CFP1049 (S) |
| – | Sweden |
|
|
| MQ24-HRL4325 |
| — | Canada |
|
|
| HMJAU44340 |
|
| China |
|
|
| HMJAU48681 |
|
| China |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSilymarin and Mushroom Poisoning · Mycorrhizal Fungi and Plant Interactions · Fungal Biology and Applications
Introduction
Cortinarius (Pers.) Gray is a genus classified within the class Agaricomycetes, order Agaricales, and family Cortinariaceae. This cosmopolitan genus demonstrates exceptional taxonomic diversity, encompassing more than 3,000 described species with a global distribution spanning all continents except Antarctica (Moser et al. 1975; Jacquemyn et al. 2010; Tedersoo et al. 2011; Harrower et al. 2015; Niskanen et al. 2018; Thoen et al. 2019; Li et al. 2025). Cortinarius is an ecologically important group of ectomycorrhizal fungi that establishes mutualistic symbioses with diverse plant taxa across various ecosystems. Some species, such as Cortinarius armillatus (Fr.) Fr., C. bovinus Fr., and C. emodensis Berk. possess both edible and medicinal value (Li et al. 2025).
Moser et al. (1983) proposed a taxonomic framework dividing the genus Cortinarius into six distinct subgenera: Cortinarius, Leprocybe, Myxacium, Phlegmacium, Sericeocybe, and Telamonia, while treating Dermocybe as an independent genus. Subsequently, Brandrud et al. (1998) refined this classification by consolidating the subgeneric structure into four subgenera: Cortinarius, Myxacium, Phlegmacium, and Telamonia, and incorporated Dermocybe within the subgenus Cortinarius. Later, Bidaud et al. (1994) established a modified system recognizing six subgenera: Cortinarius, Dermocybe, Myxacium, Phlegmacium, Telamonia, and Hydrocybe. The present study adopts the subgeneric classification proposed by Bidaud et al. (1994) with minor taxonomic modifications.
Dermocybe was originally distinguished from other subgenera of Cortinarius primarily on the basis of the presence of anthraquinone pigments, with only a few exceptions (Gruber 1970; Høiland and Holst-Jensen 2000). Nevertheless, its taxonomic status has been subject to ongoing debate (Moser 1978; Høiland 1983; Liu et al. 1997; Kuhnert-Finkernagel and Peintner 2003). Dermocybe sensu lato, hereafter referred to as dermocyboid species, comprises agaricoid fungi that are small to medium-sized and exhibit bright coloration ranging from yellow and red to brownish and olive hues. These species are characterized by a dry stipe and a pileus that varies from dry to slightly viscid, with a silky or occasionally waxy surface texture; the pileipellis displays a somewhat duplex structure with a poorly developed hypodermal layer, and the lamellae are typically adnate or adnexed, displaying yellow, red, orange, or green pigmentation. The partial veil is generally inconspicuous, while a cortina is consistently present; the odor is frequently indistinct, though some species may exhibit a radish-like scent (Liu et al. 1997; Knudsen 2008; Stefani et al. 2014; Soop 2021; Liimatainen et al. 2022; Xie et al. 2025a).
In the recent years, species of Cortinarius have been receiving much attention from mycologists, and many species were discovered from China (Wei and Yao 2013; Xie et al. 2019, 2020, 2021a, b, 2022, 2023a, b, 2024, 2025a, b; Yuan et al. 2020; Luo and Bau 2021; Xie 2022; Liu et al. 2023; Zhang et al. 2023; Zhou et al. 2023; Fan et al. 2024; Hong et al. 2024; Long et al. 2024; Dang et al. 2025; Guan et al. 2025; Jia et al. 2025; Wang et al. 2025a, b; Yang et al. 2025). In this study, a phylogenetic investigation of Cortinarius subg. Dermocybe species was identified using both morphological and molecular data (ITS+LSU). We attempt to 1) elucidate the species diversity of Cortinarius subg. Dermocybe in southwest China; 2) evaluate the phylogenetic relationships within the subgenus.
Materials and methods
Sample collection and morphological studies
From August 2014 to August 2023, seven distinct fungal collections were sourced from multiple localities across Yunnan Province, China, specifically including Lijiang, Honghe, Chuxiong, and Dali. Detailed metadata documentation was meticulously recorded for all collections following established taxonomic protocols (Rathnayaka et al. 2025). Macroscopic characteristics of the pileus, lamellae, and stipe for each collection were documented and photographed in situ. After being brought to the laboratory, the specimens were subsequently desiccated in a mushroom drier. Subsequent macroscopic morphological examination and microscopic photomicrography were conducted in the laboratory following the methodology of Zhao et al. (2015), with necessary modifications applied. The color codes referenced in the descriptions are based on Kornerup and Wanscher (1967), measurements are presented as (a–) b–c (–d), where a and d represent the absolute minimum and maximum values, and the range b–c encompasses at least 90% of all measurements. The notation [n/m/p] denotes the measurement of (n) basidiospores from (p) localities collected at (m) basidiomata. Furthermore, Q refers to the length-to-width ratio (aspect ratio) of the basidiospores, with values expressed as the mean (Q) ± standard deviation (SD). The newly collected specimens have been deposited in the Cryptogamic Herbarium of the Kunming Institute of Botany (herbarium code: KUN-HKAS).
DNA extraction, PCR amplification, and sequencing
Genomic DNA was extracted from dried basidiocarps using the “trilief” Plant Genomic DNA Kit (Tsingke Biotechnology Co., Ltd., Beijing, China). The polymerase chain reaction (PCR) was used to amplify the ribosomal internal transcribed spacer (ITS) and the large subunit of the ribosomal RNA (LSU), with primers ITS1-F/ITS4 (White et al. 1990; Gardes and Bruns 1993), and LR0R/LR5 (Vilgalys and Hester 1990; Rehner and Samuels 1994), respectively. PCR amplification was performed in a total volume of 25 μL, including 21 μL Taq PCR Master Mix (TSINGKE TSE101, Tsingke Biotechnology Co., Ltd., Beijing, China), 1 μL forward primer, 1 μL reverse primer, and 2 μL DNA template. PCR conditions: Initial denaturation at 98 °C for 5 min; followed by 35 cycles of 98 °C for 30 sec (denaturation), 53 °C for 20 sec (annealing), and 72 °C for 30 sec (extension); with a final extension at 72 °C for 10 min. PCR products were verified by 1% ethidium bromide-stained agarose gel electrophoresis, and successful amplicons were sent for sequencing at Tsingke Biotechnology Co., Ltd. (Beijing, China).
Sequence alignment and phylogenetic analyses
The newly generated sequences were carefully inspected and assembled using DNAMAN 9.0. Sequencing data from the recently collected specimens were integrated with relevant sequence data from the literature for comprehensive analysis (Huymann et al. 2024). Reference sequences of related taxa were retrieved from the NCBI database (https://www.ncbi.nlm.nih.gov/) to facilitate phylogenetic tree construction (Table 1). Multiple sequence alignment was performed using MAFFT v.7 (Katoh et al. 2019; Katoh and Standley 2013; available at: https://mafft.cbrc.jp/alignment/server/index.html). Phlegmacium boreicyanites and P. cyanites were designated as the outgroup taxa. Sequences were manually edited using BioEdit 7.0.9 (Hall 1999). The trimmed sequences were subsequently concatenated into a combined dataset using Sequence Matrix V.1.8 (Vaidya et al. 2011). The final dataset was analyzed using both Maximum Likelihood (ML) and Bayesian Inference (BI) methods. Maximum Likelihood (ML) analysis was conducted using IQ-TREE (http://iqtree.cibiv.univie.ac.at/, Trifinopoulos et al. 2016). The best-fit substitution model for each gene partition was selected using MrModeltest v.2.3 (Nylander et al. 2004). Phylogenetic analyses for all gene regions were performed under the ML criterion, with 1,000 bootstrap replicates. The best-fit substitution model for the ITS region was HKY+F+I+G4, while that for the LSU region was TN+F+I. Bayesian Inference (BI) was performed using MrBayes v3.2.6 (Ronquist et al. 2012). Four simultaneous Markov Chain Monte Carlo (MCMC) runs were performed for 2,750,000 generations, with trees sampled every 1,000 generations until the average standard deviation of split frequencies fell below 0.01. The first 25% of generations were discarded as burn-in to calculate posterior probabilities (PP) (Huelsenbeck and Ronquist 2001). The resulting phylogenetic trees were visualized using FigTree v.1.4.4 and further refined in Adobe Illustrator 2020 to optimize graphical presentation.
Results
Phylogenetic analyses
The concatenated ITS-LSU dataset comprised 87 taxa with 1,560 aligned characters (ITS: 1-670 bp; LSU: 671-1,560 bp). Following the methodology established by Huymann et al. (2024) and Xie et al. (2025a), Phlegmacium boreicyanites and P. cyanites were selected as outgroups. The phylogenetic tree generated by Maximum Likelihood (ML) analysis was highly congruent with the topology obtained from Bayesian Inference (BI). Therefore, the ML tree topology is presented along with statistical values generated by ML bootstrap support (MLBS) and Bayesian posterior probabilities (BIPP) (Fig. 1).
Phylogenetic tree inferred from the combined ITS and LSU sequence dataset using Maximum Likelihood (ML). Branch support values are indicated as “–/–/–” on the nodes (representing SH-aLRT/UFB/BIPP, respectively), with support thresholds considered significant at SH-aLRT ≥ 80%, UFB ≥ 90%, and PP ≥ 0.95. In this study, newly described specimens are highlighted in red, type specimens are indicated in bold, and collection localities are marked in blue.
In our phylogenetic study, all seven specimens of the three newly identified species were reliably grouped into three distinct, well-supported clades. Specifically, C. orienticroceus in section Dermocybe, C. orientisanguineus in sect. Sanguinei, and C. subolivaceoluteus in sect. Pauperae. Additionally, the branching into their respective sister clades was highly supported.
Taxonomy
Cortinarius
orienticroceus
Taxon classificationFungiAgaricalesCortinariaceae
Y. Zhou, Y. Zhang & Q. Zhao sp. nov.
28A621EA-6E93-5B33-B861-C8E7FA6935A8
Index Fungorum: IF904774
Fig. 2 Chinese name: 东方黄丝膜菌 (dong fang huang si mo jun)
Etymology.
Croceus means yellow, namely after its type locality from East Asia.
Holotype.
China, • Yunnan, Lijiang, Yulong Naxi Autonomous County, Jiuhe Township 26.630601°N, 99.722506°E; alt. 3236 m; in Abies forest; 28 Jul. 2016, LJ_343 (HKAS 151551).
Macrostructures.
The pileus measures 30–40 mm in diameter; it is campanulate, umbilicate, with a slightly incurved and non-striate margin; the surface is fibrillose-tomentose and non-viscid, appearing dark brown (9F6) at the disc and gradually fading to light brown (9E5) towards the margin, which is pale yellow (5D5). The lamellae are adnexed, moderately crowded, of unequal length, with even edges, and colored pale yellow to bright yellow (5D6). The stipe measures 50–80 mm long × 5–8 mm thick, cylindrical, and central. Its upper and median portions are pale yellow (5D6), while the base is reddish-brown (8B6). Initially silky and shiny, it darkens with maturity, developing slight olivaceous tones (dull pale yellow, 4D8). The veil is inconspicuous. The basal mycelium is light yellow (3A2). No distinct odor is detected from the fruiting body.
Cortinarius orienticroceus. a, b. Mature specimen (a, b. HKAS 151551 holotype); c. SEM micrographs of basidiospores; d. Basidiospores; f, g. Basidia; e, h. Pileipellis hyphae. Scale bar: 2 cm (a, b); 2 μm (c); 10 μm (d); 20 μm (e–g); 50 μm (h).
Microstructures.
Basidiospores [80/2/2] (6–) 6.5–8 (–8.5) × 4.5–6 (–6.5) μm,[Q = (1–) 1.21–1.68 (–2), Q = 1.48 ± 0.15], ellipsoid, moderately verrucose, amber-yellow. Basidia (22.5–) 23–32 × 6–9 μm, 4–spores, turning pale red to red in 10% KOH solution. Cystidia absent. Pileipellis duplex: the epicutis consisting of appressed hyphae 5–15 μm wide, cylindrical and elongated, subparallelly arranged on the epidermis, mostly hyaline and occasionally pale pink in 10% KOH, with clamp connections present; the subpellis not distinctly delimited from the context. Stipitipellis hyphae 7–18 µm wide, hyaline in 10% KOH, with clamp connections.
Habitat.
It forms ectomycorrhizal associations and grows gregariously in coniferous forests dominated by Picea spp. and Abies spp.
Distribution.
Located in northwestern Yunnan, China.
Additional material examined.
China, • Yunnan, Lijiang City, Laojunshan Nature Reserve, 26.630601°N, 99.722506°E; alt. 3846 m, in a forest of Picea spp. and Abies spp., 9 Aug. 2023, YYW_05 (HKAS 151552).
Notes.
The most distinctive feature of Cortinarius orienticroceus is its vibrant fruiting body coloration: the pileus transitions from dark brown at the disc to pale brown or light yellow towards the margin, the lamellae are pale yellow to bright yellow, the stipe is light yellow above and reddish brown below, the basidiospores are ellipsoid and ornamented with verrucose projections. Genetic studies show that C. orienticroceus is most closely related to C. subrufulus and C. hadrocroceus (Fig. 1). In comparison, C. subrufulus features a pileus that is brown to reddish-brown, often with a faint yellowish hue near the margin when young, lamellae are orange to red-orange, and its stipe is pale yellow. C. hadrocroceus has a hemispherical pileus, dark brown at the center and transitioning to brown or reddish-brown at the edges. Its lamellae are moderately spaced, starting as olive-yellow and turning pale olive-brown as they mature. The stipe is cylindrical or slightly bulbous at the base and pale yellow (Niskanen 2014).
Cortinarius
orientisanguineus
Taxon classificationFungiAgaricalesCortinariaceae
Y. Zhou, Y. Zhang & Q. Zhao sp. nov.
4CE5DA59-214D-5BB0-8A89-1E2EC9FDEEB3
Index Fungorum: IF904775
Fig. 3 Chinese name: 东方血红丝膜菌 (dong fang xue hong si mo jun)
Etymology.
Sanguineus means blood-red, namely after its type locality from East Asia.
Holotype.
China • Yunnan, Honghe Hani and Yi Autonomous Prefecture, Pingbian Miao Autonomous County, Dawei Mountain National Forest Park, 22.5306201°N, 103.722405°E, alt. 2354 m, in evergreen broad-leaved forest, 14 Jul. 2020, PB_116 (HKAS 151548).
Macrostructures.
The pileus measures 28–42 mm in diameter, hemispherical, with an initially straight margin then becomes partly erose; the surface exhibits conspicuous fibrillose squamules and is dry. It is reddish-brown (9A8) at the disc, gradually fading to light brown (9A7) toward the margin, which is orange-yellow (5A6). The lamellae are crowded, of unequal length, adnate, and reddish-brown (8C7). The stipe measures 40–60 mm long and 3–4 mm thick, central, slightly bulbous at the base; its upper and median portions are reddish-brown (8C7), while the lower part is orange-yellow (6B6). The veil is evanescent, forming a reddish-brown annulus on the upper to median stipe. The basal mycelium is yellow (3A2).
Cortinarius orientisanguineus. a, b. Mature specimen (a. HKAS 151549; b. HKAS 151548 holotype); c. SEM micrographs of basidiospores; d. Basidiospores; f, g. Basidia; e, h. Pileipellis hyphae. Scale bar: 2 cm (a, b); 2 μm (c); 5 μm (d); 20 μm (e–g); 50 μm (h).
Microstructures.
Basidiospores [120/3/1] (5–) 5.5–7.5 (–8) × 3.5–5.5 (–6) μm [Q = 1–1.5 (–1.65), Q = 1.39 ± 0.14], amygdaliform to ellipsoid, moderately verrucose, yellowish-brown. Basidia (18–)21–32 × 5–8 μm, 4–spores, containing granular contents, hyaline to pale pink in 10% KOH solution. Cystidia absent. Pileipellis exhibits a purplish-red coloration in KOH solution; epicutis well-developed, 8–15 μm wide, highly gelatinized, and the pileipellis exhibits elongate-ellipsoidal elements, closely interwoven and appressed to the epidermal layer, with clamp connections present, hyaline in 10% KOH, the hypodermal hyphae are inconspicuous. Stipitipellis hyphae 5–11 µm wide, hyaline in 10% KOH, with clamp connections.
Habitat.
It forms ectomycorrhizal associations and grows gregariously in evergreen broad-leaved forests.
Distribution.
It is situated in southern Yunnan, specifically in the southeastern part of the Honghe Hani and Yi Autonomous Prefecture.
Additional material examined.
China, • Yunnan, Honghe, Pingbian Miao Autonomous County, Dawei Mountain National Forest Park, 22.8616201°N, 103.523405°E, alt. 2341 m, in evergreen broad-leaved forest, 14 Jul. 2020, PB_68 (HKAS 151549), 15 Jul. 2020, PB_115 (HKAS 151550).
Notes.
Cortinarius orientisanguineus is distinguished by its pileus, which displays a reddish-brown center, transitioning to light brown towards the edges, and finally to orange-yellow at the margin. Its stipe is slightly enlarged at the base, with reddish-brown hues dominating the upper and middle sections, fading to orange-yellow near the base. The lamellae are closely spaced and uneven. The species also features moderately warted basidiospores, which are almond-shaped to elliptical, and basidia with granular contents. Phylogenetic studies indicate that C. orientisanguineus is closely related to the C. sanguineus complex, with the strongest similarity to C. neosanguineus (Fig. 1).
Morphologically, C. neosanguineus differs by having a uniformly bright red pileus, moderately spaced, deep purplish-red lamellae, and a stipe that matches the color of the lamellae (Niskanen et al. 2013). Additionally, the basidiospores of C. neosanguineus are larger, 7–8.5 (9) × 4.5–5. 5 μm. In comparison, C. orientisanguineus has smaller basidiospores, ranging from 5.5–7.5 × 4–5.5 (6) μm.
Cortinarius
subolivaceoluteus
Taxon classificationFungiAgaricalesCortinariaceae
Y. Zhou, Y. Zhang & Q. Zhao sp. nov.
F13A19C1-0350-5972-BFB5-3C070D584806
Index Fungorum: IF904776
Fig. 4 Chinese name: 拟橄榄绿丝膜菌 (ni gan lan lü si mo jun)
Etymology.
Olivaceoluteus means olive-green, named for the pileus color which resembles olive green but is distinctly different.
Holotype.
China, • Yunnan, Chuxiong, Zixishan Provincial Nature Reserve, 25.1301281°N, 101.221405°E; alt. 2502 m, in semi-humid evergreen broad-leaved forest, 18 Aug. 2014. ZXS_01_31 (HKAS 151553)
Macrostructures.
The pileus measures 20**–40 mm in diameter, papillate at the disc when young and becoming applanate at maturity. The disc is tawny (7E7), gradually paling toward the margin, which is pale olivaceous or cinnamomeous (7D5), slightly incurved, and non-striate. The entire surface is fibrillose and dry. The lamellae are moderately crowded to moderately spaced, unequal in length, and cinnamomeous to pale olivaceous (7F8). The stipe is 30–**45 mm long, cinnamomeous (7F8) on the upper and median parts, with reddish-brown (8D4) fibrils and patches below, and has a distinctly bulbous base. The veil is evanescent, cinnamomeous or pale olivaceous (7F8). The basal mycelium is pale grayish-white (7A1).
Cortinarius subolivaceoluteus. a, b. Mature specimen (a. HKAS 151553 holotype; b. HKAS 151554); c. SEM micrographs of basidiospores; d. Basidiospores; e, f. Basidia; g. Pileipellis hyphae. Scale bar: 2 cm (a, b); 2 μm (c); 10 μm (d); 20 μm (e, f); 50 μm (h).
Microstructures.
Basidiospores [80/2/2] (5.64–) 6–7.5 × 3.5–4.5 (–5) μm [Q = 1–1.5 (–1.65), Q = 1.39 ± 0.16], ellipsoid, moderately verrucose, amber-yellow. Basidia 18–27 (–28) × 6–8 µm, 4–spores, hyaline to pale pink in 10% KOH solution. Pileipellis duplex: epicutis consisting of cylindrical to inflated, appressed hyphae 7–11 µm wide, hyaline in 10% KOH, the hypodermal hyphae 14.5–20 μm wide, broadly ellipsoid. Stipitipellis hyphae 10–14 µm wide, hyaline in 10% KOH, with clamp connections present.
Habitat.
It forms ectomycorrhizal associations and grows gregariously in semi-humid evergreen broad-leaved forests.
Distribution.
It is situated in the western part of Chuxiong City, Yunnan, China.
Additional material examined.
China, • Yunnan, Dali, Yongping County, Jinguangsi Provincial Nature Reserve, 25.0013201°N, 99.172805°E, alt. 2441 m, in evergreen broad-leaved forest, 7 Sep. 2009, JGS_22_35 (HKAS 151554).
Notes.
Cortinarius subolivaceoluteus is distinguished by its pileus which displays a tawny central disc that gradually transitions to a paler margin, exhibiting pale olivaceous to cinnamomeous coloration, the lamellae are cinnamomeous-colored, moderately spaced, and exhibit variable lengths, the stipe presents striking color variation: the upper and middle portions are cinnamomeous, while the lower section is decorated with reddish-brown fibrillose structures and patches, terminating in a prominently bulbous base. The species possesses an evanescent, pale olivaceous partial veil.
Phylogenetic analyses demonstrate that C. subolivaceoluteus forms a sister lineage to C. olivaceoluteus (Fig. 1). Morphologically, C. olivaceoluteus exhibits a uniformly cinnamomeous pileus ornamented with fine fibrillose squamules, densely arranged olive-yellow lamellae, and a clavate stipe that is concolorous with the lamellae and lacks surface squamulation (Niskanen 2014). Additionally, spore morphometry provides a reliable diagnostic character, C. olivaceoluteus basidiospores 7–8(–8.5) × 4–5(–5.5) μm, while C. subolivaceoluteus has smaller basidiospores6–7.5 × (3.5–)4–4.5(–5) μm.
Discussion
China’s southwestern region harbors abundant fauna, flora, and fungal diversity. Over the past several decades, researchers have documented numerous novel fungal taxa from this biogeographically significant area (Yu et al. 2024; Dang et al. 2025; Wang et al. 2025; Xie et al. 2025a, b; Xu et al. 2025; Yang et al. 2025). However, the subgenus Dermocybe within the genus Cortinarius has received comparatively limited taxonomic attention, resulting in a substantial knowledge gap regarding the species diversity of Cortinarius subg. Dermocybe in this region. This study introduces three newly identified species within Cortinarius subg. Dermocybe: C. orienticroceus, C. orientisanguineus, and C. subolivaceoluteus. These discoveries are supported by a detailed morphological examination and robust molecular phylogenetic analyses. The identification of these species significantly expands the known diversity of the subgenus and offers valuable insights into the subtle differences and evolutionary relationships among its various groups.
Morphologically, C. orienticroceus stands out due to its strikingly multicolored pileus, which is dark brown at the center, transitioning to light brown and culminating in a pale yellow margin, paired with a notably long stipe. It closely resembles C. subrufulus, though the latter displays a more vibrantly colored pileus and narrower spores. In contrast, C. orientisanguineus is distinct with its reddish-brown center fading to light brown with an orange-yellow margin, densely spaced lamellae, and a stipe that changes color. These traits clearly differentiate it from the similar C. sanguineus, supported by a consistent set of features. Additionally, C. orientisanguineus has smaller spores and basidia compared to its nearest relative, C. neosanguineus. Meanwhile, although C. subolivaceoluteus shows brighter hues, it can be reliably separated from C. olivaceoluteus by the latter’s unique olive tones and larger spores. These observations align with the taxonomic framework proposed by Moser et al. (1983), emphasizing that consistent microscopic differences, such as spore size and surface patterns, are essential for distinguishing species within this group.
Currently, six recognized sections exist within the subg. Dermocybe: sect. Dermocybe, sect. Aureifolii, sect. Cruentoides, sect. Malicoriae, sect. Pauperae, and sect. Sanguinei (Liimatainen et al. 2022). Based on molecular phylogenetic analysis, the newly discovered species Cortinarius orienticroceus, C. orientisanguineus, and C. subolivaceoluteus belong to sect. Dermocybe, sect. Sanguinei, and sect. Pauperae, respectively. Species in subg. Dermocybe has considerable morphological variation in its fruiting bodies, ranging from small to medium-sized specimens. The pileus may be dry or moist, occasionally exhibiting hygrophanous properties or a sticky surface, with textures ranging from smooth to fibrillose or scaly. Molecular phylogenetic analyses have fundamentally transformed our understanding of this group’s taxonomic classification. Previously recognized as an independent genus based solely on morphological characteristics, subg. Dermocybe has been conclusively demonstrated, based on molecular evidence, to constitute a natural, monophyletic lineage within the genus Cortinarius, thereby confirming its evolutionary unity (Liu et al. 1997; Peintner et al. 2004; Garnica et al. 2005). Comprehensive molecular investigations of section Sanguinei have yielded particularly significant findings, not only validating its monophyletic status but also revealing substantially greater species diversity than morphology-based classifications had previously indicated (Niskanen et al. 2013). Conversely, section Pauperae exemplifies the numerous traditional morphology-based taxonomic groups within this subgenus that await molecular validation. The monophyletic nature and legitimate taxonomic standing of this section remain unresolved, underscoring the need for comprehensive type-specimen-based molecular phylogenetic research to reassess the many morphologically circumscribed groups within species-rich genera such as Cortinarius (Liimatainen 2013).
The taxonomic investigation of Subg. Dermocybe within China remains in its incipient phase (Yuan et al. 2020; Dang et al. 2025; Xie et al. 2025a), presenting substantial lacunae in our current understanding of this mycological assemblage. This nascent state of knowledge consequently presents considerable opportunities for advancing systematic mycology. Future investigations of Dermocybe should incorporate phylogenomic approaches to elucidate cryptic taxonomic diversity, thereby facilitating comprehensive systematic revision and enabling the description of novel taxa to augment the known biodiversity within this cortinarioid lineage.
Supplementary Material
XML Treatment for Cortinarius orienticroceus
XML Treatment for Cortinarius orientisanguineus
XML Treatment for Cortinarius subolivaceoluteus
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bidaud A, Moënne-Loccoz P, Reumaux P (1994) Atlas des Cortinaires. Cle generale des sous-genres, sections et series. Fédérat. Mycol. Dauphiné-Savoie, 102 pp.
- 2Brandrud TE, Lindström H, Marklund H, Melot J, Muskos S (1998) Cortinarius Flora Photographica I. Cortinarius HB, Sweden, 38 pp.
- 3Dang Y, Zhu M, Keyhani NO, Wu Z, Qiu C, Xiong J, Pu H, Lin L, Xiong C, Zhao Z, Zheng M, Mu T, Lin Y, Huang Y, Yuan X, Qiu C, Ji X, Qiu J, Chen Y (2025) Discovery of two new Cortinarius species in Southern China. Frontiers in Microbiology 16: e 1558935. 10.3389/fmicb.2025.1558935 PMC 1213649040469734 · doi ↗ · pubmed ↗
- 4Danks M, Lebel T, Vernes K (2010) ‘Cort short on a mountaintop’–Eight new species of sequestrate Cortinarius from sub-alpine Australia and affinities to sections within the genus. Persoonia 24(1): 106–126. 10.3767/003158510 X 512711 PMC 289016320664764 · doi ↗ · pubmed ↗
- 5Fan L, Zhong X, Ma T, Zhou H, Wang B, Ji X (2024) Four new species of Cortinariaceae (Agaricales) from Northwestern China. Frontiers in Microbiology 15: e 1454736. 10.3389/fmicb.2024.1454736 PMC 1146135539386370 · doi ↗ · pubmed ↗
- 6Filippova N, Zvyagina E, Rudykina E, Dobrynina A, Bolshakov S (2023) The diversity of macromycetes in peatlands: Nine years of plot-based monitoring and barcoding in the raised bog“Mukhrino”, West Siberia. Biodiversity Data Journal 11: e 105111. 10.3897/BDJ.11.e 105111 PMC 1061211537901679 · doi ↗ · pubmed ↗
- 7Gardes M, Bruns TD (1993) ITS primers with enhanced specificity for basidiomycetes‐application to the identification of mycorrhizae and rusts. Molecular Ecology 2(2): 113–118. 10.1111/j.1365-294X.1993.tb 00005.x 8180733 · doi ↗ · pubmed ↗
- 8Garnica S, Weiß M, Oertel B, Oberwinkler F (2005) A framework for a phylogenetic classification in the genus Cortinarius (Basidiomycota, Agaricales) derived from morphological and molecular data. Botany 83(11): 1457–1477. 10.1139/b 05-107 · doi ↗