Three new species of Picipes (Polyporales, Basidiomycota) from China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
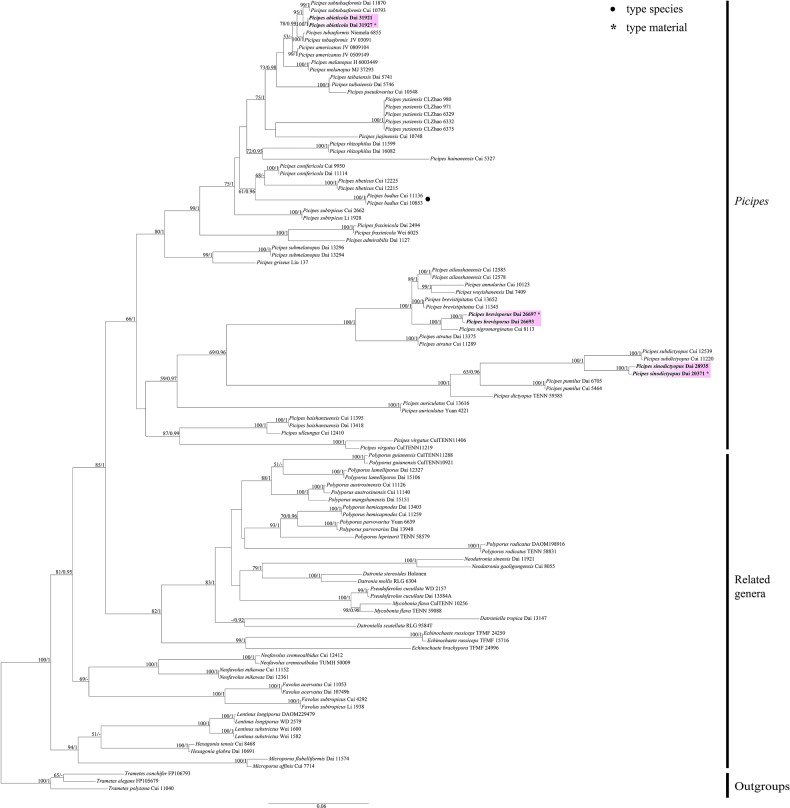 Figure 1
Figure 1 Figure 2
Figure 2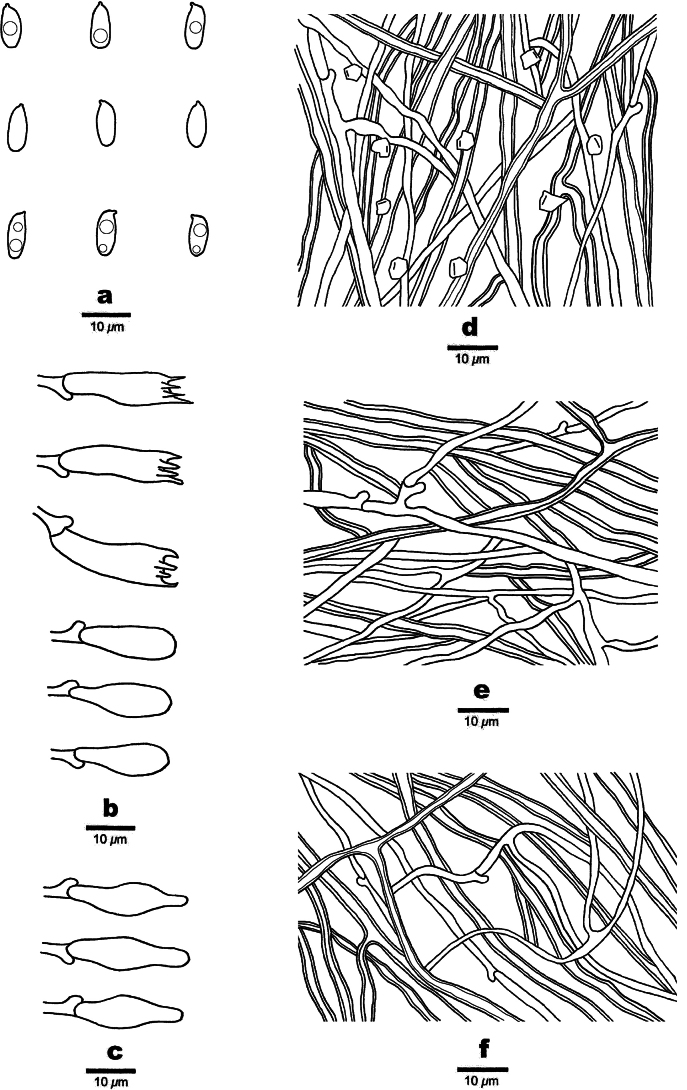 Figure 3
Figure 3 Figure 4
Figure 4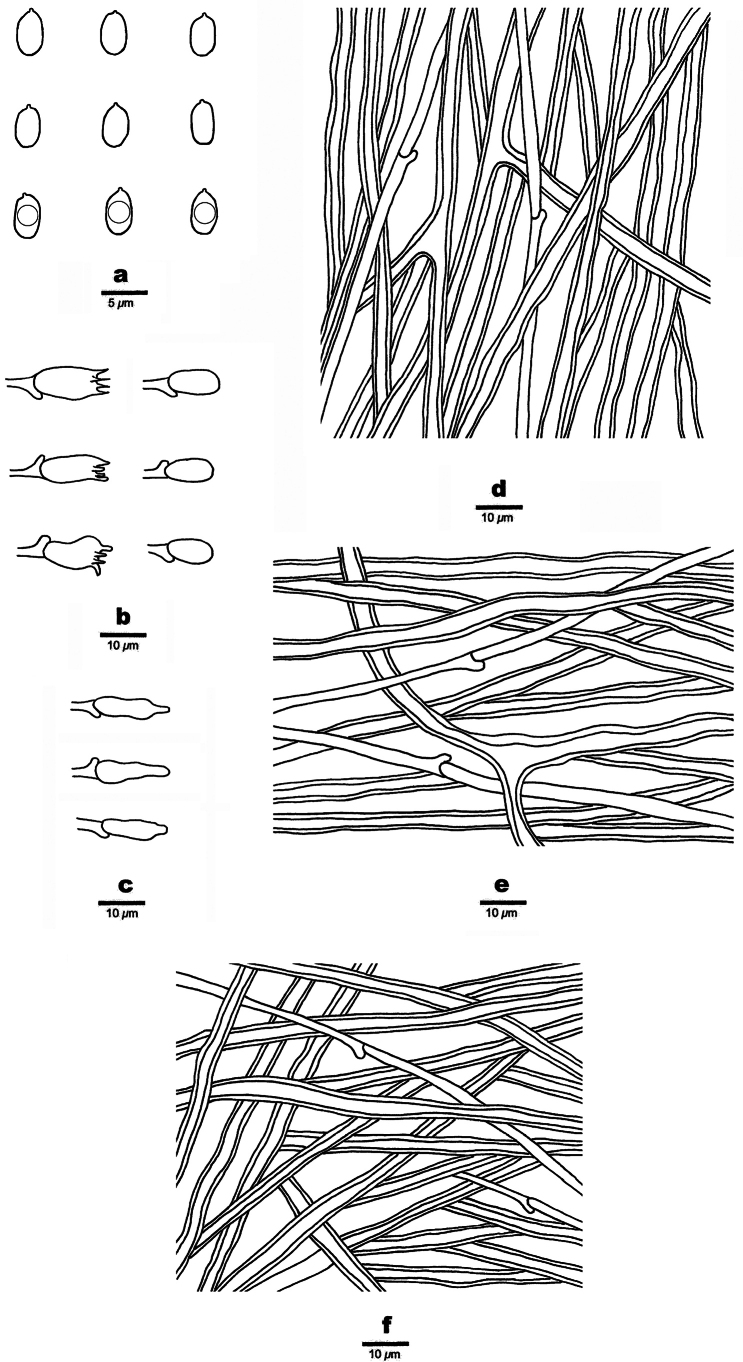 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7| Species | Sample No. | GenBank Accession No. | References | |
|---|---|---|---|---|
|
|
| |||
|
| RLG 6304 |
|
|
|
|
| Holonen |
|
|
|
|
| RLG 9584 T |
|
|
|
|
| Dai 13147 |
|
|
|
|
| TFMF 24996 |
|
|
|
|
| TFMF 15716 |
|
|
|
|
| TFMF 24250 |
|
|
|
|
| Cui 11053 |
|
|
|
|
| Dai 10749b |
|
|
|
|
| Cui 4292 |
|
|
|
|
| Li 1938 |
|
|
|
|
| Dai 10691 |
|
|
|
|
| Cui 8468 |
|
|
|
|
| DAOM229479 |
|
|
|
|
| WD2579 |
|
|
|
|
| Wei 1582 |
|
|
|
|
| Wei 1600 |
|
|
|
|
| Cui 7714 |
|
|
|
|
| Dai 11574 |
|
|
|
|
| CulTENN 10256 |
|
|
|
|
| TENN 59088 |
|
|
|
|
| Cui 8055 |
|
|
|
|
| Dai 11921 |
|
|
|
|
| Cui 12412 |
|
|
|
|
| TUMH50009 |
|
|
|
|
| Cui 11152 |
|
|
|
|
| Dai 12361 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| Dai 1127 |
| — | Unpublished |
|
| Cui 12578 |
|
|
|
|
| Cui 12585 |
|
|
|
|
| JV 0509-149 |
|
|
|
|
| JV 0809-104 |
|
|
|
|
| Cui 10123 |
|
|
|
|
| Cui 11289 |
|
|
|
|
| Dai 13375 |
|
|
|
|
| Cui 13616 |
|
|
|
|
| Yuan 4221 |
|
|
|
|
| Cui 10853 |
|
|
|
|
| Cui 11136 |
|
|
|
|
| Cui 11395 |
|
|
|
|
| Dai 13418 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| Cui 11345 |
|
|
|
|
| Cui 13652 |
|
|
|
|
| Cui 9950 |
|
|
|
|
| Dai 11114 |
|
|
|
|
| TENN 59385 |
|
|
|
|
| Dai 2494 |
|
|
|
|
| Wei 6025 |
|
|
|
|
| Liu 137 |
|
|
|
|
| Cui 5327 |
| — |
|
|
| Cui 10748 |
|
|
|
|
| H 6003449 |
|
|
|
|
| MJ 372-93 |
|
|
|
|
| Cui 8113 |
|
|
|
|
| Cui 10548 |
|
|
|
|
| Cui 5464 |
|
|
|
|
| Dai 6705 |
|
|
|
|
| Dai 11599 |
|
|
|
|
| Dai 16082 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| Cui 11220 |
|
|
|
|
| Cui 12539 |
|
|
|
|
| Dai 13294 |
|
|
|
|
| Dai 13296 |
|
|
|
|
| Cui 2662 |
|
|
|
|
| Li 1928 |
|
|
|
|
| Cui 10793 |
|
|
|
|
| Dai 11870 |
|
|
|
|
| Dai 5741 |
|
|
|
|
| Dai 5746 |
|
|
|
|
| Cui 12215 |
|
|
|
|
| Cui 12225 |
|
|
|
|
| Niemela 6855 |
|
|
|
|
| JV 0309-1 |
|
|
|
|
| Cui 12410 |
|
|
|
|
| CulTENN11219 |
|
|
|
|
| CulTENN11406 |
|
|
|
|
| CLZhao 971 |
|
|
|
|
| CLZhao 980 |
|
|
|
|
| CLZhao 6329 |
| — |
|
|
| CLZhao 6332 |
| — |
|
|
| CLZhao 6735 |
| — |
|
|
| Dai 7409 |
|
|
|
|
| Cui 11140 |
|
|
|
|
| Cui 11126 |
|
|
|
|
| TENN 58404 |
|
|
|
|
| TENN 59093 |
|
|
|
|
| Cui 11259 |
|
|
|
|
| Dai 13403 |
|
|
|
|
| Dai 15106 |
|
|
|
|
| Dai 12327 |
|
|
|
|
| TENN 58579 |
|
|
|
|
| Dai 15151 |
|
|
|
|
| Yuan 6639 |
|
|
|
|
| Dai 13948 |
|
|
|
|
| DAOM198916 |
|
|
|
|
| TENN 58831 |
|
|
|
|
| Dai 13584A |
|
|
|
|
| WD 2157 |
|
|
|
|
| FP106793sp |
|
|
|
|
| FP105679sp |
|
|
|
|
| Cui 11040 |
|
|
|
| Species Name | Marker | Top Match Species | GenBank Accession No. | Max Score | Total Score | Query Cover | E-value | Identity |
|---|---|---|---|---|---|---|---|---|
|
|
|
|
| 1002 | 1002 | 90% | 0.0 | 97.77% |
|
|
|
| 2459 | 2459 | 100% | 0.0 | 99.63% | |
|
|
|
|
| 889 | 889 | 88% | 0.0 | 92.00% |
|
|
|
| 1411 | 1411 | 99% | 0.0 | 98.74% | |
|
|
|
|
| 767 | 767 | 77% | 0.0 | 92.74% |
|
|
|
| 2357 | 2357 | 99% | 0.0 | 97.67% |
| 1 | Growing on grass roots |
|
| – | Growing on woods or ground |
|
| 2 | Generative hyphae bearing simple septa |
|
| – | Generative hyphae bearing clamp connections |
|
| 3 | Pores 2–3 per mm; basidiospores 8–10 × 3–3.9 µm |
|
| – | Pores 5–6 per mm; basidiospores 6.5–8 × 3–3.8 µm |
|
| 4 | Growing on coniferous woods |
|
| – | Growing on hardwoods or monocotyledon |
|
| 5 | Pores 2–5 per mm; cystidioles absent |
|
| – | Pores ≥ 6 per mm; cystidioles present |
|
| 6 | Basidiospores cylindrical, usually > 6 µm in length |
|
| – | Basidiospores oblong to cylindrical, usually < 6 µm in length |
|
| 7 | Pores 7–10 per mm; stipe slender, up to 5 cm |
|
| – | Pores 5–7 per mm; stipe short, up to 1.5 cm |
|
| 8 | Basidiospores mainly oblong; growing in plateau temperate regions |
|
| – | Basidiospores mainly cylindrical; growing in subtropical regions |
|
| 9 | Pileal surface grey |
|
| – | Pileal surface other colour rather than grey |
|
| 10 | Pores ≥ 7 per mm |
|
| – | Pores < 7 per mm |
|
| 11 | Cystidioles absent |
|
| – | Cystidioles subulate |
|
| 12 | Pileal surface azonate |
|
| – | Pileal surface zonate |
|
| 13 | Growing on monocotyledon |
|
| – | Growing on angiosperm |
|
| 14 | Stipe up to 1.5 cm long and 1 cm in diam. |
|
| – | Stipe up to 5 mm long and 4 mm in diam. |
|
| 15 | Cystidioles absent |
|
| – | Cystidioles present |
|
| 16 | Stipe central |
|
| – | Stipe lateral |
|
| 17 | Basidiospores with one to two guttules |
|
| – | Basidiospores without guttules |
|
| 18 | Stipe without a black cuticle |
|
| – | Stipe with a black cuticle |
|
| 19 | Stipe up to 4 mm long and 3.5 mm in diam. |
|
| – | Stipe up to 2 cm long and 1.5 cm in diam. |
|
| 20 | Basidia ≥ 25 µm in length |
|
| – | Basidia < 25 µm in length |
|
| 21 | Pores circular to subcircular |
|
| – | Pores angular |
|
| 22 | Stipe up to 8 cm |
|
| – | Stipe up to 2.5 cm |
|
| 23 | Cystidioles absent |
|
| – | Cystidioles present |
|
| 24 | Stipe without a black or terra-brown cuticle |
|
| – | Stipe with a black or terra-brown cuticle |
|
| 25 | Stipe slender |
|
| – | Stipe short or with a flattened base |
|
| 26 | Pore surface grey-beige or pearl-beige upon drying |
|
| – | Pore surface tan or buff upon drying |
|
| 27 | Hyphae in cuticle bearing clamp connections |
|
| – | Hyphae in cuticle with simple septa |
|
| 28 | Basidia < 16 µm in length |
|
| – | Basidia > 16 µm in length |
|
- —National Natural Science Foundation of China 501100001809 https://ror.org/01h0zpd94 http://doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Fungal Biology and Applications · Building materials and conservation
Introduction
Macrofungi are important natural resources, the diversity of the Chinese macrofungi has been extensively studied and a large number of species new to science and many taxa new to China have been recently discovered (Wang et al. 2021, 2023, 2024; Zhang et al. 2023; Zhou et al. 2023, 2026; Dong et al. 2024, 2025; Hongsanan et al. 2025; Song et al. 2025; Wijesinghe et al. 2025; Yang et al. 2025; Zhao et al. 2025).
Polypores, as a major group of macrofungi, play important ecological roles in forest ecosystems (Hibbett et al. 2014), in which 1337 species have been found in the Northern Hemisphere (Justo et al. 2017; Wu et al. 2022a) and 1902 species in the tropical forests of Africa, America and Asia (Zhao et al. 2024). In addition, a total of 4,026 polypores has been found worldwide, suggesting its extremely high species diversity (Zhao et al. 2026).
Recently, on the basis of studying the lentinoid and polyporoid fungi, Zmitrovich and Kovalenko (2016) described a new genus Picipes Zmitr. & Kovalenko typified by Pi. badius (Pers.) Zmitr. & Kovalenko (= Polyporus badius) according to analyses of nLSU, ITS and EF1-α sequences. This genus is characterised by the following features: basidiomata annual, stipitate, hymenophore polyporoid; pilei infundibuliform, covered with a cuticle, without scales, smoke-grey to castaneous or deeply brown; stipe covered with brownish to black cuticle; pores small (more than 5 per mm); hyphal system dimitic with skeletal-binding hyphae; clamp connections present in most species; basidiospores cylindrical to oblong ellipsoid, smooth, hyaline, thin-walled, inamyloid, acyanophilous; on both angiosperm and gymnosperm wood, causing a white rot. Polyporus badius, P. melanopus (Pers.) Fr. and P. tubaeformis (P. Karst.) Ryvarden & Gilb. were segregated from Polyporus and transferred into Picipes as Pi. badius, Pi. melanopus (Pers.) Zmitr. & Kovalenko and Pi. tubaeformis (P. Karst.) Zmitr. & Kovalenko (Zmitrovich and Kovalenko 2016).
Based on multi-gene phylogenetic analyses, species in Polyporus group were proved to be distributed in two different clades viz. picipes clade and squamosus clade (Zhou et al. 2016). The picipes clade has been described as Picipes Zmitr. et Kovalenko (Zmitrovich and Kovalenko 2016) and sixteen species with hard basidiomata in dried condition, brownish to black stipes and strongly branched skeleto-binding hyphae were included in this genus (Zhou et al. 2016). Based on the MycoBank database (http://www.mycobank.org, accessed on 6 March 2026) and the Index Fungorum (http://www.indexfungorum.org, accessed on 6 March 2026), 32 species have been accepted in the genus Picipes (Zhou et al. 2016; Zmitrovich and Kovalenko 2016; Cui et al. 2019; Bhunjun et al. 2022; Ji et al. 2022; Liu et al. 2024).
In the current study, the species diversity of Polyporus**sensu lato in China was investigated, phylogenetic analyses, based on ITS + nLSU were carried out and three new species matching the concept of Picipes are described and illustrated.
Materials and methods
Morphological studies
The studied six specimens were collected from nature forests, including Guangxi Autonomous Region, Xizang Autonomous Region and Yunnan Province of China. These are deposited in the Fungarium of the Institute of Microbiology, Beijing Forestry University (BJFC, China). Morphological descriptions are based on field notes and voucher specimens. The microscopic analysis follows Spirin et al. (2017) and Wu et al. (2022b). Freehand sections were made from dried basidiomata and mounted in 2% (w/v) potassium hydroxide (KOH) to observe colour changes. Sections were studied at a magnification of up to 1000× using a Nikon Eclipse 80i microscope and phase contrast illumination. Microscopic features and measurements were made from slide preparations stained with Cotton Blue and Melzer’s reagent. To represent the variation in the size of spores, 5% of measurements were excluded from each end of the range and are given in parentheses. In the description: KOH = 5% potassium hydroxide, IKI = Melzer’s reagent, IKI– = neither amyloid nor dextrinoid, CB = Cotton Blue, CB+ = cyanophilous in Cotton Blue, CB– = acyanophilous in Cotton Blue, L = arithmetic average of spore length, W = arithmetic average of spore width, Q = L/W ratios and n = number of basidiospores measured from a given number of specimens. Colour terms follow Anonymous (1969) and Petersen (1996).
DNA extraction, amplification and sequencing
A CTAB rapid plant genome extraction kit-DN14 (Aidlab Biotechnologies Co., Ltd, Beijing) was used to obtain DNA from dried specimens and to perform the polymerase chain reaction (PCR) according to the manufacturer’s instructions with some modifications (Shen et al. 2019; Sun et al. 2020). The internal transcribed spacer (ITS) and large subunit nuclear ribosomal RNA gene (nLSU) were amplified using the primer pairs ITS5/ITS4 and LR0R/LR7 (White et al. 1990; Hopple and Vilgalys 1999, https://sites.duke.edu/vilgalyslab/rdna_primers_for_fungi/).
The PCR procedure for ITS was as follows: initial denaturation at 95 °C for 3 min, followed by 34 cycles at 94 °C for 40 s, annealing at 54 °C for 45 s and extension at 72 °C for 1 min, with a final extension of 72 °C for 10 min. The PCR procedure for nLSU was as follows: initial denaturation at 94 °C for 1 min, followed by 34 cycles of denaturation at 94 °C for 30 s, annealing at 50 °C for 1 min and extension at 72 °C for 1.5 min, with a final extension at 72 °C for 10 min. The PCR products were purified and sequenced at the Beijing Genomics Institute (BGI, China), with the same primers. The newly-generated sequences were deposited in GenBank (Sayers et al. 2024) and listed in Table 1.
Sequences generated were aligned manually with additional sequences downloaded from GenBank using AliView version 1.27 (Larsson 2014). The final ITS and nLSU datasets were subsequently aligned using MAFFT v.7 under the E-INS-i strategy with no cost for opening gaps and equal cost for transformations (command line: mafft –genafpair –maxiterate 1000, Katoh and Standley (2013) and visualised in AliView. Alignments were spliced and transformed formats in Mesquite v.3.2. (Maddison and Maddison 2017). Multiple sequence alignments were trimmed by trimAI v.1.2 using the -htmlout-gt 0.8 -st option to deal with gaps, when necessary (Capella-Gutierrez et al. 2009).
Phylogenetic analyses
The two-marker DNA multiple sequence alignment (ITS + nLSU) was used to determine the phylogenetic position of the new species, with Trametes conchifer (Schwein.) Pilát, T. elegans (Spreng.) Fr. and T. polyzona (Pers.) Justo as the outgroup (Ji et al. 2022). The phylogenetic analyses followed the approach of Han et al. (2015) and Zhu et al. (2019). Maximum Likelihood (ML) and Bayesian Inference (BI) analyses were performed, based on the ITS + nLSU datasets.
Sequences were analysed using Maximum Likelihood (ML) with RAxML v.8.2.10 (Stamatakis 2014). Branch supports for all parsimony analyses were estimated by performing 1,000 bootstrap replicates with a heuristic search of 10 random-addition replicates for each bootstrap replicate. Bayesian phylogenetic inference and Bayesian Posterior Probabilities (BPP) were computed with MrBayes 3.2.6 with a GTR + I + G model of DNA substitution and a gamma distribution rate variation across sites (Ronquist and Huelsenbeck 2003). Four Markov chains were run for 1 million generations (two-marker dataset) until the split deviation frequency value was less than 0.01 and trees were sampled every 100 generations. The first 25% of the sampled trees were discarded as burn-in and the remaining ones were used to reconstruct a majority rule consensus and calculate Bayesian Posterior Probabilities (BPP) of the clades. All trees were viewed in FigTree v. 1.4.3 (http://tree.bio.ed.ac.uk/software/figtree/). Branches that received bootstrap support for ML (≥ 75% (ML-BS)) and BPP (≥ 0.95 BPP) were considered as significantly supported. The ML bootstrap (ML) ≥ 50% and BBP (BPP) ≥ 0.90 are presented on topologies from ML analysis, respectively.
Results
Molecular phylogeny
The combined two-marker dataset (ITS + nLSU) included sequences from 108 samples representing 65 taxa. The phylogenetic reconstruction performed with Maximum Likelihood (ML) and Bayesian Inference (BI) analyses for the combined dataset showed similar topology and few differences in statistical support. The best model-fit applied in the Bayesian analysis was GTR + I + G, lset nst = 6, rates = invgamma and prset statefreqpr = dirichlet (1, 1, 1, 1). Bayesian analysis and ML analysis resulted in a similar topology to the MP analysis, with an average standard deviation of split frequencies of 0.006865 (BI).
The phylogenetic tree inferred from the ITS + nLSU sequences indicated that the three new species belonged to Picipes (Fig. 1). In addition, Picipes abieticola grouped together with P. subtubaeformis J.L. Zhou & B.K. Cui and P. tubaeformis with high support (ML = 100, BPP = 1.00); P. brevisporus grouped together with P. nigromarginatus B.K. Cui, X. Ji & J.L. Zhou with a high support (ML = 100, BPP = 1.00); and P. sinodictyopus was sister to P. subdictyopus (H. Lee et al.) B.K. Cui, X. Ji & J.L. Zhou with high support (ML = 100, BPP = 1.00).
ML analysis of Picipes, based on dataset of ITS + nLSU. ML bootstrap values equal to or higher than 50% and Bayesian posterior probabilities values equal to or higher than 0.90 are shown. New taxa are in bold, * represents type material.
Based on the ITS and nLSU sequences of the type material, BLAST queries against the authoritative sequences in NCBI yielded alignments, which are shown in Table 2.
Taxonomy
Picipes
abieticola
Taxon classificationFungiPolyporalesPolyporaceae
K.Y. Luo, Y.C. Dai & Y.J. Cui sp. nov.
3876E9CC-E238-5714-A364-2239B5705B79
861751
Holotype.
China • Xizang Autonomous Region, Yadong County, Pasha Falls to Yadong Customs. GPS coordinates: 27°25'N, 88°55'E; altitude: 2800 m a.s.l. On fallen branch of Abies fabri, 18 October 2024, Dai 31927 (BJFC052186).
Basidiomata of Picipes abieticola (Holotype, Dai 31927). Scale bar: 2 cm.
Diagnosis.
Picipes abieticola is identified by eccentrically stipitate, fan-shaped pilei, circular to subcircular pores (5–7/mm); a dimitic hyphal structure and cylindrical basidiospores with dimensions of 7.5–9.3 × 2.8–3.5 µm.
Microscopic structures of Picipes abieticola (Holotype, Dai 31927). a. Basidiospores; b. Basidia and basidioles; c. Cystidioles; d. Hyphae from trama; e. Hyphae from context; f. Hyphae from stipe.
Etymology.
Abieticola (Lat.) refers to the species inhabiting Abies.
Description.
Basidiomata annual, eccentrically stipitate, solitary, coriaceous and without odour or taste when fresh, hard corky when dry. Pilei fan-shaped, projecting up to 7 cm long, 5 cm wide, and 2 mm thick at base. Pileal surface blackish-red to black, darkening to more or less the centre when fresh, becoming brownish-red to yellowish-brown towards the margin when dry, glabrous, with radially aligned stripes; margin incurved upon drying. Pore surface white when fresh, light ivory to buff when dry, shining; pores circular to subcircular, 5–7 per mm; dissepiments thin, entire. Context light ivory to buff when dry, hard corky upon drying, up to 0.5 mm thick. Tubes concolorous with context, up to 1.5 mm long. Stipe very short, bearing a black cuticle, up to 1.5 cm long and 8 mm in diam.
Hyphal structure: Hyphal system dimitic; generative hyphae bearing clamp connections; skeletal-binding hyphae IKI–, slightly CB+; tissues unchanged in KOH.
Context: Generative hyphae infrequent, colourless, thin-walled, moderately branched, 2–3 μm in diam.; skeletal-binding hyphae dominant, colourless, thick-walled with a wide to narrow lumen, occasionally branched, interwoven, 3–4 μm in diam.
Tubes: Generative hyphae infrequent, colourless, thin-walled, occasionally branched, 1.8–2.7 μm in diam.; skeletal-binding hyphae colourless, thick-walled with a wide to narrow lumen, occasionally branched, interwoven, 3–4 μm in diam. Irregular crystals present. Cystidia absent, cystidioles subulate, 20–25 × 4–6 μm. Basidia mostly clavate, with a basal clamp connection and four sterigmata, 17–25 × 5–8 μm; basidioles in shape similar to basidia, but slightly smaller.
Stipe: Generative hyphae infrequent, colourless, thin-walled, moderately branched, 1.5–2 μm in diam.; skeletal-binding hyphae dominant, colourless, thick-walled with a wide to narrow lumen, moderately branched, interwoven, 3.5–5.5 μm in diam.
Basidiospores—Cylindrical, hyaline, thin-walled, smooth, some with one or two guttules, IKI–, CB–, (7–)7.5–9.3(–9.5) × (2.7–)2.8–3.5(–3.8) µm, L = 8.51 µm, W = 3.18 µm, Q = 2.64–2.72 (n = 60/2).
Type of rot.
White rot.
Additional specimen (paratype) examined.
China • Xizang Autonomous Region, Yadong County, Pasha Falls to Yadong Customs. GPS coordinates: 27°25'N, 88°55'E; altitude: 2800 m a.s.l. On fallen branch of Abies fabri, 18 October 2024, Dai 31921 (BJFC052180).
Picipes
brevisporus
Taxon classificationFungiPolyporalesPolyporaceae
K.Y. Luo, Y.C. Dai, Y.J. Cui & Xin Zhang sp. nov.
6BCC9C04-6205-5D0F-B266-330500A0227B
861752
Holotype.
China • Xizang Autonomous Region, Motuo County, the road 219 from Motuo County to Jiefang Bridge. GPS coordinates: 29°18'N, 95°18'E; altitude: 1000 m a.s.l. On dead Miscanthus, 24 October 2023, Dai 26697 (BJFC 044247).
Basidiomata of Picipes brevisporus (Holotype, Dai 26697). Scale bar: 1 cm.
Diagnosis.
Picipes brevisporus is identified by laterally stipitate, glabrous, with radially aligned stripes pileal surface, short stipe; bearing clamp connections on generative hyphae, subulate cystidioles and oblong ellipsoid basidiospores with dimensions of 3.8–5.7 × 2.3–3.1 µm.
Microscopic structures of Picipes brevisporus (Holotype, Dai 26697). a. Basidiospores; b. Basidia and basidioles; c. Cystidioles; d. Hyphae from trama; e. Hyphae from context; f. Hyphae from stipe.
Etymology.
Brevisporus (Lat.) refers to the species having short basidiospores in the genus Picipes.
Description.
Basidiomata annual, laterally stipitate, solitary, coriaceous and without odour or taste when fresh, hard corky when dry. Pilei auricular to fan-shaped, about 1–1.5 cm long, 0.4–0.6 cm wide and up to 1 mm thick at base. Pileal surface yellow-brownish to brownish when fresh, becoming brown to black towards the margin when dry, glabrous, with radially aligned stripes; margin straight. Pore surface bluish-grey when fresh, light ivory to buff when dry; pores circular to subcircular, 7–9 per mm; dissepiments thin, entire. Context light ivory to buff when dry, hard corky upon drying, up to 0.5 mm thick. Tubes concolorous with pore surface, up to 0.5 mm long. Stipe short, bearing a black cuticle, up to 2 mm long and 2 mm in diam.
Hyphal structure: Hyphal system dimitic; generative hyphae bearing clamp connections; skeletal hyphae IKI–, slightly CB+; tissues unchanged in KOH.
Context: Generative hyphae infrequent, colourless, thin-walled, rarely branched, 2–3 μm in diam.; skeletal-binding hyphae dominant, colourless, thick-walled, occasionally branched, interwoven, 3.5–4.5 μm in diam.
Tubes: Generative hyphae dominant, colourless, thin-walled, rarely branched, 2–3 μm in diam.; skeletal-binding hyphae colourless, thick-walled, moderately branched, interwoven, 3–4.5 μm in diam. Cystidia absent, cystidioles subulate, 12–16 × 3–4 μm. Basidia more or less barrel-shaped, with a basal clamp connection and four sterigmata, 10–15 × 5–7 μm; basidioles in shape similar to basidia, but slightly smaller.
Stipe: Generative hyphae infrequent, colourless, thin-walled, rarely branched, 2–3 μm in diam.; skeletal-binding hyphae dominant, colourless, thick-walled, rarely branched, interwoven, 3.5–4.5 μm in diam.
Basidiospores: Short oblong ellipsoid, hyaline, thin-walled, smooth, some with one large guttule, IKI–, CB–, (3.5–)3.8–5.7(–6.6) × (2.0–)2.3–3.1(–3.4) µm, L = 4.75 µm, W = 2.67 µm, Q = 1.71–1.85 (n = 60/2).
Type of rot.
White rot.
Additional specimen (paratype) examined.
China • Xizang Autonomous Region, Motuo County, the Road 219 from Motuo County to Jiefang Bridge. GPS coordinates: 29°18'N, 95°18'E; altitude: 1000 m a.s.l. On dead Miscanthus, 24 October 2023, Dai 26693 (BJFC 044243).
Picipes
sinodictyopus
Taxon classificationFungiPolyporalesPolyporaceae
K.Y. Luo, Y.C. Dai & G.Y. Zeng sp. nov.
A0FFC84A-0FBD-528D-A4AC-A50ED60A91FE
861753
Holotype.
China • Yunnan Province, Wuding County, Shizishan Nature Reserve. GPS coordinates: 25°33'N, 102°21'E; altitude: 2000 m a.s.l. On dead branch of Litocarpus, 15 August 2019, Dai 20371 (BJFC 032039).
Basidiomata of Picipes sinodictyopus (Holotype, Dai 20371). Scale bar: 1 cm.
Diagnosis.
Picipes sinodictyopus is identified by coriaceous and without odour or taste basidiomata, cream to buff when fresh, olivaceous-buff when dry pore surface, angular pores, long stipe; infrequent generative hyphae in context, tubes and stipe and cylindrical, thin-walled basidiospores with dimensions of 5.8–7.3 × 2.3–2.8 µm.
Microscopic structures of Picipes sinodictyopus (Holotype, Dai 20371). a. Basidiospores; b. Basidia and basidioles; c. Hyphae from trama; d. Hyphae from context; e. Hyphae from stipe.
Etymology.
Sinodictyopus (Lat.) refers to the species similar to Picipes dictyopus, but having a distribution in China.
Description.
Basidiomata annual, laterally stipitate, solitary, coriaceous and without odour or taste when fresh, hard corky when dry. Pilei auricular to fan-shaped, about 2–2.5 cm long, 1.5–2 cm wide and up to 2 mm thick at base. Pileal surface reddish-brown to dark brown when fresh, becoming brown to reddish-brown when dry, glabrous; margin straight. Pore surface cream to buff when fresh, olivaceous-buff when dry; pores angular, 4–6 per mm; dissepiments thin, entire. Context light ivory to cream when dry, woody hard upon drying, up to 1 mm thick. Tubes concolorous with pore surface, up to 1 mm long. Stipe long, bearing a black cuticle, up to 1.5 cm long and 0.6 cm in diam.
Hyphal structure: Hyphal system dimitic; generative hyphae bearing clamp connections; skeletal-binding hyphae IKI–, CB+; tissues unchanged in KOH.
Context: Generative hyphae infrequent, colourless, thin-walled, rarely branched, 1–2 μm in diam.; skeletal-binding hyphae dominant, colourless, thick-walled, rarely branched, interwoven, 2.5–4 μm in diam.
Tubes: Generative hyphae infrequent, colourless, thin-walled, occasionally branched, 1.5–3 μm in diam; skeletal-binding hyphae colourless, thick-walled, occasionally branched, interwoven, 3–4.5 μm in diam. Cystidia and cystidioles absent. Basidia subclavate, with a basal clamp connection and four sterigmata, 12–22 × 4–8 μm; basidioles in shape similar to basidia, but slightly smaller.
Stipe: Generative hyphae infrequent, colourless, thin-walled, rarely branched, 1–2 μm in diam.; skeletal-binding hyphae dominant, colourless, thick-walled, rarely branched, interwoven, 3–4 μm in diam.
Basidiospores: Cylindrical, hyaline, thin-walled, smooth, some with one or two medium guttules, IKI–, CB–, (5.2–)5.8–7.3(–7.4) × (2.1–)2.3–2.8(–3.2) µm, L = 6.47 µm, W = 2.52 µm, Q = 2.57 (n = 30/1) (the specimen Dai 28935 is sterile).
Type of rot.
White rot.
Additional specimen (paratype) examined.
China • Guangxi Autonomous Region, Jinxiu County, Liugang, Daling Village. GPS coordinates: 23°59'N, 110°05'E; altitude: 800 m a.s.l. On rotten angiosperm wood, 15 July 2024, Dai 28935 (BJFC049194).
Discussion
Phylogenetically, three new species are nested in Picipes clade, based on ITS + nLSU topology (Fig. 1). Amongst them, P. abieticola grouped together with P. subtubaeformis and P. tubaeformis; however, P. subtubaeformis is delimited from P. abieticola by having shorter basidiospores (5.7–6.8 × 2.7–3.1 μm vs. 7.5–9.3 × 2.8–3.5 µm, Zhou et al. (2016); P. tubaeformis differs from P. abieticola by having circular to infundibuliform pilei and shorter basidiospores (6–7.8 × 2.3–3.2 μm vs. 7.5–9.3 × 2.8–3.5 µm, Dai et al. (1996). Picipes brevisporus grouped together with P. nigromarginatus; however, P. nigromarginatus is different from P. brevisporus by having larger pilei (2.2 × 3.4 cm vs. 1.5 × 0.6 cm), larger stipe (5 × 4 mm vs. 2 × 2 mm) and longer basidiospores (5.6–6.9 × 2.3–3 μm vs. 3.8–5.7 × 2.3–3.1 µm, Ji et al. (2022). Picipes sinodictyopus was sister to P. subdictyopus, but P. subdictyopus differs from P. sinodictyopus by its smaller pores (8–10 per mm vs. 4–6 per mm Tibpromma et al. (2017).
Morphologically, Picipes abieticola resembles P. dictyopus (Mont.) B.K. Cui, X. Ji & J.L. Zhou and P. subtropicus J.L. Zhou & B.K. Cui by sharing similar pores (5–7 per mm). However, P. dictyopus is different from P. abieticola by its longer tubes (3 mm vs. 1.5 mm) and longer stipe (5 cm vs. 1.5 cm) and growing on angiosperm wood in tropical and subtropical areas (Ryvarden and Johansen 1980); P. subtropicus is distinguished from P. abieticola by its smaller stipe (5 × 5 mm vs. 15 × 8 mm) and smaller basidiospores (5.1–6.2 × 2.2–2.7 μm vs. 7.5–9.3 × 2.8–3.5 µm, Zhou et al. (2016).
Picipes brevisporus resembles P. badius, P. baishanzuensis J.L. Zhou & B.K. Cui and P. conifericola (H.J. Xue & L.W. Zhou) J.L. Zhou & B.K. Cui by sharing subulate cystidioles. However, P. badius is different from P. brevisporus by larger basidiospores (5.5–8 × 3–3.8 μm vs. 3.8–5.7 × 2.3–3.1 µm) and lacking clamp connections (Cui et al. 2019); P. baishanzuensis is distinguished from P. brevisporus by having larger pores (3–6 per mm vs. 7–9 per mm) and longer basidiospores (6.6–7.9 × 2.5–3.1 μm vs. 3.8–5.7 × 2.3–3.1 µm) and growing on angiosperm wood (Cui et al. 2019); P. conifericola is different from P. brevisporus by its larger basidiomata (7 cm diam. vs. 1–1.5 cm long, 0.4–0.6 cm wide) and longer basidiospore, and growing on conifers (6.5–8.3 × 2.9–3.5 μm vs. 3.8–5.7 × 2.3–3.1 µm, Cui et al. (2019).
Picipes sinodictyopus is similar to P. annularius B.K. Cui et al., P. pseudovarius J.L. Zhou & B.K. Cui and P. taibaiensis (Y.C. Dai) J.L. Zhou & B.K. Cui by lacking cystidia and cystidioles. However, P. annularius is distinguished from P. sinodictyopus by having buff pileal surface and smaller stipe (4 mm × 3.5 mm vs. 15 mm × 6 mm, Ji et al. (2022); P. pseudovarius is different from P. sinodictyopus by its shorter stipe (5 mm vs. 15 mm) and longer basidiospores (7.7–9.3 × 2.6–3.4 μm vs. 5.8–7.3 × 2.3–2.8 µm, Cui et al. (2019); P. taibaiensis is distinguished from P. sinodictyopus by having shorter stipe (5 × 4 mm vs. 15 × 6 mm) and larger basidiospores (7.6–10.8 × 3.3–4.1 μm vs. 5.8–7.3 × 2.3–2.8 µm, Cui et al. (2019).
This paper enriches our knowledge of fungal diversity in the genus Picipes and it is likely that more new taxa will be found with further fieldwork and molecular analyses. There is still a lot of room to explore the diversity of Picipes species in China.
Key to species of Picipes in China
**: **
Supplementary Material
XML Treatment for Picipes abieticola
XML Treatment for Picipes brevisporus
XML Treatment for Picipes sinodictyopus
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bhunjun CS, Niskanen T, Suwannarach N, Wannathes N, Chen YJ, Mc Kenzie EHC, Maharachchikumbura SSN, Buyck B, Zhao CL, Fan YG, Zhang JY, Dissanayake AJ, Marasinghe DS, Jayawardena RS, Kumla J, Padamsee M, Chen YY, Liimatainen K, Ammirati JF, Phukhamsakda C, Liu JK, Phonrob W, Randrianjohany E, Hongsanan S, Cheewangkoon R, Bundhun D, Khuna S, Yu WJ, Deng LS, Lu YZ, Hyde KD, Lumyong S (2022) The numbers of fungi: are the most speciose genera truly diverse? Fungal Diversity 114: 387–462. 10.1007/s 132 · doi ↗
- 2Capella-Gutierrez S, Silla-Martinez JM, Gabaldon T (2009) Bioinformatics 25: 1972–1973.10.1093/bioinformatics/btp 348PMC 271234419505945 · doi ↗ · pubmed ↗
- 3Cui BK, Li HJ, Ji X, Zhou JL, Song J, Si J, Yang ZL, Dai YC (2019) Species diversity, taxonomy and phylogeny of Polyporaceae (Basidiomycota) in China. Fungal Diversity 97: 137–392. 10.1007/s 13225-019-00427-4 · doi ↗
- 4Dai YC (1996) Changbai wood-rotting fungi 5. Study on Polyporus mongolicus and P. tubaeformis. Annales Botanici Fennici 33(2): 153–163.
- 5Dong JH, Chen ML, Chen M, Li Q, Zhu YJ, Zhang XC, Zhou CQ, Li W, Muhammad A, Zhou HM, Jabeen S, Zhao CL (2025) Notes, outline, taxonomy and phylogeny of wood-inhabiting Agaricales. Mycosphere 16(1): 2599–2711. 10.5943/mycosphere/16/1/16 · doi ↗
- 6Dong JH, Li Q, Yuan Q, Luo YX, Zhang XC, Dai YF, Zhou Q, Liu XF, Deng YL, Zhou HM, Muhammad A, Zhao CL (2024) Species diversity, taxonomy, molecular systematics and divergence time of wood-inhabiting fungi in Yunnan-Guizhou Plateau, Asia. Mycosphere 15(1): 1110–1293. 10.5943/mycosphere/15/1/10 · doi ↗
- 7Han ML, Vlasák J, Cui BK (2015) Daedalea americana sp. nov. (Polyporales, Basidiomycota) evidenced by morphological characters and phylogenetic analysis. Phytotaxa 204: 277–286. 10.11646/phytotaxa.204.4.4 · doi ↗
- 8Hibbett D, Bauer R, Binder M, Giachini AJ, Hosaka K, Justo A, Larsson E, Larsson KH, Lawrey JD, Miettinen O, Nagy LG, Nilsson RH, Weiss M, Thorn RG (2014) Agaricomycetes. In: Mc Laughlin, D.J. & Spatafora, J.W. (Eds) The Mycota. Part A. Systematics and Evolution 2nd edn, vol. VII. Springer Verlag, 373–429. 10.1007/978-3-642-55318-9_14 · doi ↗