Dynamic modeling in neurocognitive frameworks of childhood ADHD: a review of inhibitory control and reward systems
Weidong Cai, Yoshifumi Mizuno

TL;DR
This review explores how ADHD in children involves problems with impulse control and reward processing, using new dynamic models to better understand brain mechanisms.
Contribution
The paper introduces dynamic modeling approaches to traditional ADHD neurocognitive frameworks, revealing novel neurodynamic mechanisms.
Findings
Children with ADHD show impairments in inhibitory control and reward processing.
Dynamic models reveal altered brain states and adaptive response strategies in ADHD.
Pharmacological studies show how medication modulates inhibitory and reward systems.
Abstract
Cognitive models of attention deficit hyperactivity disorder (ADHD) have traditionally centered on deficits in inhibitory control and motivation. A substantial body of empirical research supports impairments in both domains: children with ADHD exhibit elevated commission error rates, prolonged stop-signal reaction time, and a strong preference for smaller immediate rewards over larger delayed ones. In the first part of this review, we synthesize key findings from behavioral and task-state functional neuroimaging studies that characterize these deficits, highlighting how deficits in inhibitory control and motivation/reward systems manifest across tasks and contexts. We further examine evidence on the interplay between the two systems and how their interaction is altered in children with ADHD. In the second part, we introduce recent advances in cognitive and computational neuroscience…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
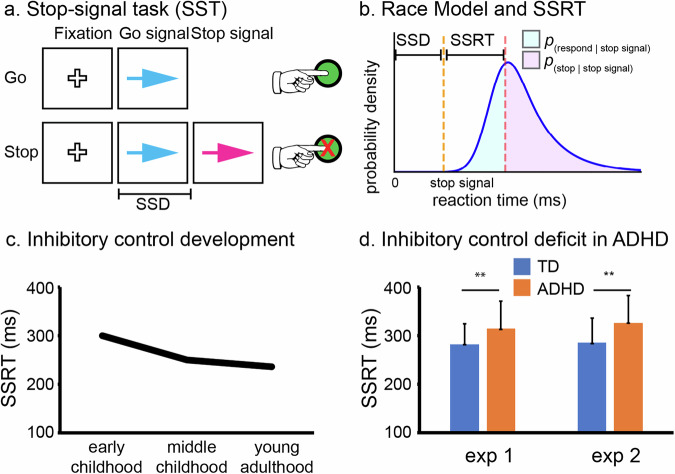 Figure 1
Figure 1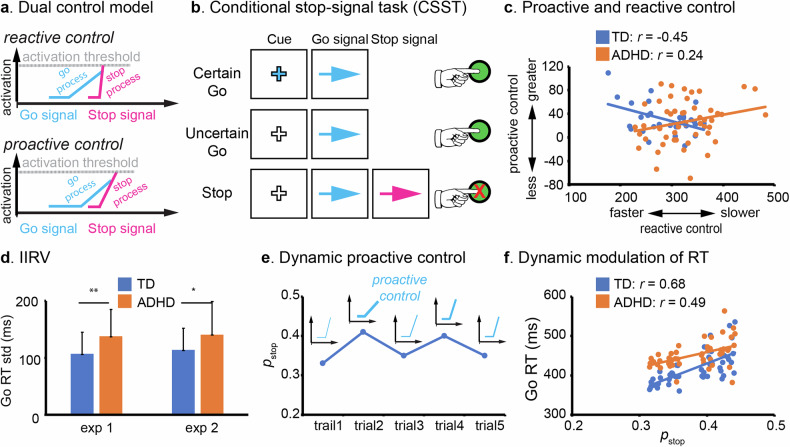 Figure 2
Figure 2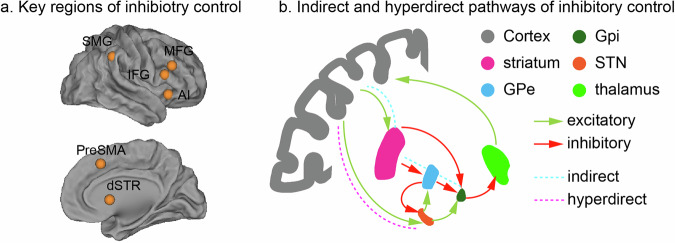 Figure 3
Figure 3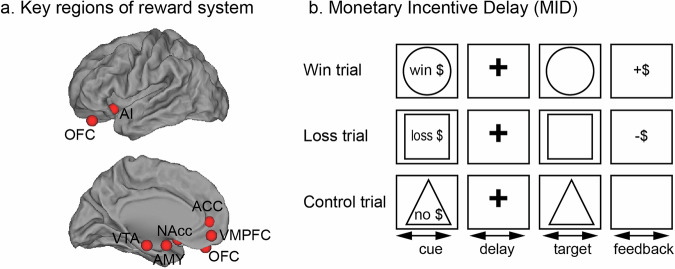 Figure 4
Figure 4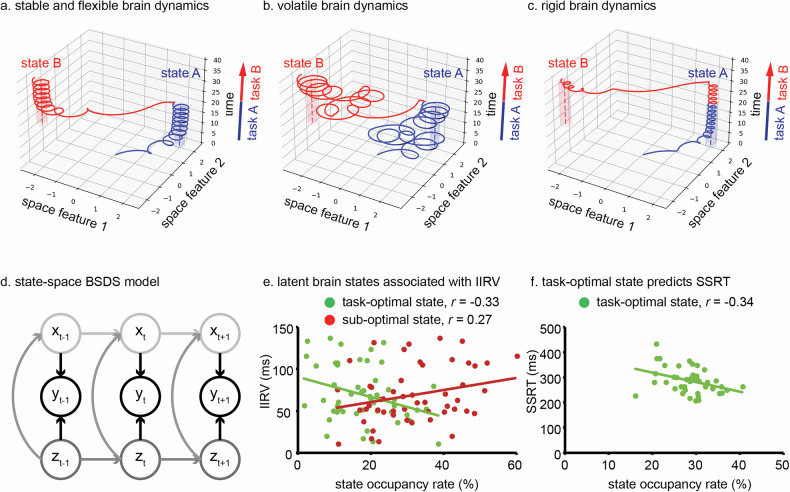 Figure 5
Figure 5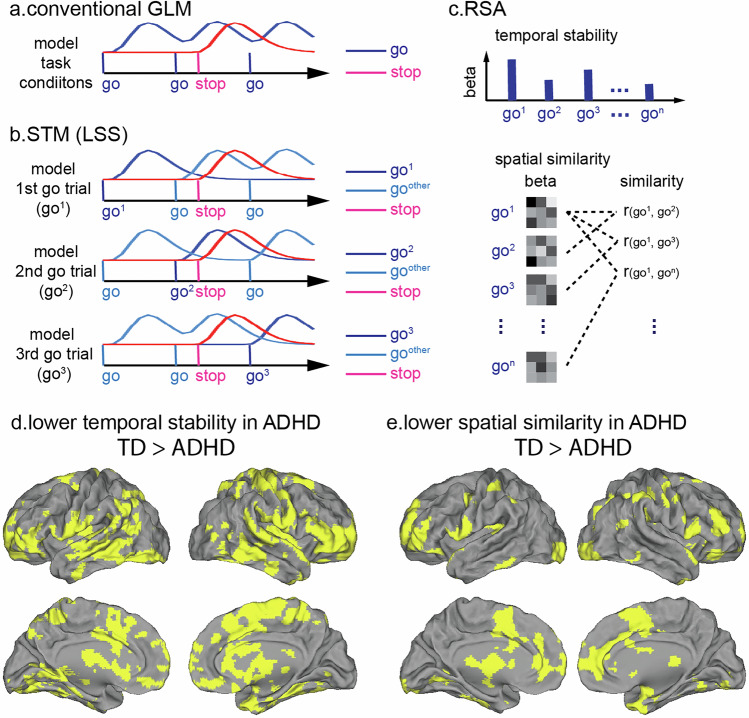 Figure 6
Figure 6- —https://doi.org/10.13039/100000025U.S. Department of Health & Human Services | NIH | National Institute of Mental Health (NIMH)
- —U.S.-Japan Brain Research Cooperative Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAttention Deficit Hyperactivity Disorder · Neural and Behavioral Psychology Studies · Functional Brain Connectivity Studies
Attention deficit hyperactivity disorder (ADHD) is a neurodevelopmental disorder with a significant global prevalence, estimated between 5–10% in the pediatric population [1–5]. In the United States, the diagnosis of childhood ADHD has risen markedly over the past three decades [6, 7]. According to the 2022 National Survey of Children’s Health, 11.4% of U.S. children aged 3–17 years had ever received an ADHD diagnosis, with 10.5% meeting criteria for current diagnosis based on parents’ report that the child has been diagnosed [8]. This substantial increase has spurred concerns and ongoing debate regarding the underlying pathophysiology of ADHD and the etiological factors contributing to these diagnostic trends [9].
Early diagnosis of childhood ADHD most commonly occurs between ages 5 and 7, when persistent and developmentally inappropriate behaviors, such as difficulty in remaining seated, frequent task interruptions, and consistent failure to follow instructions, become increasingly apparent and disruptive in structured settings like school. These behaviors are not isolated incidents but rather frequent and pronounced manifestations of the disorder’s core symptoms: inattention and hyperactivity/impulsivity. They often lead to academic underachievement, challenges in peer relationships, and increased family stress, which commonly prompt caregivers or educators to seek professional evaluation. Early identification is therefore critical, as it enables timely interventions that can improve long-term functional outcomes [10, 11].
Theoretical models of ADHD have long proposed that behavioral symptoms arise from underlying deficits in inhibitory control and motivation systems [12–21], both of which are essential for goal-directed behaviors [22, 23]. Empirical research has shown that children with ADHD tend to perform worse than typically developing (TD) peers on tasks involving inhibitory control and reward-related decision making [24, 25]. Findings from functional neuroimaging studies further reveal atypical activity in brain networks related to these cognitive functions during both rest and task engagement [26, 27]. However, despite these promising results, weak and inconsistent effect sizes have been reported [28, 29], casting doubt on the explanatory power of these neurocognitive models, thus limiting their clinical utility. From a clinical standpoint, these neurocognitive models have yet to yield reliable biomarkers that can be used for diagnostic, prognostic, or treatment monitoring purposes.
Human cognition relies on the brain’s capacity to flexibly and effectively allocate cognitive resources in response to moment-to-moment changes in task demands [30–36]. Understanding the underlying brain dynamics, encompassing transient and time-varying interactions among distributed brain areas, during task performance is thus essential for developing a comprehensive neurocognitive framework to investigate cognitive deficits and behavioral dysfunctions. Early neuroscience models of ADHD posited that unstable brain states may underlie attentional difficulties in affected children [37]. However, most of the existing research has focused on average neural responses, overlooking critical features of functional brain dynamics that contribute to clinical symptoms of ADHD. Integrating dynamic perspectives into clinical research offers the potential to uncover previously unrecognized neurocognitive mechanisms that drive behavioral problems associated with the disorder.
In this review, we examine behavioral and neuroimaging evidence supporting inhibitory control and motivation/reward models of ADHD. Our focus is specifically on task-state fMRI findings, as they capture brain responses to cognitive demands. While resting-state fMRI has generated a large body of research, it lacks the ability to probe how the brain dynamically responds to goal-directed tasks. We also highlight emerging research that integrates neurodynamic frameworks into these models, aiming to provide a more comprehensive and mechanistic understanding of the disorder’s neurobiological underpinnings.
Inhibitory control and childhood ADHD
Inhibitory control is a fundamental function that regulates an individual’s actions, emotions, and thoughts. This ability develops progressively throughout childhood and adolescence, reaching maturity in early adulthood [38, 39]. Notably, middle childhood (8–10 years old) represents a particularly critical period for the development of inhibitory control [40] and coincides with the typical diagnosis of childhood ADHD [41]. According to Barkley’s inhibitory control model of ADHD, many behavioral issues observed in children with the disorder stem from difficulties in suppressing urges and impulses that are inappropriate or no longer relevant for the context [12].
Inhibitory control is hypothesized to be a domain-general, cross-modality process, with similar mechanisms underlying inhibition of both motor and cognitive processes [42–45]. Because of the practical advantage in validation and measurement, motor response inhibition is most commonly studied in an experimental setting. Various task paradigms, such as Go/NoGo task (GNGT), Stop-signal task (SST), Flanker task, Stroop task, and anti-saccade task have been developed to examine inhibitory control over actions. These tasks typically include two trial types: low-demand and high-demand of inhibitory control. The high-demand trials require participants to cancel or override prepotent, reflexive, or automatic responses that are inappropriate in a given context, such as in no-go, stop, incongruent, and anti-saccade conditions (Fig. 1a). Among these paradigms, the SST is unique as it allows estimation of stop-signal reaction time (SSRT) via the Race Model, providing a quantitative measure of how quickly an individual can inhibit dominant response tendency [46, 47] (Fig. 1b). Notably, neurophysiological evidence from non-human primate studies has demonstrated strong alignment between SSRTs and the timing of neuronal activity associated with action cancellation, supporting the neurobiological validity of SSRT [48, 49].Fig. 1. Inhibitory control paradigm, model, development and deficits in ADHD.a Stop-signal task (SST) involves frequent Go trials and infrequent Stop trials. Participants are instructed to make accurate and speedy responses (e.g. button press) to go signals (e.g. a blue arrow) and withhold their response tendency when a go signal is shortly followed by a stop signal (e.g. the blue arrow turns to red). The critical parameter in the SST is the stop-signal delay (SSD), the interval between the onsets of go and stop signals, whose values are typically adjusted trial-by-trial with a performance-based staircase procedure. b The Race model can estimate stop-signal reaction time (SSRT), which quantifies how fast one can cancel a prepotent response [46]. c SSRTs become shorter through childhood, indicating development of inhibitory control functions. The plot illustrates averaged SSRTs from early and middle childhood to young adulthood. Data were obtained from the two previous studies [38, 39]. d Children with ADHD have elongated SSRTs in comparison to TD children, suggesting inhibitory control deficits in ADHD. The plot is adapted from a previous study in which two experiments (exp1 and exp2) show similar inhibitory control deficits in children with ADHD (9–12 years old) [50].
The SSRT improves steadily from childhood through adolescence, typically reaching a plateau in early adulthood [38, 39] (Fig. 1c). Numerous studies have shown that children with ADHD exhibit longer SSRTs compared to their typically developing (TD) peers (Fig. 1d) and slower SSRTs are associated with more severe ADHD symptoms [50–52]. Meta-analytic research confirms that SSRT yields a median effect size (~0.6) in distinguishing children with ADHD versus TD children, surpassing the effect size of commission errors on the GNG task (~0.5), another widely used behavioral index of inhibitory control [25]. These findings suggest that children with ADHD have relatively weaker inhibitory control over actions, aligning with the inhibitory control model of the disorder [12].
Dynamic dual control model and ADHD
Despite the long-standing view that inhibitory control is a single cognitive process, more recent dual control models propose that inhibitory control operates through two distinct modes: reactive and proactive control [53, 54] (Fig. 2a). Reactive inhibitory control refers to the suppression of a prepotent motor, cognitive, or emotion response upon detecting conflict or interference. In a classroom setting, this is exemplified by a child who impulsively starts to shout out an answer but manages to stop immediately upon realizing they did not raise their hand. In SST, reactive control is typically quantified using the SSRT. Calculated via the Race Model, the SSRT provides an estimate of the latency of the unobservable inhibitory process, representing the speed at which an individual can reactively halt a response once a stop signal is presented.Fig. 2. Dynamic dual inhibitory control model.a The dual inhibitory control model distinguishes between reactive and proactive control processes. Reactive control involves the suppression of a dominant motor, cognitive, or emotional response triggered by the detection of conflict or interference, whereas proactive control involves the anticipatory deployment of inhibitory mechanisms based on prior knowledge or contextual cues that signal a high likelihood of upcoming conflict. In the stop-signal task, reactive control corresponds to the stop process initiated by the infrequent stop signal, aiming to reach its activation threshold before the go process completes; whereas proactive control is reflected in strategically slowing of the go process, delaying its progress toward the activation threshold in anticipation of a potential stop signal. b The conditional stop-signal task (CSST) is an experimental paradigm designed to probe dual mechanisms of inhibitory control. In this task, the Certain Go cue (blue cross) signals a go trial with no probability of a stop signal, thus involving no inhibitory control demand. In contrast, the Uncertain Go cue (white cross) introduces the possibility of a stop signal appearing shortly after the go signal, thereby engaging proactive inhibitory control mechanisms due to increased demand for response inhibition. Proactive control is typically measured by increased reaction time (RT) in the Uncertain Go than Certain Go conditions. c A previous study has revealed that greater proactive control is associated with faster reactive control functions in TD children (r = −0.45) but not in children with ADHD [50]. Reactive control is quantified using the SSRT. Proactive control is calculated as the difference in RT between Uncertain Go and Certain Go trials. d Children with ADHD have higher intra-individual response variability (IIRV) than TD children. The plot is adapted from a previous study in which two experiments (exp1 and exp2) show heightened IIRV in children with ADHD (9–12 years old) [50]. e Based on trial history, participants continuously update their belief about the probability of stop signal (p_stop_) in the coming trial and adjust response strategies (proactive control) accordingly. f A previous study has shown that trial-wise anticipation of stop signal (p_stop_) is strongly associated with RT in TD children and such modulation is weaker in children with ADHD [50]. Two exemplar participants’ data are illustrated (1 TD and 1 ADHD).
In contrast, proactive inhibitory control involves the anticipatory deployment of inhibitory mechanisms based on prior knowledge or contextual cues that signal the likelihood of upcoming conflict. For instance, a student acts proactively when they consciously slow down their movements and quiet their voice before entering a library, anticipating the requirement for silence. Although both processes contribute to performance on inhibitory control tasks, most behavioral measures, such as commission errors, stop accuracy, and SSRT, are primarily designed to assess reactive inhibitory control. Proactive inhibitory control, on the other hand, is examined by manipulating task demand, such as varying the probability of inhibitory signals (e.g. stop or no-go signals) (Fig. 2b). Operationally, proactive control is calculated as the magnitude of response slowing, specifically, the strategic prolongation of reaction time on Go trials in contexts with a high probability of stop signals compared to low probability contexts. A great degree of response slowing indicates a robust engagement of proactive strategies to prevent commission errors.
Support of the dual control model comes from multiple lines of evidence: (1) correlation between SSRT and response slowing [55], (2) similar suppression effects on motor excitability in both control modes [56, 57], and (3) overlapping neural substrates underlying reactive and proactive inhibitory control processes [58]. Like reactive inhibitory control, proactive inhibitory control is under development during childhood [59–62].
Although numerous studies have examined reactive inhibitory control in childhood ADHD, research on proactive inhibitory control remains limited and findings are mixed [50, 63–65]. For example, Van Hulst et al. used a modified SST incorporating anticipatory cues to examine reactive and proactive inhibitory control in TD children and those with ADHD symptoms and other neurodevelopmental disorder symptoms [65]. The cues indicated varying probability of stop signals, and proactive inhibitory control was assessed via the interaction between stop signal probability and reaction time on go trials. Although they found a robust proactive control effect, no significant between-group differences were observed. In another study that manipulated stop signal probabilities within the classic SST framework, researchers reported that children with ADHD exhibited similar levels of response slowing as TD children when provided with explicit task cues [50]. However, when proactive control strategies had to be learned implicitly through experience, by tracking trial history and action consequences, children with ADHD were less efficient in utilizing such strategies [50]. Interestingly, TD children with shorter SSRTs also showed greater response slowing, suggesting a coordinated interaction between proactive and reactive inhibitory control mechanisms, similar to patterns seen in adults [50, 55]. This coordinated engagement was absent in children with ADHD [50] (Fig. 2c), implying not only deficits in reactive control, but also challenges in integrating and coordinating both inhibitory control modes even though proactive inhibitory control can be implemented under explicit instruction.
Inhibitory control tasks often engage implicit learning processes, as local probability of task-relevant signals (e.g., no-go or stop signals) varies from trial to trial, even though the overall task structure remains constant. As a result, dynamically updating expectation based on recent trial history is critical for accurate anticipation, strategic adjustment, and optimal behavioral performance. Traditional behavioral metrics, such as mean accuracy and reaction time (RT), are static measures that fail to capture trial-by-trial variability in cognitive processing. Measures of intra-individual response variability (IIRV), such as the standard deviation, sigma, or tau of RT [66, 67], have been used to quantify behavioral fluctuation during task performance. IIRV is often used as an index of attentional lapses, and studies consistently show that children with ADHD exhibit greater IIRV than TD children, reflecting less consistent task engagement and potentially poorer sustained attention [25, 68–71] (Fig. 2d). However, IIRV has notable limitations. First, it is a coarse metric that aggregates data across trials and lacks the temporal resolution to capture dynamic processes as they evolve over time. Second, variability in RT can stem from multiple sources, such as arousal regulation, fatigue, attentional lapses, or strategic adjustment [71–73], but IIRV cannot disentangle these contributing factors.
To better understand the dynamic cognitive mechanisms underlying inhibitory control, computational modeling approaches have been employed to estimate trial-by-trial latent processes driving behavioral variability [74, 75]. One such model is the Dynamic Belief Model (DBM), which estimates a subject’s belief about the chance of an upcoming inhibitory signal (p_stop_) based on trial history [75]. According to this model, individuals are expected to exert greater proactive control when they believe the probability of an inhibitory signal is increasing (Fig. 2e). This prediction has been supported by strong positive correlations between trial-wise p_stop_ and RT in adult participants [75–77], indicating strategic modulation of behavior in anticipation of inhibitory demands. A similar association has been observed in children, suggesting that they are capable of making strategic, history-based adjustment in proactive control. However, children with ADHD exhibit weaker correlation between trial-wise p_stop_ and RT compared to their TD peers [50], indicating a reduced capacity to update belief based on trial history and/or adjust proactive control strategies accordingly (Fig. 2f).
The dynamic dual control model distinguishes between proactive and reactive inhibitory control mechanisms, characterizing their continuous adaptation to changing task demands. Integrating this framework into models of inhibitory control offers a more comprehensive understanding of cognitive deficits in children with ADHD by capturing the temporal variability, multifaceted cognitive processes, and strategic adjustment that underlie inhibitory control functions.
Neural mechanism of inhibitory control and ADHD
Inhibitory control engages a widely distributed network of cortical and subcortical regions. Meta-analytic research has identified consistent activation in key areas, including the anterior insula, inferior frontal gyrus, middle frontal gyrus, pre-supplementary motor cortex, supramarginal gyrus and dorsal striatum, with a dominant involvement of the right hemisphere during response inhibition [40, 78, 79] (Fig. 3a). In particular, the indirect and hyper-direct pathways of basal ganglia have been implicated in inhibitory control of motor responses [80, 81] (Fig. 3b). It has been suggested that the subthalamic nucleus, a critical node of the hyper-direct pathway, plays a vital role in rapid inhibition [82]. Although the subthalamic nucleus is not frequently reported in task-state neuroimaging studies of inhibitory control, its importance has been substantiated by both animal studies and human clinical research [40, 83–86]. Task-dependent connectivity analyses further demonstrate that inhibitory control modulates interactions both within canonical cortical networks and between cortical and subcortical regions [40, 84, 87–91], highlighting the system-wide communication necessary for effective inhibitory control. However, few studies have examined task-related activation and connectivity of the subthalamic nucleus in children with ADHD.Fig. 3. Cortical-subcortical regions involved in inhibitory control.a Inhibitory control tasks, such as stop-signal task and go/no-go task, commonly recruit the anterior insula (AI), inferior frontal gyrus (IFG), middle frontal gyrus (MFG), pre-supplementary motor area (preSMA), supramarginal gyrus (SMG) and dorsal striatum (dSTR), with activation dominance in the right hemisphere. The coordinates of these plotted regions were obtained from meta-analysis of 70 functional neuroimaging studies in which the stop-signal task and go/no-go task were used to examine brain activation during inhibitory control in adult participants [78]. b Indirect and hyper-direct pathways of cortical-basal ganglia circuits have been implicated in inhibitory control of actions.
Inhibitory control paradigms, such as SST and GNGT, have been widely used to probe abnormal brain activation in ADHD. Numerous studies have reported hypoactivation in core inhibitory control regions, including inferior frontal gyrus and dorsomedial prefrontal cortex, in children and adolescents with ADHD compared to TD peers [51, 92–106]. However, findings are mixed, with some studies reporting no significant between-group differences [107–109]. Notably, many earlier neuroimaging studies had small sample sizes (n < 20 per group), limiting their generalizability. A larger study involving 74 adolescents with ADHD and 74 controls found hypo-activation in lateral and medial prefrontal cortex and striatum during a Go/No-Go task [102]. When making commission errors (i.e. failed inhibition), adolescents with ADHD also showed reduced activation in the anterior insula and anterior cingulate cortex, core nodes of the salience/error detection system, and failed to downregulate default mode network, unlike the controls. These results, replicated in other studies [107, 110], suggest that children with ADHD are less likely to engage salience/error detection system in response to errors. Meta-analytic research further confirmed consistent hypoactivation of the salience and frontoparietal networks across cognitive tasks requiring motor inhibition in individuals with ADHD [26, 111], linking the disorder to impaired recruitment of inhibitory control networks.
Fewer studies have investigated how inhibitory control modulates functional connectivity in children with ADHD [107, 112]. Using generalized psychophysiological interaction analysis [113], Cai et al. examined task-dependent connectivity during a Go/NoGo task in 46 children and adolescents with ADHD and 51 TD children and adolescents acquired in the International Study to Predict Optimized Treatment in ADHD trial [107]. They found that connectivity between dorsolateral prefrontal cortex and posterior parietal cortex, two key nodes in the frontoparietal network, was significantly weaker in children with ADHD than in TD children during No-Go trials. Similarly, van Rooji et al. reported reduced connectivity between inferior frontal gyrus and basal ganglia, as well as between superior frontal gyrus and thalamus and operculum, during successful stop trials in children with ADHD [112]. These findings highlight altered cortical-cortical and cortical-subcortical connectivity patterns in response to inhibitory control demand in children with ADHD, although further research is needed to confirm their reproducibility.
Overall, early task-state functional neuroimaging studies generally agree that children with ADHD show deficits in upregulating inhibitory control system and/or downregulating default mode system in response to increased demand of inhibitory control.
Motivation, reward-related processes and ADHD
Despite its popularity, the inhibitory control theory has faced considerable scrutiny within the field. First, deficits in inhibitory control are not unique to ADHD; they are also observed in other neurodevelopmental disorders, such as autism [114] and learning disorder [115]. Second, several studies have reported comparable performance on inhibitory control tasks between individuals with and without ADHD, raising questions about the universality of such deficits among those diagnosed. Third, some behavioral difficulties commonly attributed to ADHD may be better explained by impairments in other cognitive domains. As a result, the inhibitory control hypothesis, at least when considered in isolation, does not fully account for the diverse range of behavioral problems observed in children with ADHD.
An alternative and influential theory suggests that many behavioral challenges in ADHD may stem from reduced motivation, especially in tasks requiring sustained cognitive or physical efforts [116–118]. Some researchers have proposed that impairments in inhibitory control may, in fact, be secondary to more fundamental deficits in motivation [17, 119, 120]. This view is supported by a growing body of evidence. Studies using self- or teacher-reported questionnaires and observation methods have found that children with ADHD tend to be less motivated to engage in academic tasks compared to their TD peers [121–123], although findings are somewhat mixed [124]. For example, one recent study found that children and adolescents with ADHD reported particularly low motivation for cognitively demanding tasks [125]. Another study involving word puzzles found that children with ADHD were more likely to abandon the task than TD children, indicating reduced persistence and motivation in academic context [126]. Furthermore, the severity of core ADHD symptoms, such as inattention and hyperactivity, has been linked to diminished academic motivation among kindergarten students [127].
Motivation to complete a task, however, is influenced by a range of factors, including perceived task difficulty relative to one’s abilities and the individual’s self-belief or confidence. It is plausible that children with ADHD may develop low self-efficacy due to repeated academic failure, which in turn reduces their motivation to engage in learning activities [128]. Although experimental research examining motivational processes in non-academic contexts remains limited, several studies offer valuable insights. For instance, one investigation found that adults with ADHD were less willing than controls to engage in repeated manual effort tasks [129]. In another experiment requiring participants to choose between varying levels of cognitive or physical effort, individuals with ADHD showed a general reluctance to invest effort in both types of tasks [130]. However, findings are not entirely consistent: one study reported that children with ADHD were just as likely as TD peers to select high-effort tasks, although their engagement was less consistent [131]. Another study found no significant group differences in physical effort-related discounting [132].
Importantly, both extrinsic and intrinsic motivation, central to human behavior, are strongly influenced by external factors, such as reward [133–135]. Motivation is closely tied to reward sensitivity, and the motivational deficit theory posits that children with ADHD may exhibit altered sensitivity to rewards [119], a feature broadly implicated across various mental health conditions [136]. Numerous studies have examined whether children with ADHD respond differently to rewards compared to their TD peers. One of the most widely used paradigms in this area is the delay discounting task (DDT), which assesses impulsive decision-making and reward sensitivity. In the classical DDT, participants choose between smaller, immediate rewards and larger, delayed ones. Children with ADHD tend to prefer smaller, immediate rewards [137–139], indicating a steeper discounting of delayed gratification. Meta-analyses of DDT studies have reported moderate effect sizes (approximately 0.4–0.5) in distinguishing individuals with ADHD from controls [24, 140, 141].
Taken together, these behavioral findings suggest that children with ADHD exhibit reduced motivation and altered reward sensitivity, particularly characterized by a heightened preference for immediate rewards and difficulty with delayed gratification, relative to their TD peers.
Neural mechanism of reward-related processes and ADHD
A broad network of cortical and subcortical regions, including the ventromedial prefrontal cortex, orbital frontal cortex, anterior insula, anterior cingulate cortex, nucleus accumbens, amygdala, and ventral tegmental area, play central roles in motivational and reward-related processes [142–147] (Fig. 4a). One of the most widely used experimental paradigms to investigate these processes in neuroimaging research is the monetary incentive delay task (MID) [145] (Fig. 4b). In this task, participants are presented with cues that signal the potential to win money, avoid losing money, or receive no incentive, followed by a target that requires a rapid response. To maintain a pre-defined success rate, the task difficulty is adaptively adjusted, typically by changing the response time thresholds, yielding different reward outcomes such as win, no-win, loss, and no-loss. The delay period between cue and stimulus presentations is critical in functional neuroimaging studies because it enables the dissociation of brain signals associated with reward anticipation from those related to reward outcome. Meta-analyses of MID studies have consistently found that the nucleus accumbens, anterior insula, and amygdala are strongly activated during reward anticipation, whereas ventromedial prefrontal cortex and orbital frontal cortex are more engaged during reward outcome [148, 149]. In a large-scale adolescent neuroimaging dataset (n = 1510) [150], Cao and colleagues revealed widespread activation during reward anticipation including the medial prefrontal cortex, insula, dorsal and ventral striatum, thalamus, and ventral tegmental area. In contrast, activation during reward outcome was more limited, primarily involving the ventromedial prefrontal cortex. Moreover, using psychophysiological interaction methods, the study demonstrated that reward increases functional connectivity between ventral striatum and cortical regions such as the dorsomedial prefrontal cortex, anterior cingulate cortex, and anterior insula during reward anticipation, highlighting the extensive communication between reward and salience networks during reward processing.Fig. 4. Reward system and task paradigm.a Reward processing tasks commonly recruit the ventromedial prefrontal cortex (VMPFC), orbital frontal cortex (OFC), anterior insula (AI), anterior cingulate cortex (ACC), nucleus accumbens (NAcc), amygdala (AMY) and ventral tegmental area (VTA). Coordinates of these regions of interests are obtained from previous meta-analyses of reward-related task fMRI studies [146, 147]. b The monetary incentive delay (MID) task is widely used to examine neural substrates underlying reward anticipation and reward outcome [145]. The variable delays between cues and targets allow dissociation between reward anticipation and reward outcome processes.
The MID has also been extensively employed to examine ADHD-related functional abnormalities in the brain’s reward circuitry [102, 151–161]. Although behavioral differences between ADHD and controls are often minimal in the MID task, a consistent neuroimaging finding is reduced activation in ventral striatum during reward anticipation in individuals with ADHD [102, 151–161]. This hypoactivation is thought to reflect deficits in reward anticipation mechanisms. Additionally, reduced ventral striatum activation during reward anticipation has been linked to hyperactivity in both the MID [151, 157, 159] and DDT [162] tasks, although some studies have reported weak or even opposite effects [156, 161]. Results related to reward outcome have been even more variable. For example, Strohle et al. observed increased activation in right orbitofrontal cortex, left middle frontal gyrus, and inferior frontal gyrus in adults with ADHD compared to controls [159]; Plichta et al. reported greater striatal activation in males with ADHD than controls in response to success outcome [162]; and Stoy et al. found reduced activation in bilateral insula during outcome of loss avoidance [158]. Despite these findings, little is known about task-dependent connectivity during reward-related processes in ADHD.
Together, functional neuroimaging studies provide converging evidence that individuals with ADHD exhibit deficits in engaging key components of the brain’s reward circuit, particularly the ventral striatum, during reward-related processes. These findings underscore the importance of motivational and reward-related functional abnormality in the neurobiology of ADHD.
Interaction between cognitive control and reward systems
Crucially, inhibitory control and motivation are interrelated processes, rather than isolated functions [22, 163, 164]. The precise mechanism through which changes in motivational states and reward influence inhibitory control are not fully understood. Behavioral research has shown that reward significantly influences individuals’ performance on inhibitory control tasks [165, 166]. Generally, when reward contingencies emphasize response inhibition, individuals tend to adjust their response strategies, leading to greater response slowing (indicative of proactive control) and faster stopping speed (reflecting reactive control) [165, 166]. A substantial body of research has examined the interplay between motivation and inhibitory control in children with ADHD [157, 167–189]. Most of these studies employ classic inhibitory control paradigms such as GNG or SST, incorporating reward manipulation (e.g. varying reward magnitudes or contrasting reward versus loss conditions). Although significant reward effects and group differences in inhibitory control measures are commonly observed, findings regarding the interaction between reward manipulation and ADHD diagnosis are inconsistent [169–190]. The majority of studies report no significant interaction, but a few have found that reinforcement enhances inhibitory control function more in children with ADHD than TD or clinical control groups [171, 177, 182, 185, 187].
Using a reward-modulated inhibitory control paradigm, Boehler et al. found increased activation in the ventral striatum, anterior insula, and anterior cingulate cortex in reward-related than reward-unrelated stop trials, along with enhanced functional connectivity between anterior cingulate cortex and dorsomedial prefrontal cortex [191]. Other studies have similarly shown that reward modulation of inhibitory control engages not only key regions of the reward circuitry but also areas involved in inhibitory control [192–194], a finding supported by meta-analytic research [23]. Despite these insights, few studies have examined the neural mechanisms of reward-modulated inhibitory control in ADHD. Using a Go/NoGo task with low- and high-incentive conditions, Liddle et al. found attenuated deactivation of the default mode network in children with ADHD relative to TD children under low incentive conditions, a difference that disappeared under high incentive conditions [178].
Whereas there is a general agreement that reward enhances inhibitory control, even for children with ADHD, it remains unclear whether this enhancement is greater, less, or equivalent in magnitude compared to children without ADHD. To better understand this variability, it is important to investigate the underlying neural mechanisms of reward-modulated inhibitory control, particularly the dynamic aspects of these mechanisms, which may offer critical insights into individual differences in response to rewards among children with ADHD.
Neurodynamic models of cognition and ADHD
Cognition relies on the brain’s capacity to sustain both stable and flexible communication across large-scale networks, facilitating the timely engagement and maintenance of task-optimal brain states while resisting the intrusion of sub-optimal brain states (Fig. 5a) [30–36]. Disruptions in this dynamic balance, manifesting as excessive volatility (i.e., difficulty maintaining task-optimal brain states; Fig. 5b) or excessive rigidity (i.e., delayed disengagement from sub-optimal brain states; Fig. 5c), have been linked to cognitive instability and inflexibility [35, 68, 195]. In a seminal review [37], Sonuga-Barke and Castellanos proposed the default mode interference hypothesis, which posits that inadequate suppression of default mode network activity may allow task-irrelevant states to intrude during goal-directed cognitive processing. These intrusions can compete for limited cognitive resource, leading to attentional lapse and impaired performance in individuals with ADHD. This hypothesis provides a neurobiologically plausible account of the elevated IIRV, a behavioral hallmark in ADHD [69–71, 196], and is supported by neuroimaging evidence showing dysregulated default mode network activity during cognitive tasks and its association with IIRV [178, 197–199]. Consistent with the role of default mode network in moment-to-moment attentional fluctuation, Esterman et al. found that increased commission error rates were preceded by elevated default mode network activity, particularly when participants were in a stable performance state [72].Fig. 5. Schematic illustration of stable and flexible, volatile and rigid brain dynamics during cognitive performance.The optimal state for Task A is denoted as latent state A, and the optimal state for Task B is denoted as latent state B. a Stable and flexible dynamics: During task A, brain activity remains in latent state A. Upon switching to Task B, the brain transits efficiently from state A to state B, maintaining task-optimal dynamics. b Volatile dynamics: brain activity fails to stabilize in the task-optimal state, exhibiting frequent deviations from the optimal latent state for the current task to sub-optimal states. c Rigid dynamics: Following a switch from Task A to Task B, the brain remains persistently in latent state A, showing delayed disengagement and transition to the new task-optimal state B. d Schematic illustration of the state-space Bayesian Dynamic Switching System (BSDS) model: y_t_ denotes the observed fMRI timeseries at time point t; z_t_ denotes the latent states that are interdependent through a first order Markov chain; x_t_ denotes the latent space variables. Details of the model are described in a previous study [35]. e A previous study [68] has found that higher occupancy rates of the task-optimal brain state are associated with lower intra-individual response variability (IIRV) whereas higher occupancy rates of sub-optimal brain states are associated with higher IIRV in TD children and children with ADHD. f A previous study [31] has revealed that high occupancy rates of the task-optimal brain state are associated with better inhibitory control function (shorter SSRT) TD children and children with ADHD. Occupancy rate is calculated by dividing the number of time points assigned to a state by the total number of time points in the time series.
An open question is whether cognitive deficits in ADHD arise from difficulty in sustaining task-optimal states, increased susceptibility to sub-optimal state intrusion, or both, particularly within neural systems governing inhibitory control and motivation. The traditional general linear model (GLM), which estimates averaged condition-specific activation pattern (Fig. 6a), is incapable of capturing continuous fluctuation in neural activity during task performance, especially when such fluctuations are not tightly time-locked to experimental events. Moreover, model-driven analytic approaches, such as GLM, typically assume that similar behaviors are generated by similar underlying neural processes. However, this assumption often fails in practice. For instance, a slower response may result from attentional lapse, fatigue, or effortful proactive control, with distinct brain mechanisms producing superficially similar outcomes. As such, neurodynamic modeling approaches are essential for detecting ongoing changes in brain activity driven not only by experimental manipulation (e.g., cognitive demand) but also by non-experimental influences (e.g., practice effect, motivation, fatigue or attentional lapse).Fig. 6. Single trial model (STM) and representational similarity analysis (RSA).a The conventional general linear model (GLM) estimates condition-specific brain activation pattern, i.e. task-evoked activation across all the go trials. b STM estimates trial-specific brain activation pattern. For example, in the STM for the 1^st^ go trial, the Least-Squares Separate (LSS) technique separate the vector of the 1^st^ go trial from the vector of the rest of the go trials and the vectors of other condition. The same approach is applied for estimation of activation pattern elicited by each go trial. c RSA can examine temporal stability and spatial similarity of trial-wise brain activation. A recent study has used STM + RSA methods to demonstrate children with ADHD have (d) lower temporal stability and (e) lower spatial similarity of trial-wise activation pattern than TD children (p < 0.05, TFCE corrected) during the conditional stop-signal task [108].
Although widely applied in resting-state fMRI studies, the sliding window approach, which computes dynamic measures by moving a fixed-size window across the time series [200], is less suited for task-state fMRI research because of the difficulties in selecting an appropriate window size and the risk of crossing task structure boundaries. In contrast, state-space models have proven effective for uncovering latent brain dynamics that are not necessarily time-locked to task events [35, 201–204]. These models typically define states as unique patterns of brain activation and connectivity [205], and describe their temporal evolution in probabilistic frameworks such as hidden Markov models.
In state-space models, latent brain states are characterized by distinct spatial configurations of brain activation or functional connectivity. The temporal dynamics of these states are typically quantified using two key metrics: occupancy rate (or fractional occupancy) and mean lifetime (or dwell time). The occupancy rate represents the overall dominance of a state, defined as the proportion of the total task duration that an individual spends in that specific latent state. It is calculated by dividing the number of time points assigned to a state by the total number of time points in the time series. The mean lifetime reflects state stability, defined as the average continuous duration a state persists before transitioning to a different state.
Yamashita et al. used an energy landscape analysis to identify optimal and sub-optimal brain states during a sustained attention task [204]. Their findings showed that adults with ADHD spent more time in the sub-optimal state and less time in the optimal state compared to controls. However, the method relies on binarization of brain activation, which limits the interpretability of the resulting brain state features and may contribute to inconsistency in the findings. In another study, Cai et al. applied the state-space Bayesian Switch Dynamic System model (Fig. 5d) to identify latent brain states during a choice response task [68]. They found that more time spent in a task-optimal state is related to decreased IIRV whereas more time spent in sub-optimal states is associated with increased IIRV (Fig. 5e). Moreover, time spent in non-optimal state, the state characterized by weaker functional connectivity between regions in salience and frontoparietal networks relative to the task-optimal state, predicted inattention symptoms. Similarly, another study found that higher probabilities of engaging task-optimal brain state were linked to better inhibitory control function (i.e., faster SSRTs) and less severe inattention problem [31] (Fig. 5f). Together, these findings highlight aberrant brain dynamics in ADHD during cognitive performance and also demonstrate the utility of state-space modeling in revealing dynamic neural signatures of ADHD.
Despite the strengths of state-space modeling, aligning state-space-inferred dynamics with discrete task events can be challenging, particularly in fast event-related design. To better capture trial-wise neural dynamics, single-trial models (STMs) have emerged as a powerful alternative [206, 207] (Fig. 6b). When combined with representation similarity analysis (RSA), STMs enable the quantification of both activation magnitude and spatial pattern similarity across trials (Fig. 6c), which have been widely used to study memory, learning, pain and social function [208–211]. In a recent study, Gao et al. innovatively employed the STM-RSA framework to study dynamic dual control deficits in children with ADHD [108]. They developed two novel metrics to characterize temporal and spatial properties of brain dynamics: temporal and spatial stability. Temporal stability measures the consistency of activation strength across trials, whereas spatial stability quantifies the consistency of activation patterns among neighboring voxels across trials (Fig. 6c). They found that children with ADHD exhibit reduced temporal and spatial stability of task-evoked activation patterns during a conditional stop-signal task compared to TD children (Fig. 6d-e) [108]. Notably, although trial-by-trial variation in RT was tracked by activation fluctuations in key inhibitory control regions, such as anterior insula and pre-supplementary motor area, in TD children, this relation was markedly attenuated in children with ADHD [108]. These findings highlight not only diminished stability in neural activity but also compromised neurocognitive mechanisms underlying trial-wise response modulation in ADHD.
By leveraging advanced computational approaches such as state-space modeling and single-trial models, neurodynamic frameworks can offer novel insights into the brain mechanisms underlying cognitive deficits associated with ADHD.
Pharmacological effect on cognitive control and reward systems
ADHD medications, both stimulant and non-stimulant, are widely prescribed to alleviate core clinical symptoms of the disorder [212, 213]. A meta-analysis has shown that these treatments can have a protective effect across various domains of daily functioning, ranging from academic performance to road accidents [214]. However, therapeutic response is heterogeneous, with some individuals showing minimal benefit [215, 216]. This variability is likely to reflect underlying neurobiological differences, underscoring the need for biomarkers that can predict individual treatment response.
In line with cognitive models of ADHD, first-line stimulant medications have been consistently shown to enhance inhibitory control functions in affected individuals [217–219]. For instance, a recent study demonstrated that a single dose of methylphenidate improved inhibition accuracy and reduced behavioral variability in children with ADHD [220, 221]. Meta-analytic findings support these results, revealing that methylphenidate significantly improves performance on response inhibition tasks, such as stop-signal and go/no-go paradigms [217–219].
Beyond inhibitory control, stimulant medication may also enhance motivation. For example, amphetamine treatment has been shown to increase willingness to exert physical efforts [130], and methylphenidate has been found to enhance engagement in effortful tasks in adults with ADHD [129]. Additional behavioral studies report that methylphenidate increases reward sensitivity, reduces delay discounting, and decreases effort-related avoidance [222–224]. These findings suggest that stimulants can promote goal-directed behavior, particularly in tasks requiring sustained effort or delayed gratification. Nonetheless, motivational and reward-related effects have been less systematically studied than cognitive outcomes.
Neuropharmacological imaging studies provide mechanistic insights into these effects. Methylphenidate has been shown to modulate activity in brain regions implicated in both inhibitory control and reward processing, including the anterior insula, anterior cingulate cortex, and striatum [225, 226]. Although relatively few neuroimaging studies have investigated medication-induced changes in brain activation and connectivity during inhibitory control and reward processing tasks in children with ADHD [98, 178, 227, 228], a meta-analysis revealed that stimulant reliably enhances activation in the bilateral inferior frontal cortex and anterior insula during successful inhibition compared to placebo [229]. These findings support a neuropharmacological basis for the cognitive improvements observed with stimulant treatment.
At a cellular level, stimulant medications such as methylphenidate and amphetamine exert their effects by increasing extracellular dopamine and norepinephrine levels in the brain [230, 231]. Methylphenidate achieves this by blocking their respective transporters and inhibiting reuptake [232, 233]. Amphetamines not only inhibit reuptake but also promote presynaptic release of these catecholamines [234]. These mechanisms are thought to enhance the efficiency of frontostriatal and prefrontal cortical circuits involved in cognitive control, and likely contribute to increased reward sensitivity through their effects on mesolimbic dopamine pathways, key systems for motivation, reward anticipation, and reinforcement learning [118, 235].
In contrast, non-stimulant medications such as atomoxetine and guanfacine exert their effects through different neurochemical mechanisms. Atomoxetine is a selective norepinephrine reuptake inhibitor that primarily increases norepinephrine levels in the prefrontal cortex by blocking the norepinephrine transporter, thereby enhancing attention and inhibitory control [236–238]. Guanfacine, an α2A-adrenergic receptor agonist, enhances prefrontal cortical network regulation by inhibiting cAMP signaling, stabilizing neuronal firing, and supporting circuits underlying attention and working memory [239–241]. This suggests that stimulant medications may have broader effects encompassing both cognitive control and reward systems, whereas non-stimulants primarily target prefrontal executive dysfunction.
Recent research has begun to uncover how methylphenidate modulates atypical brain dynamics in children with ADHD, offering promising bridges to clinical application. For example, Mizuno et al. demonstrated that methylphenidate reduces aberrant dynamic interactions among the salience, frontoparietal, and default mode networks, effectively alleviating atypical dynamic network interactions in children with ADHD [221]. These findings were further replicated using advanced brain dynamic modelling approaches, reinforcing the robustness of this remediation effect [242]. Additionally, methylphenidate was shown to enhance dynamic neural activity in the nucleus accumbens, a key node in the reward circuitry [220]. Critically, these medication-induced alterations in the inhibitory control and reward systems were associated with corresponding improvements in inhibitory control performance, suggesting that changes in brain state dynamics translate to behavioral gains [220, 221, 242]. These findings revealed novel neurodynamic mechanisms of methylphenidate, offering circuit-level insights that can inform precision psychiatry.
Limitations, challenges and future research
Emerging research increasingly leverages computational models to characterize the dynamic latent cognitive and neural processes that underlie cognitive functioning. These models hold significant promise for uncovering abnormality in neurocognitive dynamics. By capturing fine-grained, time-resolved patterns of brain activity and behavior, computational frameworks such as state-space and single-trial models can offer mechanistic insights into the temporal dynamics of cognitive processes in ADHD.
Motivational deficit is mostly assessed through self-report and investigated using reward-dependent action-outcome contingency in experimental setting. However, these approaches serve only as indirect proxies rather than direct, quantitative measures of motivation, limiting mechanistic understanding of motivational deficits. Future effort should focus on developing novel experimental paradigms and computational models to accurately characterize human motivation [243, 244], potentially leveraging insights from advancements in animal research [245].
Neuroimaging research in ADHD faces several significant challenges. A key issue is the high drop-out rate, largely due to the young age and excessive head motion. Earlier studies were limited by small sample sizes. Large-scale neuroimaging initiatives, e.g. ABCD, HCP-D and IMAGEN, have begun to address this limitation. Furthermore, children with more severe symptoms are more likely to be excluded, leading to sampling bias. Although it is acknowledged in the field, a clear solution has yet to emerge. One potential avenue is the dimensional analysis, which may offer a way to mitigate bias by focusing on symptom continua.
Another major challenge is the intervention effect. A common approach is to implement a washout period for stimulant medications; however, the long-term effects of these treatments remain under investigation. Washout periods for non-stimulant medicines are longer and less feasible, further complicating study design. In recent years, neurostimulation techniques have emerged as promising new intervention options. However, these methods are still in their early stage and lack standardized parameters. Developing appropriate washout protocols for participants undergoing neurostimulation remains an open question.
No studies have systematically investigated how non-stimulant medications influence large-scale brain dynamics in children with ADHD. A deeper understanding of how different pharmacological agents modulate brain function could inform precision psychiatry, enabling treatment to be tailored to individuals’ neurobiological profiles.
Conclusion
Decades of behavioral and neuroimaging research have provided substantial evidence of impairments in both inhibitory control and motivation/reward systems in children with ADHD. However, the precise nature of these disrupted neural mechanisms and how they contribute to specific behavioral problems still remains poorly understood. We propose incorporating dynamic perspectives into existing neurocognitive models to examine how task-dependent brain dynamics influence downstream cognitive functioning and dysfunction in children with ADHD. Further research is critically needed to elucidate the dynamic interplay between motivation/reward and inhibitory control systems, which could yield valuable insights into the underlying heterogeneity of the disorder and developing effective and personalized interventions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Xu G, Strathearn L, Liu B, Yang B, Bao W. Twenty-year trends in diagnosed attention-deficit/hyperactivity disorder among US children and adolescents, 1997-2016. JAMA Pediatrics. 2018;1:e 181471.10.1001/jamanetworkopen.2018.1471 PMC 632428830646132 · doi ↗ · pubmed ↗
- 2Mistry, P, Branigan, NK, Gao, Z, Cai, W & Menon, V Computational modeling of proactive, reactive, and attentional dynamics in cognitive control. bio Rxiv [Preprint]. 2024. Available from: https://www.biorxiv.org/content/10.1101/2024.10.01.615613 v 1
- 3Tamm L, Menon V, Ringel J, Reiss AL. Event-related f MRI evidence of frontotemporal involvement in aberrant response inhibition and task switching in attention-deficit/hyperactivity disorder. J Am Acad Child Adolesc Psychiatry. 2004;43:1430–40. 10.1097/01.chi.0000140452.51205.8d.10.1097/01.chi.0000140452.51205.8d 15502603 · doi ↗ · pubmed ↗
- 4Cai W, Griffiths K, Korgaonkar MS, Williams LM, Menon V. Inhibition-related modulation of salience and fronto-parietal networks predicts cognitive control ability and inattention in children with ADHD. Mol Psychiatr. 2021;26:4016–25. 10.1038/s 41380-019-0564-4.10.1038/s 41380-019-0564-4PMC 718859631664176 · doi ↗ · pubmed ↗
- 5Misra J, Pessoa L. Brain dynamics and spatiotemporal trajectories during threat processing. e Life. 10.7554/e Life.102539.3.10.7554/e Life.102539 PMC 1285467341609638 · doi ↗ · pubmed ↗