Genome-wide identification and characterization of WOX gene family in saffron (Crocus sativus L.) and their roles in stress response, development and callus formation
Yuanyuan Tao, Jing Li, Jing Chen, Xiaoyuan Xi, Shuhui Yang, Feng Qiu, Xingchang Zhang, Mengqing Feng, Xiaodong Qian, Liqin Li

TL;DR
This study identifies and characterizes WOX genes in saffron, revealing their roles in stress response, development, and callus formation.
Contribution
The study provides the first genome-wide analysis of WOX genes in saffron and their functional roles in stress and development.
Findings
Twenty CsWOX genes were identified in saffron, with collinear and syntenic relationships revealed.
CsWUS and CsWOX13B expression decreased under cold stress but showed recovery trends.
CsWOX11 and CsWUS were upregulated in callus formation, with distinct expression patterns observed.
Abstract
The shoot apical meristem (SAM) of saffron (Crocus sativus L.) plays a critical role in floral transition and development. WUSCHEL-related homeobox (WOX) genes are key regulators of stem cell activity and SAM size across diverse species, including Arabidopsis thaliana, Oryza sativa Japonica, Gossypium hirsutum L. and the hybrid poplar clone 84K (Populus alba × P. glandulosa). WOX genes were identified and analyzed for phylogeny, gene structure, conserved motifs, cis-regulatory elements, chromosomal location, collinearity and duplication. Expression patterns were examined in different tissues, cold stresses, hormonal stress and callus formation, with key genes validated by real-time quantitative PCR. In this study, we identified 20 CsWOX genes based on the triploid genome of saffron. Intraspecific collinearity analysis revealed two collinear pairs (CsWOX7/CsWOX5 and CsWOX13A/CsWOX13B)…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
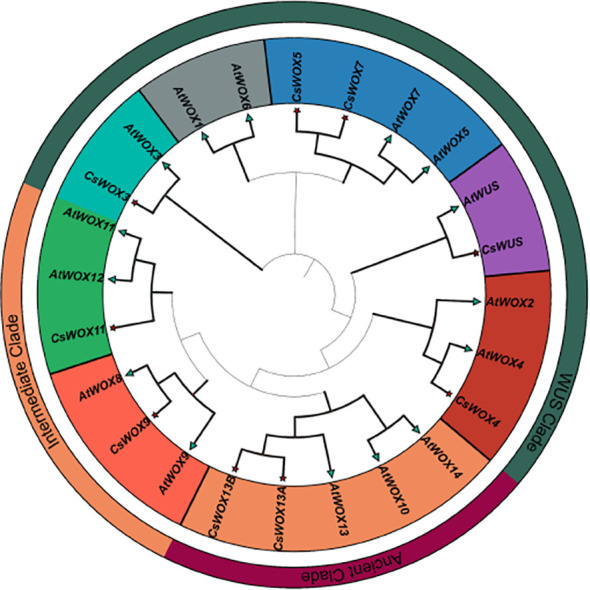 Figure 1
Figure 1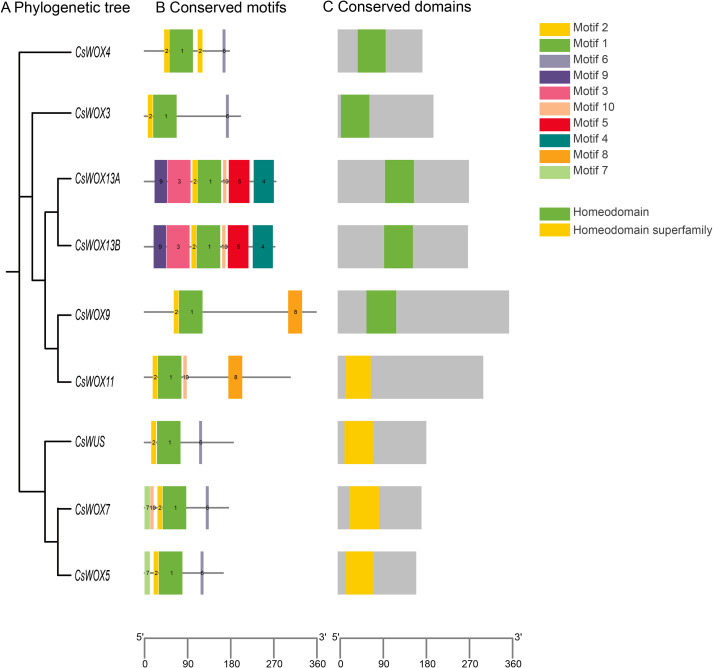 Figure 2
Figure 2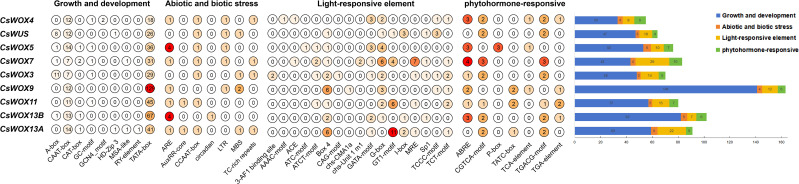 Figure 3
Figure 3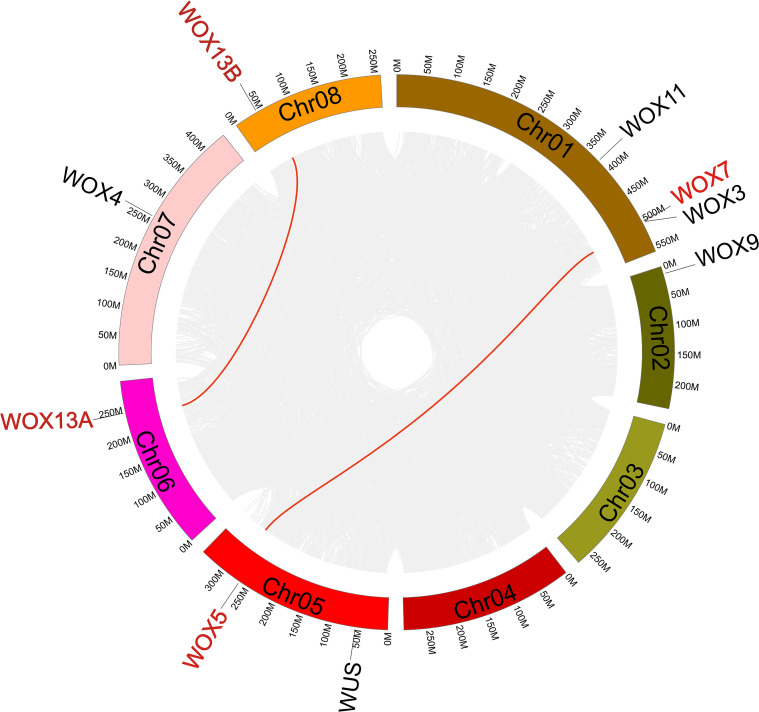 Figure 4
Figure 4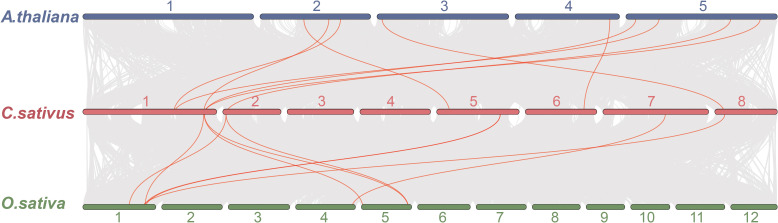 Figure 5
Figure 5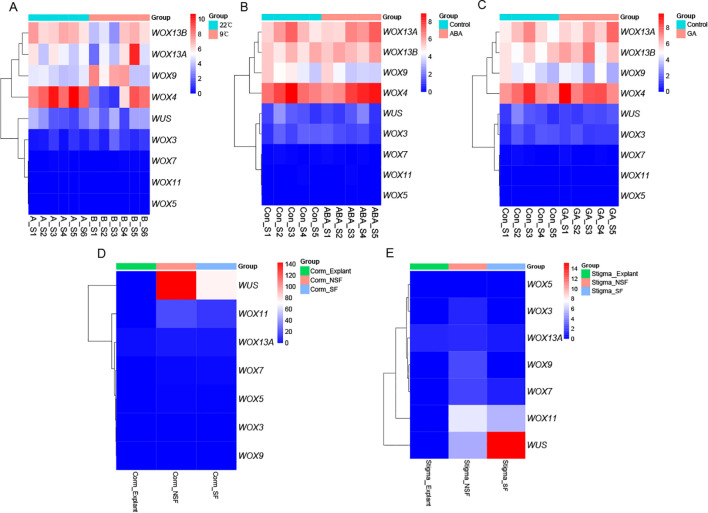 Figure 6
Figure 6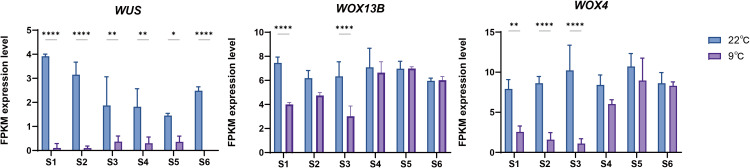 Figure 7
Figure 7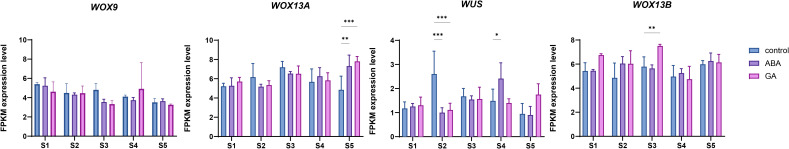 Figure 8
Figure 8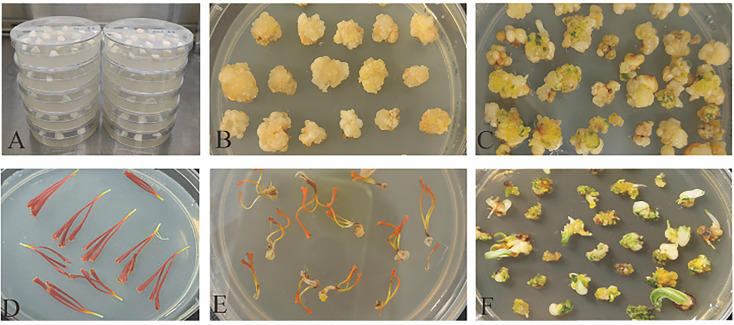 Figure 9
Figure 9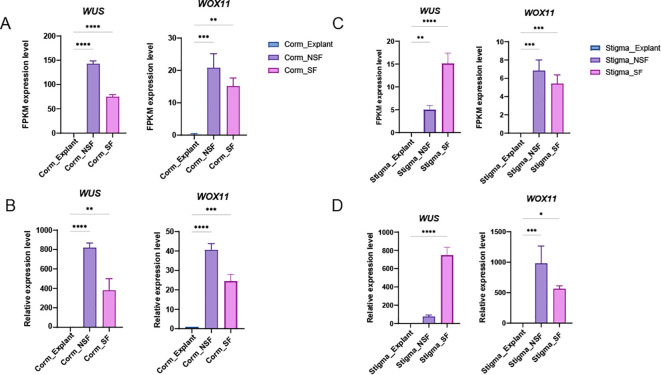 Figure 10
Figure 10Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Gene Expression Analysis · Saffron Plant Research Studies
Introduction
1
Saffron (Crocus sativus L.) is an autotriploid species that propagates exclusively through asexual reproduction. The shoot apical meristem (SAM) stem cells in its aerial part can differentiate into leaves, petals, stigmas and stamens, of which the stigma is of important medicinal value (Li et al., 2025). The WOX family is a group of plant specific transcription factors. Each member contains a highly conserved homeodomain (HD) composed of 65–66 amino acid. By regulating target gene expression at the transcriptional level, WOX genes regulate virtually every aspect of plant growth and development, including roots, stems, leaves, flowers, fruits, seeds and embryogenesis (Ueda et al., 2011; Romera-Branchat et al., 2013; Costanzo et al., 2014). Among them, WUS and WOX5 are essential for maintaining the balance between stem-cell self-renewal and differentiation in the SAM and root apical meristem (RAM), respectively, ensuring continuous development throughout the plant life cycle (Zhou et al., 2015). As per our knowledge, no studies on the WOX gene family in saffron have been reported. Investigating the WOX gene family may provide insights into improving the quality and yield of saffron.
The WOX gene family is classified into three major clades based on phylogenetic relationships and structural characteristics: the ancient clade (WOX10, WOX13, and WOX14), the intermediate clade (WOX8, WOX9, WOX11 and WOX12), and the modern clade (WUS, WOX1-WOX7). Members of these distinct clades exhibit significant functional diversification. Within the ancient clade, AtWOX13 inhibited the regeneration of pluripotent callus by suppressing WUS and STM, thereby directing cell differentiation toward turgid spherical cells (Ogura et al., 2023). In the intermediate clade, OsWOX11 recruits the ADA2-GCN5 histone acetyltransferase module to activate downstream targets in the crown root meristem (Zhou et al., 2017), and, together with CRL1, maintains cytokinin homeostasis to promote crown root development (Geng et al., 2023). Additionally, PagWOX11/12a in poplar confers salt tolerance by directly activating PagCYP736A12, attenuating ROS-induced damage (Wang et al., 2021). In the modern clade, AtWUS, the founding member, is expressed in the organizing center (OC) of the SAM, where the WUS–CLV3 negative feedback loop maintains the homeostasis of stem cell pool (Yadav et al., 2011). AtWUS cooperates with LEAFY (LFY) to activate AGAMOUS (AG) in floral meristems, which in turn terminates WUS expression to ensure floral determinacy (Lenhard et al., 2001). WOX1 and WOX3 act redundantly to control lateral leaf expansion (Nakata et al., 2012) and are crucial for leaf morphogenesis and margin patterning (Zhang et al., 2020; Xu et al., 2024a). AtWOX1 temporally modulates the number and size of leaf serrations through the known margin regulators CUC3 and BZR1 (Drieberg, 2025). AtWOX4 regulates vascular stem cell proliferation via the TDIF peptide signaling (Hirakawa et al., 2010), while OsWOX4 is cytokinin-inducible and regulates early leaf development (Cheng et al., 2014). Additionally, WOX5 and WOX7 regulate root development by sustaining callus proliferation through driving the cell cycle (Zhang et al., 2025). WOX5 and WOX7 are specifically expressed in the quiescent center (QC) across species (e.g., Arabidopsis, rice, and maize) and physically interact with PAT1 to modulate transcription of core cell-cycle genes (Yang et al., 2024).
WOX genes play crucial regulatory roles in vegetative organogenesis, reproductive organogenesis, tissue regeneration, callus formation and abiotic stress responses across diverse plant species (Zhou et al., 2024). To comprehensively characterize the structural and functional features of the WOX family in saffron, based on saffron genome (Li et al., 2025), we identified WOX family members and analyzed their gene structure, cis-regulatory elements, duplication events and collinearity. Furthermore, we profiled the expression patterns of CsWOX genes across diverse tissues and under abiotic stress conditions. Given that saffron is a high-value medicinal plant whose active compound accumulation is closely linked to its developmental processes, investigating the WOX gene family provides crucial insights into the molecular mechanisms governing saffron growth and establishes a foundation for molecular breeding strategies to enhance both the quality and yield of saffron.
Materials and methods
2
Identification of WOX gene family members in saffron
2.1
WOX protein sequences of Arabidopsis thaliana and Oryza sativa Japonica were retrieved from PlantTFDB v4.0 (https://planttfdb.gao-lab.org/) and used as queries for BLAST (E-value < 1e^−5^) against the saffron genome (Li et al., 2025). Additionally, a Hidden Markov Model (HMM) profile search was performed using HMMER 3.0, based on the conserved homeodomain of AtWOX proteins. The candidate CsWOX sequences identified by both BLAST and HMM searches were integrated. Subsequently, the conserved homeodomain was verified using the NCBI Conserved Domain Database (CDD) and SMART. Sequences missing the conserved HB (homeodomain) or redundant sequences were manually removed. Multiple sequence alignment and visualization of conserved motifs were performed using the DNAMAN 9 software.
Phylogenetic analysis and classification of CsWOX genes
2.2
Phylogenetic analysis was performed on non-redundant WOX protein sequences from C. sativus and A. thaliana. Multiple sequence alignment was performed using MUSCLE with default parameters, and a maximum-likelihood phylogenetic tree was constructed via FastTree based on the JTT model (Price et al., 2009). The resulting tree was visualized using the iTOL (https://itol.embl.de/).
Gene structure, conserved motif and cis-regulatory element analyses
2.3
Conserved motifs were identified with MEME Suite (https://meme-suite.org), with the maximum number of motifs set to 10. Homeodomain were further validated and annotated using the CDD. Cis-acting regulatory elements in the 2,000 bp promoter regions upstream of the start codon (ATG) were predicted using the PlantCARE database (Lescot et al., 2002). Visualizations of gene structures, conserved motifs and cis-regulatory elements were integrated using TBtools-II. Statistical analyses and visualization were performed using GraphPad Prism 10.
Chromosomal location, collinearity and gene duplication analysis of CsWOX
2.4
The chromosomal distributions of CsWOX genes were mapped using TBtools-II. Intraspecific collinearity and duplication events within saffron were identified using MCScanX with default parameters (Wang et al., 2024). The same methodology was applied to identify the duplication types of interspecific syntenic pairs between C. sativus and two model species, A. thaliana and O. sativa. All syntenic relationships and chromosomal localizations were visualized using Circos and TBtools-II.
Gene expression pattern in different tissues and under multiple stresses
2.5
Healthy, dormant corms (25 ± 1 g; Huzhou, China) were initially placed at 9 °C and 22 °C. Following dormancy release, the corms were transferred to 25 °C and 17 °C to induce the transition to reproductive growth. Apical buds were collected at six sequential developmental stages (S1-S6): including dormant (S1), dormancy-released (S2), early (S3), mid (S4) and late (S5) floral-bud differentiation, and floral-organ emergence (S6) (Xi et al., 2024). For hormonal treatments, corms (18–22 g; Bozhou, China) were treated with abscisic acid (ABA, 5 mg/L) and gibberellic acid (GA, 5 mg/L) for three consecutive days starting on 17 June and 8 July. The control group received an equal volume of distilled water. Apical buds were then collected at 3, 10, 13, 16 and 19 days after the initial treatment (Chen et al., 2025). Callus culture protocols: corms were subjected to a two-step sterilization procedure (Chib et al., 2020) and cultured on complete MS medium supplemented with 2,4-D (2 mg/L), BAP (1 mg/L) and NAA (1 mg/L). The cultures were maintained at 20 ± 2 °C with 45% relative humidity under a 16/8 h (light/dark) photoperiod. Stigmas explants were directly inoculated under the same conditions. Expression of CsWOX genes was quantified as Fragments Per Kilobase of transcript per Million mapped reads (FPKM) and visualized using heatmaps on the Microbial Bioinformatics platform. Statistical evaluation and line chart were performed using GraphPad Prism 10.
Real-time quantitative PCR validation
2.6
Total RNA was reverse-transcribed into cDNA using the PrimeScript RT Master Mix (Takara, Japan) from corm and stigma tissues of three distinct stages. Quantitative real-time PCR (qRT-PCR) was performed using TB Green Premix Ex Taq II (Takara, Japan) on an ABI 7500 Real-Time PCR System (Applied Biosystems, USA). All primers utilized in this study were listed in Supplementary Table 1. Three biological replicates were performed for each sample. The relative expression levels of the target genes were normalized and calculated using the 2^−ΔΔCT^ method.
Results
3
Whole-genome identification of the WOX genes family in saffron.
3.1
WOX proteins are characterized by a conserved homeodomain comprising three α-helices linked by loops and a turn. Conservative residues in Helix 1 include Q, L, and E; in Helix 2, P and L; and in Helix 3, N, V, W, F, Q, N, and R (Yu et al., 2022). By integrating Hidden Markov Model (HMM) profiles and BLASTP searches against the saffron genome database, a total of 20 CsWOX genes were identified (Supplementary Table 2). Multiple sequence alignment confirmed the high conservation of the characteristic homeodomain across all identified proteins (Supplementary Figure 1). Saffron is a triploid species, and our analysis revealed that the 20 CsWOX sequences represent nine distinct genes with varying homeologous copy numbers. Specifically, four genes harbored three homeologous copies across the three subgenomes (totaling 12 sequences), while three genes possessed two copies (totaling 6 sequences), and the remaining two genes were identified as single copy sequences. Homeologous copies shared >95% sequence similarity. Only one pair of copies exhibited a 51 amino acid gap (CSa_05_1G0005550/CSa_05_3G0005550), with the remaining regions maintaining high conservation. To ensure comprehensive coverage of key conserved motifs and functional domains for subsequent analyses, we selected one representative sequence for each of the nine CsWOX genes (Supplementary Table 3). These representative sequences, which exhibited the highest alignment with the NCBI database, were utilized for all subsequent structural, evolutionary, and expression profiling analyses. The amino acid sequences of CsWOX genes were subjected to systematic bioinformatic analyses, including molecular weight, theoretical pI, instability index, aliphatic index, and grand average of hydropathicity (GRAVY). As shown in Supplementary Table 3, CsWOX proteins comprised 163 (CsWOX5) to 357 (CsWOX9) amino acids, with molecular weights ranging from 18.49 kDa (CsWOX5) to 38.31 kDa (CsWOX9). Theoretical pI values ranged from 5.69–9.65, aliphatic index from 54.36–79.50, and instability index from 41.18–68.02, classifying them as unstable. CsWOX proteins exhibited negative GRAVY scores, indicating their intrinsic hydrophilicity.
Phylogenetic analysis and subfamily classification of CsWOX genes
3.2
To elucidate the evolutionary relationships within the WOX gene family between saffron and Arabidopsis, a phylogenetic tree was constructed using WOX protein sequences from A. thaliana and C. sativus (Figure 1). The results demonstrated that CsWOX members clustered into three conserved clades, consistent with the classification in Arabidopsis: the ancient clade (WOX10/13/14), the intermediate clade (WOX8/9, WOX11/12), and the modern/WUS clade (WOX1–7, WUS). Notably, orthologs corresponding to AtWOX1, AtWOX2, AtWOX6, AtWOX8, AtWOX10, AtWOX12, and AtWOX14 were not identified in the saffron genome.
Phylogenetic tree analysis of AtWOX (A. thaliana) and CsWOX (C. sativus) proteins. Green triangles and red stars represent A. thaliana and C. sativus, respectively. They were classified into three major phylogenetic clades: ancient, intermediate, and WUS, which were represented by distinct colors.
Conserved motifs and cis−regulatory element analysis of CsWOX genes
3.3
To further investigate the structural and functional diversity of CsWOX genes, we analyzed their conserved motifs (Figure 2) and cis-acting regulatory elements (Figure 3). Ten distinct motifs were identified across CsWOX proteins. Different motif combination modes suggested potential functional differences among family members. All CsWOX members shared Motif 1 and Motif 2 (Figure 2B), which together constituted the characteristic HD (Figure 2C). All CsWOX genes shared Motif 1 and Motif 2 (Figure 2B), which together constituted the characteristic HD (Figure 2C). Specifically, CsWUS and CsWOX3 contained Motifs 1, 2, and 6. CsWOX4 contained an additional Motif 2. CsWOX5 possessed Motifs 1, 2, 6, and 7, CsWOX7 carried Motifs 1, 2, 6, 7 and 10. CsWOX9 contained Motifs 1, 2, and 8, whereas CsWOX11 additionally included Motif 10. CsWOX13A/B lacked Motifs 6, 7, and 8, but retained the other seven motifs. These distinct motif patterns potentially underpin the functional specialization of different CsWOX clades.
Phylogenetic tree, conserved motifs and conserved domain analysis of CsWOX gene family members. (A) Phylogenetic tree. (B) Distribution of conserved motifs of CsWOX proteins. A total of 10 motifs were predicted, represented by colored boxes. The scale bar represents 90 aa. (C) Conserved domains of CsWOX proteins. The scale bar represents 90 aa.
Classification and statistics of predicted CREs on the putative promoter region of CsWOX genes. Four major CRE categories were identified: growth/development, stress response (abiotic and biotic), light response, and phytohormone response. Numbers and colors indicate the count of specific elements per gene.
Analysis of the 2,000 bp promoter regions using the PlantCARE database identified 40 types of CREs across the CsWOX genes (Figure 3; Supplementary Tables 4-1, 4-2). CREs were classified into four major categories, including abiotic stress responsive (7), hormone responsive (7), light responsive (17) and growth and development related (9). Among development related CREs, HD-Zip3 (vascular/embryo specificity) was present in CsWUS and CsWOX13A; MSA-like (meristematic root tips/callus) was detected in CsWOX7 and CsWOX13A; the CAT-box (root hair/meristem) occured in CsWOX7, CsWOX5, CsWOX13A/B. Regarding stress responses, LTR elements (low-temperature response) were found in CsWOX4/5/7/9 and CsWUS, suggesting their involvement in thermal adaptation. MBS elements (drought inducibility) were present in CsWOX3/9/13A and CsWUS. Light-responsive CREs (Box 4, G-box) were universally distributed across all CsWOX genes promoters. Notably, each CsWOX gene contained at least two hormone-responsive elements, including ABRE (ABA response) and MeJA-responsive elements (CGTCA-motif or TGACG-motif), which showed extremely high co-occurrence. This pattern suggested that CsWOX genes may be co-regulated by ABA and MeJA signaling pathways (Soma et al., 2021; Xu et al., 2024b). Additionally, GA (TATC-box, P-box), IAA (TGA-element), and SA (TCA-element) responsive elements were differentially distributed among family members, indicating diverse regulatory potentials.
Chromosomal location, collinearity and evolution analysis of CsWOX genes
3.4
The CsWOX genes were distributed across all chromosomes except for Chr03 and Chr04. Chr01 and Chr05 each contained three members, whereas the remaining three chromosomes each contained a gene (Figure 4). Gene duplication events in the saffron genome were classified into five categories using the MCScanX method, including singleton (1,584 genes), dispersed (5,372 genes), proximal (947 genes), tandem (1,415 genes), and segmental (73,439 genes). The high proportion of segmental events indicated that saffron may have undergone whole genome duplication (WGD) or large segment duplication events. The identification of 5,372 dispersed genes further suggested frequent small segment genomic rearrangements or transposition events, while the paucity of proximal and tandem duplications suggested that local gene amplification was not a primary driver of genome expansion. Among the nine CsWOX genes, two WGD gene pairs (CsWOX13A/CsWOX13B and CsWOX7/CsWOX5) and one dispersed gene pair (CsWOX11/CsWOX3) were identified (Supplementary Table 5), with no tandem duplication events detected within the family.
Distribution and colinearity of CsWOX genes within the saffron genome. Colinearity genes were highlighted in red. CsWOX genes lacking colinearity were marked in black.
To further explore the evolutionary history of the WOX family, we performed interspecific collinearity analysis between C. sativus and two model species, A. thaliana and O. sativa (Figure 5). One segmental duplication pair (AtWUS/CsWUS) and eight dispersed duplication pairs were identified in A. thaliana and C. sativus. Three segmental duplications pairs (OsWOX9/CsWOX5, OsWOX9/CsWOX7, and OsWOX2/CsWOX3) and six dispersed duplications pairs were identified in O. sativa and C. sativus (Supplementary Table 4). No tandem duplications were found in either comparison. These syntenic relationships suggested that CsWUS, CsWOX5, CsWOX7, and CsWOX3 are evolutionarily conserved and likely retain functions analogous to their orthologs in A. thaliana and O. sativa.
Collinearity analysis of WOX genes between C. sativus both A. thaliana and O. sativa. Colinearity genes were highlighted in red.
Expression patterns of CsWOX genes
3.5
The SAM development of saffron daughter corms progresses through distinct stages, beginning with dormancy and its subsequent release, followed by floral primordium differentiation, and culminating in floral organ maturation. Because the activity of SAM stem cells was influenced by temperature (de Jonge et al., 2016), we investigated CsWOX genes expression under different temperatures. As previously reported by our group, corms were stored at 9 °C and 22 °C until dormancy termination. Corms stored at 9 °C exhibited leaf development but failed to differentiate floral buds, whereas those maintained at 22 °C successfully produced both fully developed leaves and floral buds (Xi et al., 2024; Li et al., 2025). To investigate the response of CsWOX genes to cold stress, we analyzed RNA-seq data to characterize expression profiles in the SAM under 9 °C and 22 °C treatments (Figure 6A). At 22 °C, CsWUS showed a gradual downregulation from stages S1 to S6, with a slight increase at S6, whereas the remaining family members exhibited no significant changes. CsWX13B and CsWOX4 were highly expressed at 22 °C, while CsWOX11, CsWOX7, and CsWOX5 were nearly silent. Notably, cold treatment (9 °C) significantly suppressed CsWUS expression throughout stages S1-S6 compared to the 22 °C. Although CsWUS showed slight recovery during stages S4–S5, its transcripts were undetectable by stage S6. CsWOX13B and CsWOX4 were markedly downregulated in S1–S3, but recovered during stages S4–S6 (Figure 7). These results indicate that cold treatment irreversibly downregulates WUS expression, whereas its inhibitory effects on CsWOX13B and CsWOX4 are reversible.
CsWOX genes expression patterns. (A) CsWOX genes expression patterns of SAM with different temperatures treatments. (B) The response of CsWOX genes in the SAM to ABA. (C) The response of CsWOX genes in the SAM to GA. (D) CsWOX genes expression patterns with corm derived callus. (E) CsWOX genes expression patterns with stigma derived callus. NSF and SF represent the non-shoot-forming and shoot-forming callus stages, respectively. Expression levels are represented by color, with red and blue indicating high and low expression, respectively.
*Expression levels of CsWUS, CsWOX13B and CsWOX4 in SAM under 9 °C and 22 °C treatment. Asterisks indicate signifcant differences in transcript levels compared with 22 °C treatment (*P < 0.05, **P < 0.01, ***P < 0.0001).
Our previous studies have indicated that GA and ABA promote the transition of apical meristem differentiation to floral bud differentiation (Chen et al., 2025). During the dormant period, only CsWOX9 declined gradually in the control group, with no significant trends observed for other CsWOX members. ABA treatment specifically upregulated CsWOX13A at S5 and CsWUS at S4. GA induced a progressive increase in CsWOX13B at S3 and CsWOX13A at S5. In summary, although GA and ABA promoted the transition to floral meristems, most CsWOX genes were either unresponsive or respond only transiently to GA and ABA (Figures 6B, C, 8).
*Expression levels of CsWOX9, CsWOX13A, CsWUS and CsWOX13B in SAM under ABA and GA treatment. Asterisks indicate signifcant differences in transcript levels compared with control group (*P < 0.05, **P < 0.01, **P < 0.001).
The WOX gene family has been widely utilized to enhance plant tissue and organ regeneration (Jia et al., 2025). Callus induction from corm and stigma explants was shown in Figure 9. In corm-derived callus, all nine CsWOX genes were nearly undetectable at explant, with only seven CsWOX genes recovered at the non-shoot-forming (NSF) and shoot-forming (SF) callus stages (Figure 6D). Among these, WOX3, WOX5 and WOX9 exhibited the lowest expression levels, while WOX7 and WOX13A showed slight expression. In corm-derived callus, CsWUS and CsWOX11 were significantly upregulated at the NSF and SF stages. Notably, while CsWOX11 exhibited a similar expression pattern across both corm and stigma sources, CsWUS showed a distinct, progressive increase in expression during the NSF and SF stages of stigma-derived callus (Figures 6E, 10). These expression profiles were further validated via qRT–PCR (Figure 10). These results suggest that CsWUS and CsWOX11 coordinately regulate callus proliferation, and their divergent expression patterns across explant types indicate functional specialization.
Corm and stigma induced callus formation. (A) Corm explants. (B) Corm-induced callus (no buds). (C) Corm-induced callus (with buds). (D) Stigma explants. (E) Stigma-induced callus (no buds). (F) Stigma-induced callus (with buds).
*Expression level of CsWUS and CsWOX11 in corm and stigma derived callus and qRT–PCR validation. (A) Expression level of CsWUS and CsWOX11 in corm derived callus and (B) qRT–PCR validation. (C) Expression level of CsWUS and CsWOX11 in stigma derived callus and (D) qRT–PCR validation. NSF and SF represent the non-shoot-forming and shoot-forming callus stages, respectively. Asterisks indicate signifcant differences in transcript levels compared with explant group (*P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001).
We expand the expression profiles of CsWOX genes, we examined additional tissues including roots at the flowering stage, four developmental stages of stigma (1–4), and vegetative organs from the propagation stage: mother corm, leaf, and daughter corm (Supplementary Figure 2). Results showed that only CsWOX13B, CsWOX13A, CsWOX4 and CsWOX9 were detectable across these tissues, while other members were not detected. CsWOX13B exhibited progressive upregulation during stigma development, peaking at the stigma 4 stage, where expression was significantly higher than in roots. Conversely, CsWOX13B expression remained minimal in mother corms, leaves, and daughter corms. In contrast, CsWOX13A exhibited marked upregulation in roots, mother corms, leaves, and daughter corms. Although its overall abundance in stigmas was relatively lower, CsWOX13A still demonstrated a significant upward trend from stigma 1 to stigma 4 during stigma development. CsWOX4 and CsWOX9 were expressed in flowering roots and stigmas 1, with their expression in the stigma 1 significantly exceeding that observed in roots.
Discussion
4
The WOX gene family is crucial for maintaining the balance between stem cell self-renewal and differentiation, ensuring continuous plant growth throughout the life cycle (Dolzblasz et al., 2016). In C. sativus, we identified 20 WOX genes. Several genes harbored three homeologous copies across the three subgenomes, while others exhibit gene loss or pseudogenization. This phenomenon may stem from specific subgenome gene deletions, pseudogenization, or regional assembly gaps. Representative copies with the most complete ORFs and conserved domains were selected from each homeologous group to minimize redundancy and ensure reliable evolutionary reconstruction. Transcriptome analysis confirmed that these highly similar copies (>95% identity) displayed nearly identical expression profiles across developmental stages (Supplementary Figures 3–8), indicating functional redundancy and validating our representative-based approach.
Phylogenetic analysis revealed that CsWOX genes were distributed across the ancient, intermediate, and WUS clades, consistent with Arabidopsis (Figure 1). However, orthologs corresponding to AtWOX1, AtWOX2, AtWOX6, AtWOX8, AtWOX10, AtWOX12, and AtWOX14 were not identified in the saffron genome. The absence of these orthologs may reflect functional redundancy among clade members or evolutionary gene loss. For example, AtWOX5 and AtWOX7 are the closest homologs (Yang et al., 2024). AtWOX11 and AtWOX12 function redundantly (Wang et al., 2021). WOX4 acts redundantly with WOX14 downstream of the PXY signaling pathway to promote vascular cell proliferation (Etchells et al., 2013). AtWOX10 may be a pseudogene, as no transcripts have ever been detected (Deveaux et al., 2008). Intriguingly, two WOX13 copies were presented in saffron, which also observed in Akebia trifoliata and Gossypium raimondii (Yang et al., 2017; Han et al., 2024). Genes with analogous functions may have been eliminated during evolution (Chen et al., 2009).
Collinearity analysis revealed that 88.74% of saffron genes originated from WGD and segmental duplications (Supplementary Table 5). This likely stemmed from two critical whole-genome triploidy events during plant evolution (Xu et al., 2024c). Within the CsWOX family, CsWOX7/CsWOX5 and CsWOX13A/CsWOX13B were identified as WGD-derived duplicates, indicating that saffron has experienced a WGD event. Additionally, the collinear relationships of OsWOX9/CsWOX5, OsWOX9/CsWOX7, OsWOX2/CsWOX3 and AtWUS/CsWUS further confirmed the conserved homology of WOX family genes among C. sativus, A. thaliana and O. sativa.
The SAM in bulbous species like Lilium candidum and C. sativus is highly sensitive to low temperatures that suppress bolting and flowering (Mazor et al., 2021). Our previous study revealed that cold exposure (9 °C) during corm dormancy enabled leaf differentiation but entirely suppressed floral organ formation (Li et al., 2025). Transcriptome analysis of the SAM revealed that cold treatment significantly suppressed CsWUS expression from stages S1 to S6. Similarly, in tomato, SlWUS expression decreases significantly under cold stress. Enhanced SlWUS expression is associated with the MF formation (Wu et al., 2023). This suggests that the low-temperature mediated inhibition of CsWUS likely disrupts the stem cell niche required for floral organ differentiation, thereby terminating flowering. Intriguingly, while CsWOX13B and CsWOX4 were initially suppressed by cold (S1–S3), their expression levels recovered during later stages (S4–S6). Given that WOX4 and WOX13 are involved in leaf primordia vascular development and grafting success (Ji et al., 2010; Yue et al., 2020), their resilient expression profiles may explain why leaf development remains unaffected by cold stress while floral organs fail to form.
During corm dormancy, we exogenously applied ABA and GA to SAM of saffron. GA significantly enhanced floral primordium differentiation, ABA produced positive but non-significant effect, whereas CK showed no promotion at all. Nevertheless, no significant trend changes were observed in CsWOX genes, with almost no variation detected. GA and ABA drive floral transition and floral organ primordia differentiation in Orchid Cymbidium Sinense by activating the expression of floral meristem specific genes AP1 (Ahmad et al., 2024). CK and WUS genes synergistically interact through complex feedback loops in SAMs, ensuring the balance between stem cell homeostasis and organogenesis (Lee et al., 2016). This suggest that CsWOX genes may maintaining stem cell homeostasis and remain less responsive to hormone induced floral primordia initiation during dormancy.
Callus culture is valued as a core tool in agriculture and horticulture (Guo et al., 2015; Efferth, 2019), enabling the development of transgenic lines with enhanced tolerance to abiotic and biotic stresses (Benhayyim and Goffer, 1989; Chavarriaga-Aguirre et al., 2016), and improved yield and nutritional quality (Bahgat et al., 2009). Studies have shown that AtWOX11 is rapidly activated under high auxin conditions to initiate callus formation via LBD16 and WOX5/7 (Sheng et al., 2017), whereas cytokinin signaling triggers WUS expression through chromatin remodeling and HD-ZIP III–type-B ARR complex formation to establish shoot primordia (Zhang et al., 2017; Wan et al., 2023; Liu et al., 2025). Our results suggest that CsWUS and CsWOX11 were concurrently and significantly upregulated at the NSF stage and downregulated at the SF stage in corm-derived callus. Notably, while CsWOX11 exhibited similar expression pattern both corm and stigma callus, CsWUS showed a distinct, progressive increase during the NSF and SF stages of stigma-derived callus. These results indicate that the roles of CsWUS and CsWOX11 in callus formation and differentiation (Ikeda and Ohme-Takagi, 2014) are conserved across multiple model plants, including Arabidopsis thaliana, Gossypium hirsutum L. and the hybrid poplar clone 84K (Populus alba × P. glandulosa) (Liu et al., 2014, 2018; Wei et al., 2022). Additionally, variations in the expression of CsWOX11 and CsWUS across different plant species and explant types may reflect distinct regeneration pathways, regulatory variations within a conserved pathway, or species-specific expression programs. In this study, callus developmental stages were defined solely by morphological criteria, which may not fully capture the underlying molecular heterogeneity. Comprehensive characterization of callus development requires integrating morphological, molecular, and functional approaches. Future studies are needed to determine whether CsWOX11 and CsWUS act coordinately or sequentially in the saffron regeneration pathway.
Following flowering, saffron transitions to a vegetative phase where mother corms supply carbon to developing daughter corms (Jose-Santhi et al., 2024). Most CsWOX genes were exhibited either no expression or low expression levels during this stage, similar to the expression patterns of SsuWOX genes (Jia et al., 2025). Notably, CsWOX13B expression increased progressively throughout the four stages of stigma development, peaking at stage stigma 4. GhWOX13 was highly expressed at 10 and 15 DPA (days post anthesis) and involved in phytohormone-mediated fiber development in cotton (He et al., 2019). Similarly, AtWOX13 plays important role in controlling the medio lateral pattern of the fruit development, which is fundamental for seed dispersal (Romera-Branchat et al., 2013). This indicated the functional diversity of CsWOX13.
Conclusion
5
Based on the saffron genome, the study identified 20 CsWOX gene copies. Phylogenetic analysis revealed that the saffron WOX gene family can be classified into three conserved clades: the ancient, intermediate, and WUS clades. Analyses of conserved motif and cis−regulatory elements indicated that genes within the same clade exhibit similar structures and functions. Collinearity analysis identified two WGD derived duplicate pairs (CsWOX7/5 and CsWOX13A/B). Interspecific synteny analysis further confirmed the conserved homology of WOX family genes among Saffron, Arabidopsis, and Rice. During daughter corm development following saffron flowering, most CsWOX genes exhibited low or undetectable expression in roots, mother corms, leaves, and daughter corms. Notably, CsWOX13A and CsWOX13B showed progressively increasing expression concomitant with stigma maturation, peaking at stage stigma 4. Crucially, the downregulation of CsWUS in the SAM following cold treatment was identified as a potential molecular trigger for floral primordia abortion. No significant expression changes were observed in CsWOX genes after ABA or GA treatment of SAM. During callus induction from corm and stigma explants, both CsWOX11 and CsWUS were significantly upregulated in the NSF and SF callus, supporting their conserved roles in cell dedifferentiation and regeneration. In summary, this study provides a systematic analysis of the WOX gene family in saffron, offering valuable insights for subsequent functional studies of CsWOX genes and establishing a theoretical foundation for practical applications in saffron meristem regulation, callus culture optimization, and molecular breeding.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahmad S. Lu C. Gao J. Wei Y. Xie Q. Jin J. . (2024). Integrated proteomic, transcriptomic, and metabolomic profiling reveals that the gibberellin–abscisic acid hub runs flower development in the Chinese orchid Cymbidium sinense. Hortic. Res. 11, uhae 073. doi: 10.1093/hr/uhae 073. PMID: 38738212 PMC 11088716 · doi ↗ · pubmed ↗
- 2Bahgat S. Shabban O. A. El-Shihy O. Lightfoot D. A. El-Shemy H. A. (2009). Establishment of the regeneration system for Vicia faba L. Curr. Issues Mol. Biol. 11, I 47–i 54. doi: 10.21775/cimb.011.i 47. PMID: 19193964 · doi ↗ · pubmed ↗
- 3Benhayyim G. Goffer Y. (1989). Plantlet regeneration from a Na Cl-selected salt-tolerant callus culture of Shamouti orange (Citrus sinensis L. Osbeck). Plant Cell Rep. 7, 680–683. doi: 10.1007/BF 00272060. PMID: 24240461 · doi ↗ · pubmed ↗
- 4Chavarriaga-Aguirre P. Brand A. Medina A. Prias M. Escobar R. Martinez J. . (2016). The potential of using biotechnology to improve cassava: a review. In Vitro Cell. Dev. Biology-Plant 52, 461–478. doi: 10.1007/s 11627-016-9776-3. PMID: 27818605 PMC 5071364 · doi ↗ · pubmed ↗
- 5Chen J. Yang S. Qian X. Zhang X. Tao Y. Li J. . (2025). The impact of phytohormones on the number and quality of flowers in Crocus sativus. BMC Plant Biol. 25, 683. doi: 10.1186/s 12870-025-06712-6. PMID: 40410691 PMC 12100840 · doi ↗ · pubmed ↗
- 6Chen S. K. Kurdyukov S. Kereszt A. Wang X. D. Gresshoff P. M. Rose R. J. (2009). The association of homeobox gene expression with stem cell formation and morphogenesis in cultured Medicago truncatula. Planta 230, 827–840. doi: 10.1007/s 00425-009-0988-1. PMID: 19639337 PMC 2729979 · doi ↗ · pubmed ↗
- 7Cheng S. Huang Y. Zhu N. Zhao Y. (2014). The rice WUSCHEL-related homeobox genes are involved in reproductive organ development, hormone signaling and abiotic stress response. Gene 549, 266–274. doi: 10.1016/j.gene.2014.08.003. PMID: 25106855 · doi ↗ · pubmed ↗
- 8Chib S. Thangaraj A. Kaul S. Dhar M. K. Kaul T. (2020). Development of a system for efficient callus production, somatic embryogenesis and gene editing using CRISPR/Cas 9 in Saffron (Crocus sativus L.). Plant Methods 16, 47. doi: 10.1186/s 13007-020-00589-2. PMID: 32280363 PMC 7137501 · doi ↗ · pubmed ↗