AAV2-Retro–Mediated Gene Transfer Selectively Targets Outer Retinal Cells Following Intravitreal Injection
Chaimaa Kinane, Moxa Panchal, Pantelis Tsoulfas, Venu Talla, Kevin K. Park

TL;DR
This study shows that AAV2-retro can efficiently deliver genes to outer retinal cells in mice after a simple eye injection.
Contribution
AAV2-retro enables outer retina-specific gene delivery via intravitreal injection, offering a less invasive alternative to subretinal delivery.
Findings
AAV2-retro targets photoreceptors and retinal pigment epithelium with strong efficiency.
Transduction is limited to outer retinal layers, with minimal inner nuclear layer involvement.
Sequential injections enhance reporter expression and spatial coverage.
Abstract
To characterize the cellular tropism and temporal dynamics of adeno-associated virus 2 (AAV2)-retro–mediated gene delivery in the adult mouse retina following intravitreal injection. Adult C57BL/6J mice received single or sequential intravitreal injections of AAV2-retro carrying the mGreenLantern (mGL) reporter gene. Retinas were collected at 1, 3, and 14 days post-injection (dpi) and processed for immunofluorescence analysis. Transduced cell types were identified using cell-type markers, including cone arrestin, RNA-binding protein with multiple splicing (RBPMS), and activator protein 2α (AP-2α). The number and distribution of mGL-positive cells were quantified on whole retinas or retinal cross-sections to assess transduction efficiency, specificity, and spatial coverage. Reporter expression was detected in the outer retina at 1 dpi and increased markedly at 3 and 14 dpi. AAV2-retro…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
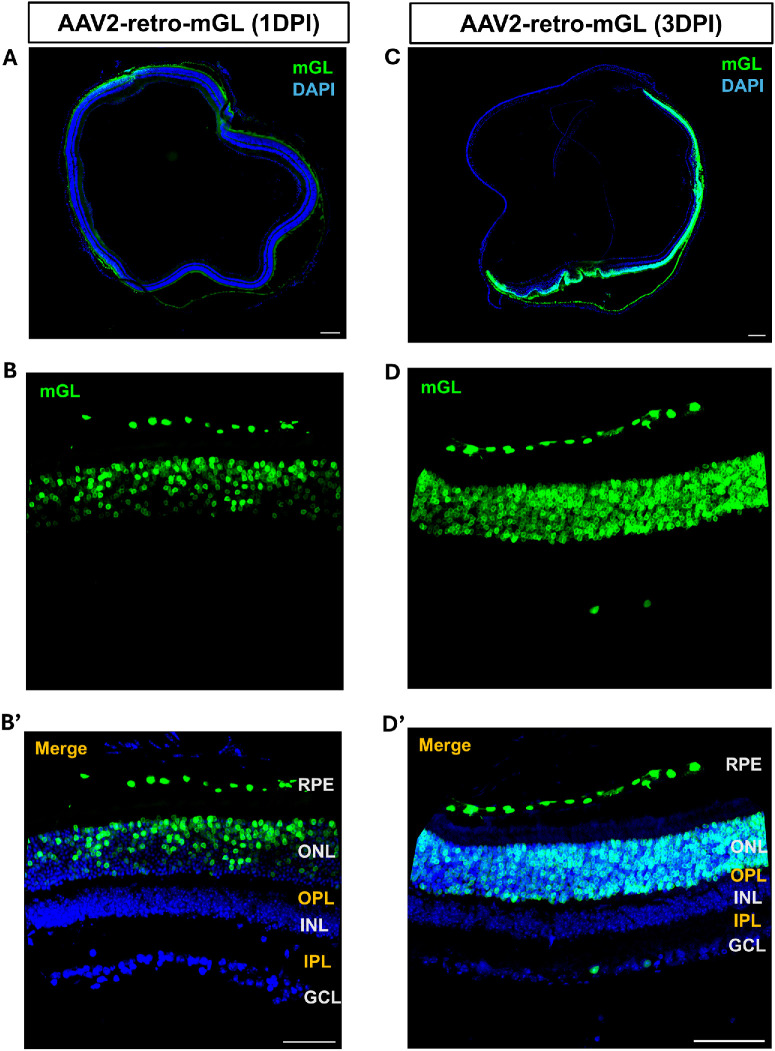 Figure 1
Figure 1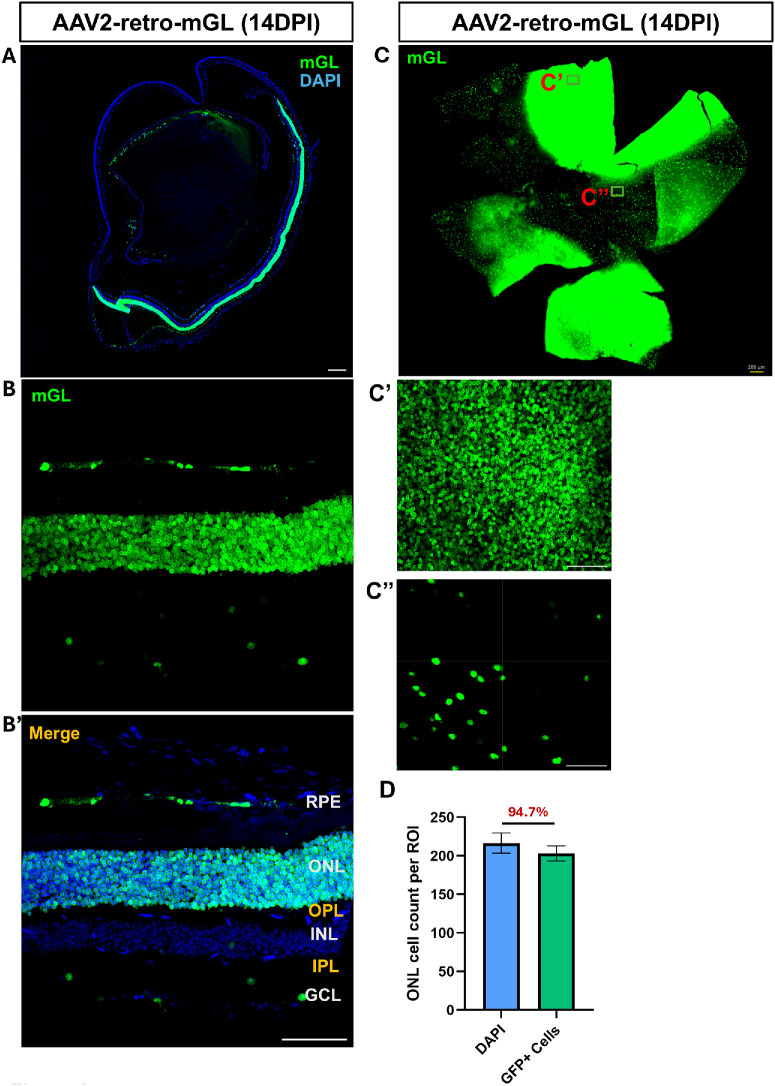 Figure 2
Figure 2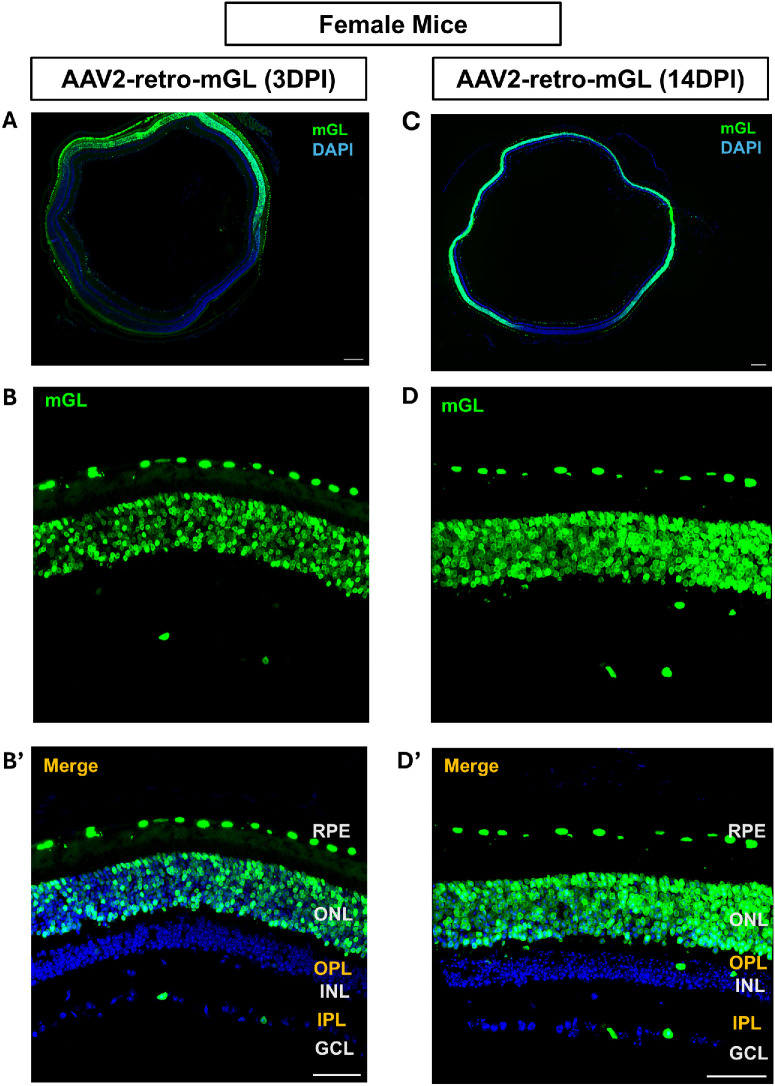 Figure 3
Figure 3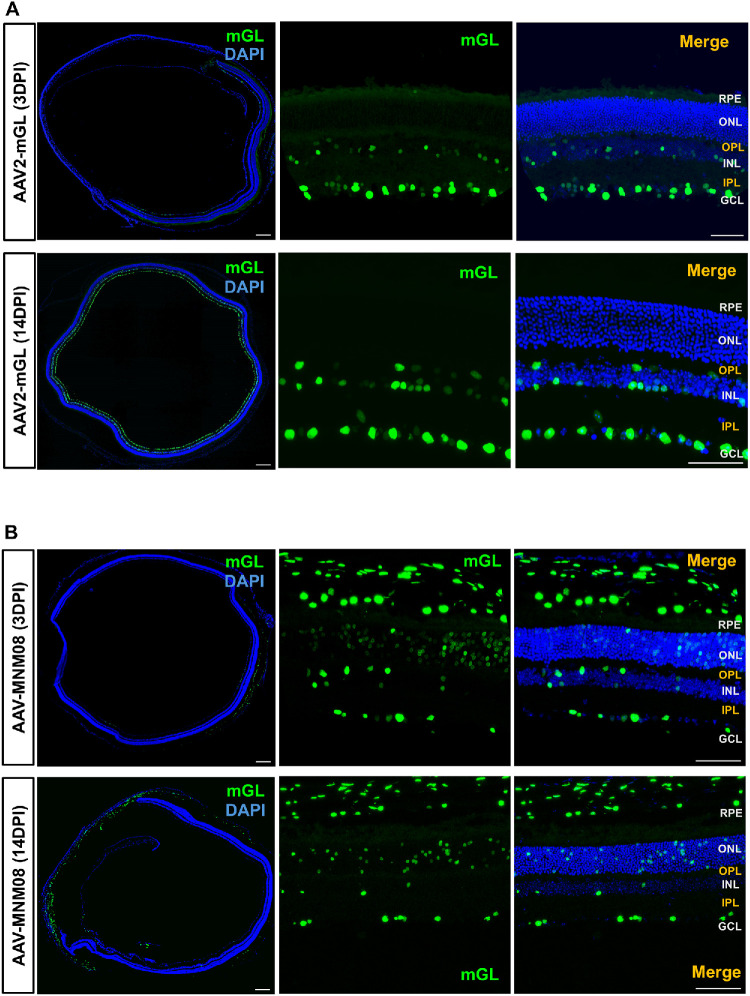 Figure 4
Figure 4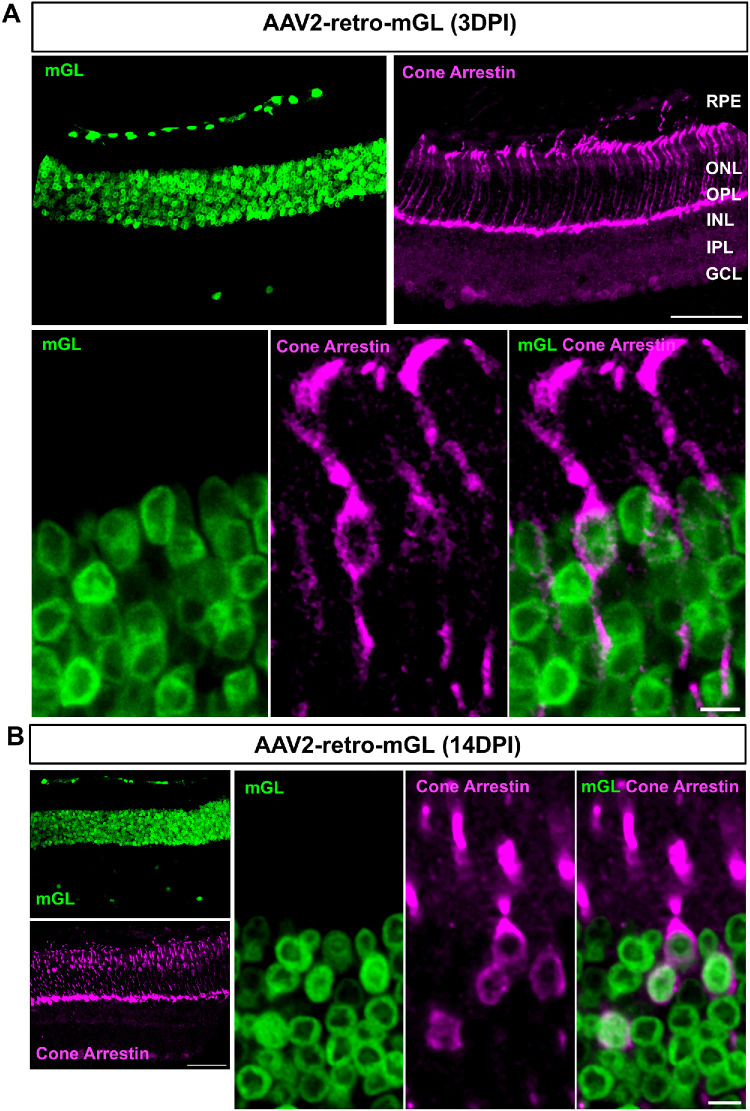 Figure 5
Figure 5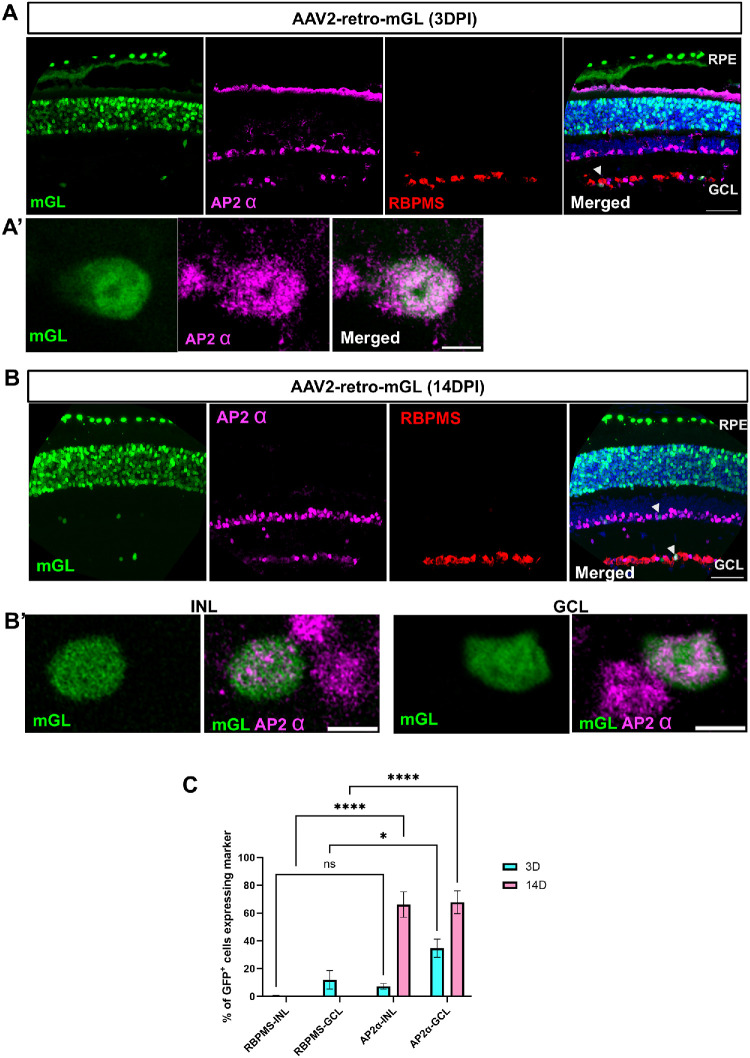 Figure 6
Figure 6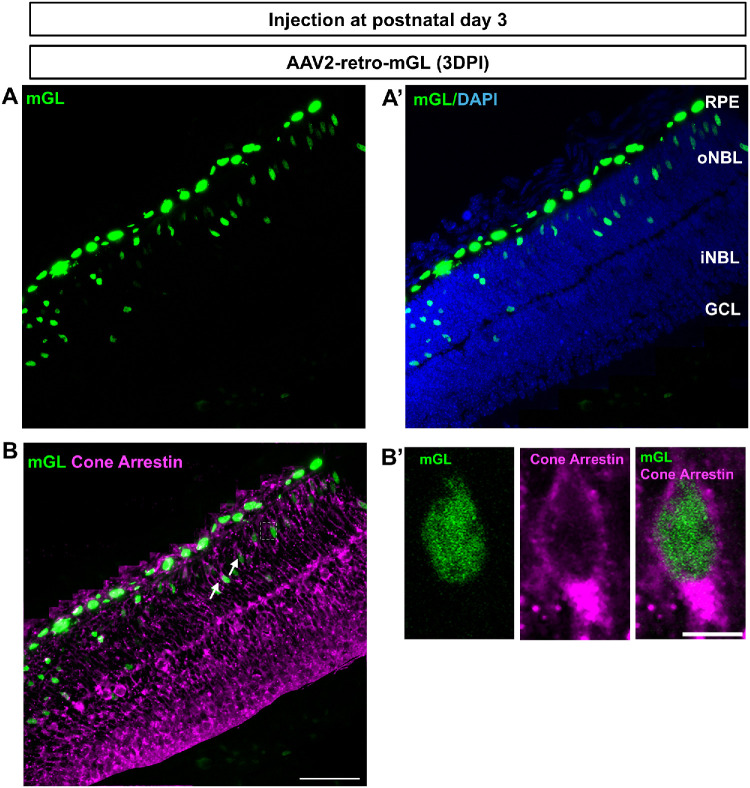 Figure 7
Figure 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRetinal Development and Disorders · Virus-based gene therapy research · CRISPR and Genetic Engineering
Inherited and acquired retinal degenerative diseases are a leading cause of irreversible blindness worldwide, primarily due to the loss of photoreceptors or dysfunction of the supporting retinal pigment epithelium (RPE).1^–^3 Gene therapies targeting these cells offer a promising avenue for restoring vision or halting disease progression.4^–^7 Recombinant adeno-associated virus (rAAV) vectors have emerged as a key platform in this field, demonstrating sustained gene expression, low immunogenicity, and a strong safety profile in both ocular and non-ocular applications. In particular, rAAV-mediated gene transfer to photoreceptors and RPE has shown significant therapeutic potential in preclinical models and human clinical trials.7^–^11 Currently, a widely used method for delivering AAV vectors to photoreceptors is subretinal injections, which place the vector in direct contact with the outer retina.12^–^14 This approach is exemplified by the U.S. Food and Drug Administration–approved gene therapy voretigene neparvovec (Luxturna), which uses AAV serotype 2 (AAV2) to restore RPE65 function in patients with inherited retinal dystrophy.15 However, subretinal delivery requires a complex surgical procedure involving retinal detachment and the creation of a transient subretinal bleb.16 This is not only technically demanding but also poses risks to an already fragile retina, particularly in late-stage disease when retinal structure is severely compromised.17^,^18 Moreover, subretinal injections typically result in localized gene expression confined to the area of the bleb, as conventional AAV capsids are limited in their ability to diffuse laterally through retinal tissue.4^,^19 Consequently, only a portion of the retina is transduced, which may be insufficient to halt disease progression or restore functional vision, especially in conditions that require widespread retinal coverage. These limitations underscore a critical need for alternative delivery strategies that are less invasive and more broadly effective.
Intravitreal injection, by contrast, is a well-established, minimally invasive technique commonly used for the delivery of pharmacological agents in clinical ophthalmology. Its accessibility and repeatability make it an attractive route for ocular gene therapy. However, the inner limiting membrane (ILM) and other physical barriers of the inner retina hinder AAV access to deeper retinal layers, including photoreceptors and the RPE, limiting the utility of traditional AAV serotypes when delivered intravitreally.20^–^23
Recent studies have identified engineered AAV capsids with enhanced ability to traverse retinal barriers and reach the outer retina from the vitreous cavity.23^–^26 Others have identified promoters that confer highly selective gene expression in the outer retina.27 These novel AAV variants and promoters are under active investigation and hold significant promise, although their efficiency, safety, and translational readiness for broad clinical application are being established through ongoing studies.
AAV2-retro is an engineered AAV capsid variant derived from AAV2, specifically developed to enhance retrograde transport in neurons. Originally described by Tervo et al.28 in 2016, AAV2-retro enables robust gene delivery to projection neurons by traveling against the direction of synaptic transmission. In this study, we initially sought to determine whether AAV2-retro, when delivered via intravitreal injection, exhibits a preferential pattern of transduction in retinal neurons that reflects retrograde transport through the visual circuitry and, if so, identify which neuronal cell types in the retina are most efficiently transduced. Here we report that intravitreal delivery of AAV2-retro resulted in robust transduction of photoreceptors and RPE cells within 1 day, demonstrating a rapid onset and striking cell-type specificity for these cell types in the adult mouse retina. These findings highlight the unexpected capability of AAV2-retro to bypass inner retinal barriers. The capacity to target outer retinal cells non-invasively in adult animals offers major advantages for preclinical studies, enabling more accessible, reproducible, and less damaging gene delivery in animal models of retinal degeneration and repair.
Materials and Methods
Ethical Review
All animal procedures were conducted in compliance with protocols approved by the Institutional Animal Care and Use Committee at University of Texas Southwestern Medical Center (UTSW) and in accordance with the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research.
Animals
Male and female C57BL/6J mice (6–8 weeks old, 000664; The Jackson Laboratory, Bar Harbor, ME, USA) were used for this study. Postnatal day 3 (P3) pups, regardless of sex, were used for the postnatal injection experiment. Mice were housed in individually ventilated cages under a 12-hour light/dark cycle. Food and water were provided ad libitum, and nesting material and shelters were added to all mice cages for environment enrichment.
Intravitreal Injection
Mice were anesthetized using isoflurane (3% for induction and 1.5%–2% for maintenance) vaporized in oxygen. Intravitreal injections were performed using pulled glass micropipettes under a stereomicroscope as described previously.29^,^30 A total volume of 2 µL of viral vector was injected into the vitreous chamber of the right or left eye through the sclera just posterior to the limbus. Mice received intravitreal injections of AAV2-retro–cytomegalovirus (CMV)–histone 2B (H2B)–mGreenLantern (mGL), AAV2–CMV–H2B–mGL, or AAV–MNM008–CAGIG–H2B–V5–mGL (approximately 1.8 × 10^10^ vector genomes [vg]/eye). The original titers of these AAVs were AAV2-retro–CMV–H2B–mGL (3.46 × 10^13^ vg/mL), AAV2–CMV-H2B–mGL (0.9 × 10^13^ vg/mL), and AAV carrying a capsid variant MNM008 with a chicken β‑actin intron containing a splice donor (I) and splice acceptor (G) promoter (CAGIG)–H2B fused V5 tag (i.e., AAV–MNM008–CAGIG–H2B–V5–mGL) (7.82 × 10^13^ vg/mL). Buprenorphine SR was administered subcutaneously as postoperative analgesia.
For the postnatal injection study, intravitreal injections were performed at P3. Pups were anesthetized by hypothermia on ice, and viral vectors were delivered into the vitreous chamber using pulled glass micropipettes under a stereomicroscope. After injection, pups were allowed to recover on a warming pad before being returned to the dam.
AAV Vectors and Virus Production
AAVs were produced at the UTSW Ophthalmology Gene Editing and Viral Production Core or at the University of Miami Viral Vector Core. The reporter plasmid pAAV.CMV.H2B.mGL.WPRE contains AAV2 inverted terminal repeats (ITRs), a CMV enhancer/promoter, a chimeric intron, the mGL reporter fused to histone H2B, a human growth hormone (hGH) polyadenylation signal, and a woodchuck hepatitis virus posttranscriptional regulatory element (WPRE) element (Supplementary Fig. S1). AAVs were produced with two different capsids: AAV2-retro (rAAV2-retro helper was a gift from Alla Karpova and David Schaffer; Addgene plasmid #81070; RRID:Addgene_81070) and MNM008.11 The MNM008 capsid was constructed on the AAV2-retro backbone plasmid by substituting the sequence between N587 and R602 of the AAV2-retro VP1 capsid gene with the amino acid sequence SFTSPLHKNENTVS. The viruses were purified following previously described protocols,31 and viral titers were quantified using either quantitative polymerase chain reaction (qPCR) or digital PCR.
Tissue Processing
At designated time points (1 day, 3 days, and 2 weeks post-injections), mice were humanely euthanized in accordance with the American Veterinary Medical Association guidelines by CO_2_ inhalation followed by a confirmatory secondary method. When the absence of reflexes was confirmed, animals were transcardially perfused with ice-cold phosphate-buffered saline (PBS, 1×), followed by 4% paraformaldehyde (PFA) in PBS. Eyes were then enucleated and post-fixed in 4% PFA at room temperature (RT) for 2 to 3 hours before further processing. For flatmount retinal preparation, retinas were carefully dissected out from the eye cup and were washed three times in PBS. Four radial cuts were made at the ora serrata to create a cloverleaf shape before mounting. For cryosections, eyes were cryoprotected by immersion in 30% sucrose at 4°C overnight. Tissues were embedded in optimal cutting temperature (O.C.T.) compound using cryomolds and sectioned at a thickness of 10 µm on a Leica CM3050 S cryostat (Leica, Wetzlar, Germany).
Immunohistochemistry
To perform the immunofluorescent staining, samples were blocked with PBS containing 10% normal goat serum (NGS) and 0.3% Triton X-100 (0.3% phosphate-buffered saline with Triton x-100 [PBST]) for 1 hour at RT. Samples were then incubated with anti–RNA-binding protein with multiple splicing (RBPMS) antibody (1:2000; custom made at ProSci, Fort Collins, CO, USA), anti-cone arrestin (1:200; AB15282; Sigma-Aldrich, St. Louis, MO, USA), and anti–activator protein 2α (AP-2α) antibody (1:50; 3B5; Developmental Studies Hybridoma Bank, Iowa City, IA, USA) that were diluted in PBST blocking solution and incubated at 4°C overnight. After three washes (10 minutes each) in PBST, sections were incubated with anti-guinea pig Alexa Fluor 594 (1:500; 106-585-003; Jackson ImmunoResearch Laboratories, West Grove, PA, USA), anti-rabbit Alexa Fluor 647 (1:500; ab150079; Abcam, Cambridge, UK), and anti-mouse Alexa Fluor 647 (1:500; ab150115; Abcam) for 2 hours at RT. Samples were rinsed three times with PBST and then mounted onto a glass coverslip using Fluoromount-G Mounting Medium (Thermo Fisher Scientific, Waltham, MA, USA).
Imaging and Quantification
Retinal sections were imaged using a confocal microscope (Zeiss 780 Inverted). Images were acquired using 20× and 40× objectives. Sequential acquisition was performed to avoid spectral overlapping between fluorophores. Z-stacked images were acquired, and maximum intensity projections were generated using ImageJ Fiji (National Institutes of Health, Bethesda, MD, USA). Layer identification and qualitative assessments were performed based on 4′,6-diamidino-2-phenylindole (DAPI) staining and anatomical landmarks. For quantification, single-plane images were used, and maximum-intensity projections were generated for visualization. Cell quantification was performed on at least two fixed rectangular regions of interest (ROIs) measuring approximatively 59 × 198 µm, applied consistently across sections and animals. At least three animals were used for quantification. Data are presented as mean ± SEM and were analyzed using Prism (GraphPad, Boston, MA, USA). P < 0.05 was considered significant (*P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001).
Results
Intravitreal Injection of AAV2-Retro Leads to Rapid and Preferential Transduction of Photoreceptors and RPE Cells
To evaluate the transduction profile of AAV2-retro following intravitreal injections, we examined retinal tissue at 1, 3, and 14 dpi in adult male mice. At 1 dpi, the reporter mGL expression was already detectable, predominantly localized to the outer retina (Figs. 1A, 1B), suggesting early onset of transgene expression in photoreceptors and RPE cells. By 3 dpi, transduction intensity increased markedly, with a broader distribution across the photoreceptor layer and consistent labeling of RPE cells (Figs. 1C, 1D). Fluorescence signal intensity and cellular coverage were both elevated compared to the 1-day time point, indicating active viral entry and transcription. At 3 dpi, only a few green fluorescent protein (GFP)^+^ cells were seen in the ganglion cell layer (GCL) (Fig. 1D).
AAV2-retro preferentially transduces photoreceptors and RPE 1 and 3 days after intravitreal injection. (A, C) Stitched 10× images of retinal cross-sections showing mGL expression at 1 dpi (A) and at 3 dpi (C). Blue indicates DAPI-labeled nuclei; green indicates mGL reporter expression. (B, D) Higher-magnification images of representative retinal regions showing mGL expression at 1 dpi (B) and at 3 dpi (D). (B’, D’) Merged images corresponding to panels B and D, illustrating mGL and DAPI expression across retinal layers. Retinal layers are indicated: RPE, ONL, OPL, INL, inner plexiform layer (IPL), and GCL. Scale bars: 200 µm (A, C); 50 µm (B’, D’).
At 14 dpi, a robust GFP expression was observed throughout the photoreceptor layer, with strong labeling of RPE cells. (Figs. 2A, 2B) Minimal signal was observed in the GCL and the inner nuclear layer (INL) (Fig. 2B), confirming the tropism of AAV2-retro for outer retinal cell types under these conditions. These results demonstrate that intravitreal delivery of AAV2-retro in adult mice leads to rapid, preferential, and progressively enhanced transduction of photoreceptors and RPE cells within 2 weeks post-injection.
AAV2-retro maintains selective transduction in the outer layers 14 days after intravitreal injection. (A) Stitched 10× images of retinal cross-sections showing mGL expression at 14 dpi. (B) Higher-magnification view of a representative retinal region with mGL expression. (B’) Merged image from B highlighting layer-specific distribution (RPE, ONL, OPL, INL, IPL, and GCL), mGL, and DAPI across the layers. (C) Retinal flatmount at 14 dpi after sequential injections, showing widespread mGL expression. (C’, C’’) Higher magnification images of the boxed areas in C. Scale bars: 200 µm (A, C); 50 µm (B’, C’, C’’). (D) Quantification of mGL-positive cells within the regions of high transduction in the ONL (i.e., ROI). The percentage shown indicates the proportion of transduced cells among total DAPI-positive ONL cells. N = 3 animals. Scale bars: 200 µm (A, C); 50 µm (C’, C’’).
Normally, AAV-mediated transduction following a single injection is spatially concentrated, with robust expression near the presumed injection site. To assess whether sequential AAV2-retro delivery improves transduction efficiency in wider areas, adult mice received two intravitreal injections spaced 3 days apart. Retinal tissues were analyzed 14 days after the initial injection. Compared to a single injection, sequential dosing resulted in markedly increased reporter expression across the retina, with broader distribution and higher intensity in both photoreceptors and RPE cells (Fig. 2C). In some retinas, reporter expression extended to nearly the entire retina, and this widespread coverage was already observed by 3dpi (data not shown). In regions with strong mGL expression, over 90% of DAPI-stained cells in the photoreceptor layer were positive for mGL 14 days post-injection (Fig. 2D).
AAV2-Retro Exhibits Similar Photoreceptors and RPE Transduction in Female Mice
So far, we had used male mice. To assess whether the AAV2-retro transduction pattern observed in males was conserved across sex, intravitreal AAV2-retro injections were performed in adult female mice. Analysis of retinal cross-sections revealed robust reporter expression predominantly localized in the RPE and outer nuclear layer (ONL), with strong photoreceptor and RPE transduction. This ONL transduction pattern was evident at both 3 and 14 dpi and closely resembled the distribution observed in male retinas (Fig. 3). Minimal reporter expression was detected in the INL and GCL. These findings were consistently observed in three female animals, indicating that AAV2-retro–mediated transduction of RPE and outer retinal cell types is comparable between sexes under these conditions.
AAV2-retro leads to similar preferential transduction in female mice after intravitreal injection. (A, C) Stitched 10× images of retinal cross-sections showing mGL expression at 3 dpi (A) and at 14 dpi (C). Blue indicates DAPI-labeled nuclei; green indicates mGL reporter expression. (B, D) Higher magnification images of representative retinal regions showing mGL expression at 3 dpi (B) and at 14 dpi (D). (B’, D’) Merged images corresponding to panels B and D, illustrating mGL and DAPI expression across retinal layers. Retinal layers are indicated: RPE, ONL, OPL, INL, IPL, and GCL. Three female mice were examined per time point with similar results. Scale bars: 200 µm (A, C); 50 µm (B’, D’).
To rule out the possibility that the observed outer layer specificity resulted from unintended deep injections rather than true intravitreal delivery, we administered wild-type AAV2-mGL via intravitreal injection. This vector is well established to transduce cells primarily in the GCL under these conditions. Three days after intravitreal AAV2-mGL injection, mGL expression was detected primarily in the GCL, with no labeling detected in the inner or ONL (Fig. 4A). By day 14, transduction was markedly enhanced in the GCL, with dense and specific labeling of ganglion cells; expression remained absent in the outer retina, indicating that AAV2-mGL selectively and efficiently targets ganglion cells over time but does not reach the photoreceptors (Fig. 4A).
Distinct tropism is seen after intravitreal injection of AAV2–CMV-mGL and AAV2-MNM08-CMV-mGL. (A) Retinal cross-sections showing mGL expression at 3 dpi (top row) and 14 dpi (bottom row) following AAV2–CMV–mGL delivery. Blue indicates DAPI-labeled nuclei; green indicates mGL reporter expression. (B) Retinal cross-sections showing mGL expression at 3 dpi (top row) and 14 dpi (bottom row) following AAV2–MNM08–CMV–mGL delivery. The far-left panels show stitched 10× overviews. Retinal layers are indicated: RPE, ONL, OPL, INL, IPL, and GCL. The total number of animals examined was as follows: AAV2-mGL (3 dpi, n = 5; 14 dpi, n = 5) and AAV-MNM08 (3 dpi, n = 3; 14 dpi, n = 3). Scale bars: 200 µm (left panels); 50 µm (right panels).
Another Known Retrograde AAV Lacks Specificity for the Outer Retinal Layers
In addition to the original AAV2-retro,28 other engineered AAV capsids have been identified that exhibit efficient retrograde transport. Among these, MNM008 capsid, derived from canine adenovirus domains, showed exceptional retrograde infectivity for multiple neuronal types.11 To determine whether outer layer–specific gene delivery is a common feature among retrograde AAVs, we compared AAV-MNM008 with AAV2-retro following intravitreal injection. Mice received AAV-MNM008 expressing the mGL reporter, and retinal tissues were analyzed at 3 and 14 dpi. We observed that intravitreal injection of AAV-MNM008 resulted in a distinct transduction profile compared to that of AAV2-retro. At day 3 post-injection (Fig. 4B), mGL expression was detected in the photoreceptor layer, but with a lower signal intensity than what we observed with AAV2-retro. At 14 dpi, AAV-MNM008 mGL expression was persistent in the GCL and extended to the photoreceptor layer (PRL), although only a limited number of photoreceptor cells were transduced. (Fig. 4B). Together, these findings confirm that AAV2 robustly transduces the retinal ganglion cells (RGCs) without photoreceptors, whereas AAV2-MNM08 enables modest photoreceptor transduction with restricted efficiency and a limited spread. Neither vector matched the outer layer–specific transduction profile of AAV2-retro.
Identification of the Retinal Cell Types Transduced by the Vectors Using Cell Markers
To identify the retinal cell types transduced by AAV2-retro, we first performed immunostaining with an antibody against cone arrestin (a marker for cone photoreceptors). At 3 and 14 dpi, mGL expression colocalized with cone arrestin immunoreactivity, confirming efficient transduction of cones (Figs. 5A, 5B). In addition, mGL^+^ cells lacking cone arrestin immunoreactivity were also observed in the ONL, reflecting rod photoreceptor transduction (Figs. 5A, 5B). At both 3 and 14 dpi, a few scattered mGL^+^ cells were also detected in the GCL. To determine the identity of these cells, we performed immunohistochemistry with two cell markers: AP2-α, a marker for amacrine cells, and RBPMS, a marker for RGCs. Staining with AP2-α revealed that most transduced cells in the GCL were amacrine cells. (Figs. 6A, 6B). At 14 dpi, over 60% of mGL^+^ cells in the GCL and INL were immunoreactive for AP2-α. In contrast, fewer than 10% of mGL⁺ cells were immunoreactive for RBPMS (Fig. 6C). These results suggest that, in the inner retinal layers, where overall transduction was limited, AAV2-retro preferentially targeted amacrine cells, and this tropism persisted over time.
AAV2-retro effectively delivers transgene to both rods and cones. (A, B) Representative retinal cross-sections showing mGL expression (green) at 3 dpi (A) and at 14 dpi (B) following intravitreal injection of AAV2-retro–CMV–mGL. Cone arrestin (magenta) was used to identify cone photoreceptors. Bottom panel in A shows higher magnification images highlighting the colocalization of mGL with cone arrestin immunoreactivity. Scale bars: 50 µm (A, B); 5 µm (higher magnification images).
AAV2-retro transduces only a small number of amacrine cells and RGCs. (A, B) Retinal cross-sections show mGL expression (green) at 3 dpi (A) and at 14 dpi (B). AP-2α immunoreactivity is shown in magenta and RBPMS immunoreactivity in red. (A’, B’) Higher magnification images showing cells that appear to be co-labeled with mGL and AP-2α. (C) Quantification shows that the percentage of mGL+ cells that were positive for either RBPMS (in GCL and INL) or AP-2α at each time point. N = 3 per group. Error bar, ± SEM Two-way ANOVA with Tukey's multiple comparisons: * < 0.05, **** < 0.0001, ns = not significant. Scale bars: 50 µm (A, B); 5 µm (A’, B’).
AAV2-Retro Transduces Cells in the RPE and Outer Neuroblastic Layer of Postnatal Mouse Retina
To assess whether AAV2-retro can transduce retinal cells during early postnatal development, intravitreal injections were performed on P3 mice, and retinas were analyzed 3 days later. Reporter expression was detected predominantly within the RPE and the outer neuroblastic layer (oNBL) (Fig. 7). At this developmental stage, rods and cones are already present; however, their maturation is not yet complete, and they progressively contribute to formation of the ONL. Immunostaining with a cone arrestin antibody showed overlap with reporter expressing cells within the oNBL,32^,^33 indicating transduction of cone lineage cells during postnatal development (Fig. 7).
Early postnatal intravitreal delivery of AAV-retro results in viral transduction of RPE cells and cells within the outer neuroblastic layer of the retina. (A) Higher magnification view of a retinal region from a P3-injected pup showing mGL+ cells expressed in the oNBL, consistent with photoreceptor localization. (A’) Higher magnification view of the same region showing mGL and DAPI. (B) Immunostaining for cone arrestin (magenta) shows colocalization of mGL and arrestin signals, indicating early cone transduction when the AAV was injected at P3. (B’) Higher magnification images highlight cells co-labeled with mGL and cone arrestin. Retinal layers are indicated: RPE, oNBL, inner neuroblastic layer (iNBL), and GCL. AAV was injected intravitreally in four mice with similar results. Scale bars: 50 µm (B); 5 µm (B’).
Discussion
Our results demonstrate that AAV2-retro rapidly and robustly transduces the outer retina, with reporter expression detectable as early as 1 dpi and progressively increasing by 3 and 14 dpi. We also observed transduction of a few cells in the GCL, where the majority co-expressed the AP-2α, consistent with the identity of amacrine cells. The pronounced specificity of AAV2-retro for the outer retinal layers is striking, given that intravitreal delivery has traditionally been considered inadequate for targeting these regions, with most vectors preferentially transducing cells in the INL and GCL.34^,^35 Other engineered AAV capsids, including AAV2.7m8, AAV2.GL, and AAV2.NN, have demonstrated markedly improved retinal transduction efficiency compared with wild-type AAV2.24^,^36 However, these variants generally enhance transduction across multiple retinal layers rather than conferring specificity to the outer retina. Consequently, photoreceptor-restricted gene expression in these systems has relied on the use of rod-specific promoters, such as the rhodopsin (RHO) promoter.24 In contrast, the ability of AAV2-retro to overcome these limitations underscores its unique efficiency and distinguishes it from other tested capsids. The absence of a similar expression pattern in MNM08 further underscores the unique characteristics of AAV2-retro.
The specificity and rapid onset of AAV2-retro provide distinct advantages for preclinical animal studies of retinal degeneration. When delivered intravitreally, AAV2-retro efficiently and selectively transduces photoreceptors and RPE within a brief period, eliminating the need for invasive subretinal injection. This property allows for more consistent and reproducible gene delivery across the retina and reduces surgical variability and tissue damage. The rapid onset of transgene expression further facilitates early evaluation of gene function and therapeutic efficacy, which is a key advantage in disease models characterized by fast progressing degeneration. Taken together, these attributes make AAV2-retro a potent tool for accelerating preclinical testing of gene therapies targeting photoreceptor and RPE dysfunction. Importantly, confining gene manipulation to photoreceptors and RPE minimizes off-target effects from transgene expression in inner retinal neurons or glial cells, which could otherwise perturb retinal circuitry and impair visual function. Gene expression in non-targeted retinal cell types can alter synaptic signaling, disrupt homeostatic support functions, or induce unintended stress responses, ultimately confounding the interpretation of experimental outcomes. By confining transgene delivery to the outer retina, AAV2-retro enables more precise assessment of gene function within disease-relevant cell populations while preserving the physiological integrity of the inner retinal network. Thus, the unique tropism of AAV2-retro raises opportunities for studying outer retinal physiology in vivo. By restricting transgene expression to photoreceptors and RPE, researchers can manipulate signaling pathways, visual cycle components, or metabolic processes without disrupting inner retinal neuronal networks, enabling clearer mechanistic insights. Although single intravitreal injections resulted in strong photoreceptor and RPE transduction, retinal coverage was not uniform; second intravitreal injections further enhanced both the efficiency and spatial coverage of transduction, suggesting that sequential injections can be used to overcome local variability in viral entry and improve gene delivery outcomes.
In this study, we also demonstrated that AAV2-retro exhibits similarly robust tropism for photoreceptors and RPE in female mice, paralleling the strong and selective outer retinal transduction observed in males. The consistency of vector performance across sexes is an important finding, as several retinal degenerative diseases, including certain forms of age-related macular degeneration, autoimmune retinopathies, and mitochondrial optic neuropathies, show higher prevalence, faster progression, or distinct clinical manifestations in females. The ability of AAV2-retro to reliably target the outer retina in female animals underscores its utility as a gene-delivery tool for modeling and potentially treating these sex-skewed conditions.
We further showed that AAV2-retro delivered intravitreally at P3 can efficiently transduce the RPE and the oNBL, including cone lineage cells, during a critical window of early retinal development. Many inherited retinal degenerations such as Leber congenital amaurosis and early-onset retinitis pigmentosa may cause photoreceptor dysfunction or death beginning in early postnatal or even prenatal stages. Thus, the ability of AAV2-retro to access and transduce photoreceptor precursors at P3 suggests that this vector may be particularly valuable in animal models where therapeutic intervention must occur before maturation of the ONL. By providing efficient early developmental gene delivery via a simple intravitreal injection, AAV2-retro offers a practical tool for probing disease pathogenesis and evaluating therapeutic rescue in inherited retinal disorder (IRD) models where early photoreceptor targeting is essential.
The mechanism underlying the selective tropism of AAV2-retro for outer retinal cells remains unclear. One possibility is that its capsid structure facilitates enhanced penetration through the ILM, a barrier that has historically limited the efficacy of intravitreal gene transfer.37 Alternatively, AAV2-retro may possess capsid motifs that preferentially interact with cell-surface receptors expressed on photoreceptors or RPE, enabling rapid internalization and transgene expression. Given the original design of the vector for retrograde transport in central nervous system neurons,28^,^38 its adaptation to the retinal environment may involve unexpected cross-compatibility with outer retinal cellular entry pathways.
Despite the promise of AAV2-retro, several limitations warrant consideration. Although our results demonstrate rapid and efficient transgene expression, the long-term stability of this transduction remains to be determined. Comprehensive evaluation of expression durability, potential immune responses, and the safety of repeated dosing in larger cohorts may be needed. In a previous study, several AAV capsids, including AAV2-retro, were evaluated for their ability to transduce cells in the human retina ex vivo. In this experiment, human retinal explants were cultured in Transwells with the photoreceptor layer facing downward, and AAVs were applied from the vitreal side (ILM/RGC layer) to mimic intravitreal injection conditions. Under these conditions, AAV2-retro produced only limited transduction of the human retina.39 Although ex vivo and in vivo environments differ substantially, these findings suggest that the robust outer retinal transduction observed in mouse eyes in the present study may not directly translate to other species.
Conclusions
Our work has established AAV2-retro as a markedly efficient virus for selectively targeting photoreceptors and RPE following intravitreal injection. Its rapid onset of expression, superior efficiency compared with established serotypes, and enhanced distribution with sequential dosing underscore its potential for experimental applications.
Supplementary Material
Supplement 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kerschensteiner D. Losing, preserving, and restoring vision from neurodegeneration in the eye. Curr Biol. 2023; 33(19): R 1019–R 1036.37816323 10.1016/j.cub.2023.08.044PMC 10575673 · doi ↗ · pubmed ↗
- 2Sanie-Jahromi F, Nowroozzadeh MH. RPE based gene and cell therapy for inherited retinal diseases: a review. Exp Eye Res. 2022; 217: 108961.35092717 10.1016/j.exer.2022.108961 · doi ↗ · pubmed ↗
- 3Azam M, Jastrzebska B. Mechanisms of rhodopsin-related inherited retinal degeneration and pharmacological treatment strategies. Cells. 2025; 14(1): 49.39791750 10.3390/cells 14010049 PMC 11720364 · doi ↗ · pubmed ↗
- 4Russell S, Bennett J, Wellman JA, et al. Efficacy and safety of voretigene neparvovec (AAV 2-h RPE 65v 2) in patients with RPE 65-mediated inherited retinal dystrophy: a randomised, controlled, open-label, phase 3 trial. Lancet. 2017; 390(10097): 849–860.28712537 10.1016/S 0140-6736(17)31868-8PMC 5726391 · doi ↗ · pubmed ↗
- 5Nuzbrokh Y, Ragi SD, Tsang SH. Gene therapy for inherited retinal diseases. Ann Transl Med. 2021; 9(15): 1278.34532415 10.21037/atm-20-4726 PMC 8421966 · doi ↗ · pubmed ↗
- 6Lonfat N, Moreno-Leon L, Punzo C, Khanna H. Update on gene therapy clinical trials for eye diseases. Hum Gene Ther. 2025; 36(19-20): 1287–1300.40965251 10.1177/10430342251379824 PMC 13089423 · doi ↗ · pubmed ↗
- 7Smith AJ, Schlichtenbrede FC, Tschernutter M, Bainbridge JW, Thrasher AJ, Ali RR. AAV-mediated gene transfer slows photoreceptor loss in the RCS rat model of retinitis pigmentosa. Mol Ther. 2003; 8(2): 188–195.12907141 10.1016/s 1525-0016(03)00144-8 · doi ↗ · pubmed ↗
- 8Wang D, Gao G. State-of-the-art human gene therapy: part I. Gene delivery technologies. Discov Med. 2014; 18(97): 67–77.25091489 PMC 4440413 · pubmed ↗