Molecular weight-based profiling of sperm proteins and their association with semen quality in Kokok Balenggek roosters
Jaswandi Jaswandi, Harif Gusdinal, Tulus Maulana, Ekayanti Mulyawati Kaiin, Rusfidra Rusfidra, Ananda Ananda

TL;DR
This study explores how the molecular weight of sperm proteins in Kokok Balenggek roosters relates to semen quality, identifying potential markers for fertility.
Contribution
The study introduces molecular weight-based profiling of sperm proteins as a novel approach to assess semen quality in Kokok Balenggek roosters.
Findings
SDS-PAGE identified 46 sperm protein bands with molecular weights from 8 to ≥245 kDa in Kokok Balenggek roosters.
High molecular weight proteins (≥218–247 kDa) negatively correlate with sperm motility and viability.
Low molecular weight proteins (<30–40 kDa) show positive associations with sperm quality parameters.
Abstract
The study characterized the sperm proteins of Kokok Balenggek roosters based on molecular weight patterns to investigate the relationship between sperm composition and semen quality. Semen from 15 Kokok Balenggek roosters was collected and tested for standard quality indicators, including volume, motility, viability, abnormalities, and plasma membrane integrity (PMI). The protein content was measured, and the sperm protein profile was established by applying SDS-PAGE on a 10% polyacrylamide gel. In the next phase, the statistical correlation between the indicators of semen quality and the total protein levels, as well as the protein fractions defined by the molecular weight, was determined. Through the SDS-PAGE analysis, a total of 46 protein bands with molecular weights ranging from 8 to ≥ 245 kDa were identified in KBR semen. The average volume of fresh semen was 597.67 ± 325.20 µl,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
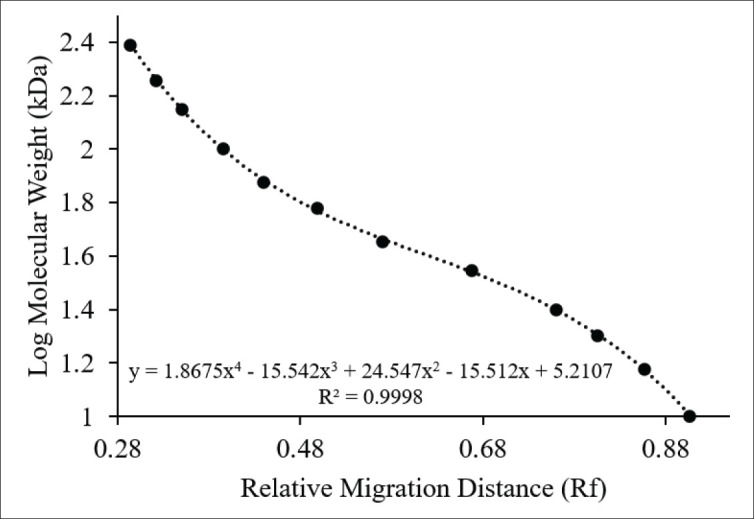 Figure 1
Figure 1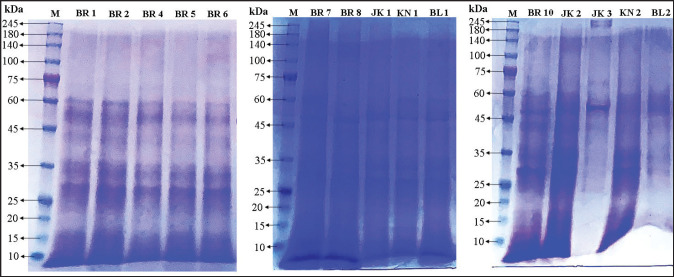 Figure 2
Figure 2| Semen evaluation parameters | Mean |
|---|---|
| Macroscopic characteristics | |
| Volume (µl) | 597.67 ± 325.20 |
| Odor | Rooster-specific |
| pH | 7.11 ± 0.24 |
| Color | Milky White |
| Consistency | Thick |
| Microscopic characteristics | |
| Mass Movement | ++ |
| Motility (%) | 80.78 ± 12.92 |
| Concentration (10⁹/ml) | 1.99 ± 1.14 |
| Viability (%) | 87.56 ± 5.33 |
| Abnormality (%) | 10.60 ± 2.30 |
| Plasma Membrane Integrity (%) | 85.06 ± 5.37 |
| Rooster ID | Sperm Motility (%) | Sperm Viability (%) | Sperm Abnormality (%) | Sperm PMI (%) | Sperm PC (mg/ml) |
|---|---|---|---|---|---|
| BR1 | 90.00 ± 0.00a | 92.67 ± 0.76a | 11.33 ± 0.76a | 89.67 ± 0.29a | 0.22 |
| BR2 | 85.00 ± 5.00abc | 89.00 ± 1.00bcd | 9.00 ± 1.00a | 87.33 ± 1.26bc | 0.57 |
| BR4 | 85.00 ± 0.00abc | 85.67 ± 1.04e | 10.50 ± 2.50a | 83.33 ± 0.29e | 0.81 |
| BR5 | 86.67 ± 2.89ab | 88.67 ± 0.29bcd | 10.67 ± 1.76a | 85.33 ± 0.29cde | 0.99 |
| BR6 | 71.67 ± 2.89d | 88.50 ± 2.00bcd | 9.00 ± 2.00a | 86.33 ± 1.76bcd | 0.25 |
| BR7 | 86.67 ± 2.89ab | 90.67 ± 1.26b | 11.33 ± 2.25a | 88.33 ± 1.26ab | 0.72 |
| BR8 | 81.67 ± 2.89bc | 88.00 ± 0.50cd | 10.33 ± 1.26a | 86.00 ± 0.50cd | 0.71 |
| JK1 | 80.00 ± 0.00c | 87.50 ± 1.50de | 10.50 ± 3.00a | 84.17 ± 1.26de | 0.72 |
| KN1 | 81.67 ± 2.89bc | 88.33 ± 1.76cd | 10.33 ± 0.76a | 86.33 ± 1.26bcd | 0.69 |
| BL1 | 86.67 ± 2.89ab | 89.50 ± 0.50bcd | 9.67 ± 2.75a | 87.00 ± 1.00bc | 0.72 |
| BR10 | 81.67 ± 2.89bc | 88.00 ± 1.00cd | 12.00 ± 1.00a | 86.00 ± 1.50cd | 0.66 |
| JK2 | 86.67 ± 2.89ab | 90.00 ± 0.50bc | 9.33 ± 0.76a | 86.50 ± 1.00bc | 0.66 |
| JK3 | 36.67 ± 5.77e | 69.00 ± 1.50f | 16.33 ± 1.76b | 66.33 ± 1.76f | 0.70 |
| KN2 | 86.67 ± 2.89ab | 88.17 ± 1.76cd | 9.33 ± 1.26a | 85.67 ± 1.76cd | 0.72 |
| BL2 | 85.00 ± 5.00abc | 89.67 ± 0.29bcd | 9.33 ± 0.76a | 87.50 ± 1.00bc | 0.73 |
| Mean ± SD | 80.78 ± 12.92 | 87.56 ± 5.33 | 10.60 ± 2.30 | 85.06 ± 5.37 | 0.66 ± 0.19 |
| MW (kDa) | Rooster ID | Protein Absence* | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BR1 | BR2 | BR4 | BR5 | BR6 | BR7 | BR8 | JK1 | KN1 | BL1 | BR10 | JK2 | JK3 | KN2 | BL2 | ||
| ≥ 245 | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − | 1/15 (6,7) |
| 244–180 | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − | 1/15 (7,7) |
| 179–140 | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 0/15 (0) |
| 139–100 | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 0/15 (0) |
| 99–75 | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 0/15 (0) |
| 74–60 | + | − | − | − | − | + | − | − | − | − | − | − | − | − | − | 2/15 (13,3) |
| 59–45 | + | + | + | + | + | + | + | + | + | + | + | + | + | − | + | 14/15 (93,3) |
| 44–35 | + | − | − | + | − | − | − | − | − | − | + | + | − | − | − | 4/15 (26,7) |
| 34–25 | + | + | + | + | + | + | − | + | + | + | + | − | − | + | − | 11/15 (73,3) |
| 24–20 | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 0/15 (0) |
| 19–15 | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 0/15 (0) |
| 14–10 | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | 1/15 (6,7) |
| ≤ 9 | + | + | + | + | + | + | + | + | + | + | + | + | − | − | − | 12/15 (80) |
| Σ Bands | 5 | 3 | 3 | 4 | 3 | 3 | 4 | 2 | 3 | 3 | 4 | 3 | 3 | 2 | 1 | 46/195 (0,3) |
| Parameter / Protein MW (kDa) | Motility ( | Viability ( | Abnormality ( | PMI ( |
|---|---|---|---|---|
| Protein concentration (mg/ml) | 0.042, 0.881 | −0.199, 0.478 | 0.103, 0.715 | −0.213, 0.446 |
| Protein bands ( | 0.091, 0.746 | 0.133, 0.636 | 0.314, 0.254 | 0.100, 0.723 |
| 247 kDa | −0.943**, 0.000 | −0.956**, 0.000 | 0.867**, 0.000 | −0.958**, 0.000 |
| 218 kDa | −0.943**, 0.000 | −0.956**, 0.000 | 0.867**, 0.000 | −0.958**, 0.000 |
| 61 kDa | 0.237, 0.394 | 0.311, 0.259 | 0.162, 0.563 | 0.297, 0.283 |
| 54 kDa | −0.075, 0.789 | 0.018, 0.949 | 0.144, 0.609 | 0.024, 0.932 |
| 42 kDa | 0.263, 0.344 | 0.264, 0.341 | 0.079, 0.779 | 0.210, 0.452 |
| 31 kDa | 0.403, 0.136 | 0.399, 0.141 | −0.229, 0.412 | 0.408, 0.132 |
| 11 kDa | 0.126, 0.655 | 0.032, 0.911 | −0.192, 0.493 | 0.031, 0.911 |
| 8 kDa | 0.435, 0.105 | 0.490, 0.063 | −0.326, 0.236 | 0.487, 0.066 |
| Molecular weight (kDa) | Candidate protein | Reported function | Species studied | References |
|---|---|---|---|---|
| ~28–32 | BSP proteins (Binder of Sperm, e.g., BSP-A1/A2, BSP-A3) | Capacitation, sperm storage, sperm–egg binding | Bull, boar, suggested in poultry | [ |
| ~40–42 | Glycoproteins (e.g., sperm–egg binding proteins) | Sperm activation, perivitelline layer penetration | Chicken, quail | [ |
| ~60–75 | HSP70/HSP90 variants | Stress response, protein folding, sperm protection | Bull, chicken, human | [ |
| ~72 | SPAG6 (Sperm-associated antigen 6) | Flagellar structure and function, motility regulation | Rooster, human | [ |
| ~200–250 | Fibronectin, large HSPs | Structural support, sperm–oviduct interaction, capacitation | Chicken, mammal | [ |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSperm and Testicular Function · Reproductive Biology and Fertility · Animal Genetics and Reproduction
Introduction
In poultry, the quality of semen and the efficiency of reproduction are the two primary factors that directly influence the breeding outcomes, such as fertility, productivity, and the preservation of genetic diversity, among others. For male chickens, evaluation of semen typically involves assessing plasma membrane integrity (PMI), motility, morphological abnormalities, and viability, all of which are closely related to the fertilization potential of females [1]. Of the semen quality parameters, sperm motility is the most important, as it indicates the sperm’s ability to reach the oocyte within the female reproductive tract. It is essential to keep the sperm alive and, at the same time, maintain the membrane integrity if the fertilization potential of the functional spermatozoa is to be preserved. On the other hand, changes in morphology may severely hinder reproductive performance [2–4].
The use of artificial insemination in studies with Kokok Balenggek roosters (KBRs) has revealed that semen quality is the primary characteristic responsible for determining both fertility and hatchability[5,6].
In the primary semen assays, which are predominant, still considerable disadvantages that could lead to unfavorable results. Using a microscope to assess the movement of sperm, their concentration, and their shape provides the researcher with a basic description of the condition; however, this alone is not enough to give a complete picture of the sperm's functional capacity. As a result, some males of bird species, such as chickens, which exhibit normal semen characteristics, may still be sterile due to undetected molecular abnormalities during routine analyses [7,8].
Furthermore, the use of manual microscopy may lead to subjective interpretation and observer variability, resulting in the dullness of both reproducibility and predictive reliability in semen assessments[9,10]. The use of technologies such as computer-assisted sperm analysis (CASA), flow cytometry, and fluorescence microscopy, among others, has significantly standardized sperm analysis[11. However, CASA still primarily focuses on evaluating kinematic traits and clarifying the mechanistic basis of sperm function, although it is costly and less accessible in low-resource settings, and remains restricted. Sperm dysfunction has been increasingly attributed to oxidative stress, which serves as a critical mediator influencing motility, viability, and fertilization ability[12–15]. This trend calls for molecular methods, which not only provide conventional assessments but also capture the underlying biological processes beyond these.
One of the factors that affects the variation in semen quality is its regulation by hormones. The correlation between fecal testosterone levels and semen parameters has been observed in different KBR phenotypes, supporting this finding [16]. To clarify the role of testosterone, it first supports the process of spermatogenesis, and secondly, it impacts the production of some sperm-associated proteins related to energy and capacitation. Antioxidant supplementation studies in avian semen provide strong evidence for the interplay of hormones, redox balance, and protein expression, especially when oxidative stress prevails [17,18].
Proteomic analysis enables the identification of proteins that are indispensable for essential reproductive functions, such as fertilization, motility, the acrosome reaction, and capacitation. The proteins identified as potential fertility biomarkers include proAKAP4, phosphatidylethanolamine-binding protein 4, mitochondrial enzymes, and heat shock proteins, which are significant in both avian and mammalian species [19–21]. Sperm proteins that exhibit different amounts in high-fertility and low-fertility males can serve as effective molecular markers in breeding programs for livestock species [22,23]. A recent study conducted by us defined sperm proteins and seminal plasma in relation to different Kokok Balenggek rooster phenotypes, revealing a significant diversity in protein expression within this local breed [24]. However, the protein molecular weights and their quality traits in KBRs have not been studied yet.
The technique of SDS-PAGE, which is a combination of sodium dodecyl sulfate and polyacrylamide gel electrophoresis, allows for the separation of proteins based on their molecular weight, which is a simple and cheap method that helps researchers to acknowledge the distinct color bands of the proteins that might indicate the quality of the semen [24–26].
Mass spectrometry, a resolution technique, is still considered a high-resolution method for the precise identification of species with little prior research; nevertheless, SDS-PAGE is still recognized as a worthy method for initial protein analysis 27]. In the case of birds, the sperm protein profiles linked to fertility traits have been revealed through the use of SDS-PAGE, thus providing users with insights that extend beyond traditional semen evaluation. The native chicken breeds are not only significant from a genetic perspective, but they also provide a solid foundation for sustainable poultry production in developing countries, primarily due to their ability to adapt, resistance to diseases, and socio-economic importance [28,29].
On the other hand, the distribution of commercial breeds and crossbreeding poses a threat to their genetic diversity and may lead to genetic erosion [30]. Conservation measures need to be implemented, and an extensive characterization of native breeds should be conducted to protect biodiversity and preserve traits that can be utilized for future breeding strategies. In Indonesia, the Kokok Balenggek rooster is distinguished by its unique crowing, adaptability, and reproductive power [31,32].
Research on KBR sperm has provided information about its quality parameters and storage conditions in various diluents, indicating that it is a suitable candidate for artificial insemination applications [33]. However, there has been limited molecular data available on the sperm protein composition.
The researchers of this study proposed that there is a connection between certain molecular weights of sperm proteins and quality traits of semen in KBRs. The research focused on characterizing sperm proteins through SDS-PAGE and their relationship to semen parameters. Thus, new molecular insights into the reproductive biology of roosters were gained, and at the same time, the conservation of native chicken breeds and their genetic improvement were supported.
Materials and Methods
Ethical approval
The Research Ethics Committee of the Faculty of Medicine, Universitas Andalas, Indonesia, reviewed and approved all procedures involving animals conducted in this study (Approval No: 473/UN.16.2/KEP-FK/2024) approval date: 3 December 2024.
Experimental animals and management
This study utilized 15 KBR (Gallus gallus domesticus) aged 2–3 years, with an average body weight of 1.4 ± 0.3 kg. All roosters were healthy and free from any physical defects. Each bird was housed individually in a battery cage (45 × 50 × 60 cm) at the Poultry Unit, Universitas Andalas, Padang, Indonesia. The birds were fed a commercial breeder feed (ABS Crumble, JAPFA Comfeed Indonesia) supplemented with vitamins in the drinking water, provided ad libitum. Feeders and drinkers were maintained and cleaned daily, ensuring constant access to fresh water.
Semen collection and evaluation
The semen of each rooster was collected three times at 3-day intervals using the dorso-abdominal massage method [34]; thus, a total of 45 ejaculates was produced. Collected ejaculates were placed in sterile microtubes, labeled, and analyzed for macroscopic traits (consistency, volume, pH, color, and odor) as well as microscopic traits, which included PMI, mass movement, concentration, abnormality, motility, and viability, according to established protocols [35–38].
Sperm motility was quantified as the percentage of progressively motile sperm using a light microscope (Axiocam HRc, Carl Zeiss, Germany) at 400x magnification, while mass movement was scored from—(absent) to +++ (excellent). After semen dilution in 3% NaCl, sperm concentration was determined using a hemocytometer. Viability and abnormal morphology were examined through eosin–nigrosin staining, and PMI of 200 spermatozoa was assessed via the hypo-osmotic swelling test under phase-contrast microscopy.
Protein extraction from sperm cells
Ejaculates with motility of 70% or more were subjected to centrifugation after three consecutive washes with (phosphate-buffered saline) at 4°C to separate sperm cells from seminal plasma. The sperm pellets from each collection were stored at –20°C until needed. At the end of the collection period, three frozen sperm pellets from each rooster were thawed and combined to create a sample that represents each individual, thereby yielding 15 final sperm samples for protein analysis. Sperm pellets were preserved using the freezing method at −20°C without any cryoprotectants until pooling, simulating the typical conditions of semen storage prior to artificial insemination research processing, while also acknowledging that freezing-thawing could affect certain membrane proteins. Using PRO-PREP™ Protein Extraction Solution (iNtRON Biotechnology, Seoul, Korea), samples were resuspended, homogenized, and incubated at –20°C for 20 min, followed by centrifugation at 13,000 rpm for 5 min at 4°C; the supernatant enriched with extracted proteins was harvested and preserved at –20°C until further use [37,38].
Protein quantification
Using a spectrophotometer (UV-1800, Shimadzu, Kyoto, Japan), absorbance at 595 nm was measured after determining the total protein concentration via the Bradford assay, with bovine serum albumin (BSA; Sigma-Aldrich, St. Louis, MO, USA) used as the reference standard [35].
SDS-PAGE and molecular weight estimation
Using SDS-PAGE, proteins extracted from spermatozoa were resolved on precast 10% polyacrylamide gels (Q-PAGE™, SMOBIO, Taiwan). The 10% gel concentration was selected because it provides reliable resolution for proteins in the 10–245 kDa range, which was the primary focus of this study. Equal amounts of protein (approximately 25 µg in 20 µl per lane) were loaded to ensure comparable band intensity across samples, based on normalization of protein concentrations obtained from the Bradford assay. After denaturing the sample proteins at 70°C for 10 min in NuPAGE™ LDS Sample Buffer (Thermo Fisher Scientific, Waltham, MA, USA), they were loaded into gel wells along with a prestained molecular weight marker (ExcelBand™, SMOBIO, Hsinchu, Taiwan). Electrophoresis was carried out for 90 min at 110 V using a BIO-RAD vertical system (Hercules, CA, USA). Following overnight staining with Fast Coomassie Blue (Elabscience, Wuhan, China) and rinsing in distilled water, the gels were visualized using the iBright 1,500 imaging system (Thermo Fisher Scientific, Waltham, MA, USA). The relative migration of each protein band was quantified using ImageJ (NIH, Bethesda, MD, USA) and compared to the molecular weight standards. Protein molecular weights were subsequently calculated with a polynomial regression formula:
y = 1.8675x⁴ − 15.542x³ + 24.547x² − 15.512x + 5.2107; R² = 0.9998
where y corresponds to log molecular weight (kDa) and x represents the measured Rf. The regression curve was generated specifically from the marker bands (Fig. 1).
Standard calibration curve for SDS-PAGE. Protein markers (PM2700, SMOBIO, Taiwan) were used to correlate the relative migration distance (Rf) with the logarithm of molecular weight (kDa). Using ImageJ software, the molecular weights of the sperm protein bands in KBr were calculated based on a polynomial regression equation (R² = 0.9998), confirming the high precision of the estimation.
Data analysis
The various traits of semen quality were initially summarized with descriptive statistics defined as mean ± SD. To detect significant differences between single roosters, a one-way ANOVA was performed, followed by Duncan's multiple range test to separate means where significant differences were found (p < 0.05). A Pearson's correlation test, conducted using SPSS version 25.0 (IBM Corp., Armonk, NY, USA), was utilized to investigate the relationships among semen quality factors, protein concentration, and protein bands of different molecular weights. The consideration of effect sizes was also made to enhance the statistical interpretation [38].
Results and Discussion
Quality of fresh semen from Kokok Balenggek roosters
The average semen volume of KBR was 597.67 ± 325.20 µl, as shown in Table 1, which presents the characteristics of fresh semen. This value exhibits high variability, with the standard deviation accounting for almost 54% of the mean. This variability indicates the possibility of outliers or individual physiological differences among roosters; if not using statistical methods such as outlier diagnostics, this may skew the interpretation of correlations. However, the values observed were within the wide range reported in other indigenous breeds, such as 0.05–1.0 ml in Castellana chickens [39], 0.22–0.36 ml in Thai natives [40], and ~0.6 ml in Tanzanian Horasi ecotypes. The semen was characterized by a neutral to slightly alkaline environment, having a mean pH of 7.11 ± 0.24, which was considered optimal for sperm viability. The averages of the motility, viability, and PMI were 80.78% ± 12.92%, 87.56% ± 5.33%, and 85.06% ± 5.37%, respectively, while the sperm abnormality was 10.60% ± 2.30%, which is quite low. These characteristics exceed the threshold for artificial insemination, where a motility of 70% is required for acceptable fertility [41]. In Kedu, Sentul, and Kampung chickens, similar values have been reported with motility of 75%–85% and viability of nearly 90% [42,43].
The variation among individuals (Table 2) is anticipated in native breeds, wherein genetic diversity and environmental adaptation promote phenotypic variation [44]. Climatic and environmental factors are of great importance, as high temperatures and humidity reduce motility and sperm concentration in tropical birds 45]. These factors underscore the need for multiple sampling points rather than single-point sampling in semen studies. Moreover, the quality of the semen must be assessed in conjunction with the management practices: over-restraining, inefficient collection techniques, or stress during handling may reduce the quality of the ejaculate without affecting the inherent sperm characteristics. An increase in corticosterone due to stress has been demonstrated to affect testicular function and sperm physiology in birds and mammals, indicating the need to consider behavioral and management factors when determining semen quality.
Protein concentration and SDS-PAGE profiles of sperm proteins
Table 2 illustrates that the concentration of sperm protein in Kokok Balenggek male chickens varied between 0.22 and 0.99 mg/ml, with an average value of 0.66 ± 0.19 mg/ml. The nearly a five-fold difference in concentrations among males indicates a significant physiological variation among the birds. Such variations might be due to differences in testicular activity, spermatogenic efficiency, or sperm protein turnover. For instance, the intake of dietary antioxidants, the burden of oxidative stress, and the age of males have been associated with the level of sperm protein in poultry and livestock [14,23].
Another factor that might influence sperm protein levels is the season. Chickens subjected to heat stress develop a different seminal protein profile, with an increased amount of chaperones and stress proteins [45]. Hence, such variations in sperm, indicated by changes in protein concentration, are considered biologically acceptable, and at the same time, they provide insight into the complexity of regulating the sperm proteome.
During the SDS-PAGE analysis, a total of 46 protein bands were detected with an approximate weight ranging from 10 kDa to 245 kDa, as depicted in Table 3 and Figure 2.
SDS-PAGE analysis of sperm protein profiles from Kokok Balenggek roosters. A 10% polyacrylamide gel was used to separate the protein bands. Lanes BR1–BL2 corresponded to individual Kokok Balenggek roosters, and PM2700 (SMOBIO, Taiwan) served as the molecular weight reference in the leftmost lane, ranging from 10 to 245 kDa. Gels were documented using the iBright imaging system (Thermo Fisher Scientific).
The main groups that were expressed the most consistently were from the middle weight ranges: 59–45 kDa (93.3% of the samples), 44–35 kDa (73.3%), and 34–25 kDa (73.3%). The mentioned ranges have been previously associated with sperm motility and fertilization in birds and mammals [26,46].
The low-molecular-weight proteins (~28–32 kDa) are probably those that belong to BSPhomologs, which are responsible for sperm-egg binding and capacitation [4], while the proteins in ~40–42 kDa are those glycoproteins that help in perivitelline penetration [48]. The proteins at the intermediate level (~60–75 kDa) are composed of HSP70/90 chaperones and SPAG6, with the former being the dominant ones in the flagellum's operation and the latter playing a role in resistance to stress [49]. Fibronectin and large heat shock proteins, for example, high-molecular-weight proteins exceeding 200 kDa, may be indicative of structural or stress-related processes; however, their high levels have been negatively associated with semen quality in several species [50]. The existence of the proteins in the sample, however, is still uncertain. SDS-PAGE simply gives a separation based on the approximate molecular weight, and without densitometry, Western blotting, or mass spectrometry, the identities of the candidate proteins cannot be confirmed. Therefore, the correlations stated here must be viewed as suggestive rather than conclusive. Rooster fertility will be a topic for which the specific proteins involved and their functional roles will be validated by future studies using LC-MS /MS or targeted immunodetection.
Correlations between protein profiles and semen quality traits
A correlation analysis conducted on the dataset from the experiment yielded three key findings (Table 4. First, a moderate correlation was observed between protein concentration and motility (r = 0.42); meanwhile, the correlations to viability (r = −0.199), PMI (r = −0.213), and abnormality (r = 0.103) were all of a weak nature. It follows that the total protein content supports motility, perhaps through energy metabolism, but does not have a direct impact on cell survival or morphology. In fact, motility is dependent on and supported by glycolytic and oxidative phosphorylation pathways that are activated by adenosine triphosphate (ATP) synthase, GAPDH, and other metabolic proteins [23,51]. On the other hand, viability is predominantly influenced by the level of oxidative stress, age, and environmental conditions [4,45]. The results suggest that it is crucial to exercise caution in not overemphasizing protein concentration as a predictor of fertility. Its moderate impact on motility is of biological importance but not enough to qualify it as a standalone biomarker.
Second, the number of protein bands presented a very strong positive correlation with motility (r = 0.91), indicating that the diversity of the proteome is a supporting factor in sperm function. Roosters exhibiting a wider spectrum of protein expression were more often characterized by increased motility, which likely indicates that their metabolic and structural pathways are robust and exhibit some degree of overlap. Similar trends have been observed in indigenous poultry and livestock, where protein diversity has been associated with predicting higher fertility outcomes [52]. Nevertheless, correlations with viability (r = 0.133), PMI (r = 0.100), and abnormality (r = 0.314) were weak and statistically non-significant, thus emphasizing that traits other than motility are less reliant on proteomic variation and more controlled by intrinsic genetics or spermatogenic fidelity. The analysis of molecular weight categories, the third point, indicated that high molecular weight proteins (218–247 kDa) had a negative correlation with sperm motility, viability, and PMI counting, whereas they had a positive correlation with abnormality. This suggests that stress-related proteins being overexpressed may lead to impairment of sperm function. On the contrary, proteins in the range of 30–75 kDa, including HSP70/90, SPAG6, and glycolytic enzymes, were those for which semen quality was favorable throughout the study. The study results have categorized mid-range proteins as potential biomarker candidates, but the final identification is dependent on mass spectrometry validation. It should always be taken into account that these were purely associative findings and not causal ones. The non-triviality of the Type I error risk arises from the fact that no adjustments were made to multiple correlations tested at once. Moreover, the lack of replicates and the use of densitometry make the band quantification less robust. It is necessary to conduct further studies using regression modeling, mediation analysis, and a larger sample size to determine whether some proteins directly influence motility or if both are simply co-regulated by broader physiological processes.
Biological interpretation and integration with reproductive physiology
The mentioned relationships correspond to the broader reproductive mechanisms. Proteins with molecular weights of 30–75 kDa are functionally connected to energy metabolism and flagellar movement. GAPDH and ATP synthase are the entry points of the glycolytic and oxidative phosphorylation pathways, respectively, supplying the ATP necessary for motility. SPAG6 stabilizes the axonemal structure, while HSP70/90 facilitates protein folding and protects proteins from oxidative stress. Thus, the positive relationships between these proteins and motility signal the very well-acknowledged functional pathways (Table 5)
On the other hand, the negative correlations with high-MW proteins point towards a stress-related phenotype. The expression of large chaperones, such as HSP90A, has been associated with impaired sperm motility in bulls and infertility in poultry [50,53].
At around 250 kDa, fibronectin is the protein that facilitates sperm-oviduct interactions. However, its change in expression may disrupt both capacitation and sperm storage [54]. These claims suggest that the mid-MW proteins, despite being mostly supportive of sperm function, might get excessively expressed and thus signal cellular stress or dysregulation.
The role of oxidative stress becomes very significant within this context. Reactive oxygen species deteriorate the membrane and weaken the DNA, leading to the death of the sperm and a decrease in the PMI even when motility is still at an acceptable level. Some of the proteins that have been tentatively identified, such as HSP90 and antioxidant enzymes, are all part of the defense mechanism against oxidative damage, thus connecting the proteomic profile to sperm survival pathways [12,15].
This raises an issue that has not been fully examined yet: the transition from motility to diverse protein expression is the primary reason for the latter's vulnerability to oxidative imbalance, which also explains the weak negative correlations with total protein concentration observed in this study. The breeding process not only has to take into account the psychological and behavioral factors. It is possible that the indigenous roosters that were subjected to the repeated collection of semen experienced the stress of handling, which in turn raised the levels of corticosterone and negatively affected the testicles. It has been known that stress can affect protein expression, regardless of fertility level, resulting in a misleading interpretation of the observed associations. Stress-mitigation strategies should be incorporated into breeding management, as they will help ensure that the proteomic biomarkers reflect the true reproductive potential and not just handling artifacts.
Limitations and future perspectives
This research is the first systematic evaluation of sperm protein profiles in Kokok Balenggek roosters; however, it is limited by a few factors. The high variation in semen traits, first of all, necessitates larger sample sizes and multiple measures to account for outliers and environmental effects in the case. Secondly, SDS-PAGE gives only approximate protein separation; without densitometry or replicates, quantification is limited, and protein identification remains speculative. The associations presented here are therefore provisional and require confirmation using LC-MS/MS or Western blotting with specific antibodies. Thirdly, multiple correlation testing increases the risk of Type I error. Although effect sizes like r = 0.91 are very persuasive, stronger inferences require statistical adjustments and scatter plot validation. The study, however, despite these limitations, has laid down the foundation that motility is the semen characteristic most consistently related to protein profiles, especially in the 30–75 kDa range. The future research is suggested to be carried out in the manner of (i) validating the candidate proteins through proteomics, (ii) incorporating oxidative stress markers and hormonal profiles, and (iii) examining protein expression influenced by season and management practice. Such strategies will enhance the mechanistic understanding of rooster fertility and, simultaneously, increase the applicability of proteomic markers for conservation and breeding programs.
Conclusion
The first characterization of sperm protein profiles in the Kokok Balenggek roosters and their correlation with semen quality formed the basis of this study. The ejaculates presented high motility, viability, and plasma membrane integrity, while the SDS-PAGE analysis exhibited protein bands in the range of 30–75 kDa, which were positively correlated with motility. On the contrary, the high-molecular-weight proteins (> 200 kDa) were negatively correlated with semen quality traits. Thus, it can be inferred that mid-range proteins may contribute to reproductive performance, while large proteins may indicate stress-related processes. Nonetheless, the results need to be interpreted with caution, as the correlations are associative and not causal, and the identification of proteins by SDS-PAGE remains tentative without validation via LC-MS/MS or Western blotting. Additionally, differences between males and the potential effects of stress and environmental factors may obscure the observed relationships. In conclusion, the study provided a valuable basis for further research into the fertility of indigenous cocks. However, verifying the selected candidates and enhancing their use in breeding and conservation programs would necessitate the integration of state-of-the-art proteomic tools, oxidative stress markers, and seasonal aspects in future research.
List of abbreviations
AI, artificial insemination; ANOVA, analysis of variance; ATP, adenosine triphosphate; BSP, binder of sperm protein; DNA, deoxyribonucleic acid; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; HMW, high molecular weight; HOST, hypo-osmotic swelling test; HSP, heat shock protein; KBR, Kokok Balenggek rooster; LC-MS/MS, liquid chromatography–tandem mass spectrometry; MW, molecular weight; PBS, phosphate-buffered saline; PMI, plasma membrane integrity; SD, standard deviation; SDS-PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis; SPAG6, sperm-associated antigen 6; SPSS, statistical package for the social sciences.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zhang J Kwon T Kim S Jeong D Plasma farming: non-thermal dielectric barrier discharge plasma technology for improving the growth of soybean sprouts and chickens Plasma 20181228596 https://doi.org/10.3390/plasma 1020025
- 2Ruiz ED Bermejo JVD Jurado JML González FJN Arbulu AA Guzmán JFB Effect of supplementation of a cryopreservation extender with Pectoliv 30 on post-thawing semen quality parameters in rooster species Antioxidants 20241381018 https://doi.org/10.3390/antiox 130810183919926210.3390/antiox 13081018 PMC 11351633 · doi ↗ · pubmed ↗
- 3Getachew T Goshu G Lemma A Effects of commercial and homemade extenders on post-thaw sperm quality and fertility of semen from Horro chicken breed World Vet J 20231323417 https://doi.org/10.54203/scil.2023.wvj 37
- 4Juiputta J Loengbudnark W Koedkanmark T Chankitisakul V Boonkum W Determining the priority semen characteristics and appropriate age for genetic improvement in Thai native roosters P Lo S One 2025204 https://doi.org/10.1371/journal.pone.031953710.1371/journal.pone.0319537 PMC 1196425040173143 · doi ↗ · pubmed ↗
- 5Jaswandi J Ananda A Rusfidra R Subekti K Gusdinal H Sri Wahyuni R Fertility rate, fertility period, and DOC sex ratio of Kokok Balenggek chicken after artificial insemination Adv Anim Vet Sci 2023115795801 https://doi.org/10.17582/journal.aavs/2023/11.5.795.801
- 6Ananda A Jaswandi J Rusfidra R Gusdinal H Sperm longevity and motility in Ringer’s Lactate solution with addition of egg yolk among five phenotypes of Kokok Balenggek chicken Bul Peternakan 202347312735 https://doi.org/10.21059/buletinpeternak.v 47i 3.83647
- 7Li WN Jia MM Peng YQ Ding R Fan LQ Liu G Semen quality pattern and age threshold: a retrospective cross-sectional study of 71,623 infertile men in China, between 2011 and 2017 Reprod Biol Endocrinol 201917107 https://doi.org/10.1186/s 12958-019-0551-23181562910.1186/s 12958-019-0551-2PMC 6902580 · doi ↗ · pubmed ↗
- 8Boeri L Pozzi E Capogrosso P Fallara G Belladelli F Candela L Infertile men with semen parameters above WHO reference limits at first assessment may deserve a second semen analysis: challenging the guidelines in the real-life scenario P Lo S One 2023181280519 https://doi.org/10.1371/journal.pone.028051910.1371/journal.pone.0280519 PMC 985154436656872 · doi ↗ · pubmed ↗