Effects of dietary incorporation of Moringa oleifera leaf meal on hatching characteristics and serum parameters of local Guinea fowl (Numida meleagris) in Togo
Patrik Nukunu Komi Atitso, Kokou Voemesse, Aduayi Akue, Hèzouwè Tchilabalo Meteyake, Kafui Amivi Tete-Benissan

TL;DR
Adding Moringa oleifera leaf meal to Guinea fowl diets improves hatching rates and keet health in Togo.
Contribution
The study demonstrates that Moringa oleifera leaf meal at 0.5-1.5% improves hatching and serum parameters in local Guinea fowl.
Findings
Hatchability and fertility rates improved with Moringa inclusion at 0.5-1.5%.
Embryo mortality decreased in treatments with Moringa leaf meal.
Serum parameters like total proteins and albumin increased in Moringa-fed groups.
Abstract
This study evaluated the effects of Moringa oleifera leaf meal incorporation in local Guinea fowl breeders’ diet on the hatching rate, quality, and serum parameters of Keet in Togo. For this study, 512 breeder Guinea fowls (128 males and 384 females) were assigned randomly to four dietary groups (with four replicates each), named DT0, DT1, DT2, and DT3, containing, respectively, 0%, 0.5%, 1%, and 1.5% of M. oleifera leaf powder in diets. Guinea fowls are raised in confinement. Four incubations (I38, I42, I46, and I50) of 560 eggs were carried out each (Incubated at 37.7°C, with a relative humidity of 55%, the eggs were turned once per hour at a 90° angle until 23 days before transferred to hatching baskets for 3–4 days). These eggs were collected from 384 local Guinea fowls at 38, 42, 46, and 50 weeks of age. Egg weight loss, the duration of hatching events, fertility, hatchability,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
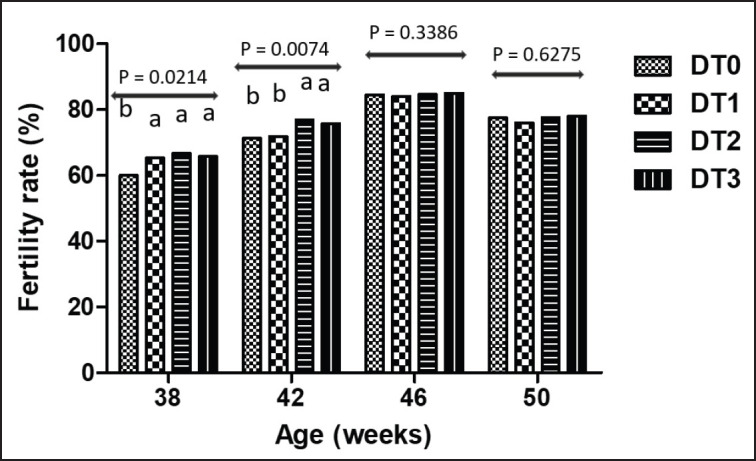 Figure 1
Figure 1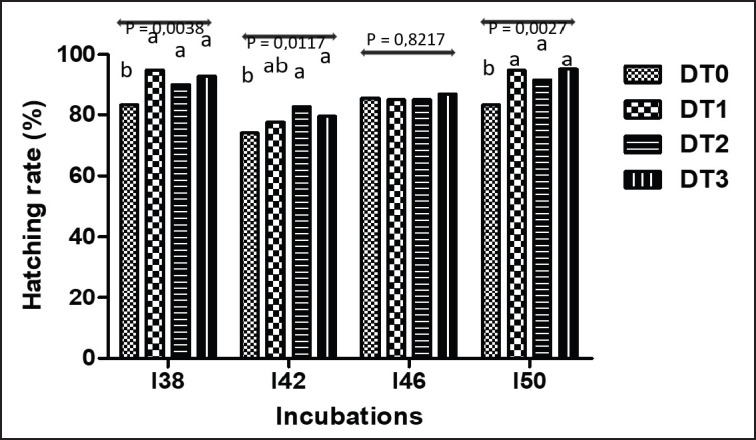 Figure 2
Figure 2| Phytochemical group | Concentrations |
|---|---|
| Total phenols (μg Eq Gallic Acid/mg) | 125.63 ± 2.07 |
| Tannins (μg Eq Gallic Acid/mg) | 87.60 ± 2.65 |
| Flavonoids (μg Eq Rutin/mg) | 176.44 ± 10.45 |
| Polysaccharides (μg Eq Glucose/mg) | 402.80 ± 12.07 |
| Mineral element | Composition (mg/100g) |
| Calcium | 2455 ± 151 |
| Phosphor | 354 ± 22 |
| Magnesium | 510 ± 45 |
| Potassium | 2062 ± 118 |
| Zinc | 2.92 ± 0.6 |
| Copper | 2.15 ± 1.2 |
| Manganese | 12 ± 12.07 |
| Iron | 20 ± 5.7 |
| Selenium | 0.32 ± 0.03 |
| Ingredient | Feed composition according to age and group | |||||||
|---|---|---|---|---|---|---|---|---|
| 22–31 week of age | 32–50 week of age | |||||||
| DT0 | DT1 | DT2 | DT3 | DT0 | DT1 | DT2 | DT3 | |
| Maize | 54 | 53.5 | 53.5 | 53 | 59 | 59 | 59 | 59 |
| Wheat bran | 17 | 17 | 17 | 17 | 13 | 12.5 | 12 | 12 |
| Roasted soybean | 19 | 19 | 18.5 | 18.5 | 18 | 18 | 18 | 17.5 |
| Laying concentrate | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| oyster shell | 7 | 7 | 7 | 7 | 7 | 7 | 7 | 7 |
| Methionine | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Lysine | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
|
| 0 | 0.5 | 1 | 1.5 | 0 | 0.5 | 1 | 1.5 |
| Total | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Calculated analysis | ||||||||
| ME (Kcal/Kg) | 2787 | 2791 | 2795 | 2799 | 2849 | 2860 | 2870 | 2873 |
| Crude protein (%) | 17.72 | 17.67 | 17.45 | 17.40 | 17.06 | 17.13 | 17.21 | 17.13 |
| Calcium (%) | 2.27 | 2.26 | 2.26 | 2.26 | 2.27 | 2.28 | 2.29 | 2.30 |
| Phosphorus (%) | 0.53 | 0.52 | 0.52 | 0.52 | 0.49 | 0.48 | 0.48 | 0.48 |
| Methionine (%) | 0.78 | 0.78 | 0.78 | 0.78 | 0.78 | 0.78 | 0.78 | 0.78 |
| Lysine (%) | 1.07 | 1.07 | 1.05 | 1.05 | 1.03 | 1.03 | 1.03 | 1.03 |
| Methionine + Cysteine (%) | 1.02 | 1.02 | 1.01 | 1.01 | 1 | 1 | 0.99 | 0.99 |
| Parameters | Groups |
| |||
|---|---|---|---|---|---|
| DT0 | DT1 | DT2 | DT3 | ||
| Egg weight (gm) | 39.79 ± 1.65 | 40.31 ± 0.95 | 39.82 ± 0.97 | 39.99 ± 1.07 | 0.9781 |
| Egg weight loss (%) | 11.2 ± 0.31 | 12.84 ± 0.48 | 10.92 ± 0.41 | 11.27 ± 0.32 | 0.0758 |
| Incubation duration (h) | 553.67 ± 1.44 | 553.33 ± 0.44 | 554.67 ± 1.11 | 553.33 ± 0.88 | 0.9695 |
| Duration of internal pipping (h) | 12.5 ± 1a | 12.5 ± 0.53a | 10.5 ± 0.7b | 10.5 ± 0.53b | 0.0157 |
| Duration of external pipping (h) | 22 ± 0.58a | 21 ± 1.02a | 18.5 ± 0.88ab | 17 ± 0,62b | 0.0145 |
| Duration of hatch (h) | 34.5 ± 0.67a | 33.5 ± 1.18a | 29 ± 0.67b | 27.5 ± 0.58b | 0.0050 |
| Keets weight (gm) | 25.35 ± 1.6 | 25.61 ± 1.46 | 25.81 ± 0.79 | 26.59 ± 2.14 | 0.2896 |
| Embryo mortality (%) | 12.84 ± 1.59a | 9.92 ± 0.24b | 11.08 ± 0.34b | 11.12 ± 0.71b | 0.0065 |
| Parameters | Groups |
| ||||
|---|---|---|---|---|---|---|
| DT0 | DT1 | DT2 | DT3 | |||
| Average weight (gm) | Heart | 0.32 ± 0.04 | 0.27 ± 0.02 | 0.27 ± 0.015 | 0.28 ± 0.01 | 0.1090 |
| Liver | 0.61 ± 0.01c | 0.65 ± 0.026bc | 0.65 ± 0.03ab | 0.68 ± 0.03a | 0.0039 | |
| Yolk sac | 5.3 ± 0.16a | 3.43 ± 0.35b | 3.5 ± 0.46b | 3.53 ± 0.23b | 0.0088 | |
| Keet without yolk | 18.57 ± 1.14b | 23.03 ± 0.38a | 23.83 ± 0.67a | 23.8 ± 0.83a | < 0.0001 | |
| Ratio of organ (%) | Heart | 1.26 ± 0.03 | 1.05 ± 0.02 | 1.05 ± 0.02 | 1.05 ± 0.017 | 0.1370 |
| Liver | 2.41 ± 0.01 | 2.54 ± 0.022 | 2.52 ± 0.04 | 2.56 ± 0.035 | 0.0957 | |
| Yolk sac | 20.91 ± 0.18a | 13.39 ± 0.4b | 13.56 ± 0.8b | 13.28 ± 0.4b | 0.0079 | |
| Keet without yolk | 73.25 ± 1.48b | 89.93 ± 0.48a | 92.33 ± 0.59a | 89.53 ± 1.34a | < 0.0001 | |
| Parameters | Groups |
| |||
|---|---|---|---|---|---|
| DT0 | DT1 | DT2 | DT3 | ||
| Total protein (gm/l) | 11.3 ± 0.29c | 32.8 ± 1.1a | 23.83 ± 0.84b | 20.5 ± 0.52b | < 0.0001 |
| Albumin (gm/l) | 5.53 ± 0.33b | 9.37 ± 1.67a | 9.67 ± 0.1a | 7.47 ± 0.1ab | 0.0271 |
| AST (U/l) | 171 ± 8.51 | 195.67 ± 11.57 | 188.67 ± 18.24 | 170.33 ± 5.3 | 0.1986 |
| ALT (U/l) | 7.47 ± 0.93 | 8.73 ± 0.63 | 9.07 ± 0.88 | 7.51 ± 0.34 | 0.5738 |
| Creatinine (mg/dl) | 0.25 ± 0.03 | 0.37 ± 0.03 | 0.27 ± 0.03 | 0.30 ± 0.06 | 0.2503 |
| Uric Acid (mg/dl) | 5.37 ± 0.44 | 6.35 ± 1.1 | 5.75 ± 1.6 | 5.62 ± 0.8 | 0.9327 |
| Urea (gm/l) | 0.25 ± 0.02 | 0.25 ± 0.01 | 0.33 ± 0.01 | 0.28 ± 0.02 | 0.7184 |
| Triglycerides (gm/l) | 1.26 ± 0.13 | 1.47 ± 0.11 | 1.18 ± 0.2 | 1.19 ± 0.06 | 0.1509 |
| Cholesterol (gm/l) | 5.95 ± 0.21a | 4.25 ± 1b | 4.63 ± 0.6b | 4.45 ± 0.23b | 0.0395 |
| HDL-CH (gm/l) | 1.31 ± 0.05 | 1.37 ± 0.1 | 1.80 ± 0.12 | 1.39 ± 0.15 | 0.2324 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMoringa oleifera research and applications · Animal Nutrition and Physiology · Rabbits: Nutrition, Reproduction, Health
Introduction
The breeding of local Guinea fowls (Numida meleagris) is considered a sector for poverty reduction in West African countries, especially in Togo, and contributes to meeting the population’s animal protein requirements [1,2]. Despite its nutritional and cultural significance [1,3], Guinea fowl breeding still faces many difficulties, such as seasonal reproduction of local Guinea fowl, the low availability of fertile eggs and day-old keets (post-hatch juveniles), and their low growth and high mortality rate during the start [1,3,4]. However, Sodjedo et al. [2] reported that Guinea fowl can lay throughout the year if fed ad libitum with adequate feed.
In poultry, several factors, including genetics, breeders’ diet, egg storage duration, and incubation parameters, affect hatchability and the quality of day-old chicks [5–7]. Thus, studies have shown the effects of breeders’ diet on the quality of hatching eggs, hatchability, and the quality of day-old chicks or keets [7-10]. Mustafa et al. [11] showed an improvement in the hatching rate, blood parameters, and immunity of Japanese quails by using feed additives. A positive effect on hatching rate and weight at hatch was also observed by Aberbour et al. [12] using rosemary essential oil in quails.
Moringa oleifera, a leguminous plant rich in quality nutrients and secondary metabolites (carotenoids, flavonoids, tannins, saponins, terpenoids, glycosides, phenolic compounds, and so on) which, by different mechanisms of action, give it antioxidant, antimicrobial, anti-inflammatory, hepatoprotective, hypolipidemic, and other properties [8,9,13,14]. It is used for both human and animal consumption due to its nutritional and phytotherapeutic properties [15-17]. In poultry, an improvement in fertility and hatching rate was observed in aged broiler breeders with dietary supplementation of Moringa leaf extract [18]. Habibi et al. [19] also demonstrated an increase in hatchability with 1% dietary supplementation of either the powder or hydroalcoholic extract of M. oleifera seeds in Chukar partridges. When incorporated up to 15% in the diet of Japanese quail (partially replacing soybean meal), M. oleifera had no detrimental effects on egg quality, fertility, and hatchability of quail eggs [20]. Hatching rate and day-old chick weights increased with the injection of 0.5 μg/ml of M. oleifera leaf extracts into fertile eggs of Isa Brown breeders [8].
Despite all the earlier reports on M. oleifera leaves as a phytogenic feed additive, scientific reports regarding its dietary incorporation by local Guinea fowl breeders on the productive and reproductive performances are scarce. Hence, this study aimed to investigate the effect of incorporating M. oleifera leaf powder into the diets of local Guinea fowl breeders on the hatchability of eggs, the performance of keets, and their blood parameters.
Material and Methods
Ethical approval
All experimental procedures were conducted in accordance with the guidelines approved by the Animal Ethics Committee of the University of Lomé, Togo (Approval No. 008/2021/BC-BPA/FDS-UL).
Study areas
The rearing phase of the Guinea fowl breeders that provided the eggs for incubation was studied by the same authors in the same study areas, and the results were published [10]. The University of Lomé in Togo, through the experimental unit of the Regional Center of Excellence in Poultry Science, served as the setting for our study. The Poultry Production Techniques laboratory of the Center provided the Guinea fowl used for the study.
Moringa oleifera leaves collection and experimental diet formulation
The collection of M. oleifera leaves (MOL) for the rearing phase of Guinea fowl breeders, which provided the eggs for incubation, was studied by the same authors, and the results were published [10]. Thus, before their incorporation into feed, the leaves were air-dried on a clean surface. Table 1 presents the phytochemical group concentrations and mineral element composition of M. oleifera leaf powder. The M. oleifera leaf meal was incorporated into the tested diets at rates of 0%, 0.5%, 1%, and 1.5%. The composition of the different experimental diets (iso-nitrogenous and iso-caloric) is presented in Table 2.
Experimental design and data collection
For this study, four incubations (I38, I42, I46, and I50) of 560 hatching eggs each were carried out. Each incubation was considered a repetition. These eggs were collected from 384 local Guinea fowl breeders at 38, 42, 46, and 50 weeks of age, divided randomly into four dietary treatments (the Guinea fowl had an average weight of 1176.7 ± 2.9 gm [10]), with four replicates each, 24 females for 8 males [2] per replicate. These are the treatments DT0 (fed with a diet containing 0% MOL meal), DT1 (fed with a diet containing 0.5% MOL meal), DT2 (fed with a diet containing 1% MOL meal), and DT3 (fed with a diet containing 1.5% MOL meal) [10]. After 1 week of acclimatization, the experimental diets were served ad libitum to the Guinea fowl starting at 23 weeks of age [10]. The Guinea fowls were reared on litter at a density of 6 per square meter [21] in an open-sided poultry house, partitioned with wood (2.7 m × 2 m) [10]. During the experiment, water was provided ad libitum, and natural light was used (with an average temperature of 25°C during the dry season and 27.3°C during the rainy season) [10].
Thus, for each incubation, 140 eggs per treatment group, stored for 3–7 days, were weighed and individually numbered before being placed in the incubator. The eggs were incubated at 37.7°C and 55% relative humidity. During incubation, the eggs were turned at a 90° angle every hour for 23 days in a Petersime^®^ Vision 96 incubator [22]. The eggs were weighed and candled after 23 days of incubation. Those showing signs of live embryos were transferred to hatching baskets [22,23]. The transferred eggs were individually checked every 3 h to identify internal pipping (IP), external pipping (EP), and hatching [22,23]. Eggs in which the embryo’s beak had pierced the inner shell membrane IP were transferred to a new basket (and examined again individually every 3 h) to detect the time at which the shell (above the air cell) cracked EP [22,23]. Cracked eggs were then placed in new baskets to determine the time of hatching [22,23]. At the end of the hatching period, individual incubation, pipping, and hatching times were recorded to determine their average duration [22,23]. Depending on the treatments, unhatched eggs were recorded, broken open, visually examined, and classified as either infertile or containing dead embryos [22,23]. Organ samples (heart, liver, yolk sac) were collected and weighed from 8 keets per treatment (randomly selected) at hatching. The keets were stunned and slaughtered for the samples [10]. For biochemical analyses, venous blood samples (approximately 2 ml) were collected (from 8 Guinea fowl per treatment) in dry tubes [10]. The collected blood was centrifuged (3,000 rpm) to obtain the serum. Albumin, total protein, alanine aminotransferase (ALT), aspartate aminotransferase (AST), urea, uric acid, creatinine, high-density lipoproteins-cholesterol (HDL), total cholesterol, and triglycerides were then measured [10]. The colorimetric method (using a Mindray BS automatic biochemical analyzer, China) was used to determine these blood parameters. The determinations were done in triplicate.
Calculated parameters
The relative weight loss of the eggs during incubation was determined according to the following formula:
Egg weight loss = 100 × (egg weight at the start of incubation—egg weight on the 23rd day of incubation) / egg weight at setting.
The durations of IP (time of EP—time of IP), EP (hatching time—time of EP), and hatching (hatching time—time of IP) [22] were calculated, as well as the incubation period (time elapsed between the start of incubation and internal pipping) [22].
The number of infertile eggs, dead embryos, and hatched Guinea fowl chicks per treatment was used to calculate the fertility, embryonic mortality, and hatching rates relative to the total number of eggs according to the following formulas:
Fertility = 100 × (Number of fertile eggs / Number of eggs set);
Embryo mortality rate = 100 × (Number of dead embryos / Number of fertile eggs);
Hatching rate = 100 × (Number of Guinea keet hatched / Number of fertile eggs) [21].
The relative weight of the organ was also calculated (Organ weight × 100 / live weight) [9,10].
Statistical analysis
After the homogeneity and normality tests with Bartlett’s test, data analysis was performed using ANOVA with GraphPad Prism 8.1 software. Comparison of means (expressed as mean ± standard deviation and as a percentage) between the different treatments was performed using Tukey’s test. Differences were considered statistically significant when p < 0.05.
Results
Incubation outcomes and embryo mortality
Egg weight, egg weight loss after 23 days of incubation, and incubation duration were similar between treatments (p > 0.05). Concerning hatching events, IP, EP, and hatching duration were higher in DT0 and DT1 groups compared to DT2 and DT3 (p < 0,05), as shown in Table 3. The weight of Guinea fowl keets at hatch and the embryo mortality rate are presented in Table 3. Although the difference was not significant (p > 0.05), keet weight was higher in groups DT1, DT2, and DT3 compared to DT0. Concerning the embryo mortality rate (%), it was higher in the DT0 treatment (12.84 ± 1.59) compared to DT1, DT2, and DT3 (9.92 ± 0.24, 11.08 ± 0.34, and 11.12 ± 0.71, respectively) (p < 0.05).
Fertility
Fertility rate increased in all treatments until 46 weeks of age before decreasing at 50 weeks. At 38 weeks, fertility was higher in DT1, DT2, and DT3 groups compared to DT0, and at 42 weeks of age, the fertility of treatments DT2 and DT3 was higher than that of treatments DT0 and DT1 (p < 0.05). The fertility was similar between treatments at 46 and 50 weeks of age, as shown in Figure 1. On average, fertility increased (p < 0,05) in DT2 and DT3 groups (76.43 ± 3.8 and 76.15 ± 4.5, respectively) compared to DT0 and DT1 groups (73.3 ± 6.6 and 74.2 ± 4.7, respectively).
Fertility rate according to guinea fowl breeders’ age (38, 42, 46, and 50 weeks of age) and Moringa oleifera treatments. DT0, DT1, DT2, DT3: Treatments having received respectively 0%, 0.5%, 1% and 1.5% Moringa oleifera leavesin the diet; a, b Within columns, histograms not sharing the same letters are significantly different (p < 0.05). The rearing lasted 28 weeks.
Hatchability
Figure 2 shows the hatching rate in incubations I38, I42, I46, and I50 according to treatments. Hatchability was significantly higher in group DT2 and DT3 compared to DT0 at I38, I42, and I50 (p < 0.05). The difference was not significant between treatments at incubation I46 (p > 0.05). On average, treatments DT1, DT2, and DT3 (87.54 ± 5.5, 87.03 ± 3.2, and 88.25 ± 4.1, respectively) recorded a higher hatching rate (p < 0.05) than that of treatment DT0 (81.5 ± 3.3).
Hatching rate at I38, I42, I46, and I50 incubations according to Moringa oleifera treatments. DT0, DT1, DT2, DT3: Treatments having received, respectively, 0%, 0.5%, 1%, and 1.5% Moringa oleifera leavesin the diet; I38, I42, I46, I50: incubation of eggs from Guinea fowl breeders aged 38, 42, 46 et 50 weeks, respectively; a, b Within columns, histograms not sharing the same letters are significantly different (p < 0.05).
Average weight of heart, liver, yolk sac, and keet weight without yolk sac
Heart weight was similar between treatments (p > 0.05). Liver weight and keet weight without yolk sac increased in groups DT1, DT2, and DT3 compared to the group DT0 (p < 0.05). Yolk sac weight was higher in DT0 treatment compared to DT1, DT2, and DT3 (p < 0.05), as shown in Table 4.
Biochemical parameters
Serum parameter concentrations of keets according to treatments are presented in Table 5. Total protein and albumin were significantly higher in DT1, DT2, and DT3 treatments compared to DT0 (p < 0.05). Aspartate aminotransferase and alanine aminotransferase of all treatments were not significantly different (p > 0.05). The levels of urea, uric acid, creatinine, triglycerides, and HDL-CH were also similar between treatments (p > 0.05). Total cholesterol levels were slightly decreased in the DT1, DT2, and DT3 treatments compared to the DT0 treatment (p < 0.05).
Discussion
The improvement in fertility with the incorporation of M. oleifera leaf meal is comparable to the results of Ghadimi et al. [18], which showed that the extract of Moringa leaf in dietary supplementation (200 μl/kg) improves the fertility of aged breeder broilers. Improved fertility has also been observed in New Zealand White male rabbits with the use of an aqueous extract of M. oleifera leaves [24], and in Balinese bulls (with supplementation of M. oleifera leaves) [25]. Indeed, living organisms, as a result of normal cellular metabolism and sometimes under the influence of environmental factors, produce reactive oxygen species. Part of the antioxidant needed to balance free radicals and avoid oxidative stress is provided by the diet [9,13,16,18]. According to Diemer et al. [26], Asadi [27], Panner et al. [28], Juan et al. [29], Behnamifar et al. [30], and Mohlala et al. [31], oxidative stress has negative impacts on the reproductive functions (protein oxidation in reproductive cells, DNA damage; the reduction in hormonal balance such as FSH (Follicle-Stimulating Hormone) LH (Luteinizing Hormone) which stimulate the production of testosterone, and consequently decreasing circulating testosterone and suppression of sexual behavior), which leads to a reduction in semen quality, egg fertility, and hatchability. Reducing the negative effects of oxidative stress, thereby improving the reproductive function of roosters, can be achieved by supplementing their feed with antioxidants [32]. The antioxidant properties, stimulated by phenolic compounds, carotenoids, flavonoids, flavanols, vitamins C and E, zinc, and selenium, contained in Moringa leaves could reduce the negative effects of oxidative stress, thus improving egg fertility [13,16,33,34].
Additionally, M. oleifera leaf extract increased serum and testicular testosterone (by inhibiting 6beta-hydroxytestosterone), as well as FSH and LH, semen volume, sperm motility, and sperm viability [24,25,31,35-37]. It increases sexual desire by increasing blood flow to the reproductive organs and stimulating the nervous system [31]. Furthermore, M. oleifera reduces the percentage of abnormal sperm and increases testicular weight [38,39]. Additionally, several nutrients, such as selenium and zinc, found in Moringa leaves [40], have been shown to improve fertility. Indeed, zinc plays a role in protecting the genetic material in the nucleus of sperm cells, which is essential for successful fertility [41]. Supplementing with plants containing selenium increases fertility and hatching rates [42].
The increase in hatching rate with the incorporation of Moringa leaves is consistent with the results obtained by N’nanle et al. [8] in the Isa Brownbreeder’s fertile eggs. This increase in hatchability could be attributed to the availability of energy during the hatching process, which is derived from the metabolism of lipids contained in the yolk, stimulated by Moringa leaves [8,43]. This mechanism is confirmed by the present results, which showed a significant reduction in yolk sac weight in treatments DT1, DT2, and DT3 compared to treatment DT0. However, after detachment of the chorioamniotic membrane during pipping, oxygenation and catabolism of lipid are restricted [44,45]. At this point, the glycogen reserves are primarily used by the hatching muscle [44,45]. Likewise, according to De Oliveira [46], M. oleifera contains substances promoting the storage of glycogen in the liver. The hydrolysis of this glycogen could produce more energy necessary for muscle contraction [8,47]. These results agree with the significant reduction in IP, EP, and hatching duration observed in groups DT2 and DT3 compared to DT0 and DT1 in this study. The increase in the hatching rate of groups DT1, DT2, and DT3 could also be attributed to several nutrients, such as selenium, zinc, and vitamin E, contained in Moringa leaves [14,39] and essential for egg hatching [42,48,49]. Indeed, an increase in the hatching rate with the level of zinc in the diet of laying hens has been reported by Durmus et al. [50]. In addition, the improvement in egg yolk quality because of carotenoids contained in Moringa leaves [9,13] could also increase hatching rate. Indeed, Surai and Sparks [51] showed that birds transfer large quantities of carotenoids into the yolk of their eggs. The tissues developing during embryogenesis are, therefore, protected against the damaging effects of free radicals and peroxides by the transferred carotenoids [52].
The similarity in incubation time and egg weight loss could be linked to the similarity in shell ratio of hatching eggs observed in previous work [10]. The similar weight of keets at hatch may have resulted from the high weight of the yolk sac of keets from treatment DT0 compared to DT1, DT2, and DT3; because keet weight without yolk sac of DT1, DT2, and DT3 groups was significantly higher compared to DT0. The high keet weight without yolk sac in DT1, DT2, and DT3 groups could be attributed to the high albumen ratio of eggs from these groups, as shown in our previous study [10], and could be confirmed by the increase in total proteins and albumin of these groups observed in serum parameters results. This improvement in albumen ratio, likely linked to the quality of protein, energy, vitamins, and mineral salts in Moringa leaves [10,14], would have, through a positive correlation, improved keet weight without a yolk sac at hatch [5,53].
The reduction in embryonic mortality rate in the MOL incorporated treatments compared to the control treatment can be linked to the reduction in oxidative stress at the end of incubation by the activity of antioxidant enzymes stimulated by flavonoids, polyphenols, flavonols, proanthocyanidins, carotenoids, vitamins C and E, zinc, and selenium contained in Moringa leaves [13,34,52].
Concerning serum parameters, the increase in total proteins and albumin in the M. oleifera leaves incorporated treatments compared to the control group may be due to the high albumen ratio of their eggs [10], resulting in more protein compounds available for embryo development. The similarity in AST and ALT levels between the different treatments was also reported by Yuangsroi et al. [54] and shows that M. oleifera did not have a negative influence on the organs (especially the liver) of Guinea fowl chicks whose parents consumed the leaves. Indeed, the release of transaminases (such as ALT and AST) into the blood indicates liver damage [55,56]. Moreover, the slight reduction in AST and ALT levels (although the difference is not significant) in treatment DT3 compared to DT1 and DT2 could be attributed to the hepatoprotective properties of M. oleifera leaves [57]. The reduction in total cholesterol level could be attributed to hypocholesterolemic agents, such as β-sitosterol, contained in Moringa leaves [58].
Conclusion
Results from this study showed that M. oleifera leaf meal, incorporated in local Guinea fowl breeders’ diets, improved egg fertility, embryonic viability during incubation, hatchability, and keet quality at hatch. Thus, M. oleifera leaves powder, when incorporated into the diets of local Guinea fowl breeders at 0.5%, 1%, and 1.5%, can improve keet production. However, according to the results of our previous study on egg production performance of Guinea fowl breeders, the use of 0.5% and 1% M. oleifera leaves in the diet should be encouraged. Further research needs to be done to measure nutrients and bioactive components (amino acids, flavonoids, polyphenols, flavonols, proanthocyanidins, carotenoids, vitamins, and mineral salts, such as zinc, selenium) in local Guinea fowl breeders or their eggs, fed with a diet containing M. oleifera leaf powder. Additionally, an investigation is necessary to explore the post-hatch growth performance and blood parameters of keets from Guinea fowl breeders fed a diet containing M. oleifera leaves, and to elucidate the mechanisms of action by which M. oleifera influences the productive performance of local Guinea fowl.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abdallah N Oyebamiji OA Guinea fowl production in Africa: economic importance and constraints Egypt J Vet Sci 2024 p. 1–15https://doi.org/10.21608/EJVS.2024.320900.2374
- 2Sodjedo C Pitala W LaréL Lombo Y Effect of the season and the sex ratio on the laying and reproductive performance of indigenous guinea fowl (Numida meleagris) in South Togo J Anim Plant Sci 2022522950311
- 3Abdallah N Oluwaseun OA Socio-economic and production dynamics of guinea fowl farming in northern Ghana: insights into health management, challenges, and climate change impacts Trop Anim Health Prod 202557181 https://doi.org/10.1007/s 11250-025-04427-24025911410.1007/s 11250-025-04427-2PMC 12011904 · doi ↗ · pubmed ↗
- 4Ousseini MH Nouri B Laminou SGM Production characteristics of local guinea fowl (Numida meleagris) in the urban commune of Tessaoua, Niger Asian J Adv Agricult Res 2024247709 https://doi.org/10.9734/ajaar/2024/v 24i 7524
- 5Romero-Sanchez H Enting H Van Eck L Van Emous R Kroetz Neto F Leentfaar E et al.Achieving reproductive performance and quality chicks with modern broiler breeders J Appl Poult Res 2025343100539 https://doi.org/10.1016/j.japr.2025.100539
- 6Gregrova M Lichovnikova M Foltyn M Hampel D Interaction between broiler parent stock age and egg pre-incubation duration: effects on embryo development, hatchability, day-old chick weight, and yolk sac weight Animal 20241811101333 https://doi.org/10.1016/j.animal.2024.1013333939641410.1016/j.animal.2024.101333 · doi ↗ · pubmed ↗
- 7Chang A Hallez J Silva MM Can feeding the broiler breeder improve chick quality and offspring performance?Anim Prod Sci 2016568125462 https://doi.org/10.1071/AN 15381
- 8N’nanle O Tété-Bénissan A Tona K Teteh A Voemesse K Decuypere E et al.Effect of in ovo inoculation of Moringa oleifera leaf extract on hatchability and chicken growth performance Eur Poult Sci 20178119 https://doi.org/10.1399/eps.2017.213