Lactobacillus acidophilus alleviates slow transit constipation by modulating 5-HT pathway and gut microbial composition
Yunhe Fan, Hao Qin, Jiayao Liu, Mureed Abbas, Chuanli Yang, Haixia Cheng, Xiushan Dong

TL;DR
Lactobacillus acidophilus helps treat slow transit constipation by improving gut movement and reducing inflammation through changes in gut microbes and the 5-HT pathway.
Contribution
This study reveals a novel mechanism by which L. acidophilus alleviates STC via modulation of the 5-HT pathway and gut microbiota.
Findings
L. acidophilus improved intestinal motility and reduced inflammation by regulating Occludin, TNF-α, and IL-1β.
L. acidophilus influenced 5-HT synthesis and release by modulating TPH1 and Piezo1 expression.
L. acidophilus reshaped the gut microbial community and its interaction network, correlating with improved motility.
Abstract
Slow transit constipation (STC) is a chronic disease characterized by delayed intestinal transit and weakened spontaneous contractions of colonic smooth muscle. Current pharmacological treatments are often associated with adverse effects, highlighting the need for safe and more effective therapeutic strategies. This study investigated the potential role of Lactobacillus acidophilus (L. acidophilus) in regulating intestinal motility and alleviating STC, as well as the underlying mechanism. A humanized mouse model was established by intragastric administration of fecal bacterial suspension from STC patients on alternate days, in order to evaluate the effect of L. acidophilus on constipation. The regulatory effect of L. acidophilus on intestinal motility was evaluated using defecation parameters. Colon histopathology was assessed by hematoxylin-eosin (H&E) staining. Immunohistochemistry…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
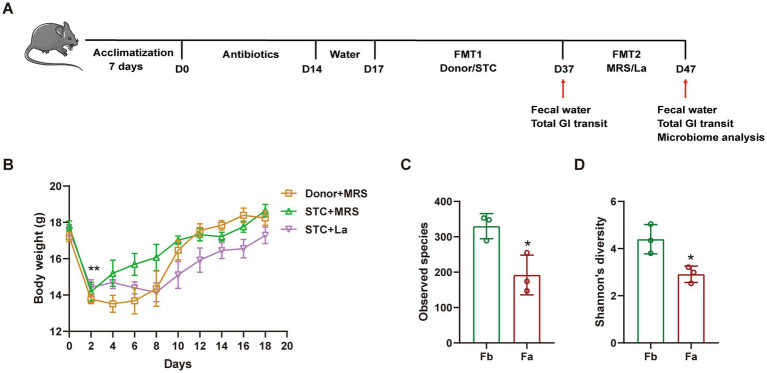 Figure 1
Figure 1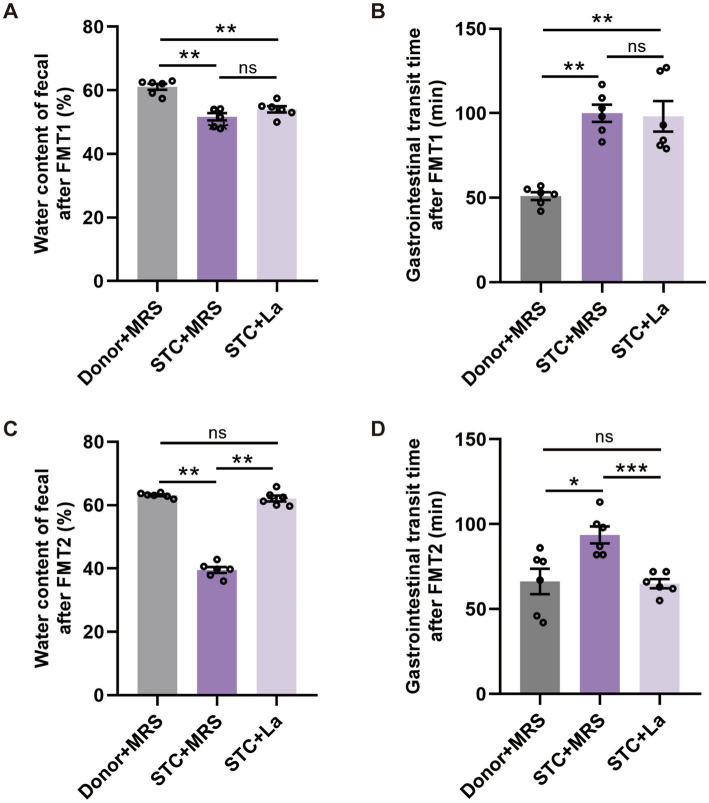 Figure 2
Figure 2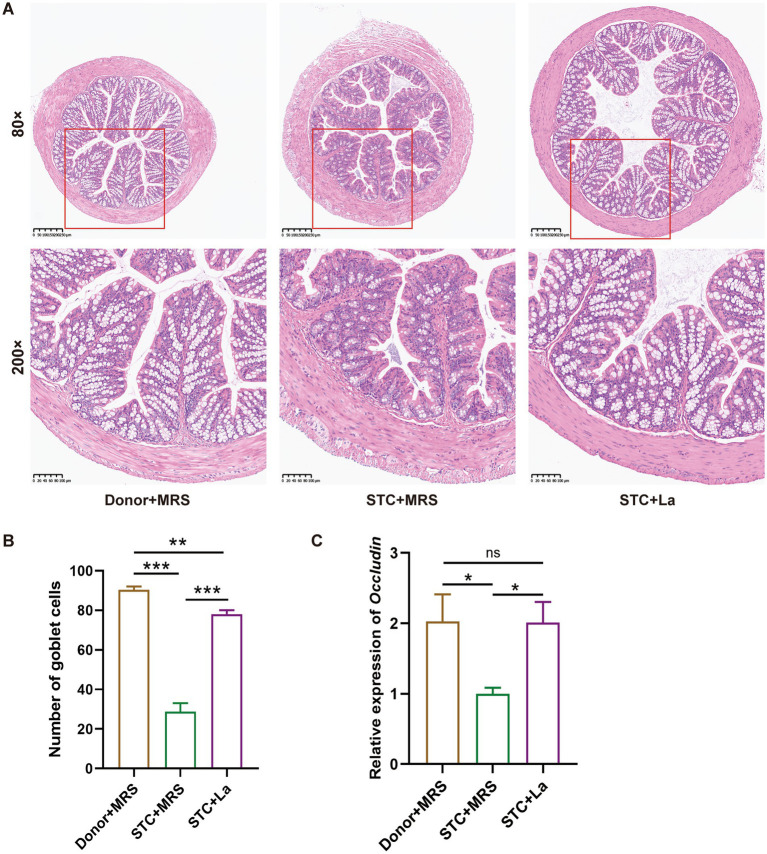 Figure 3
Figure 3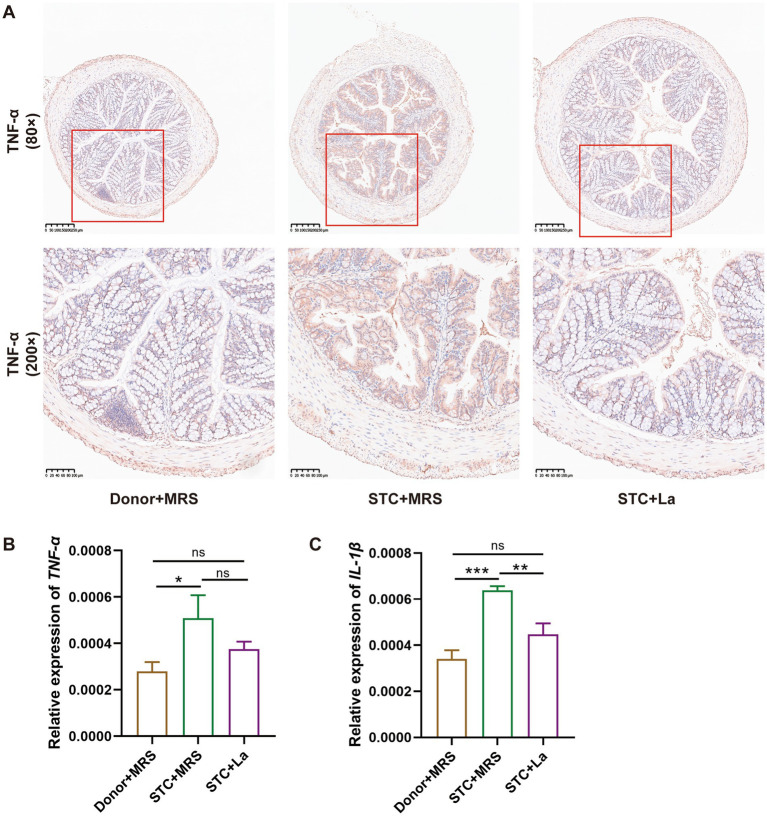 Figure 4
Figure 4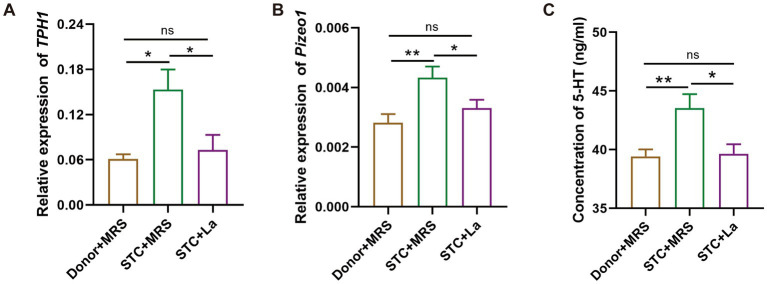 Figure 5
Figure 5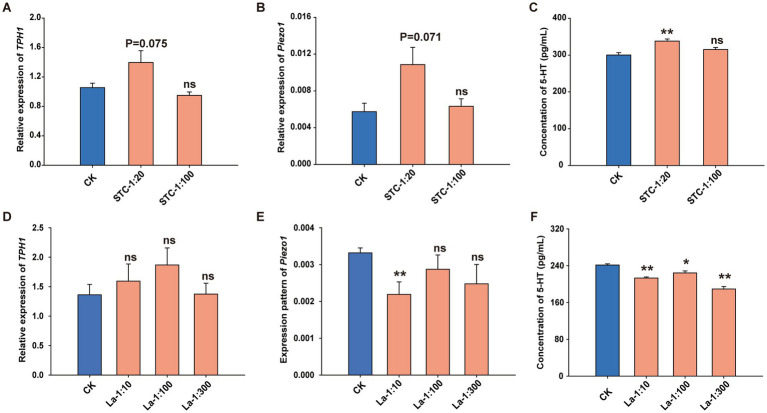 Figure 6
Figure 6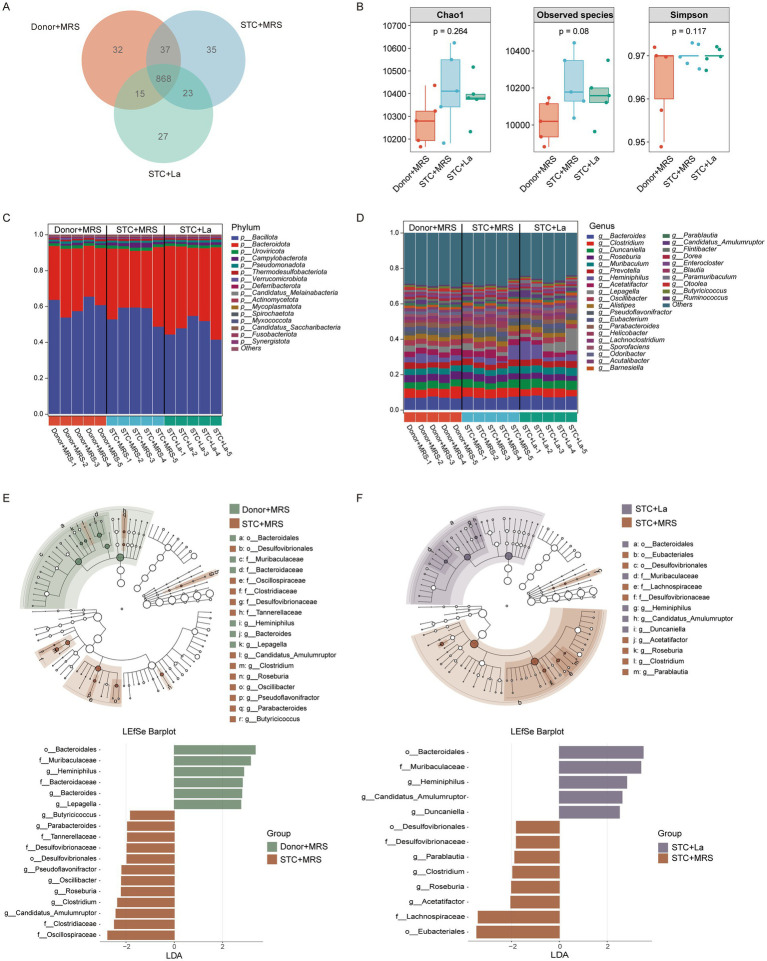 Figure 7
Figure 7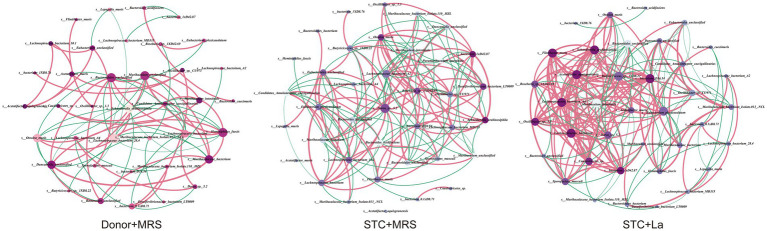 Figure 8
Figure 8| Gene name | Species | Sequence of primers (5–3) |
|---|---|---|
| GAPDH | Mouse | F-ATGGGAAGCTTGTCATCAACG |
| R-AAGACACCAGTAGACTCCACG | ||
| TNF-α | Mouse | F-CGGGCAGGTCTACTTTGGAG |
| R-CAGGTCACTGTCCCAGCATC | ||
| IL-1β | Mouse | F-TGGGAAACAACAGTGGTCAG |
| R-CCATCAGAGGCAAGGAGGAA | ||
| Occludin | Mouse | F-GCGGAAGAGGTTGACAGTCC |
| R-ACTCCCCACCTGTCGTGTAG | ||
| TPH1 | Mouse | F-CTGCGACATCAGCCGAGAA |
| R-TGGTCGGCGTCAAGTTCG | ||
| Piezo1 | Mouse | F-GCTGTCCTCACCCGAATCCA |
| R-GATAGGGCAAGGCGAAGG | ||
| RPL22 | Human | F-TGATTGCACCCACCCTGTAG |
| R-GGTTCCCAGCTTTTCCGTTC | ||
| TPH1 | Human | F-TGCAAAGGAGAAGATGAGAGAATTTAC |
| R-CTGGTTATGCTCTTGGTGTCTTTC | ||
| Piezo1 | Human | F-ACCTGCGTCATCATCGTGTG |
| R-AGTTGGTGCTGTTGGGGAAG |
| Network indices | Donor + MRS | STC + MRS | STC + La |
|---|---|---|---|
| Total nodes | 38 | 38 | 38 |
| Total edges | 110 | 172 | 222 |
| Positive edges (%) | 69.44% | 61.62% | 69.37% |
| Negative edges (%) | 30.56% | 38.38% | 30.63% |
| Average degree | 5.789 | 9.053 | 11.684 |
| Network diameter | 10 | 4 | 5 |
| Average path length | 3.713 | 2.006 | 1.921 |
| Density | 0.156 | 0.245 | 0.316 |
| Average clustering coefficient | 0.752 | 0.673 | 0.828 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGastrointestinal motility and disorders · Gut microbiota and health · Gastroesophageal reflux and treatments
Introduction
Slow transit constipation (STC) is a subtype of functional constipation characterized by impaired colonic motility, resulting in delayed fecal transit and prolonged defecation time. Clinically, STC presents with infrequent bowel movements, difficulty in defecation, and a sense of incomplete evacuation (1, 2). Epidemiological studies indicate that approximately 37.9 to 55% of patients experience STC (3, 4). With the changes in dietary structure and lifestyle, the incidence of STC has shown a progressive annual increase with alarming trend towards younger age groups. The prevalence of constipation in the general population is estimated at around 20% (5) with STC incidence rising with age, and significantly higher among females (4). In the early stages, most patients do not pay sufficient attention and do not receive standardized diagnosis and treatment, leading to disease progression, chronicity and reduced treatment efficacy.
STC not only manifests as gastrointestinal discomforts such as abdominal distension and loss of appetite but also contributes to extragastrointestinal symptoms such as anxiety and depression, which seriously affect the quality of life. Although oral laxatives are used to treat constipation, prolonged use may disrupt intestinal microbiota balance and impair intestinal nerves and smooth muscle function, ultimately reducing intestinal sensitivity (6). Consequently, discontinuation of medication often results in symptom relapse or worsening constipation severity.
Accumulating evidence suggests that probiotics play a beneficial role in relieving constipation. Probiotics are active micro-organisms that colonize the host and alter the composition of a certain part of the host’s flora. Numerous studies have demonstrated that probiotics can alleviate constipation in various ways, including altering the composition of the intestinal flora, improving intestinal environment, and maintaining microbial balance (6, 7). Lactic acid bacteria (LAB) are one of the most common groups of probiotics naturally inhabiting the gastrointestinal tract and are also widely present in fermented foods (8). Various LAB strains possess proven probiotic properties and have been shown to provide diverse physiological benefits to the host (9, 10). Several studies have shown that certain strains of lactobacillus can improve intestinal function and relieve constipation (6, 11, 12). Lactobacillus plantarum PS128 enhances intestinal peristalsis, mucin production and serotonin signal transduction, thereby exerting a laxative effect in mice (13). Clinical trials have also shown that supplementation of Bifidobacteria spp. significantly increases weekly bowel movement frequency and improves stool consistency in patients with chronic constipation (14).
Lactobacillus acidophilus, which was first isolated from infant feces in 1900, is a non-spore-forming Gram-positive bacterium belonging to the genus Lactobacillus with the family Lactobacillaceae. It is recognized as an important intestinal probiotic closely associated with human health (15). L. acidophilus exhibits a broad range of biological functions, including enhancing immunomodulatory, nutritional, regulating the balance of intestinal flora, antioxidant, anti-carcinogenic, and cholesterol lowering effects (16–19). Previous studies have demonstrated that L. acidophilus can regulate the intestinal movement in zebrafish (2). Moreover, a probiotic mixture containing L. acidophilus have been shown to alleviate constipation symptoms in mice (20). Although these findings indicate the potential of L. acidophilus in regulating intestinal motility, its underlying molecular mechanism remains poorly understood. Therefore, the present study aimed to elucidate the regulatory effect of L. acidophilus on intestinal motility by evaluating fecal parameters and intestinal peristalsis in constipated mice, and to further investigate its involvement in the 5-HT signaling pathway.
Materials and methods
Preparation of Lactobacillus acidophilus B636 culture
The bacterial strain was purchased from BeNa Culture Collection (BNCC). After receiving the bacteria, the culture was inoculated into the MRS medium and cultured in an anaerobic incubator at 37 °C for 12 h. When the bacterial concentration reached approximately 10^10^ colony forming units (CFUs)/mL, the bacteria were stored at 4 °C for subsequent use. For cell-based assay the bacterial supernatant was obtained by centrifuging the refrigerated culture at 3000 rpm for 15 min, followed by filtration of supernatant through a 0.22 μm membrane filter.
Animals and cells
Six-week-old adult female C57BL/6J mice were obtained from the Charles River (Beijing, China). Animals were placed in a temperature-controlled (22 ± 2 °C) and humidity-controlled (55–65%) facility at the Medical Experiment Center of Shanxi Bethune Hospital. Mice were maintained under a 12 h light/12 h dark cycle with free access to standard laboratory feed and tap water.
QGP-1 cell line was purchased from COBIOER (Nanjing, China). This cell line is a well-established and internationally recognized in vitro model for studying EC cell biology and the mechanisms of 5-HT synthesis and secretion. The cells were cultured in RPMI 1640 medium (Thermo Fisher Scientific, Shanghai, China), supplemented with 10% fetal bovine serum, and maintained in a humidified incubator at 37 °C with 5% CO_2._
Experimental group design
The experimental design is illustrated in Figure 1A. Following a one-week acclimatization period, mice were randomly divided into three groups: Control group (Donor + MRS), Model group (STC + MRS) and L. acidophilus group (STC + La). All mice were received an antibiotic cocktail (consisting ampicillin [1 g/L], vancomycin [0.5 g/L], neomycin [1 g/L], and metronidazole [1 g/L]) in their drinking water supplemented with 10% sucrose for 2 weeks to deplete the gut microbiota. Afterward, mice had access to normal drinking water for 3 days. Subsequently, the first fecal microbiota transplantation (FMT) was performed. Mice in the Donor + MRS group received donors’ fecal bacterial suspension via oral gavage, whereas the STC + MRS and STC + La groups received fecal bacterial suspension derived from STC patients for 20 consecutive days. Afterwards, a second FMT was conducted: The Donor + MRS and STC + MRS groups received MRS medium by gavage, while the STC + La group received L. acidophilus. All animal procedures were approved by the Animal Care and Use Committee of Shanxi Bethune Hospital.
*The experimental design (A) animal experimental flowchart. (B) Body weight. The alpha diversity of microbiomes identified in mice on (C) observed species and (D) Shannon’s diversity. Fb, before FMT; Fa, after FMT. p < 0.05, n = 6.
Fecal moisture content test
After the first FMT (FMT1) and the second FMT (FMT2), each mouse was individually placed in a separate cage. Fresh fecal samples were collected from each mouse within 2 h. The wet weight of freshly collected fecal samples was recorded immediately. Samples were then dried in an oven at 60 °C to a constant weight. The fecal moisture content (%) was calculated as follows: (wet weight − dry weight)/wet weight × 100 (21).
Measurement of total gastrointestinal transit time (TGTT)
To assess TGTT, mice were fasted for 6 h and then gavaged with 0.3 mL of a non-absorbable dye solution (6% carmine red in 0.5% methylcellulose). The interval between gavage and the excretion of the first red-stained fecal pellet was documented as TGTT. During the measurement, mice were housed in separate cages lined with white filter paper to enhance visualization of excreted dye (22).
Histopathology and immunohistochemistry
The colon tissues of each group of mice were dissected and placed separately into different 1.5 mL EP tubes. Samples were fixed in 4% paraformaldehyde, followed by standard dehydration, paraffin embedding and sectioning at 5 μm. The sections were stained with H&E, dehydrated and mounted with neutral mounting medium. Histopathological changes were examined using an OLYMPUS BX51 microscope.
Immunohistochemistry (IHC) was performed to assess the localization and relative expression level of TNF-α. Paraffin-embedded mouse colon sections underwent antigen retrieval by pressure heating in citrate buffer (pH 6.0). After which, endogenous peroxidase activity was blocked by 3% hydrogen peroxide for 10 min. Sections were then incubated with 5% (w/v) bovine serum albumin for 25 min at room temperature for protein blocking. Primary antibody incubation against TNF-α (Proteintech, Wuhan, China) (60291-1-IG) (dilution 1:1000) was performed overnight at 4 °C. After washing, sections were incubated with a horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature. Hematoxylin counterstaining, dehydration, clearing, and mounting were subsequently performed. Stained slides were visualized by light microscopy, and the IHC quantification was performed using ImageJ software.
Expression analysis of related genes
Transcript levels of target genes were measured by reverse transcription quantitative PCR (RT-qPCR). Total RNA was extracted from mouse colon tissues or QGP-1 cells using the RNAiso Plus kit (Takara, Dalian, China) according to the manufacturer’s instructions. 1 μg of total RNA was reverse-transcribed into first-strand cDNA using M-MLV reverse transcriptase (TaKaRa, Dalian, China). Primers were designed using Primer Premier 5.0 software (Table 1).
RT-qPCR was performed in a Light Cycler^®^ 480II (Roche, Basel, Switzerland) using SYBR Green Real-time PCR Master Mix (Promega, Madison, WI, USA). The 20 μL reaction mixture comprised: 10 μL 2× SYBR Green PCR Master Mix, 0.8 μL of each primer (0.4 μmol/L), 8 μL 10-fold diluted cDNA template, and nuclease-free water. Thermal cycling conditions included: 95 °C for 60 s (initial denaturation); 40 cycles of 95 °C for 5 s (denaturation) and 55 °C for 31 s (annealing/extension). Amplification specificity was confirmed by melt curve analysis. Relative mRNA expression was calculated using the method and normalized to an internal reference gene.
Enzyme-linked immunosorbent assay (ELISA)
After FMT2 treatment, blood samples were collected from the retro orbital sinus into 1.5 mL centrifuge tubes. Samples were centrifuged at 4000 rpm for 20 min, after which the supernatant was transferred to fresh 1.5 mL tubes and stored at −80 °C for later use. The collected mouse serum samples were two-fold dilution with 1× PBS. Then, the 5-HT concentration in the serum was detected using commercial mouse 5-HT enzyme-linked immunosorbent assay kit (Ruixin Biotech, Guangzhou, China) according to the manufacturer’s instructions.
After 24 h of adding varying concentrations of bacterial supernatant, the culture media from QGP-1 cells were collected and centrifuged it at 4000 rpm for 20 min to remove cell debris. The commercial human 5-HT enzyme-linked immunosorbent assay kit (Ruixin Biotech, Guangzhou, China) was used to detect 5-HT in the supernatant of QGP-1 cells according to the manufacturer’s protocol. The absorbance of the final product was measured on a microplate at a wavelength of 450 nm.
16S rRNA gene sequencing and analysis
16S rRNA sequencing was performed to analyze intestinal microbiota of mice before and after antibiotic treatment, as described previously (23). Total genomic DNA was extracted from mice fecal samples using the QuickGene DNA tissue kit (Kurabo, Neyagawa, Japan) following the manufacturer’s protocol. The V3-V4 hypervariable regions of the bacterial 16S rRNA gene were amplified using primers 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′). Purified PCR amplicons were paired-end sequenced on an Illumina MiSeq platform following standard protocols. Raw sequences were demultiplexed, quality-filtered, and analyzed using QIIME2. Denoising, chimera removal, and amplicon sequence variant (ASV) inference were performed with DADA2. Taxonomic assignment was conducted against the SILVA database with a 97% similarity threshold. Alpha diversity indices (Shannon, Chao1) were calculated from rarefied ASV tables.
Metagenomic sequencing and analysis
The gut microbiome of mice was investigated using the metagenomic shotgun sequencing method as described previously (24). Genomic DNA was extracted from fecal samples using the Magnetic Bead-Based Stool DNA Extraction Kit (BioTeKe, Beijing, chain). DNA libraries were constructed using the TruSeq Nano DNA Library Preparation Kit-Set (Illumina, USA) following the manufacturer’s instructions. Metagenome libraries were then sequenced on an Illumina NovaSeq 6000 platform with PE150 at Shanghai Biotree Biotech Co., Ltd. (Shanghai, China). Fastp software (v0.23.4) was used to remove the reads that contained adaptor contamination, low quality bases and undetermined bases. Then sequence quality was also verified using fastp. Quality filtered reads were first aligned to mice genome1 by using bowtie (v2.2) to filter out host contaminations. Then, the remaining reads were subjected to de novo assembly for each sample using MEGAHIT (v1.2.9) and used to assign microbial functions and taxonomy. Metaprodiga (v2.6.3) was used to predict the coding regions (CDS) of the assembled contigs, and CDS sequences of all samples were clustered using MMseq2 (v15-6f452) to obtain unigenes. DIAMOND (v0.9.14) was used to perform a taxonomic assessment of the microbiota based on the NR database. The wilcoxon rank-sum test was used to identify the differentially abundant species, and significances were declared at p < 0.05 and |log2-fold change| > 1. An assignment of microbial functions was done using the Kyoto Encyclopedia of Genes and Genomes (KEGG). Network analysis was performed using R 3.2.6 and Gephi v.0.9.2 software; p < 0.05, |R| > 0.6 were considered statistically significant.
Statistical analysis
Statistical analyses were performed using Student’s t-test for comparisons between two groups. Data are presented as mean ± standard deviation (SD). Statistical significance was defined as *p < 0.05, **p < 0.01 and ***p < 0.001.
Results
Effect of L. acidophilus on fecal excretion parameters in STC mice
Following a 14-day antibiotic pretreatment regimen, constipation was induced in mice through FMT using samples obtained from STC patients. L. acidophilus was subsequently administered, and defecation related parameters were evaluated on days 37 and 47 to assess its therapeutic effects on STC-associated symptoms (Figure 1A). A significant reduction in body weight was observed in all mice on the second day after antibiotic administration (p < 0.01). Subsequently, the body weight of the mice gradually increased, indicating that the impact of antibiotics on body weight was transient and diminished over time (Figure 1B). Alpha diversity analysis demonstrated a significant reduction in gut microbial richness in antibiotic-treated mice compared to pre-treatment baselines (p < 0.05), suggesting that antibiotic intervention depleted a portion of the intestinal microbiota (Figures 1C,D).
After FMT1, mice receiving STC patient-derived microbiota exhibited markedly altered defecation parameters compared to control group (Donor + MRS). These mice exhibited a significant reduction in fecal water content (p < 0.01) and a significant increase in TGTT (p < 0.01), indicating successful induction of constipation phenotypes by STC patients’ fecal microbiota (Figures 2A,B). Subsequently, the FMT2 was performed. The results showed that the mice treated with L. acidophilus had significantly different fecal parameters compared to the STC + MRS group. Specifically, the water content of the fecal was significantly increased (p < 0.01), while their TGTT was significantly decreased (p < 0.001) (Figures 2C,D).
*Effect of L. acidophilus on excretion parameters in mice. (A) Water content of fecal and (B) gastrointestinal transit time after FMT1. (C) Water content of fecal and (D) gastrointestinal transit time after FMT2. *p < 0.05, **p < 0.01, **p < 0.001, n = 6.
Effect of L. acidophilus on colonic histopathology in mice
To evaluate the impact of L. acidophilus on colonic pathology, H&E staining was performed to examine the histological architecture of mouse colon. The results showed that in the control group, the structure of the colon tissue in mice was intact and clear, with orderly arranged goblet cells, and no observable pathological lesions. However, the STC + MRS group exhibited extensive intestinal mucosal layer necrosis, in which the necrotic intestinal glands were replaced by proliferating fibrous tissue, the structure of the mucosal layer disappeared, accompanied by diffuse inflammatory cell infiltration. Additionally localized enlargement of the muscle fiber gaps and loosened fiber arrangement were observed in the muscular layer. Following L. acidophilus treatment, the inflammatory infiltration was markedly reduced and the goblet cells appeared more regularly arranged (Figure 3A). The number of goblet cells in the colon tissue was calculated using the ImageJ software. The results showed that compared with the control group, the number of goblet cells in the STC + MRS group was significantly reduced (p < 0.001). Conversely, the STC + La group showed a significant increase in goblet cell numbers relative to STC + MRS group (p < 0.001) (Figure 3B). Further, expression of Occludin gene were examined using RT-qPCR as shown in Figure 3C. The results showed that compared with the Donor + MRS group, the mRNA level of Occludin in the STC + MRS group was significantly decreased (p < 0.05), and upon the addition of L. acidophilus, the expression of Occludin was significantly increased compared with the STC + MRS group (p < 0.05).
*Lactobacillus acidophilus improves intestinal barrier damage. (A) Representative images of H&E staining of the colon tissue in each group. (B) The number of goblet cells in colon tissue from each group. (C) The relative expression of Occludin in colon sections. *p < 0.05, **p < 0.01, **p < 0.001.
Effect of L. acidophilus on inflammatory cytokine levels in mice
IHC analysis demonstrated significantly elevated expression of inflammatory cytokine TNF-α in the colonic mucosa of the STC + MRS group compared with the control group, whereas no significant difference in TNF-α expression was observed between the STC + La and control group (Figure 4A). These observations were further validated by RT-qPCR analysis, which revealed significantly increased mRNA expression level of TNF-α and IL-1β in the STC + MRS group relative to the control group (p < 0.05; p < 0.01). In contrast, the expression levels of TNF-α and IL-1β in the STC + La group did not differ significantly from those in the control group (Figures 4B,C).
*Lactobacillus acidophilus modulates the expression of inflammatory cytokines. (A) Immunohistochemical analysis of TNF-α protein expression in mouse colon tissue. (B) RT-qPCR analysis of TNF-α mRNA expression in colonic tissues. (C) RT-qPCR analysis of IL-1β mRNA expression in colonic tissues. *p < 0.05, **p < 0.01, **p < 0.001.
Effect of L. acidophilus on 5-HT signal transduction pathway
Previous studies have shown that STC can affect the 5-HT signaling pathway (25, 26). Therefore, both in vivo and in vitro experiments were performed to investigate the regulatory effects of L. acidophilus on 5-HT signal transduction.
To assess the effect of L. acidophilus on the 5-HT signaling pathway in vivo, the expression of tryptophan hydroxylase 1 (TPH1) and Piezo1 in colonic tissues was quantified by RT-qPCR and the concentration of 5-HT in the serum was detected using ELISA. Compared with the control group, mice in the STC + MRS group exhibited significantly elevated expression of TPH1 and Piezo1 (p < 0.05; p < 0.01); in contrast, administration of L. acidophilus significantly reduced the expression of TPH1 and Piezo1 compared with the STC + MRS group (p < 0.05) (Figures 5A,B). Consistently, the concentration of 5-HT in the serum of mice in the STC + MRS group was significantly higher than in the control group (p < 0.05), whereas after adding L. acidophilus, 5-HT concentration was significantly decreased relative to STC + MRS group (Figure 5C).
*Lactobacillus acidophilus reduces 5-HT production by targeting TPH1 and Piezo1 in mice. (A) RT-qPCR analysis of TPH1 mRNA expression in colonic tissues. (B) RT-qPCR analysis of Piezo1 mRNA expression in colonic tissues. (C) Measurement of 5-HT levels in serum. *p < 0.05, *p < 0.01.
To further assess the regulatory effects of L. acidophilus on 5-HT signaling in vitro, QGP-1 cells were treated with supernatants of STC patients’ bacteria and L. acidophilus, respectively, at different concentrations. After 24 h of incubation, the transcript levels of TPH1 and Piezo1 were quantified by RT-qPCR, and secreted 5-HT levels in cell supernatants were measured by ELISA. Compared with the control group, after QGP-1 cells were incubated with the supernatant of STC patient cultures at a concentration of 1: 20, the expression levels of TPH1 and Piezo1 showed an upward trend (p = 0.075; p = 0.071) respectively (Figures 6A,B). The ELISA results also indicated that after the cells were treated with the supernatant of STC patient cultures at a concentration of 1: 20, the content of 5-HT in the cell culture medium significantly increased (p < 0.01) (Figure 6C). Conversely, after incubation of L. acidophilus solution with QGP-1, although the expression of TPH1 showed no significant alteration compared to the control (Figure 6D), the expression of Piezo1 was significantly down-regulated at 1: 10 (p < 0.01) (Figure 6E), and the concentration of 5-HT in the culture supernatants was significantly reduced across all tested L. acidophilus concentrations (p < 0.05, p < 0.01) (Figure 6F).
*Lactobacillus acidophilus reduces 5-HT production by targeting TPH1 and Piezo1 in QGP-1 cells. Relative expression of (A) TPH1 and (B) Piezo1 after treated with STC patients’ fecal supernatant. (C) 5-HT content in the cell supernatant after treated with STC patients’ fecal supernatant. Relative expression of (D) TPH1 and (E) Piezo1 after treated with L. acidophilus supernatant. (F) The content of 5-HT in the cell supernatant after treated with L. acidophilus supernatant. *p < 0.05, *p < 0.01.
Lactobacillus acidophilus improves gut microbiota dysbiosis in STC model mice
Growing evidence indicates that the gut microbiota plays a critical role in maintaining intestinal health, and dysbiosis of the gut microbial community is closely associated with a range of gastrointestinal dysfunctions. To further investigate the effects of L. acidophilus on gastrointestinal motility, DNA was extracted from mouse fecal samples and subjected to metagenomic sequencing analysis. Venn diagram analysis at species level showed that the numbers of intestinal bacteria in the Donor + MRS, STC + MRS, and STC + La groups were 952, 963, and 923, respectively (Figure 7A). To evaluate disparities in gut microbiota diversity, alpha diversity analyses were conducted. The analysis revealed no statistically significant differences in the Chao1, Observed species, and Simpson indices (Figure 7B), demonstrating that overall microbial richness and diversity did not differ significantly among the groups.
Effects of L. acidophilus on the composition of gut microbiota in constipation mice. (A) Venn diagram. (B) Comparison of α-diversity indices (Chao, observed species, and Simpson) among groups; (C) taxonomic distribution of bacterial communities at the phylum level; (D) taxonomic distribution of bacterial communities at the genus level; LEfSe analysis identifies the most differentially abundant bacterial taxa from phylum to species level between donor + MRS and STC + MRS groups (E); and between STC + La and STC + MRS groups (F).
To further characterize microbial community composition, taxonomic differences were analyzed at multiple levels. At the phylum level, Bacillota and Bacteroidota were the dominant phyla in all groups, collectively accounting for more than 90% of the total microbial abundance (Figure 7C). Compared with the STC + MRS group, the STC + La group exhibited a significant decrease in the relative abundance of Bacillota (STC + MRS group: 0.558 vs. STC + La group: 0.480, p = 0.043) and a significant increase in the abundance of Bacteroidota (STC + MRS group: 0.360 vs. STC + La group: 0.450, p = 0.038). At the genus level (Figure 7D), L. acidophilus intervention significantly reduced the relative abundances of Clostridium (STC + MRS group: 0.053 vs. STC + La group: 0.044, p = 0.016) and Roseburia (STC + MRS group: 0.040 vs. STC + La group: 0.033, p = 0.026), both belonging to Bacillota phylum, while increasing the relative abundance of Muribaculum (STC + MRS group: 0.037 vs. STC + La group: 0.042, p = 0.028), which is part of Bacteroidota phylum. These findings suggest that L. acidophilus alleviates constipation in mice, primarily through its association with Bacillota and Bacteroidota in the gut. Furthermore, Linear Discriminant Analysis (LDA) combined with Effect Size (LEfSe) analysis was used to identify differences in gut microbiota composition among groups. The results showed that the dominant microbiota in the Donor + MRS and STC + La groups included o_Bacteroidales, f_Muribaculaceae, and g_Heminiphilus, whereas the STC + MRS group was primarily characterized by g_Roseburia, g_Clostridium, and f_Desulfovibrionaceae (Figures 7E,F).
Network correlation analyses were conducted in three experimental groups to evaluate the competitive associations within the intestinal microbial communities of the mice at the species level. The top 38 bacterial species with the highest relative abundances were selected for analysis, with each node representing a bacterium and node size corresponding to its relative abundance. Each line represents the interaction between bacteria, with pink lines indicating positive correlations and green lines indicating negative correlations. In the Donor + MRS group, a total of 110 connections were identified among the 38 nodes, of which 69.44% represented positive correlations, The STC + MRS group exhibited 172 connections, with 61.62% being positive correlations. Notably, the STC + La group displayed the highest number of interactions with 222 connections, 69.37% of which were positive correlations (Figure 8). Overall, the microbial co-occurrence network in the STC + La group was the most complex. Furthermore, comparative analysis of network topology revealed marked differences among the three groups in key parameters, including average degree, average path length and network density (Table 2).
Co-occurrence networks of the top 38 species among different groups. Nodes represent species and their sizes indicate different relative abundance. The lines between nodes indicated the Spearman correlation (Spearman’s correlation greater than 0.6 or lower than −0.6), and the color of lines indicated the correlation coefficient (pink, positive; blue, negative).
Discussion
STC is a common gastrointestinal disorder characterized by delayed colonic peristalsis and obstruction of intestinal contents excretion. This condition substantially affects patients’ physical and psychological well-being, as well as their overall quality of life, and may potentially lead to secondary gastrointestinal complications. Laxatives are commonly used for symptomatic relief; however, they may provide rapid improvement, long-term use can easily lead to an imbalance in the intestinal homeostasis, and can impair intestinal nerves and smooth muscles function, ultimately reducing the sensitivity of the intestine (6). Notably, discontinuation of laxative therapy often results in severe constipation with exacerbated symptoms. A large number of studies have shown that the gut microbiota plays a crucial role in maintaining intestinal homeostasis and peristalsis. Patients with STC generally have an imbalance in the intestinal microbiota, with a significant reduction in beneficial bacteria (such as Bifidobacteria and Lactobacilli), and an increase in the proportion of potentially pathogenic bacteria such as Clostridium and Clostridium perfringens (27). Numerous studies have demonstrated that probiotic supplementation can alleviate constipation by modulating gut microbiota composition. However, research has predominantly focused on multi-strain probiotic formulations, such as Leven PRO-CR and SynBalance SmilinGut, rather than on the effects of individual probiotic strains (20, 26). Recent clinical studies have reported significant improvement in constipation symptoms among patients following FMT, accompanied by a marked increase in L. acidophilus abundance as revealed by metagenomic analysis. These findings suggest a potential role for L. acidophilus in the regulation of intestinal motility (24). Although L. acidophilus is a well-established probiotic, and formulations containing L. acidophilus have been shown to alleviate constipation (28), it remains unclear whether L. acidophilus alone is sufficient to exert these effects. Therefore, in the present study, we systematically investigated the impact of single-strain L. acidophilus administration on intestinal motility using a mouse model of constipation and further explored the underlying molecular mechanisms involved.
Given the crucial role of gut microbiota in regulating intestinal motility, an antibiotic cocktail was administered in this study to deplete the indigenous intestinal microbes in mice. Although transient reduction in body weight was observed during antibiotic treatment, likely due to profound diarrhea induced by the treatment, 16S rRNA gene sequencing confirmed a significant reduction in gut microbial diversity following antibiotic administration. This approach is consistent with clinical practice where bowel cleansing using antibiotics is performed prior to FMT. Following antibiotic-mediated depletion of gut microbiota, a STC mouse model was established via FMT using fecal samples from patients with STC. This approach fundamentally differs from conventional constipation model induced by loperamide hydrochloride. Although the loperamide-induced model is widely adopted due to its operational simplicity and consistent induction of stable constipated phenotypes (29–31), it fails to adequately recapitulate female-specific constipation phenotypes and exhibits poor clinical relevance to the complex etiology of human constipation. Therefore, this study established a constipation mouse model using fecal microbiota from STC patients. This approach directly recapitulates the characteristic gut dysbiosis observed in constipated patients. Model mice successfully reproduced the microbial community structure of STC patients. Consequently, it provides a more physiologically relevant representation of the complete pathogenic cascade from microbial disruption to functional impairment. Furthermore, as this model is derived directly from human clinical samples, it exhibits superior predictive validity and enhanced translational potential for evaluating the efficacy of probiotic interventions.
Previous studies have reported that loperamide-induced constipation is associated with a significant reduction in gut microbial diversity (32, 33). However, our results showed that, compared with the Donor treatment group, the intestinal microbiota diversity of mice fed with fecal suspension from STC patients did not show significant change. We propose that the intestinal flora disorder in STC patients is not simply a decrease in microbial diversity, but rather a disruption in the balance between beneficial and pathogenic bacteria. Furthermore, network correlation analysis revealed that, compared to the Donor + MRS group, the bacterial interactions in the STC + MRS group were notably more complex, with a significant increase in negative correlations among bacterial species. Supplementation with L. acidophilus modified these inter-bacterial interactions, markedly enhanced positive correlations among bacterial taxa. These findings suggest that L. acidophilus may improve intestinal motility by remodeling gut microbial interaction networks rather than by simply increasing microbial diversity.
Among probiotics, Lactobacillus and Bifidobacterium have been extensively investigated for their ability to modulate the gut microbiota composition and intestinal function. Previous studies have shown that Lactobacillus plantarum promotes intestinal motility by regulating the 5-HT signaling pathway and enhancing mucus secretion (13), whereas Bifidobacterium longum alleviates constipation by improving intestinal barrier integrity (34). In this study, we demonstrated that administration of L. acidophilus significantly ameliorated constipation symptoms in mice, as evidenced by improved excretion parameters. Furthermore, HE staining revealed substantial structural damage to the intestinal tissue in STC mice, accompanied by significantly reduced expression of Occludin and a substantial upregulation of inflammatory factors (TNF-α and IL-1β). These findings are consistent with previous reports (32, 35). Growing evidence suggests that constipation is closely associated with dysfunction of the intestinal mucosal immune system (36). Constipated patients exhibit immune activation and a certain degree of mucosal inflammation (37). Activated immune cells release a range of inflammatory cytokines and neurotransmitters, which can disrupt the intestinal mucosal barrier, ultimately impairing gut sensation and motility. For example, interferon-γ (IFN-γ) has been shown to increase intestinal permeability by downregulating Zonula Occludens-1 (ZO-1) expression and altering the distribution of tight junction proteins, such as Occludin (38). These mechanism provide a plausible explanation for our observations that STC mice showed significantly decreased Occludin expression and elevated levels of inflammatory factors in the colon. Importantly, L. acidophilus treatment increased Occludin expression and decreased expression of TNF-α and IL-1β, thereby improving intestinal barrier structure and ultimately alleviating constipation symptoms.
Furthermore, the 5-HT signal transduction pathway is also closely associated with the pathophysiology of constipation. As a key neurotransmitter and paracrine signaling molecule, 5-HT plays a crucial role in the gastrointestinal tract. Approximately 90% of 5-HT in the human body is synthesized by intestinal enterochromaffin cells (ECs) through the catalytic activity of TPH1 (39). Newly synthesized 5-HT is released into the intestinal lumen via ion channels. Studies have demonstrated that Piezo1 is a mechanosensitive ion channel expressed on ECs, capable of sensing mechanical stimuli (40, 41). Following 5-HT release, a portion of 5-HT is taken up into cells via Serotonin-selective reuptake transporter (SERT) and subsequently degraded, while the remaining fraction exerts biological effects through binding to specific 5-HT receptors on the cell membrane. Previous studies have shown reduced intestinal 5-HT level in loperamide hydrochloride-induced constipation models (32, 42). In contrast, increased TPH1 expression and elevated 5-HT level have been observed in rectal biopsy samples from patients with constipation (43). Moreover, in patient populations with constipation-predominant irritable bowel syndrome (IBS-C) and functional constipation (FC), a negative correlation between defecation frequency and 5-HT concentration has been identified (44). The discrepancies among these results may be attributed to the complex effects of 5-HT on gastrointestinal motor activity. In the present study, a constipation mouse model was established by transplanting fecal bacterial suspension from patients with STC. The results showed a significant increase in colonic 5-HT levels in constipated mice, consistent with previous findings (24). In addition, the results indicated that compared with the control group, the relative expression of TPH1 and Piezo1 in the colon of constipated mice was significantly elevated. However, L. acidophilus treatment effectively regulated and normalized the expression of TPH1 and Piezo1, and these findings were corroborated by in vitro cell experiments. These findings could be attributed to both the activation and desensitization of 5-HT receptors and the characteristics of the experimental STC model employed.
Conclusion
STC is an intractable constipation with unknown etiology and uncertain pathogenesis, significantly impacting patients’ physical and mental well-being as well as their overall quality of life. In this study, a humanized constipation mouse model was established via FMT, and the therapeutic effect of L. acidophilus on constipation was demonstrated by analyzing the defecation parameters. Through integrated in vivo and in vitro analyses, we elucidated the mechanisms by which L. acidophilus alleviates constipation. Our findings demonstrate that it not only repairs the intestinal barrier, but also modulates the 5-HT pathway by regulating the expression of TPH1 and Piezo1. Additionally, metagenomic sequencing revealed that L. acidophilus remodels the gut microbiota structure and interaction network. In summary, L. acidophilus exerts its therapeutic effect through a multi-faceted action on the gut barrier, serotonin signaling, and microbial ecology.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mazzone A Strege PR Gibbons SJ Alcaino C Joshi V Haak AJ . micro RNA overexpression in slow transit constipation leads to reduced Na(V)1.5 current and altered smooth muscle contractility. Gut. (2020) 69:868–76. doi: 10.1136/gutjnl-2019-318747, 31757880 PMC 7147984 · doi ↗ · pubmed ↗
- 2Wang J Yin L Zheng W Shi S Hao W Liu C . Lactobacillus rhamnosus GG normalizes gut dysmotility induced by environmental pollutants via affecting serotonin level in zebrafish larvae. World J Microbiol Biotechnol. (2022) 38:222. doi: 10.1007/s 11274-022-03409-y, 36100774 · doi ↗ · pubmed ↗
- 3Chaichanavichkij P Vollebregt PF Tee SZY Scott SM Knowles CH. Slow-transit constipation and criteria for colectomy: a cross-sectional study of 1568 patients. BJS Open. (2021) 5:zrab 049. doi: 10.1093/bjsopen/zrab 049, 34052848 PMC 8164778 · doi ↗ · pubmed ↗
- 4Tanner S Chaudhry A Goraya N Badlani R Jehangir A Shahsavari D . Prevalence and clinical characteristics of dyssynergic defecation and slow transit constipation in patients with chronic constipation. J Clin Med. (2021) 10:2029. doi: 10.3390/jcm 10092027, 34065116 PMC 8126009 · doi ↗ · pubmed ↗
- 5Vazquez Roque M Bouras EP. Epidemiology and management of chronic constipation in elderly patients. Clin Interv Aging. (2015) 10:919–30. doi: 10.2147/CIA.S 54304, 26082622 PMC 4459612 · doi ↗ · pubmed ↗
- 6Dimidi E Christodoulides S Scott SM Whelan K. Mechanisms of action of probiotics and the gastrointestinal microbiota on gut motility and constipation. Adv Nutr. (2017) 8:484–94. doi: 10.3945/an.116.014407, 28507013 PMC 5421123 · doi ↗ · pubmed ↗
- 7Lai H Li YF He Y Chen F Mi B Li J . Effects of dietary fibers or probiotics on functional constipation symptoms and roles of gut microbiota: a double-blinded randomized placebo trial. Gut Microbes. (2023) 15:2197837. doi: 10.1080/19490976.2023.2197837, 37078654 PMC 10120550 · doi ↗ · pubmed ↗
- 8MrvčićJ Stanzer D ŠolićE Stehlik-Tomas V. Interaction of lactic acid bacteria with metal ions: opportunities for improving food safety and quality. World J Microbiol Biotechnol. (2012) 28:22806724:2771–82. doi: 10.1007/s 11274-012-1094-222806724 · doi ↗ · pubmed ↗