Review of the southern African slender stonebashers, genus Heteromormyrus Steindachner 1866 (Teleostei: Mormyridae), with description of six new species
Tadiwa I. Mutizwa, Wilbert T. Kadye, Pedro H. N. Bragança, Albert Chakona

TL;DR
This paper describes six new species of slender stonebashers in southern Africa and provides detailed taxonomic information to better understand their diversity.
Contribution
The paper formally describes six new species of Heteromormyrus and provides redescriptions of two existing ones.
Findings
Six new species of Heteromormyrus are described, including two endemic to the Kwanza River system.
Species in the genus are morphologically similar but can be distinguished by features like scale counts and fin ray counts.
The Kwanza River system has five known species, suggesting higher taxonomic diversity in this region.
Abstract
Recent molecular studies have advanced our knowledge of the taxonomic diversity and generic placement of the slender stonebashers, previously placed in the genus Hippopotamyrus, in southern Africa. These fishes were recently transferred to the genus Heteromormyrus whose range encompasses the Kwanza, Kunene, Okavango, Zambezi, Pungwe and Buzi River systems in southern Africa, as well as the southern tributaries of the Congo River system. The present study builds on previous research that identified at least eight candidate species within the Heteromormyrus ansorgii species complex by providing formal descriptions for six new species and redescriptions of Heteromormyrus pauciradiatus and H. ansorgii s.s. The Kwanza River system is peculiar because it currently has five known species in this genus, some of which are co‐distributed, whereas the other river systems have only one or two…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
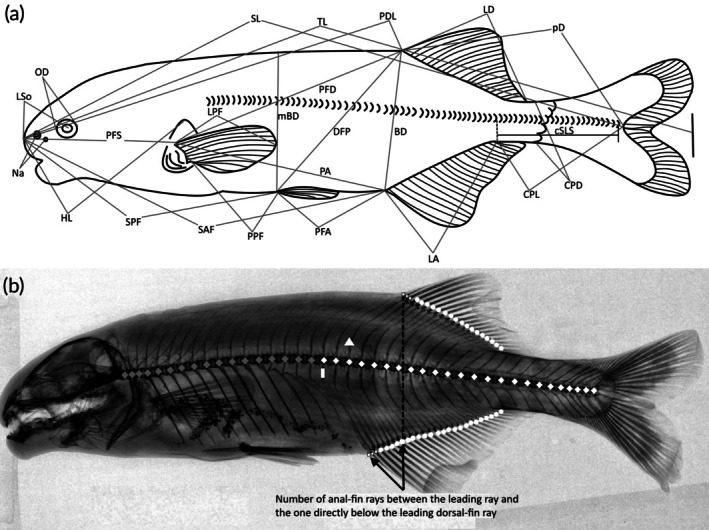 FIGURE 1
FIGURE 1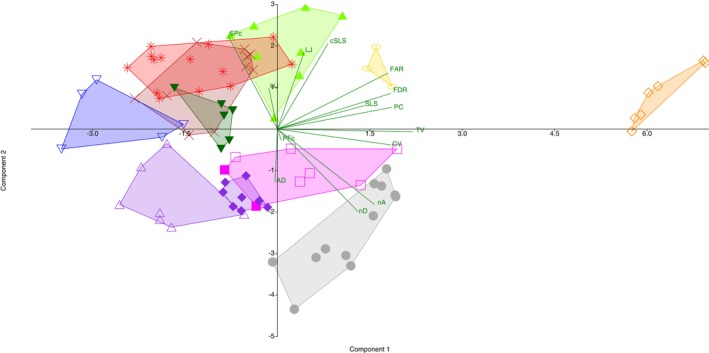 FIGURE 2
FIGURE 2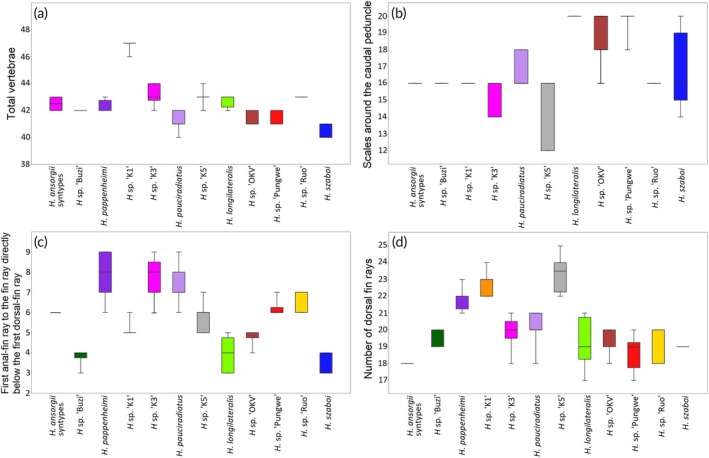 FIGURE 3
FIGURE 3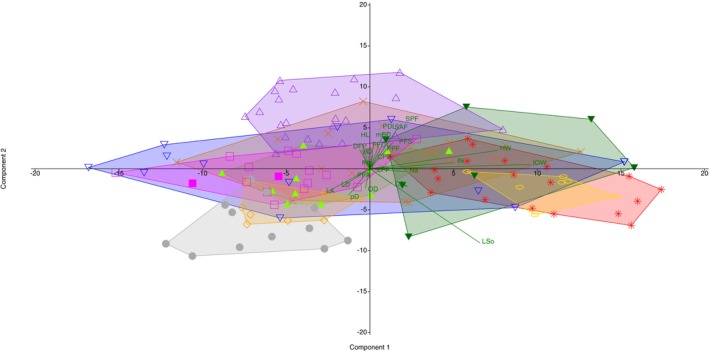 FIGURE 4
FIGURE 4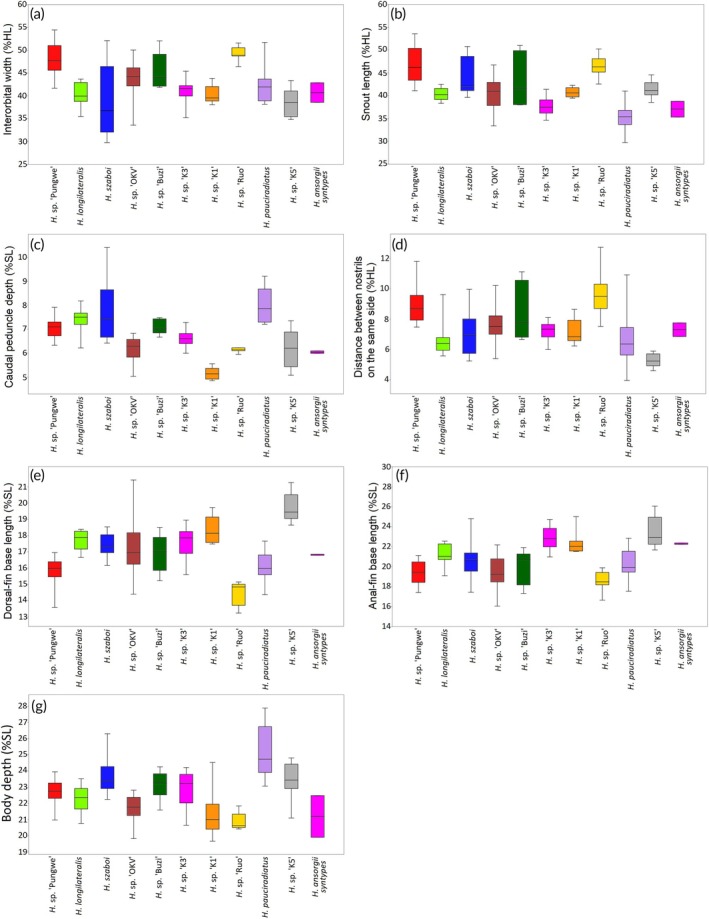 FIGURE 5
FIGURE 5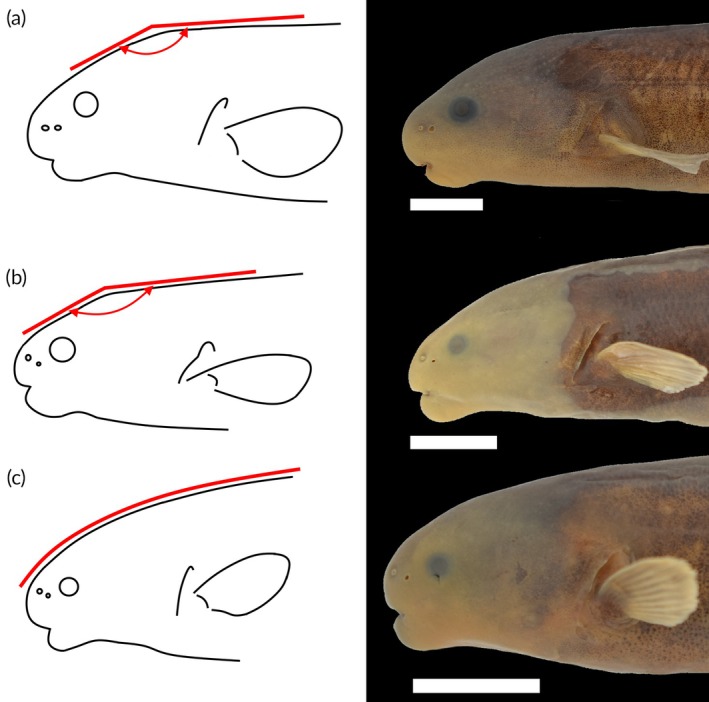 FIGURE 6
FIGURE 6 FIGURE 7
FIGURE 7 FIGURE 8
FIGURE 8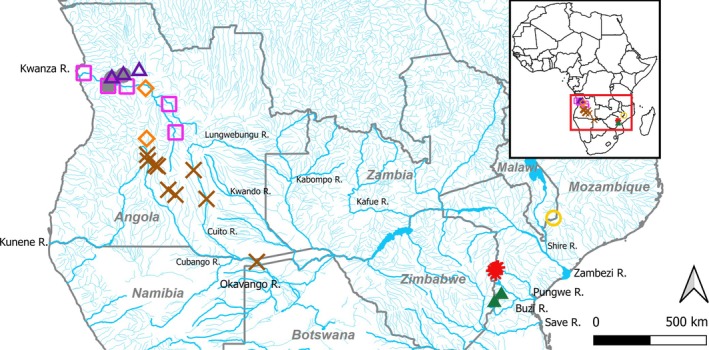 FIGURE 9
FIGURE 9 FIGURE 10
FIGURE 10 FIGURE 11
FIGURE 11 FIGURE 12
FIGURE 12 FIGURE 13
FIGURE 13 FIGURE 14
FIGURE 14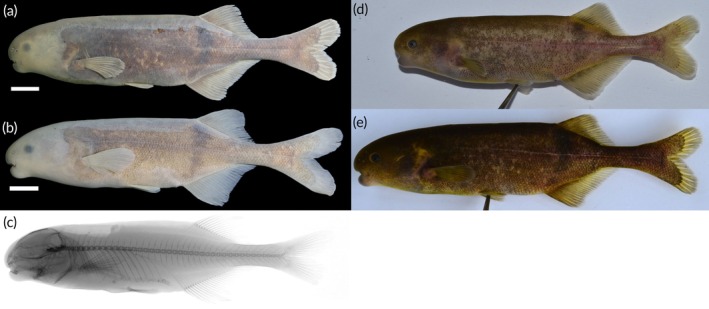 FIGURE 15
FIGURE 15 FIGURE 16
FIGURE 16| Principal component | 1 | 2 |
|---|---|---|
| Eigenvalue | 5 | 2.79 |
| % variance | 35.71 | 19.95 |
| Pectoral‐fin rays (PFc) | 0.01 | −0.01 |
| Number of scales around the caudal peduncle (SPc) | −0.15 | 0.45 |
| Number of scales along the caudal peduncle (cSLS) | 0.17 | 0.4 |
| Lateral‐line scales (SLS) | 0.28 | 0.1 |
| First anal‐fin ray to first dorsal‐fin ray (AD) | −0.01 | −0.24 |
| Pre‐caudal vertebrae (PC) | 0.36 | 0.1 |
| Caudal vertebrae (CV) | 0.36 | −0.08 |
| Number of vertebrae before the first dorsal‐fin radial (FDR) | 0.36 | 0.17 |
| Number of vertebrae before the first anal‐fin radial (FAR) | 0.35 | 0.26 |
| Total vertebrae (TV) | 0.43 | −0.01 |
| Dorsal‐fin rays (nD) | 0.26 | −0.41 |
| Anal‐fin rays (nA) | 0.31 | −0.35 |
| Lower‐jaw teeth (LJ) | 0.09 | 0.37 |
| Upper‐jaw teeth (UJ) | −0.04 | 0.17 |
| Principal component | 1 | 2 |
|---|---|---|
| Eigenvalue | 55.91 | 21.59 |
| % variance | 40.79 | 15.75 |
| Pre‐dorsal length (PDL) | 0.05 | 0.28 |
| Pre‐anal length (SAF) | 0.07 | 0.28 |
| Pre‐pelvic length (SPF) | 0.12 | 0.35 |
| Pre‐pectoral length (PFS) | 0.09 | 0.2 |
| Pectoral fin to anal fin (PA) | −0.03 | 0.06 |
| Pelvic fin to anal fin (PFA) | −0.05 | −0.05 |
| Pectoral fin to dorsal fin (PFD) | −0.01 | 0.16 |
| Dorsal fin to pelvic fin (DFP) | −0.04 | 0.15 |
| Dorsal‐fin base length (LD) | −0.11 | −0.14 |
| Anal‐fin base length (LA) | −0.16 | −0.18 |
| Dorsal fin to caudal fin (pD) | −0.08 | −0.2 |
| Caudal peduncle length (CPL) | 0.08 | −0.12 |
| Caudal peduncle depth (CPD) | 0.01 | 0.11 |
| Pectoral‐fin length (LPF) | 0.01 | 0.01 |
| Pectoral fin to pelvic fin (PPF) | 0.04 | 0.15 |
| Body depth (BD) | −0.03 | 0.15 |
| Mid body depth (mBD) | 0.02 | 0.21 |
| Head length (HL) | −0.03 | 0.19 |
| Length of the snout (LSo) | 0.42 | −0.58 |
| Distance between nostrils on the same side (Na) | 0.14 | −0.02 |
| Orbit diameter (OD) | −0.02 | −0.15 |
| Head width (HW) | 0.49 | 0.14 |
| Distance between anterior nostrils (IN) | 0.32 | 0.05 |
| Interorbital width (IOW) | 0.61 | 0.03 |
|
|
|
|
|
|
| |
|---|---|---|---|---|---|---|
| Number of specimens | 15 | 8 | 12 | 22 | ||
| Total length | 113.2 | 113.5 | 66.7–167.5 (109.4 ± 26.4) | – | – | 40.0–135.1 (78.8 ± 28.1) |
| Standard length | 98.5 | 100.1 | 59.5–142.6 (96.8 ± 22.6) | 80–180* | 53–139* | 36.3–118.7 (69.4 ± 24.8) |
| Head length | 24.5 | 21.9 | 12.8–29.4 (21 ± 4.3) | – | – | 9.5–27.2 (17.2 ± 5.5) |
| Pre‐dorsal length | 63.8 | 68.1 | 62.1–67.0 (64.9 ± 1.4) | – | 41.3–47.0* | 64.0–68.8 (65.9 ± 1.4) |
| Pre‐anal length | 60.7 | 65.1 | 56.6–61.0 (59.3 ± 2) | – | 18.0–22.0* | 58.4–63.6 (61.1 ± 1.6) |
| Pre‐pelvic length | 40.4 | 43 | 35.6–40.7 (38.9 ± 1.7) | – | – | 39.4–45.6 (42.6 ± 2) |
| Pre‐pectoral length | 24.6 | 23 | 21.8–25.3 (23.5 ± 1) | – | – | 24.1–29.1 (26.7 ± 1.3) |
| Pectoral fin to anal fin | 35.9 | 39.8 | 33.2–38.4 (36.3 ± 1.6) | – | – | 32.9–40.9 (35.4 ± 1.5) |
| Pelvic fin to anal fin | 20 | 20.7 | 17.3–21.8 (20.2 ± 1) | – | – | 16.5–21.5 (18.8 ± 1.3) |
| Pectoral fin to dorsal fin | 42.2 | 46 | 41.6–45.4 (43.4 ± 1.3) | – | – | 39.0–44.6 (42.5 ± 1.4) |
| Dorsal fin to pelvic fin | 31.3 | 33.7 | 31.4–36.0 (33.4 ± 1.2) | – | – | 30.4–37.3 (34.1 ± 1.8) |
| Dorsal‐fin base length | 16.9 | 16.8 | 15.6–19.0 (17.4 ± 1) | – | 15.2–19.1* | 14.4–17.7 (16.4 ± 1.5) |
| Anal‐fin base length | 22.2 | 22.4 | 21.0–24.7 (22.8 ± 1.1) | – | 19.3–22.3* | 17.5–22.8 (20.1 ± 1.5) |
| Dorsal fin to caudal fin | 38.8 | 38.8 | 36.8–40.3 (38.8 ± 1.1) | – | – | 34.9–40.2 (38.1 ± 1.6) |
| Caudal peduncle length | 18.7 | 16.2 | 18.2–21.4 (19.4 ± 1.2) | – | 16.4–21.1* | 17.6–23.8 (20.4 ± 1.4) |
| Caudal peduncle depth | 6 | 6.1 | 6.0–7.2 (6.6 ± 0.4) | – | 6.9–8.6* | 7.3–9.2 (8.1 ± 0.7) |
| Pectoral‐fin length | 16.2 | 16.2 | 14.5–18.0 (16.2 ± 1) | – | 17.2–20.9* | 16.6–21.5 (18.6 ± 1.3) |
| Pectoral fin to pelvic fin | 15.9 | 17.3 | 14.7–18.0 (16.6 ± 0.9) | – | – | 16.1–19.6 (17.8 ± 1.1) |
| Body depth | 19.9 | 22.5 | 20.6–24.2 (22.8 ± 1.2) | – | 22.2–24.2* | 23.1–27.9 (25.4 ± 1.6) |
| Mid body depth | 20.4 | 20.9 | 18.7–23.4 (21.3 ± 1.4) | – | – | 22.5–27.3 (24.9 ± 2.1) |
| Head length | 24.9 | 21.9 | 19.4–23.6 (21.9 ± 1.3) | – | 25.8–29.2* | 22.9–27.5 (25.1 ± 1.3) |
| Length of the snout | 35.3 | 38.8 | 34.6–41.4 (36.5 ± 4.1) | – | 24.7–30.0* | 29.7–41.0 (35.5 ± 2.6) |
| Distance between nostrils on the same side | 7.8 | 6.8 | 6.0–8.1 (7.2 ± 0.6) | – | – | 3.9–10.9 (6.5 ± 1.6) |
| Orbit diameter | 15.9 | 17.4 | 7.7–20.3 (13 ± 3.2) | – | 12.4–16.7* | 9.4–16.5 (12.6 ± 1.9) |
| Head width | 47.8 | 50.7 | 47.5–61.4 (54.1 ± 3.4) | – | – | 50.1–61.2 (55.9 ± 2.7) |
| Distance between anterior nostrils | 24.7 | 22.8 | 21.0–30.5 (24.7 ± 2.3) | – | – | 23.4–31.0 (26.1 ± 1.9) |
| Interorbital width | 28.6 | 32.9 | 35.2–45.4 (40.7 ± 2.7) | – | 24.7–30.0* | 38.2–51.7 (42 ± 3) |
| Counts | ||||||
| Number of scales around caudal peduncle | 16 | 16 | 16 (14–16) | 12–14* | 20–22* | 16 (16–18) |
| Number of scales along caudal peduncle | 17 | 16 | 18 (17–19) | – | – | 16 (14–18) |
| Number of scales along lateral line | 70 | 69 | 72 (68–77) | 71–80* | 70–77* | 70 (60–74) |
| Teeth on upper jaw | 7 | 5 | 7 (6–8) | 5–7* | 6–10* | 7 (6–7) |
| Teeth on lower jaw | 4 | 9 | 8 (7–9) | 6–8* | 8–10* | 8 (6–9) |
| Number of pectoral‐fin rays | 10 | 9 | 10 (9–11) | – | – | 10 (9–11) |
| Number of pelvic‐fin rays | 6 | 5 | 6 (6–6) | – | – | 6 |
| X‐ray counts | ||||||
| Number of specimens | _ | _ | 8 | 8 | – | 7 |
| First anal‐fin ray to first dorsal‐fin ray | 6 | 6 | 7 (6–9) | 9 (6–9) | – | 7 (6–9) |
| Total vertebrae | 42 | 43 | 43 (42–44) | 42 (42–43) | – | 41 (40–42) |
| Pre‐caudal vertebrae | 18 | 18 | 18 (18–19) | 18 (17–18) | – | 18 (18–19) |
| Caudal vertebrae | 24 | 25 | 25 (24–25) | 24 (24–25) | – | 23 (22–24) |
| Vertebrae before the first dorsal‐fin radial | 20 | 21 | 21 (21–22) | 19 (19–20) | – | 20 (20–21) |
| Vertebrae before the first anal‐fin radial | 19 | 18 | 19 (19–20) | 18 | – | 18 (18–19) |
| Dorsal‐fin rays | 18 | 18 | 20 (18–21) | 22 (21–23) | 17–20* | 20 (18–21) |
| Anal‐fin rays | 24 | 26 | 25 (23–27) | 27 (26–28) | 22–23* | 23 (23–26) |
|
|
|
|
|
|
| |
|---|---|---|---|---|---|---|
| Number of specimens | 11 | 13 | – | 8 | – | 12 |
| Total length | 64.2–183.0 (141.2 ± 36.6) | 46.0–148.5 (84 ± 30.9) | 133.1 | 66.2–173.0 (121.4 ± 34.2) | 173 | 85.0–176.0 (138.2 ± 31.7) |
| Standard length | 57.1–162.0 (125.4 ± 32.7) | 41.3–134.5 (75 ± 27.8) | 118.3 | 59.6–152.5 (108.7 ± 29.7) | 151 | 73.1–155.0 (120.6 ± 27.6) |
| Head length | 13.2–36.1 (27.3 ± 6.6) | 9.7–33.7 (18.8 ± 6.9) | 27.3 | 14.3–33.2 (24.1 ± 5.8) | 36.2 | 16.9–36.2 (26.9 ± 5.8) |
| Pre‐dorsal length | 60.8–66.5 (63.7 ± 1.9) | 62.0–66.6 (65.1 ± 0.8) | 63.7 | 61.9–63.7 (62.5 ± 0.6) | 65.3 | 58.9–65.3 (61.8 ± 2.1) |
| Pre‐anal length | 59.5–62.2 (60.6 ± 0.9) | 58.5–63.3 (61.8 ± 0.9) | 61.7 | 56.8–62.6 (58.6 ± 1.7) | 60.8 | 54.6–60.8 (57 ± 2) |
| Pre‐pelvic length | 38.4–42.8 (40.3 ± 1.3) | 40.0–45.5 (44 ± 0.9) | 40.3 | 36.7–40.3 (38.8 ± 1.1) | 40.3 | 35.0–40.3 (37.6 ± 1.7) |
| Pre‐pectoral length | 22.4–26.4 (24.1 ± 1.4) | 24.5–29.6 (27.3 ± 1.4) | 23.3 | 22.4–26.2 (23.5 ± 1.3) | 25 | 21.3–25.7 (23.7 ± 1.5) |
| Pectoral fin to anal fin | 34.6–40.2 (38.2 ± 1.5) | 34.1–38.3 (36.3 ± 1.2) | 38 | 33.1–38.0 (35.6 ± 1.6) | 36.8 | 33.1–36.8 (34.9 ± 1.1) |
| Pelvic fin to anal fin | 18.2–22.5 (20.6 ± 1.1) | 17.0–19.3 (18 ± 0.8) | 21 | 17.9–21.0 (19.9 ± 1) | 21 | 18.5–21 (19.5 ± 0.9) |
| Pectoral fin to dorsal fin | 39.1–43.8 (41.9 ± 1.6) | 38.9–41.5 (40.5 ± 0.8) | 43.5 | 38.9–43.5 (41.4 ± 1.6) | 42.3 | 38.4–42.4 (40.1 ± 1.2) |
| Dorsal fin to pelvic fin | 29.7–33.2 (31.7 ± 1.1) | 28.8–33.5 (31.2 ± 1) | 31.7 | 30.5–33.8 (31.6 ± 1.1) | 33.3 | 29.2–33.7 (31.4 ± 1.3) |
| Dorsal‐fin base length | 17.2–18.4 (17.7 ± 0.6) | 16.2–18.5 (17.4 ± 0.7) | 17.6 | 17.5–19.7 (18.4 ± 0.9) | 20.5 | 18.6–21.3 (19.8 ± 0.9) |
| Anal‐fin base length | 19.1–22.6 (21.1 ± 1.1) | 17.4–24.8 (20.6 ± 1.9) | 21.7 | 21.5–25.0 (22.4 ± 1.3) | 23.4 | 21.7–26.1 (23.6 ± 1.6) |
| Dorsal fin to caudal fin | 35.6–42.1 (39.3 ± 1.8) | 37.4–40.5 (38.7 ± 1.1) | 39.4 | 38.3–40.9 (40.1 ± 0.9) | 40.5 | 39.8–44.9 (42 ± 1.7) |
| Caudal peduncle length | 19.9–23.4 (21.2 ± 0.9) | 18.2–22.2 (20.2 ± 1.7) | 20.4 | 19.2–22.7 (20.7 ± 1.1) | 20.8 | 20.8–23.1 (21.6 ± 0.8) |
| Caudal peduncle depth | 7.0–8.2 (7.4 ± 0.5) | 6.4–10.4 (7.8 ± 1.2) | 5.3 | 4.9–5.6 (5.1 ± 0.2) | 7.4 | 5.1–7.4 (6.2 ± 0.8) |
| Pectoral‐fin length | 12.9–19.0 (17.1 ± 1.6) | 16.7–19.5 (18.1 ± 0.9) | 17 | 15.6–17.7 (16.7 ± 0.8) | 18.5 | 17.3–20.6 (18.9 ± 1) |
| Pectoral fin to pelvic fin | 17.1–19.9 (18.6 ± 0.9) | 16.5–21.7 (19.4 ± 1.7) | 18.6 | 15.6–18.6 (17 ± 1.1) | 17.1 | 14.4–17.1 (15.9 ± 0.7) |
| Body depth | 20.8–23.5 (22.3 ± 0.8) | 22.2–26.3 (23.6 ± 1.3) | 21 | 19.7–24.5 (21.3 ± 1.6) | 24.6 | 21.1–24.8 (23.4 ± 1.1) |
| Mid body depth | 19.2–23.8 (21.5 ± 1.5) | 18.8–26.1 (23 ± 1.8) | 21.1 | 19.5–22.2 (21 ± 1) | 21.9 | 18.6–23.2 (21.1 ± 1.3) |
| Head length | 19.7–23.7 (22 ± 1.1) | 22.8–27.6 (25.1 ± 1.6) | 23.1 | 21.2–24.1 (22.4 ± 1) | 24 | 20.4–24.0 (22.4 ± 1.3) |
| Length of the snout | 38.3–42.5 (40.3 ± 1.3) | 39.6–50.8 (44.2 ± 4.1) | 39.6 | 39.5–42.3 (40.7 ± 1.1) | 40.6 | 38.5–44.6 (41.4 ± 1.8) |
| Distance between nostrils on the same side | 5.6–9.6 (6.8 ± 1.3) | 5.2–10.0 (6.9 ± 1.4) | 6.2 | 6.2–8.6 (7.2 ± 0.9) | 5.9 | 4.6–5.9 (5.3 ± 0.4) |
| Orbit diameter | 10.6–16.3 (14.2 ± 1.5) | 9.0–14.2 (12 ± 1.7) | 12.8 | 11.1–18.0 (15.6 ± 2.7) | 14.9 | 10.0–19.7 (14.1 ± 2.4) |
| Head width | 51.1–61.1 (55.8 ± 2.8) | 44.2–62.6 (54.2 ± 6.8) | 51.2 | 50.1–56.8 (52.6 ± 2.5) | 54.5 | 50.4–58.2 (53.5 ± 2.3) |
| Distance between anterior nostrils | 23.6–27.6 (25.1 ± 1.4) | 16.5–31.1 (22.7 ± 4.7) | 21.7 | 21.7–26.2 (23.3 ± 1.7) | 24.8 | 20.5–27.4 (24.5 ± 2.1) |
| Interorbital width | 35.5–43.7 (40.4 ± 2.5) | 29.8–52.1 (39.5 ± 8.3) | 39.1 | 38.1–43.8 (40.4 ± 2.1) | 35.7 | 34.8–43.4 (38.5 ± 3) |
| Counts | ||||||
| Number of scales around caudal peduncle | 20 | 18 (14–22) | 16 | 16 | 16 | 16 (12–16) |
| Number of scales along caudal peduncle | 20 (19–24) | 18 (17–22) | 22 | 20 (19–24) | 18 | 17 (17–20) |
| Number of scales along lateral line | 79 (76–80) | 70 (66–78) | 84 | 78 (78–85) | 75 | 74 (69–88) |
| Teeth on upper jaw | 7 (6–7) | 7 (6–7) | 7 | 7 (6–8) | 6 | 7 (6–7) |
| Teeth on lower jaw | 8 (8–10) | 8 (6–10) | 9 | 10 (8–12) | 3 | 6 (3–8) |
| Number of pectoral‐fin rays | 10 | 10 | 10 | 10 | 10 | 10 |
| Number of pelvic‐fin rays | 6 | 6 | 6 | 6 | 6 | 6 |
| X‐ray counts | ||||||
| Number of specimens | 8 | 5 | _ | 7 | _ | 12 |
| First anal‐fin ray to first dorsal‐fin ray | 3 (3–5) | 3 (3–4) | 5 | 5 (5–6) | 5 | 6 (5–7) |
| Total vertebrae | 43 (42–43) | 40 (40–41) | 46 | 47 (46–47) | 43 | 43 (42–44) |
| Pre‐caudal vertebrae | 19 (18–19) | 18 | 21 | 20 (20–21) | 19 | 19 (18–19) |
| Caudal vertebrae | 24 (23–25) | 22 (22–23) | 25 | 27 (25–27) | 24 | 24 (24–25) |
| Vertebrae before the first dorsal‐fin radial | 20 (20–21) | 19 (19–20) | 22 | 23 (22–24) | 21 | 20 (19–21) |
| Vertebrae before the first anal‐fin radial | 19 (19–20) | 18 (18–19) | 21 | 21 (21–22) | 19 | 19 (18–19) |
| Dorsal‐fin rays | 19 (17–21) | 19 | 23 | 23 (22–24) | 23 | 24 (22–25) |
| Anal‐fin rays | 24 (22–25) | 22 (22–24) | 30 | 28 (28–30) | 27 | 27 (26–29) |
|
|
|
|
|
|
|
|
| |
|---|---|---|---|---|---|---|---|---|
| Number of specimens | – | 23 | – | 7 | – | 19 | – | 7 |
| Total length | 94.9 | 41.8–130.3 (68.5 ± 23.5) | 71.7 | 38.3–76.0 (58.5 ± 13.7) | 114.8 | 57.3–114.8 (81 ± 16.5) | 166 | 34.7–166 (103.3 ± 63.2) |
| Standard length | 84.5 | 37.0–118.1 (61.5 ± 21) | 65.1 | 33.9–69.2 (52.6 ± 12.6) | 102.2 | 49.2–102.2 (72.1 ± 14.8) | 149.2 | 30.7–149.2 (92.4 ± 56.9) |
| Head length | 20.4 | 9.3–25.7 (14.2 ± 4.3) | 14.9 | 8.4–14.9 (12.1 ± 2.5) | 23.6 | 11.7–23.6 (16.7 ± 3.6) | 31.7 | 7.6–32.2 (20.5 ± 11.8) |
| Pre‐dorsal length | 64.5 | 62.7–66.7 (64.4 ± 0.7) | 64.5 | 63.2–67.0 (64.9 ± 1.5) | 65.3 | 62.7–67.8 (65.3 ± 1.3) | 64.6 | 62.2–66.3 (64.3 ± 1.5) |
| Pre‐anal length | 61.4 | 58.1–65.3 (60.7 ± 1.6) | 59 | 57.6–62.8 (59.7 ± 1.6) | 60.7 | 59.0–62.2 (60.9 ± 1.1) | 60.5 | 58.1–64.0 (61.1 ± 2.2) |
| Pre‐pelvic length | 42.6 | 39.8–44.6 (42.3 ± 1.2) | 41.3 | 40.3–44.1 (41.4 ± 1.3) | 41 | 40.0–44.0 (42 ± 0.9) | 41.1 | 38.6–46.1 (42.4 ± 2.6) |
| Pre‐pectoral length | 24.9 | 23.5–28.8 (26.3 ± 1.5) | 23.4 | 23.4–29.1 (26.8 ± 2) | 24.6 | 22.2–28.3 (26.1 ± 1.7) | 22.6 | 22.5–30.5 (25.8 ± 3.4) |
| Pectoral fin to anal fin | 36.6 | 20.1–41.7 (35.8 ± 1.8) | 36.4 | 33.2–36.4 (34.9 ± 1.4) | 37.8 | 33.6–38.2 (36 ± 1.3) | 39.2 | 31.9–40.8 (36.5 ± 3.5) |
| Pelvic fin to anal fin | 18.7 | 16.5–22.2 (18.4 ± 1.2) | 18 | 18.0–18.6 (18.2 ± 0.2) | 20.5 | 17.3–20.9 (18.9 ± 0.9) | 20.4 | 16.4–21.1 (19.3 ± 1.7) |
| Pectoral fin to dorsal fin | 40.9 | 39.0–45.3 (41.2 ± 1.2) | 41.8 | 38.2–42.7 (40.7 ± 1.7) | 43.9 | 39.2–43.9 (41.6 ± 1.3) | 44.3 | 38.8–45.0 (41.9 ± 2.5) |
| Dorsal fin to pelvic fin | 29.9 | 29.2–32.8 (30.8 ± 1) | 31.1 | 29.3–31.1 (30.3 ± 0.8) | 33.4 | 30.1–33.4 (31.8 ± 1.1) | 34.4 | 30.3–34.4 (32.3 ± 1.6) |
| Dorsal‐fin base length | 18.2 | 14.4–21.4 (17 ± 1.5) | 15 | 13.2–15.1 (14.5 ± 0.7) | 15.6 | 13.6–17.0 (15.7 ± 0.9) | 18.5 | 15.2–18.5 (17.1 ± 1.2) |
| Anal‐fin base length | 21.3 | 16.1–22.2 (19.6 ± 1.4) | 18.5 | 16.6–19.9 (18.5 ± 1) | 20.5 | 17.4–21.1 (19.1 ± 1) | 21 | 17.3–21.9 (20.4 ± 1.6) |
| Dorsal fin to caudal fin | 39.5 | 33.0–40.3 (37.4 ± 1.5) | 37.9 | 35.7–39.3 (37.2 ± 1.2) | 37.3 | 36.1–39.9 (37.8 ± 1.1) | 38.6 | 36.0–41.6 (39 ± 1.5) |
| Caudal peduncle length | 19.5 | 18.1–23.4 (20.9 ± 1.3) | 21.5 | 21.5–25.3 (23.4 ± 1.5) | 21.9 | 19.8–23.7 (22.2 ± 1.1) | 19.1 | 19.1–22.1 (20.9 ± 1.4) |
| Caudal peduncle depth | 5.2 | 5.0–6.8 (6.1 ± 0.6) | 6.2 | 5.9–6.2 (6.1 ± 0.1) | 7 | 6.3–7.9 (7.1 ± 0.4) | 7.1 | 6.7–7.5 (7.1 ± 0.3) |
| Pectoral‐fin length | 17.7 | 15.0–21.7 (17.9 ± 1.5) | 17.1 | 16.2–18.4 (17.6 ± 0.8) | 16 | 15.2–21.3 (17.6 ± 1.6) | 19.7 | 17.4–21.0 (19.3 ± 1.3) |
| Pectoral fin to pelvic fin | 18.9 | 16.0–20.8 (17.9 ± 1.1) | 18.2 | 14.4–18.9 (17.3 ± 1.5) | 17.7 | 16.1–19.0 (17.8 ± 0.7) | 20.9 | 16.0–21.8 (18.6 ± 2.4) |
| Body depth | 22.6 | 19.8–22.8 (21.8 ± 0.8) | 21.3 | 20.4–21.8 (20.9 ± 0.5) | 23 | 21.0–23.9 (22.7 ± 0.8) | 21.6 | 21.6–24.3 (23 ± 1) |
| Mid body depth | 22 | 16.8–22.2 (20.6 ± 1.2) | 22.6 | 18.7–22.6 (20.3 ± 1.3) | 22.8 | 20.8–24.6 (22.6 ± 0.9) | 24.3 | 20.0–24.3 (22.1 ± 1.6) |
| Head length | 24.2 | 20.8–26.0 (23.4 ± 1.5) | 22.9 | 21.5–24.7 (23.2 ± 1.1) | 23 | 21.6–24.9 (23.1 ± 1) | 21.3 | 21.3–25.4 (22.9 ± 1.6) |
| Length of the snout | 38.7 | 33.4–46.7 (40.9 ± 3.1) | 45.5 | 42.6–50.2 (46.5 ± 2.5) | 43.2 | 41.1–53.5 (46.9 ± 3.9) | 45.9 | 38.0–51.0 (42.5 ± 4.6) |
| Distance between nostrils on the same side | 5.4 | 5.4–10.2 (7.7 ± 1.2) | 9.5 | 7.5–12.7 (9.6 ± 1.6) | 7.9 | 7.5–11.8 (8.9 ± 1.2) | 6.7 | 6.7–11.1 (8 ± 1.6) |
| Orbit diameter | 9.1 | 8.2–15.8 (11.6 ± 1.9) | 13.1 | 8.6–13.1 (11.3 ± 1.7) | 13.3 | 10.5–18.6 (13.8 ± 2.2) | 11.1 | 10.9–15.0 (12.6 ± 1.5) |
| Head width | 50.7 | 50.7–64.7 (56.5 ± 3.3) | 60.3 | 55.3–63.1 (59.9 ± 2.6) | 61.1 | 55.4–65.0 (59.8 ± 2.9) | 60.7 | 54.5–67.2 (61.1 ± 4.5) |
| Distance between anterior nostrils | 19.6 | 19.6–30.2 (26 ± 3.1) | 29.9 | 26.3–29.9 (28.6 ± 1.3) | 28.3 | 24.3–31.8 (28.4 ± 2.1) | 27.8 | 24.2–29.7 (26.9 ± 1.9) |
| Interorbital width | 33.6 | 33.6–50.0 (43.7 ± 4.2) | 51.6 | 46.4–51.6 (49.3 ± 1.6) | 48.9 | 41.7–54.4 (47.9 ± 3) | 46 | 41.9–52.0 (45 ± 3.8) |
| Counts | ||||||||
| Number of scales around caudal peduncle | 20 | 20 (16–20) | 16 | 16 | 20 | 20 (18–20) | 16 | 16 |
| Number of scales along caudal peduncle | 20 | 20 (16–23) | 22 | 22 (21–23) | 21 | 21 (18–22) | 16 | 18 (16–20) |
| Number of scales along lateral line | 76 | 78 (64–86) | 82 | 82 (82–84) | 83 | 71 (71–83) | 78 | 72 (72–78) |
| Teeth on upper jaw | 7 | 7 (6–7) | 7 | 7 (7–8) | 7 | 7 (4–8) | 7 | 7 (5–9) |
| Teeth on lower jaw | 8 | 8 (6–10) | 9 | 8 (8–9) | 8 | 8 (4–10) | 8 | 8 (8–9) |
| Number of pectoral‐fin rays | 10 | 10 | 10 | 10 | 10 | 10 | 10 | 10 (9–10) |
| Number of pelvic‐fin rays | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 |
| X‐ray counts | ||||||||
| Number of specimens | – | 7 | – | 3 | – | 14 | – | 3 |
| First anal‐fin ray to first dorsal‐fin ray | 5 | 5 (4–5) | 7 | 7 (6–7) | 6 | 6 (6–7) | 4 | 4 (3–4) |
| Total vertebrae | 41 | 41 (41–42) | 43 | 43 | 41 | 41 (41–42) | 42 | 42 |
| Pre‐caudal vertebrae | 19 | 19 (18–19) | 20 | 19 (18–20) | 18 | 18 (17–19) | 18 | 18 (17–18) |
| Caudal vertebrae | 22 | 22 (22–24) | 23 | 24 (24–25) | 23 | 23 (22–25) | 24 | 24 (24–25) |
| Vertebrae before the first dorsal‐fin radial | 21 | 20 (20–21) | 21 | 22 (21–22) | 20 | 20 (19–21) | 20 | 20 (19–20) |
| Vertebrae before the first anal‐fin radial | 19 | 19 (19–20) | 20 | 19 (18–20) | 18 | 19 (18–20) | 18 | 19 (18–19) |
| Dorsal‐fin rays | 18 | 19 (18–20) | 20 | 20 (18–20) | 18 | 19 (17–20) | 20 | 20 (19–20) |
| Anal‐fin rays | 22 | 23 (22–24) | 25 | 23 (23–25) | 22 | 24 (20–25) | 23 | 23 (23–24) |
- —National Research Foundation10.13039/501100001321
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFish Biology and Ecology Studies · Fish biology, ecology, and behavior · Fish Ecology and Management Studies
INTRODUCTION
1
The Mormyridae Bonaparte 1831 is the most diverse family within the order Osteoglossiformes, comprising 227 valid species in 22 genera (Fricke et al., 2024; Nelson, 2016). Fishes of this family are endemic to Africa where they are found in several rivers across the continent, with the highest diversity occurring in Central and West Africa (Peterson et al., 2022; Roberts, 1975; Skelton, 2001). These fishes are known for their ability to produce and detect weak electric organ discharges (EODs), a key evolutionary innovation that they use for electrolocation and electrocommunication (Arnegard et al., 2010; Carlson & Arnegard, 2011; Feulner et al., 2007; Kramer, 1990; Lavoué et al., 2003; Lissmann, 1958; Sullivan et al., 2000). Evidence from several mormyrid species has shown that EODs diverged more rapidly than other ecomorphological characters, thereby potentially forming early reproductive barriers between species (Arnegard et al., 2010; Arnegard & Hopkins, 2003; Carlson & Arnegard, 2011; Feulner et al., 2008, 2009). This makes the delimitation of species boundaries among some closely related mormyrid species using only traditional morphological approaches challenging. Consequently, the species‐level taxonomy in Mormyridae remains poorly resolved, as shown by the presence of taxonomic conflicts in most genera of this family (Peterson et al., 2022). For example, in southern Africa, the Mormyridae are represented by eight genera with a total of 27 species, most of which are thought to be broadly distributed across multiple river systems (Marshall, 2011; Skelton, 2001). Recent studies implementing integrated taxonomic approaches have identified several undescribed species that were previously included within a single widely distributed species, highlighting the need for further reviews of the taxonomy of the mormyrid fishes within this region (Kramer et al., 2003, 2007, 2012, 2016; Kramer & Swartz, 2010; Kramer & van der Bank, 2000; Kramer & van der Bank, 2011; Kramer & Wink, 2013; Maake et al., 2014).
Advances in molecular approaches have facilitated the resolution of mormyrid taxonomy. For example, five species of slender stonebashers from southern Africa, including Heteromormyrus ansorgii (Boulenger, 1905) and Heteromormyrus pappenheimi (Boulenger, 1910), both occurring in the Kwanza River system; Heteromormyrus tavernei (Poll, 1972) from the upper Lualaba and Lufira rivers; Heteromormyrus longilateralis (Kramer & Swartz, 2010), a species endemic to the Kunene River system; and Heteromormyrus szaboi (Kramer et al., 2004) found throughout the upper Zambezi River system, which were previously included in the genera Hippopotamyrus Pappenheim, 1906 and Brienomyrus Taverne, 1971, were transferred into the genus Heteromormyrus Steindachner, 1866. This transfer of species was based on mitogenomic data demonstrating the monophyly of the slender stonebashers from southern African (all but H. pappenheimi, which was not sequenced) with the holotype of Heteromormyrus pauciradiatus Steindachner, 1866, while showing their polyphyletic relationship with the type species of Hippopotamyrus from Cameroon (Sullivan et al., 2022).
Several previous studies on H. ansorgii indicated that it is a species complex concealing undocumented diversity across its disjunct distribution range (Chakona et al., 2018; Kramer et al., 2004; Kramer & Swartz, 2010; van der Bank & Kramer, 1996). This species, originally described based on two specimens collected from an uncertain locality broadly identified as ‘between Benguella and Bihé’, in Angola, was subsequently reported from several southern African river systems, including the Kwanza, Kunene, Okavango, Zambezi, Pungwe and Buzi rivers (Bell‐Cross & Minshull, 1988; Chakona et al., 2018; Kadye et al., 2008; Marshall, 2011; Skelton, 2001). Mutizwa et al. (2021), through detailed analysis of comprehensive molecular and morphological data, identified 10 distinct lineages within the H. ansorgii complex, and resolved that the type locality of this species was likely to be in the Kwanza River system. Five of the lineages identified by Mutizwa et al. (2021) as K1*–*K5 are distributed in the Kwanza River system. Subsequently, Sullivan et al. (2022) provided mitogenomic evidence that indicated that one of the Kwanza River lineages (K4) represents H. pauciradiatus. However, it remains unclear which of the other lineages identified from the Kwanza River represent the two species that were described from this system, H. ansorgii and H. pappenheimi.
The purpose of this study was to undertake detailed morphological examination of the lineages within the H. ansorgii complex identified by Mutizwa et al. (2021) to identify diagnostic characters that support their recognition and description as new species. In the Kwanza River system, there are five lineages within the H. ansorgii complex (K1–K5), and three names are available (H. pauciradiatus, H. ansorgii and H. pappenheimi). Sullivan et al. (2022) provided evidence showing that the K4 lineage represents H. pauciradiatus. For the present study, our first objective was to determine if any of the identified lineages from this system are conspecific with either H. ansorgii or H. pappenheimi. The second objective was to provide redescriptions of H. ansorgii s.s. and H. pauciradiatus, and describe six new species: two from the Kwanza River system, and one species each from the Okavango, Ruo, Pungwe and Buzi river systems. Two lineages included in Mutizwa et al. (2021), Heteromormyrus sp. ‘K2’ from the Kwanza River and Heteromormyrus sp. ‘UZ1’ from the upper Zambezi River, were not included in the present study because they were only represented by juvenile specimens. This study builds on the previous work that highlighted the hidden diversity in the genus Heteromormyrus (Chakona et al., 2018; Kramer et al., 2004; Kramer & Swartz, 2010; Mutizwa et al., 2021) by providing formal names for most of the known lineages in southern Africa, thus ensuring their inclusion in future studies and conservation planning for this region.
MATERIALS AND METHODS
2
2.1
This study included specimens obtained from the National Research Foundation – South African Institute for Aquatic Biodiversity (NRF‐SAIAB), in Makhanda, South Africa. Ethical clearance for this research was granted by the Rhodes University Animal Ethics Committee (reference number: 2700). The sampling approaches were approved by the NRF‐SAIAB Animal Ethics Committee (reference number: 2014/03). Permission to survey localities in the Eastern Highlands of Zimbabwe was approved by the Zimbabwe Parks and Wildlife Authority.
Examined material
2.2
The present study was based on specimens collected from across the range of the H. ansorgii species complex during surveys conducted between 1999 and 2017. Most specimens were collected using a combination of seine netting and electrofishing. Specimens were euthanized with clove oil, and a small piece of muscle tissue or fin clip for genetic analysis was dissected from some specimens and placed in a tube with 99% ethanol and stored at −80°C for long‐term storage. Voucher specimens were fixed in formaldehyde before being stored in 70% ethanol for long‐term storage. Voucher specimens and tissue samples were deposited into the National Fish Collection and the National Biobank, both at NRF‐SAIAB, Makhanda, South Africa.
This study examined 174 specimens from 50 collection sites. These specimens were obtained from NRF‐SAIAB, the Royal Museum for Central Africa in Tervuren, Belgium, (RMCA) and the Natural History Museum (BMNH) in London, UK. The specimens represented different H. ansorgii lineages identified by Mutizwa et al. (2021): Heteromormyrus sp. ‘Buzi’ (n = 7) from the Buzi River system, Heteromormyrus sp. ‘Pungwe’ (n = 18) from the Pungwe River system, Heteromormyrus sp. ‘Ruo’ (n = 7) from the Rou River, lower Zambezi system, Heteromormyrus sp. ‘OK’ (n = 36) from the Okavango River system, Heteromormyrus sp. ‘K1’ (n = 8), Heteromormyrus sp. ‘K3’ (n = 16) and Heteromormyrus sp. ‘K5’ (n = 23) from the Kwanza River system. Comparative material included H. ansorgii syntypes (n = 2), H. szaboi (n = 13), H. longilateralis (n = 11), H. pauciradiatus (n = 25) and H. pappenheimi syntypes (n = 8). In this study, we examined and measured the two syntypes of H. ansorgii. However, we could not access the syntypes of H. pappenheimi. We therefore made comparisons based on meristic data from X‐ray radiographs and the original description by Boulenger (1910). Similarly, data for H. tavernei were obtained from the original description by Poll (1972). Detailed information of the specimens examined is presented in the material examined section of each species account.
Morphological analyses
2.3
Measurements and external counts were done on the left side of each specimen. Digital Vernier callipers were used for measurements to the nearest 0.1 mm following Mutizwa et al. (2021), with the addition of six osteological characters obtained from X‐ray radiographs and the exclusion of three morphometric characters (distance from upper lip to centre of orbit, distance from mouth edge to centre of orbit, distance from mental lobe to centre of orbit) that were difficult to constantly measure. The study examined 26 point‐to‐point measurements and 15 meristic counts (Figure 1a,b). These measurements and meristic counts included total length (TL), standard length (SL), pre‐dorsal length (PDL), pre‐anal length (SAF), pre‐pelvic length (SPF), pre‐pectoral length (PFS), pectoral fin to anal fin (PA), pelvic fin to anal fin (PFA), pectoral fin to dorsal fin (PFD), dorsal fin to pelvic fin (DFP), dorsal‐fin base length (LD), anal‐fin base length (LA), dorsal fin to caudal fin (pD), caudal peduncle length (CPL), caudal peduncle depth (CPD), pectoral‐fin length (LPF), pectoral fin to pelvic fin (PPF), body depth (BD), mid body depth (mBD), head length (HL), length of snout (LSo), inter‐nostril distance on the same side (Na), orbit diameter (OD), head width (HW), distance between anterior nostrils (IN), interorbital width (IOW), dorsal‐fin rays (nD), anal‐fin rays (nA), number of anal‐fin rays between the leading ray and the one directly below the leading dorsal‐fin ray (AD), pelvic‐fin rays (nP), pectoral‐fin rays (PFc), number of scales around the caudal peduncle (SPc) and number of scales along the caudal peduncle (cSLS), the lateral‐line scales (SLS), upper‐jaw teeth (UJ), lower‐jaw teeth (LJ) and total vertebrae (TV), pre‐caudal vertebrae (PC), caudal vertebrae (CV), number of vertebrae before the first dorsal‐fin radial (FDR) and number of vertebrae before the first anal‐fin radial (FAR). Vertebrae, dorsal‐ and anal fin‐rays were counted from radiographs taken using the Inspex 20i Digital X‐ray Imaging System (Kodex Inc., New Jersey, NJ, USA) at NRF‐SAIAB, RMCA and BMNH. Vertebrae counts did not include the demi‐centrum fused to the hypural plate. In mormyrids, the first two dorsal‐ and anal‐fin rays are unbranched, unsegmented and so small that they are difficult to see without a radiograph. These two unbranched and unsegmented rays are followed by a single unbranched, segmented ray (the first long ray) followed by numerous branched, segmented rays. The last dorsal‐ and anal‐fin ray is usually branched all the way to its base. In the present study, all rays, both unbranched and branched, were included in the counts. The last ray, which is branched to its base, was counted as one.
(a) Illustration of the morphological characters examined in this study. Explanation for the morphometric characters provided in the materials and methods section. (b) Radiograph showing how osteological counts were done in the present study: pre‐caudal vertebrae (PC), caudal vertebrae (CV), vertebra before the first dorsal‐fin radial (FDR), vertebra before the first anal‐fin radial (FAR), unbranched fin rays, branched fin rays and first anal‐fin ray to first dorsal‐fin ray (AD).
RESULTS
3
Meristic data of the southern African species of Heteromormyrus
3.1
Principal component analysis of the meristic data showed separation between most of the Heteromormyrus species. The H. ansorgii and H. pappenheimi syntypes overlapped, and they were negatively associated with the first principal component (PCI), separating them from H. szaboi, which was more negatively associated with this axis, as well as Heteromormyrus sp. ‘K1’ and Heteromormyrus sp. ‘Ruo’ that were positively associated with this axis (Figure 2). The PCI accounted for 35.7% of the observed variance, and it was positively associated with the total number of vertebrae (Table 1). Consistent with the pattern shown by PCI, H. ansorgii (42–43) and H. pappenheimi (42–43) have more vertebrae than H. szaboi (40–41) but have fewer than Heteromormyrus sp. ‘K1’ (46–47) (Figure 3a).
Principal component analysis of the meristic data of the Heteromormyrus lineages from southern Africa showing components 1 and 2: pectoral‐fin rays (PFc), number of scales around the caudal peduncle (SPc), number of scales along the caudal peduncle (cSLS), lateral‐line scales (SLS), first anal‐fin ray to first dorsal‐fin ray (AD), pre‐caudal vertebrae (PC), caudal vertebrae (CV), number of vertebrae before the first dorsal‐fin radial (FDR), number of vertebrae before the first anal‐fin radial (FAR), total vertebrae (TV), dorsal‐fin rays (nD), anal‐fin rays (nA), lower jaw teeth (LJ) and upper jaw teeth (UJ). Heteromormyrus ansorgii syntype, Heteromormyrus szaboi , Heteromormyrus longilateralis , Heteromormyrus pauciradiatu , H. pappenheimi syntype, Heteromormyrus sp. ‘Buzi’, Heteromormyrus sp. ‘Pungwe’, Heteromormyrus sp. ‘OK’, Heteromormyrus sp. ‘Ruo’, Heteromormyrus sp. ‘K1’ , Heteromormyrus sp. ‘K3’, Heteromormyrus sp. ‘K5’.
Boxplots of meristic characters that distinguished some of the Heteromormyrus lineages and species from southern Africa; the colours are consistent for each lineage or species.
The H. ansorgii and H. pappenheimi syntypes overlapped and were negatively associated with the second principal component (PCII), separating them from H. longilateralis, Heteromormyrus sp. ‘Pungwe’, Heteromormyrus sp. ‘K1’, Heteromormyrus sp. ‘Ruo’ and Heteromormyrus sp. ‘OK’, which were positively associated with this axis (Figure 2). The PCII accounted for 20.0% of the observed variance, and it was positively associated with the number of scales around the caudal peduncle (Table 1). Consistent with the pattern shown by PCII, H. pappenheimi, a species with a low number of scales around the caudal peduncle (12–14), is separated from H. pauciradiatus (16–18), H. tavernei (20–22), H. longilateralis (20), Heteromormyrus sp. ‘Pungwe’ (18–20), Heteromormyrus sp. ‘K1’ (16), Heteromormyrus sp. ‘OK’ (16–20), Heteromormyrus sp. ‘Ruo’ (16) and Heteromormyrus sp. ‘Buzi’ (16, Figure 3b). Similarly, the scales around the caudal peduncle separate H. ansorgii (16) from H. longilateralis (18–20) and Heteromormyrus sp. ‘Pungwe’ (18–20, Figure 3b). Additionally, the number of anal‐fin rays between the leading ray and the one directly below the leading dorsal‐fin ray in the H. ansorgii (6) and H. pappenheimi (6–9) syntypes distinguish them from H. longilateralis (3–5), H. szaboi (3–4), Heteromormyrus sp. ‘OK’ (4–5) and Heteromormyrus sp. ‘Buzi’ (3–4, Figure 3c). The number of dorsal‐fin rays distinguish H. ansorgii syntypes (18) from the H. pappenheimi (21–23) syntypes, Heteromormyrus sp. ‘K1’ (22*–24) and Heteromormyrus sp. ‘K5’ (22–*25, Figure 3d).
Morphometric data of the southern African species of Heteromormyrus
3.2
Principal component analysis of the morphometric characters showed that the Heteromormyrus species largely overlapped. The H. ansorgii syntypes were negatively associated with PCI separating them from Heteromormyrus sp. ‘Ruo’, Heteromormyrus sp. ‘Pungwe’ and Heteromormyrus sp. ‘Buzi’, which were positively associated with this axis (Figure 4). The PCI accounted for 40.8% of the observed variance, and it was positively associated with the interorbital width (Table 2). Consistent with the pattern shown by PCI, the H. ansorgii syntypes (28.6%–32.9%SL) have a narrower interorbital width compared to Heteromormyrus sp. ‘Ruo’ (46.4%–51.6%SL), Heteromormyrus sp. ‘Pungwe’ (41.7%–54.4%SL) and Heteromormyrus sp. ‘Buzi’ (41.9%–52%SL, Figure 5a).
Principal component analysis of the morphometric data of the Heteromormyrus lineages from southern Africa showing components 1 and 2: pre‐dorsal length (PDL), pre‐anal length (SAF), pre‐pelvic length (SPF), pre‐pectoral length (PFS), pectoral fin to anal fin (PA), pelvic fin to anal fin (PFA), pectoral fin to dorsal fin (PFD), dorsal fin to pelvic fin (DFP), dorsal‐fin base length (LD), anal‐fin base length (LA), dorsal fin to caudal fin (pD), caudal peduncle length (CPL), caudal peduncle depth (CPD), pectoral‐fin length (LPF), pectoral fin to pelvic fin (PPF), body depth (BD), mid body depth (mBD), head length (HL), length of snout (LSo), distance between nostrils on the same side (Na), orbit diameter (OD), head width (HW), distance between anterior nostrils (IN) and interorbital width (IOW). Heteromormyrus ansorgii syntype, Heteromormyrus szaboi , Heteromormyrus longilateralis , Heteromormyrus pauciradiatus , Heteromormyrus sp. ‘Buzi’, Heteromormyrus sp. ‘Pungwe’, Heteromormyrus sp. ‘OK’, Heteromormyrus sp. ‘Ruo’, Heteromormyrus sp. ‘K1’, Heteromormyrus sp. ‘K3’, Heteromormyrus sp. ‘K5’ .
Boxplots of the morphometric characters that distinguished some of the Heteromormyrus lineages and species from southern Africa; the colours are consistent for each lineage or species.
The PCII accounted for 15.8% of the observed variance, and it was positively associated with the snout length (Table 2). There was minimal separation of the species along PCII; as a result snout length could only marginally separate the species examined in the study (Figure 4). The H. ansorgii syntypes (35.3%–38.8%HL) are distinguished by a shorter snout than H. szaboi (39.6%–50.8%HL), Heteromormyrus sp. ‘K1’ (39.5%–42.3%HL), Heteromormyrus sp. ‘Ruo’ (42.6%–50.2%HL) and Heteromormyrus sp. ‘Pungwe’ (41.1%–53.5%HL, Figure 5b).
Further comparison of morphometric characters, including caudal peduncle depth, distance between the nostrils on the same side, dorsal‐fin base length, anal‐fin base length and body depth, revealed that they were informative in separating the different species (Figure 5). The caudal peduncles of the H. ansorgii syntypes (6.0%*–6.1%SL) are narrower than H. pauciradiatus (7.3%–*9.2%SL), H. longilateralis (7.0%–8.2%SL), H. szaboi (6.4%–10.4%SL), Heteromormyrus sp. ‘Pungwe’ (6.3%–7.9%SL) and Heteromormyrus sp. ‘Buzi’ (6.7%–7.5%SL), while being deeper than Heteromormyrus sp. ‘K1’ (4.9%–5.6%SL, Figure 5c). The distance between nostrils on the same side distinguishes the H. ansorgii syntypes (6.8%–7.8%HL) from Heteromormyrus sp. ‘K5’ (4.6%–5.9%HL, Figure 5d). The dorsal‐fin base length of H. ansorgii syntypes (16.8%–16.9%SL) is shorter compared to H. longilateralis (17.2%–18.4%SL), Heteromormyrus sp. ‘K5’ (18.6%–21.3%SL) and Heteromormyrus sp. ‘K1’ (17.5%–19.7%SL), while being longer than that of Heteromormyrus sp. ‘Ruo’ (13.2%–15.1%SL, Figure 5e). The H. ansorgii syntypes (22.2%–22.4%SL) are distinguished by a shorter anal‐fin base length compared to Heteromormyrus sp. ‘Pungwe’ (17.4%–21.1%SL), Heteromormyrus sp. ‘Buzi’ (17.3%–21.9%SL) and Heteromormyrus sp. ‘Ruo’ (16.6%–19.9%SL, Figure 5f). A deeper body distinguishes H. pauciradiatus (23.1%–27.9%SL) from the H. ansorgii syntypes (19.9%–22.5%SL, Figure 5g).
Head shape, nostril position and body colouration
3.3
The Heteromormyrus species are reliably separated by distinct head shapes, nostril positions and colour patterns (Figure 6). Heteromormyrus sp. ‘K5’ have a flat dorsal head profile that forms an obtuse angle with the rest of the dorsal body profile and a relatively large snout (Figure 6a). Similarly, Heteromormyrus sp. ‘OK’ and H. longilateralis have a flat dorsal head profile that forms an obtuse angle with the rest of the dorsal body profile and a relatively small snout (Figure 6b). H. ansorgii, H. pappenheimi, H. tavernei, H. szaboi, Heteromormyrus sp. ‘Ruo’, Heteromormyrus sp. ‘Pungwe’, Heteromormyrus sp. ‘Buzi’, H. pauciradiatus and Heteromormyrus sp. ‘K1’ have a rounded dorsal head profile (Figure 6c). The nostrils of Heteromormyrus sp. ‘K5’ were aligned horizontally below the level of the orbit. In contrast, the anterior nostrils in the rest of the Heteromormyrus species are always positioned higher than the posterior nostrils, and both nostrils are in line with the orbit (Figure 6).
The head shapes of the Heteromormyrus species and lineages examined in this study. (a) Heteromormyrus sp. ‘K5’ (SAIAB 84645) has a straight dorsal head profile that forms an obtuse angle where it meets the rest of the dorsal body surface. The nostrils of Heteromormyrus sp. ‘K5’ were aligned horizontally below the level of the orbit, contrasting the other Heteromormyrus species/lineages, which have anterior nostrils that are always positioned higher than the posterior nostrils, and both nostrils are in line with the orbit; (b) Heteromormyrus sp. ‘OK’ (SAIAB 202504) similarly has a straight dorsal head profile that forms an obtuse angle where it meets the rest of the dorsal body surface; c. Heteromormyrus sp. ‘Pungwe’ (SAIAB 201071) has a rounded dorsal head profile that gently blends into the rest of the dorsal body surface. Scale bar = 1 cm.
Live and preserved specimens examined in this study show a similar underlying colour pattern with small differences between species. The dorsal surface of all species is darker than the ventral surface. They all have a dark vertical bar beginning at the origin of the dorsal fin and ending at the anal fin. There is a dark blotch present near the flexion point of the caudal fin, and in some species, there are additional dark vertical bars in the caudal peduncle. The species are separated into four groups based on the visibility of the markings in the caudal peduncle. The first group consists of species with barely any visible markings in the caudal peduncle that included Heteromormyrus sp. ‘K5’, Heteromormyrus sp. ‘Ruo’ and Heteromormyrus sp. ‘Buzi’ (Figure 7a). The second group consists of H. szaboi, H. longilateralis, Heteromormyrus sp. ‘Pungwe’ and Heteromormyrus sp. ‘OK’ that have a clearly visible dark blotch present near the flexion point of the caudal fin (Figure 7b). The third group consists of H. ansorgii, H. pauciradiatus and Heteromormyrus sp. ‘K1’; these species have a clearly visible dark blotch present near the flexion point of the caudal fin and a dark vertical bar just anterior of this blotch (Figure 7c). The fourth group includes specimens of Heteromormyrus sp. ‘K1’ that have a series of clearly visible thin curved vertical bars due to the presence of melanophores within the tissue over the vertical myosepta that were more conspicuous in the anterior portion of flank (Figure 7d). Similar thin curved vertical bars were also present in some H. longilateralis, Heteromormyrus sp. ‘K5’ and Heteromormyrus sp. ‘OK’ specimens although they were much less visible.
The range of colour patterns observed in Heteromormyrus lineages and species from southern Africa. (a) Heteromormyrus sp. ‘K5’ (SAIAB 84645) had barely any visible markings in the caudal peduncle. (b) Heteromormyrus sp. ‘K5’ (SAIAB 203161) had a clearly visible dark blotch present near the flexion point of the caudal fin. (c) Heteromormyrus sp. ‘K3’ (SAIAB 84790) had a clearly visible dark blotch present near the flexion point of the caudal fin and a dark vertical bar just anterior to this blotch. (d) Heteromormyrus sp. ‘K1’ (SAIAB 85039) had a series of clearly visible thin curved vertical bars that were more conspicuous in the anterior portion of the flank. Scale bar = 1 cm.
Summary of meristic and morphological data
3.4
The meristic data show that the H. ansorgii syntypes can be separated from all the species examined, except for H. pauciradiatus and Heteromormyrus sp. ‘K3’, using a combination of characters, including total number of vertebrae, number of scales around the caudal peduncle, number of dorsal‐fin rays and the number of anal‐fin rays between the leading ray and the one directly below the leading dorsal‐fin ray. The syntypes of H. pappenheimi can be separated from all the species, except for Heteromormyrus sp. ‘K3’ and Heteromormyrus sp. ‘K5’, based on the same meristic characters. Although no specimens of H. tavernei were examined in this study, Poll's (1972) original description of this species indicates that differences in the number of scales around the caudal peduncle (20–22) and the number of anal‐fin rays (22–23) distinguish this species from its congeners, except for H. longilateralis, Heteromormyrus sp. ‘OK’ and Heteromormyrus sp. ‘Pungwe’.
The morphometric data from species examined in this study show that the H. ansorgii syntypes can be separated from all the species examined, except Heteromormyrus sp. ‘K3’, using a combination of characters, including interorbital width, snout length, caudal peduncle depth, distance between the nostrils on the same side, dorsal‐fin base length, anal‐fin base length and body depth. The measurements of H. tavernei provided in Poll's (1972) original description suggest that it can be distinguished from all the species examined in this study by the combination of the interorbital width (24.7%–30.0%HL), snout length (24.7%–30.0%HL), caudal peduncle depth (6.9%–8.6%SL) and head length (25.8%–29.2%SL). Similarly, Boulenger's (1910) original description of H. pappenheimi suggests that this species has a deeper body (25.0%–27.3%TL) compared to Heteromormyrus sp. ‘K3’ (18.8%–21.5%TL) and Heteromormyrus sp. ‘K5’ (18.3%–21.7%TL).
Using a combination of meristic characters, morphometric characters, head shapes, nostril positions and variation in colour pattern, the present study was able to distinguish the H. ansorgii syntypes from the other two previously described species in the Kwanza River system, H. pauciradiatus, H. pappenheimi, as well as two candidate species, Heteromormyrus sp. ‘K1’ and Heteromormyrus sp. ‘K5’. However, the H. ansorgii syntypes cannot be separated from one of the candidate species in this system, Heteromormyrus sp. ‘K3’, based on any of these characters. Our results also show that the Heteromormyrus sp. ‘K3’ lineage is distinct from both H. pauciradiatus and H. pappenheimi. Based on these findings, we propose that the Heteromormyrus sp. ‘K3’ lineage is likely to be conspecific with H. ansorgii s.s. This species is herein redescribed based on two syntypes and 15 specimens (previously designated as Heteromormyrus sp. ‘K3’) collected between 2007 and 2008 from the Kwanza River system. H. pauciradiatus, a species that was described based on a specimen with a deformed caudal peduncle (Sullivan et al., 2022), is also redescribed in this study using 22 specimens collected between 2007 and 2008. None of the specimens examined in this study were morphologically consistent with H. pappenheimi. Based on consistent meristic and morphometric differences, six new species are described in the present study: Heteromormyrus sp. ‘K5’ as Heteromormyrus dolichorhynchus sp. nov., Heteromormyrus sp. ‘K1’ as Heteromormyrus angusticaudata sp. nov., Heteromormyrus sp. ‘OK’ as Heteromormyrus xanekweorum sp. nov., Heteromormyrus sp. ‘Ruo’ as Heteromormyrus chilembwei sp. nov., Heteromormyrus sp. ‘Pungwe’ as Heteromormyrus tangwenai sp. nov. and Heteromormyrus sp. ‘Buzi’ as Heteromormyrus ndauorum sp. nov.
TAXONOMIC ACCOUNTS
4
H. ansorgii (Boulenger 1905)
4.1
See Figure 8.
(a) Syntype of Heteromormyrus ansorgii 100.1 (BMNH 1905.5.29.62) mm standard length (SL), female, from between Benguella and Bihé, Angola, collected by WJ Ansorge. (Picture provided by Luis Moreira da Costa); (b) recently collected H. ansorgii female specimen 110.0 mm SL (SAIAB 84790) from the Kwanza River, Angola, southern Africa; (c) recently collected H. ansorgii male specimen 120.0 mm SL (SAIAB 84790) from the Kwanza River; (d) X‐ray radiograph of syntype; (e, f) live pictures of H. ansorgii. Scale bar =1 cm.
Marcusenius ansorgii Boulenger, 1905.
Hippopotamyrus ansorgii (Boulenger, 1905) new combination by Taverne, 1971.
H. ansorgii (Boulenger, 1905) new combination by Sullivan et al., 2022.
Heteromormyrus sp. ‘K3’ in Mutizwa et al. (2021).
Syntypes
4.1.1
BMNH 1905.5.29.62100.1 mm SL, between Benguella and Bihé, Angola, W. J. Ansorge. BMNH 1905.5.29.63, 98.5 mm SL, between Benguella and Bihé, Angola, W. J. Ansorge. Mutizwa et al. (2021) proposed that these specimens were likely collected from the Kwanza River system.
Non‐type specimens
4.1.2
SAIAB 84763, 1 specimen, 61.1 mm SL, Kwanza River at confluence with Kawa River, 9° 10′ 14″ S, 13° 21′ 59″ E, Kwanza River system, Angola, E. Swartz and A. Chakona, 18 October 2007, GenBank genseq‐3 COI: MW600862, S7 intron: MW756326, MW756327; SAIAB 85028, 3 specimens, 80.6*–89.1 mm SL, bridge on road east of Camacupa, Kwiva River, 11° 59′ 3″ S, 17° 43′ 21″ E, Kwanza River system, Angola, E. Swartz, 10 August 2008, BOLD genseq‐3 COI: SAFW466‐09, SAFW467‐09; SAIAB 85177; 3 specimens 79.1–142.6 mm SL, bridge between Cimanga and Capunda, Luando River, 10° 38′ 26″ S, 17° 25′ 6″ E, Kwanza River system, Angola, E. Swartz and A. Chakona, 23 August 2008, BOLD genseq‐3 COI: SAFW572‐09, SAFW577‐09, SAFW566‐09; SAIAB 84636, 4 specimens, 94.1–112 mm SL, Terra Nova, 09° 46′ 44″ S, 14° 31′ 24″ E, Kwanza River system, Angola, E. Swartz and A. Chakona, 9 October 2007, BOLD genseq‐3 COI: SAFW252‐08, SAFW250‐08; SAIAB 84790, 5 specimens 59.5–*138.6 mm SL, Posto 5, 09° 48′ 23″ S, 15° 24′ 30″ E, Kwanza River, Angola, E. Swartz and A. Chakona, 24 October 2007, GenBank genseq‐3 COI: MW600861.
Diagnosis
4.1.3
H. ansorgii has 14–16 scales around the caudal peduncle, distinguishing it from H. tavernei (20–22), H. longilateralis (20) and H. tangwenai sp. nov. (18*–20). The number of anal‐fin rays between the leading ray and the one directly below the leading dorsal‐fin ray distinguishes H. ansorgii (6–9) from H. longilateralis (3–5), H. szaboi (3–4), H. xanekweorum sp. nov. (4–5) and H. ndauorum (3–4). Fewer dorsal‐fin rays distinguish H. ansorgii (18–21) from H. angusticaudata sp. nov. (22–24) and H. dolichorhynchus sp. nov. (22–25). H. ansorgii is further distinguished from H. angusticaudata sp. nov. by having fewer total vertebrae 42–44 (vs. 46–47), fewer caudal vertebrae 24–25 (vs. 27–27) and fewer anal‐fin rays 23–27 (vs. 28–30). H. ansorgii is distinguished from H. chilembwei sp. nov. by having fewer scales along the caudal peduncle 16–19 (vs. 21–23) and fewer scales along the lateral line 68–77 (vs 82–84). H. ansorgii is distinguished from H. pauciradiatus by a slender caudal peduncle 6.0%–7.2%SL (vs. 7.3%–9.3%SL). A narrower body distinguishes H. ansorgii (18.8%–21.5%TL) from H. pappenheimi (25.0%–27.3%TL). Interorbital width further distinguishes H. ansorgii (28.6%–45.4%HL) from H. chilembwei sp. nov. (46.4%–51.6%HL). Snout length distinguishes H. ansorgii (34.6%–41.4%HL) from H. tavernei (24.7%–30.0%HL) and H. chilembwei sp. nov. (42.6%–50.2%HL). Head length further distinguishes H. ansorgii (19.4%–24.9%SL) from H. tavernei (25.8%–29.2%SL). H. ansorgii is further distinguished from H. szaboi by a greater distance between pectoral and dorsal fins 41.6%–46.0%SL (vs. 38.9%–*41.5%SL). Presence of a clearly visible dark blotch near flexion point of the caudal fin and a dark vertical bar on the caudal peduncle distinguishes H. ansorgii from H. dolichorhynchus sp. nov., H. chilembwei sp. nov. and H. ndauorum sp. nov. that lack any clearly visible marks on caudal peduncle; H. longilateralis, H. szaboi, H. tangwenai sp. nov. and H. xanekweorum sp. nov. have only a dark blotch near flexion point of the caudal fin.
Description
4.1.4
Morphometric proportions and meristics are summarised in Table 3, with meristic counts of the syntypes given in parentheses.
Rounded blunt snout, below eye level. Dorsal profile of head convex from snout to back of the head where it becomes gently inclined towards dorsal fin. Small sub‐terminal mouth in line with pectoral‐fin base. Small rounded chin swelling transitions into concave ventral profile of head. Round orbit. Anterior and posterior nostrils laterally positioned, closer to snout tip than opercular opening, anterior to and arranged horizontally in line with orbit. Anterior nostril positioned slightly higher than posterior nostril. Small gill opening with soft skin cover adjacent to pectoral‐fin base. Bicuspid teeth 5–8 (7) in upper jaw and 4–9 (4) in lower jaw.
Laterally compressed body with greatest width between gill covers. Slender fusiform body with greatest depth occurring between origin of dorsal and anal fins. Body tapers gently towards head but sharply from origins of both dorsal and anal fins to caudal fin. Caudal peduncle slender, same depth from its origin to roughly around half its length, gradually widening into two symmetrical lobes. Body covered with transparent membrane that becomes increasingly translucent to opaque towards head, dorsal and ventral surfaces in preserved specimens. Head without scales; rest of the body covered in small cycloid scales with reticulated striae. Lateral line originates above pectoral fin and extends to caudal peduncle. There are 68*–77 (70) scales along lateral line, 16–*19 (17) of them along caudal peduncle length and 14–16 (16) around caudal peduncle. Urogenital opening situated just anterior to origin of anal fin.
Elliptical pectoral fin with 9–11 (10) rays, extends just beyond origin of pelvic fin. Short pelvic fin with 6 (6) rays. The first two dorsal‐ and anal‐fin rays are unbranched, unsegmented and small. These two unbranched and unsegmented rays are followed by a single long unbranched and segmented ray, followed by numerous branched, segmented rays. The last dorsal and anal ray is usually branched all the way to its base. The first two dorsal‐ and anal‐fin rays are unbranched, unsegmented and small. Anal and dorsal fins set towards posterior of body. The last dorsal and anal ray is usually branched all the way to its base. Anal‐fin origin 6–9 (6) fin rays anterior to dorsal‐fin origin. Dorsal‐fin rays: 18*–21 (18); anal‐fin rays: 23–*27 (24). Caudal fin deeply forked with rounded lobes, with bases covered in scales; distance from caudal‐fin flexion point to caudal‐fin tips is roughly equal to caudal peduncle length.
See Figure 8. Live colour: body colour ranges from silver with a tinge of brown to a very dark brown. Dorsal surface usually darker than the ventral surface. Dark vertical bar originates from origin of dorsal fin. Dark blotch presents near the flexion point of the caudal fin and a dark vertical bar just anterior to this blotch. Fins brown, darker than the body in silver specimens.
Preserved in ethanol: body colour ranges from light to dark brown. Dorsal surface usually darker than the ventral surface. Dark vertical bar originates from origin of dorsal fin. Clearly visible dark blotch presents near the flexion point of the caudal fin and a dark vertical bar just anterior of this blotch. Fins brown like the body.
Determining the sex of adults can be done externally by examining their anal‐fin base. In males, the body wall is dorsally indented giving the dorsal margin of the anal fin a sigmoid curvature. In females, the body wall appears almost straight.
Total vertebrae: 42*–44 (42), pre‐caudal vertebrae: 18–19 (18), caudal vertebrae: 24–25 (24), number of vertebrae at first dorsal radial: 20–22 (20), number of vertebrae at first anal radial: 18–*20 (18).
Distribution
4.1.5
H. ansorgii is known from the lower, middle and upper sections of the Kwanza River mainstem and its tributary, the Lwando River, Angola (Figure 9). However, historical connectivity of these populations is likely to have been fragmented by the construction of three major hydroelectric dams, the Cambambe, Lauca and Capanda dams on the mainstem Kwanza River.
Distribution of the Heteromormyrus species described in this study. Heteromormyrus ansorgii , Heteromormyrus pauciradiatus , Heteromormyrus angusticaudata sp. nov. and Heteromormyrus dolichorhynchus sp. nov. are restricted to the Kwanza River. Heteromormyrus xanekweorum sp. nov. is restricted to the Okavango River. Heteromormyrus chilembwei sp. nov. is found in the Ruo River. Heteromormyrus tangwenai sp. nov. is endemic to the Pungwe River. Heteromormyrus ndauorum sp. nov. is endemic to the Buzi River.
Etymology
4.1.6
The species was named in honour of Dr. William John Ansorge, who made extensive collections of various animals, which include the syntypes of H. ansorgii from the Kwanza River system during his expedition in Angola from 1903 to 1909.
H. pauciradiatus Steindachner 1866
4.2
See Figure 10.
(a) Heteromormyrus pauciradiatus (SAIAB 85209), female 118.7 mm standard length (SL), from the Kwanza River; (b) H. pauciradiatus (SAIAB 85203), male 108.7 mm SL, from the Kwanza River; (c) X‐ray radiograph of holotype; (d, e) live pictures of H. pauciradiatus. Scale bar =1 cm.
Mormyrus (Heteromormyrus) pauciradiatus Steindachner, 1866.
Marcusenius pauciradiatus (Steindachner, 1866) new combination by Boulenger, 1898.
M. (Heteromormyrus) pauciradiatus (Steindachner, 1866) new subgeneric combination by Géry, 1968.
Pollimyrus pauciradiatus (Steindachner, 1866) new combination by Taverne, 1971.
Heteromormyrus pauciradiatus (Steindachner, 1866) new combination by Taverne, 1972.
Hippopotamyrus sp. ‘K4’ in Mutizwa et al. (2021).
Holotype
4.2.1
NMW 22417 lodged in the Naturhistorisches Museum Vienna, 100 mm TL, type locality only given as ‘Angola’, GenBank genseq‐1 mitogenome: ON533765.
Non‐type specimens
4.2.2
SAIAB 85209, 2 specimens, 88.0–115.0 mm SL, farm below Lucala 2 bridge, 9° 25′ 30″ S, 14° 42′ 0″ E, Kwanza River system, Angola, E. Swartz and A. Chakona, 25 August 2008, BOLD genseq‐3 COI: SAFW619‐09, GenBank genseq‐3 cyt b: MW600880, S7 intron: MW756330, MW756331; SAIAB 85203, 2 specimens 94.0–176.0 mm SL, farm below Lucala 2 bridge, 9° 25′ 30″ S, 14° 42′ 0″ E, Kwanza River system, Angola, E. Swartz and A. Chakona, 25 August 2008, BOLD genseq‐3 COI: SAFW599‐09, SAFW596‐09, GenBank genseq‐3 cyt b: MW600881, S7 intron: MW756368, MW756369; SAIAB 84683, 4 specimens 54.7–86.2 mm SL, N'dalatando farm, Lucala River, Kwanza River system, 9° 25′ 30″ S, 14° 42′ 0″ E, Kwanza River system, Angola, E. Swartz and A. Chakona, 11 October 2007, BOLD genseq‐3 COI: SAFW274‐08; SAIAB 85102, 1 specimen 79.2 mm SL, Lucala River bridge, 9° 16′ 8″ S, 15° 14′ 49″ E, Kwanza River system, Angola, E. Swartz and A. Chakona, 18 August 2008, BOLD genseq‐3 COI: SAFW493‐09; SAIAB 85120, 16 specimens, 36.3–94.4 mm SL, above Calandula Falls immediately above the major waterfall, Lucala River, 9° 4′ 26″ S, 16° 0′ 0″ E, Kwanza River system, Angola, E. Swartz and A. Chakona, 19 August 2008, BOLD genseq‐3 COI: SAFW518‐09, SAFW515‐09; SAIAB 85139, 1 specimen 97.1 mm SL, below Calandula Falls below the major waterfall, Lucala River, 9° 4′ 37″ S, 15° 59′ 59″ E, Kwanza River system, Angola, E. Swartz and A. Chakona, 19 August 2008, BOLD genseq‐3 COI: SAFW434‐08. SAIAB 84716, 1 specimen 65.0 mm SL, confluence of Kawa and Kwanza rivers, Kawa River, 09° 10′ 17″ S, 13° 22′ 5″ E, Kwanza River system, Angola, E. Swartz, 16 August 2007, GenBank genseq‐3 COI: MW600858; SAIAB 84708, 18 specimens 64.0–135.0 mm SL, Lucala 3, Lucala River, 09° 31′ 21″ S, 14° 23′ 10″ E, Kwanza River system, Angola, E. Swartz, 12 November 2007, BOLD genseq‐3 COI: MW600856, MW600857, GenBank genseq‐3 cyt b: MW600882, MW600883, S7 intron: MW756366, MW756367, MW756358, MW756359.
Diagnosis
4.2.3
A deep caudal peduncle distinguishes H. pauciradiatus (7.3%*–9.2%SL) from H. ansorgii (6.0%–7.2%SL), H. angusticaudata sp. nov. (4.9%–5.6%SL), H. xanekweorum sp. nov. (5.0%–6.8%SL) and H. chilembwei sp. nov. (5.9%–6.2%SL). A deep body further separates H. pauciradiatus (23.1%–27.9%SL) from H. xanekweorum sp. nov. (19.8%–22.8%SL) and H. chilembwei sp. nov. (20.4%–21.8%SL). H. pauciradiatus has 16–18 scales around the caudal peduncle that distinguish it from H. pappenheimi (12–14), H. tavernei (20–22) and H. longilateralis (20). Number of anal‐fin rays between the leading ray and the one directly below the leading dorsal‐fin ray in H. pauciradiatus (6–9) distinguishes it from H. longilateralis (3–5), H. szaboi (3–4), H. xanekweorum sp. nov. (4–5) and H. ndauorum sp. nov. (3–4). Fewer scales along the caudal peduncle distinguish H. pauciradiatus (14–18) from H. longilateralis (19–24), H. chilembwei sp. nov. (21–23) and H. angusticaudata sp. nov. (19–24). Fewer dorsal‐fin rays distinguish H. pauciradiatus (18–21) from H. angusticaudata sp. nov. (22–24) and H. dolichorhynchus sp. nov. (22–25). Fewer vertebrae also separate H. pauciradiatus (40–42) from H. angusticaudata sp. nov. (46–47) and H. chilembwei sp. nov. (43). Fewer anal‐fin rays further distinguish H. pauciradiatus (23–26) from H. angusticaudata sp. nov. (28–*30). Combination of a clearly visible dark blotch present near flexion point of the caudal fin and a dark vertical bar on caudal peduncle distinguishes H. pauciradiatus from H. dolichorhynchus sp. nov., H. chilembwei sp. nov. and H. ndauorum sp. nov. that do not have any clearly visible marks on caudal peduncle; H. longilateralis, H. szaboi, H. tangwenai sp. nov. and H. xanekweorum sp. nov. have only a dark blotch present near flexion point of the caudal fin.
Description
4.2.4
Morphometric proportions and meristics are summarised in Table 4.
Rounded blunt snout, below eye level. Dorsal head profile convex from snout to back of head where it becomes gently inclined towards dorsal fin. Small sub‐terminal mouth in line with pectoral‐fin base. Small rounded chin swelling transitions into the concave ventral profile of head. Round orbit. Anterior and posterior nostrils laterally positioned, closer to snout tip than opercular opening, anterior to and arranged horizontally in line with orbit. Anterior nostril positioned slightly higher than posterior nostril. Small gill opening with soft skin cover adjacent to pectoral‐fin base. Bicuspid teeth: 6–7 in upper jaw and 6–9 in lower jaw.
Laterally compressed body with greatest width between gill covers. Fusiform body with greatest depth occurring between origin of dorsal and anal fins. Body tapers gently towards head, while tapering sharply from origins of both dorsal and anal fins; the thin caudal peduncle maintains same depth from its origin to roughly around half its length, then widens into two symmetrical lobes. Body covered with transparent membrane that becomes increasingly translucent to opaque towards head, dorsal and ventral surfaces in preserved specimens. Head without scales; but rest of the body is covered by small cycloid scales with reticulated striae. Lateral line originates above pectoral fin and forms a straight line to caudal peduncle. There are 60–74 scales along lateral line, 16–18 along caudal peduncle and 14–18 around caudal peduncle. Urogenital opening situated adjacent to origin of anal fin.
Rounded pectoral fin with 10 rays, extending just beyond origin of pelvic fin. Short pelvic fin with 6 rays. The first two dorsal and anal‐fin rays are unbranched, unsegmented and small. These two unbranched and unsegmented rays are followed by a single long unbranched and segmented ray, followed by numerous branched, segmented rays. The last dorsal‐ and anal‐fin ray is usually branched all the way to its base. Anal and dorsal fins set towards posterior of body. Anal‐fin origin is 6–9 fin rays anterior to dorsal‐fin origin. Dorsal‐fin rays: 18–21; anal‐fin rays: 23–26. In both dorsal and anal fins, anterior fin rays increase in length up to fourth and fifth rays; subsequent rays get progressively shorter. Caudal fin deeply forked with rounded lobes with bases covered in scales; distance from caudal‐fin flexion point to caudal‐fin tips is roughly equal to caudal peduncle length.
See Figure 10. Live colour: body colour ranges from brown to grey. Dorsal surface usually darker than the ventral surface. Dark vertical bar originates from origin of dorsal fin. Dark vertical bar originates from origin of dorsal fin. Dark blotch present near the flexion point of the caudal fin; dark vertical bar often visible just anterior to the caudal‐fin flexion point. Fins brown. Preserved in ethanol: body colour ranges from light to dark brown. Dorsal surface usually darker than the ventral surface. Dark vertical bar originates from origin of dorsal fin. Dark blotch presents near the flexion point of the caudal fin; dark vertical bar often visible just anterior to the caudal‐fin flexion point. Fins brown.
Determining the sex of adults can be done externally by examining their anal‐fin base. In males, the body wall is dorsally indented giving the dorsal margin of the anal fin a sigmoid curvature. In females, the body wall appears almost straight.
Total vertebrae: 40–42, pre‐caudal vertebrae: 18–19, caudal vertebrae: 22–24, vertebrae at first dorsal radial: 20–21, vertebrae at first anal radial: 18–19.
Distribution
4.2.5
H. pauciradiatus was recorded from the mainstem channel of the Kwanza River and its north bank tributary, the Lucala River, in Angola (Figure 9).
H. dolichorhynchus sp. nov.
4.3
https://zoobank.org/NomenclaturalActs/d6a799cc-01cf-4175-a062-60931370a6d5; Figure 11.
(a) Holotype of Heteromormyrus dolichorhynchus sp. nov. (SAIAB 85219), female 151.0 mm standard length (SL), from the Kwanza River; (b) paratype of H. dolichorhynchus sp. nov. (SAIAB 85219), male 155.0 mm SL, from the Kwanza River; (c) H. dolichorhynchus sp. nov. (SAIAB 84645), male 151.0 mm SL, from the Kwanza River; (d) H. dolichorhynchus sp. nov. (SAIAB 84645), female 117.0 mm SL, from the Kwanza River; (e, f) live pictures of H. dolichorhynchus sp. nov.; (g) X‐ray radiograph of holotype. Scale bar =1 cm.
Hippopotamyrus sp. ‘K5’ in Mutizwa et al. (2021).
Heteromormyrus sp. ‘K5’ in Sullivan et al. (2022).
Holotype
4.3.1
SAIAB 246301, 151.0 mm SL, farm below Lucala 2 bridge, 9° 25′ 30″ S, 14° 42′ 0″ E, Kwanza River system, Angola, E. Swartz and A. Chakona, 25 August 2008. BOLD genseq‐1 COI: SAFW620‐09.
Paratypes
4.3.2
SAIAB 85219, 5 specimens 94.0–176.0 mm SL, farm below Lucala 2 bridge, 9° 25′ 30″ S, 14° 42′ 0″ E, Kwanza River system, Angola, E. Swartz and A. Chakona, 25 August 2008.
Non‐type specimens
4.3.3
SAIAB 246348, 1 specimen 86.0 mm SL, Lucala River bridge, 9° 16′ 8″ S, 15° 14′ 49″ E, Kwanza River, Angola, E. Swartz and A. Chakona, 18 August 2008, BOLD genseq‐3 COI: SAFW492‐09, GenBank genseq‐3 cyt b: MW600879, S7 intron: MW756356, MW756357; SAIAB 84645, 5 specimens 73.1*–*150.9 mm SL, Terra Nova, 9° 46′ 44″ S, 14° 31′ 24″ E, Kwanza River system, Angola, E. Swartz, 9 October 2007, BOLD genseq‐3 COI: SAFW251‐08, GenBank genseq‐3 S7 intron: MW756354, MW756355.
Diagnosis
4.3.4
H. dolichorhynchus sp. nov. has a unique elongate head which distinguishes it from its congeners. H. dolichorhynchus sp. nov. has nostrils aligned horizontally below the level of orbit readily distinguishing it from its congeners whose anterior nostrils are always positioned higher than the posterior nostrils, and both nostrils are horizontally in line with orbit. A higher number of dorsal‐fin rays further distinguish H. dolichorhynchus sp. nov. (22*–25) from most of its congeners, which include H. tangwenai sp. nov. (17–20), H. ndauorum sp. nov. (19–20), H. chilembwei sp. nov. (18–20), H. szaboi (19), H. xanekweorum sp. nov. (18–20), H. longilateralis (17–21), H. ansorgii (18–21), H. tavernei (17–20) and H. pauciradiatus (18–21). A higher number of anal‐fin rays further distinguish H. dolichorhynchus sp. nov. (26–29) from H. ndauorum sp. nov. (23–24), H. tangwenai sp. nov. (20–25), H. chilembwei sp. nov. (23–25), H. szaboi (22–24), H. xanekweorum sp. nov. (22–24) and H. longilateralis (22–25). The number of anal‐fin rays between the leading ray and the one directly below the leading dorsal‐fin ray distinguishes H. dolichorhynchus sp. nov. (5–7) from H. szaboi (3–4) and H. ndauorum sp. nov. (3–4). The total vertebrae separate H. dolichorhynchus sp. nov. (42–44) from H. angusticaudata sp. nov. (46–47) and H. szaboi (40–41). A higher number of caudal vertebrae separate H. dolichorhynchus sp. nov. (24–25) from H. szaboi (22–*23). A higher number of vertebrae before the first dorsal radial distinguish H. dolichorhynchus sp. nov. (18–19) from H. chilembwei sp. nov. (20). H. dolichorhynchus sp. nov. has 12–16 scales around the caudal peduncle, distinguishing it from H. tavernei (20–22), H. longilateralis (20) and H. tangwenai sp. nov. (18–20). H. dolichorhynchus sp. nov. (18.3%–21.7%TL) has a shallower body depth compared to H. pappenheimi (25.0%–27.3%TL). The lack of any clearly visible marks in the caudal peduncle further distinguishes H. dolichorhynchus sp. nov. from H. ansorgii, H. pauciradiatus and H. angusticaudata sp. nov. that have a clearly visible dark blotch present near the flexion point of the caudal fin and a dark vertical bar in the caudal peduncle; H. longilateralis, H. szaboi, H. xanekweorum sp. nov. and H. tangwenai sp. nov. have only a dark blotch present near the flexion point of the caudal fin.
Description
4.3.5
Morphometric proportions and meristics are summarised in Table 3. Meristic counts of the holotype are provided in parentheses.
Rounded blunt snout, below eye level. Straight dorsal head profile forms obtuse angle with the gently inclined dorsal body profile. Small sub‐terminal mouth placed below the level of pectoral‐fin base. Small rounded chin swelling transitions into deep concave ventral profile of head. Round orbit. Anterior and posterior nostrils laterally positioned, closer to snout tip than opercular opening, anterior to and arranged horizontally below orbit. Small gill opening with soft skin cover adjacent to pectoral‐fin base. Bicuspid teeth: 6*–7 (6) in upper jaw and 3–*8 (3) in lower jaw.
Laterally compressed body with greatest breadth occurring across gill covers. Fusiform body with greatest depth occurring between origin of dorsal and anal fins. Body tapers gently towards head, while tapering sharply from origins of both dorsal and anal fins to produce a relatively thin caudal peduncle. Caudal peduncle maintains same thin depth from its origin to roughly around half its length, then it gradually widens into two symmetrical lobes. Body covered with transparent membrane that becomes increasingly translucent to opaque towards head, dorsal and ventral surfaces in preserved specimens. Head without scales; but the rest of the body is covered with small cycloid scales with reticulated striae. Lateral line originates approximately above pectoral fin and forms a straight line to caudal peduncle. There are 69*–88 (75) scales along lateral line, 17–20 (18) along caudal peduncle and 12–*16 (16) around caudal peduncle. Urogenital opening situated adjacent to origin of anal fin.
Rounded pectoral fin with 10 rays, extending just beyond origin of pelvic fin. Short pelvic fin with 6 rays. The first two dorsal and anal‐fin rays are unbranched, unsegmented and small. These two unbranched and unsegmented rays are followed by a single long unbranched and segmented ray, followed by numerous branched, segmented rays. The last dorsal‐ and anal‐fin ray is usually branched all the way to its base. Anal and dorsal fins set towards posterior of body. Anal‐fin origin is 5–7 (5) fin rays anterior to dorsal‐fin origin. Dorsal‐fin rays: 22*–25 (23); anal‐fin rays: 26–*29 (27). In both dorsal and anal fins, anterior fin rays increase in length up to fourth and fifth rays; subsequent rays get progressively shorter. Caudal fin deeply forked with rounded lobes with bases covered in scales; distance from caudal‐fin flexion point to caudal‐fin tips is roughly equal to caudal peduncle length.
See Figure 11. Live colour: body colour ranges from silver to grey. Dorsal surface usually darker than the ventral surface. Dark vertical bar originates from origin of dorsal fin. Dark vertical bar originates from origin of dorsal fin, barely visible in darker specimens. Thin curved vertical bars on anterior portion of flank most conspicuous in slivery specimens. Fins grey, darker than the body in silvery specimens. Preserved in ethanol: body colour ranges from light to dark brown. Dorsal surface usually darker than ventral surface. Dark vertical bar originates from origin of dorsal fin, barely visible in darker specimens. Thin curved vertical bars conspicuous in the anterior portion of flank. No clearly visible marking in the caudal peduncle.
Determining the sex of adults can be done externally by examining their anal‐fin base. In males, the body wall is dorsally indented giving the dorsal margin of the anal fin a sigmoid curvature. In females, the body wall appears almost straight.
Total vertebrae: 42*–44 (43), pre‐caudal vertebrae: 18–19 (18), caudal vertebrae: 24–25 (25), vertebrae at first dorsal radial: 19–21(19), vertebrae at first anal radial: 18–*19 (18).
Distribution
4.3.6
H. dolichorhynchus was recorded from the mid‐reaches of the Kwanza River and its north bank tributary, the Lucala River, Angola (Figure 9).
Etymology
4.3.7
The name is the combination of the Greek words dolicho, meaning elongated or long, and rhynchos, meaning beak or snout. This is a reference to the unique elongate head of this species, which distinguishes it from its congeners.
Remarks
4.3.8
H. dolichorhynchus has two morphs, one that has a deeper caudal peduncle with 16 scales around the caudal peduncle (Figure 11a,b), and the other has a narrower caudal peduncle with 12 scales (Figure 11c,d). These two morphs are genetically and morphologically similar, except for the caudal peduncle depth and caudal scale counts. The specimens (SAIAB 85219, SAIAB 246348) with the deeper caudal peduncle and higher number of scales were all collected from the Lucala River, a north bank tributary of the Kwanza River. The specimens (SAIAB 84645) with the narrower caudal peduncle and fewer caudal scales were collected from the mainstem Kwanza River. There are no obvious natural barriers between the collection sites of the different morphs. However, the Cambambe Hydroelectric Power Station, a dam on the Kwanza River mainstem completed in 1963, currently separates these morphs. Intraspecific differences in caudal peduncle depth have been linked to differences in water conductivity. However, water quality variables were not recorded during the 2007 and 2008 surveys. This will need to be ascertained in future surveys.
H. angusticaudata sp. nov
4.4
https://zoobank.org/NomenclaturalActs/43fa6d0f-461b-4933-9ad5-be3a36e10b58; Figure 12.
(a) Holotype of Heteromormyrus angusticaudata sp. nov. (SAIAB 85039), 152.5 mm standard length (SL), from the Kwanza River; (b) paratype of H. angusticaudata sp. nov. (SAIAB 85039), 149.0 mm SL, from the Kwanza River, southern Africa; (c) X‐ray radiograph of holotype; (d, e) live pictures of H. angusticaudata sp. nov. Scale bar =1 cm.
Hippopotamyrus sp. ‘K1’ in Mutizwa et al. (2021).
Heteromormyrus sp. ‘K1’ in Sullivan et al. (2022).
Holotype
4.4.1
SAIAB 246302, 118.3 mm SL, northern track between Tchuimbo and Kutato, 12° 16′ 38″ S, 16° 21′ 3″ E, Angola, E. Swartz and A. Chakona, 12 August 2008; BOLD genseq‐1 COI: SAFW399‐08; GenBank genseq‐1 S7 intron: MW756324.
Paratypes
4.4.2
SAIAB 85039, 6 specimens 94.8*–*152.5 mm SL, northern track between Tchuimbo and Kutato, 12° 16′ 38″ S, 16° 21′ 3″ E, Kwanza River system, Angola, E. Swartz and A. Chakona, 2 August 2008, BOLD genseq‐2 COI: SAFW398‐08, GenBank genseq‐2 S7 intron: MW756325.
Non‐type specimens
4.4.3
SAIAB 85147, 1 specimen 114.8 mm SL, below Calema Falls, 09° 53′ 24″ S, 16° 18′ 16″ E, Angola, E. Swartz and A. Chakona, 21 August 2008; BOLD genseq‐3 COI: SAFW452‐08. SAIAB 85169, 1 specimen 29.6 mm SL, just above Calema Falls 09° 53′ 28″ S,16° 18′ 26″ E, Angola, E. Swartz and A. Chakona, 22 August 2008, BOLD genseq‐3 COI: SAFW545‐08, GenBank genseq‐3 S7 intron: MW756322, MW756323.
Diagnosis
4.4.4
H. angusticaudata sp. nov. possesses the highest number of vertebrae (46*–47), a character that differentiates this species from all its congeners in southern Africa: H. tangwenai sp. nov. (41–42), H. ndauorum sp. nov. (42), H. chilembwei sp. nov. (43), H. szaboi (40–41), H. xanekweorum sp. nov. (41–42), H. longilateralis (42–43), H. ansorgii (42–44), H. dolichorhynchus (42–44), H. pappenheimi (42–43) and H. pauciradiatus (40–42). The higher number of anal‐fin rays further distinguishes H. angusticaudata sp. nov. (28–30) from H. ndauorum sp. nov. (23–24), H. tavernei (22–23), H. tangwenai sp. nov. (20–25), H. chilembwei sp. nov. (23–25), H. szaboi (22–24), H. xanekweorum sp. nov. (22–24), H. longilateralis (22–25), H. ansorgii (24–27) and H. pauciradiatus (23–26). The number of anal‐fin rays between the leading ray and the one directly below the leading dorsal‐fin ray in H. angusticaudata sp. nov. (5–6) further distinguishes this species from H. szaboi (3–4) and H. ndauorum sp. nov. (3–4). A higher number of dorsal‐fin rays separate H. angusticaudata sp. nov. (22–24) from H. tavernei (17–20), H. longilateralis (17–21), H. szaboi (19), H. ansorgii (18–21), H. xanekweorum sp. nov. (18–20), H. ndauorum sp. nov. (18–20), H. tangwenai sp. nov. (17–20) and H. chilembwei sp. nov. (18–20). A lower number of scales around the caudal peduncle separate H. angusticaudata sp. nov. (16) from H. tavernei (20–22), H. tangwenai sp. nov. (18–20) and H. longilateralis (20). A narrow caudal peduncle distinguishes H. angusticaudata sp. nov. (4.9%–*5.6%SL) from H. pauciradiatus (7.3%–9.2%SL), H. longilateralis (7.0%–8.2%SL), H. szaboi (6.4%–10.4%SL), H. ansorgii (6.0%–7.3%SL), H. chilembwei sp. nov. (5.9%–6.2%SL), H. tangwenai sp. nov. (6.3%–7.9%SL) and H. ndauorum sp. nov. (6.7%–7.5%SL). H. angusticaudata sp. nov. is readily distinguished from its congeners except H. dolichorhynchus, H. longilateralis and H. xanekweorum sp. nov. by a series of thin curved vertical bars that are more conspicuous in the anterior portion of the flank. The combination of a dark blotch present near the flexion point of the caudal fin and a dark vertical bar in the caudal peduncle further distinguishes H. angusticaudata sp. nov. from H. dolichorhynchus, H. chilembwei sp. nov. and H. ndauorum sp. nov. that do not have any clearly visible marks on the caudal peduncle; H. longilateralis, H. szaboi, H. tangwenai sp. nov. and H. xanekweorum sp. nov. have only a dark blotch present near the flexion point of the caudal fin.
Description
4.4.5
Proportional measurements and meristics are summarised in Table 4. Holotype meristic counts are presented in parentheses.
Blunt snout, below eye level that gradually blends into convex dorsal profile of body. Small sub‐terminal mouth occurring below the level of pectoral‐fin base. Small rounded chin swelling transitions into concave ventral profile of head. Round orbit. Anterior and posterior nostrils laterally positioned, closer to snout tip than opercular opening, anterior to and arranged horizontally in line with orbit. Anterior nostril positioned slightly higher than posterior nostril. Small gill opening with soft skin cover adjacent to the pectoral‐fin base. Bicuspid teeth: 6*–7 (7) in upper jaw and 8–*12 (9) in lower jaw.
Laterally compressed body with greatest breadth occurring across gill covers. Fusiform body with greatest depth occurring between origin of dorsal and anal fins. Body tapers gently towards head, while tapering sharply from origins of both dorsal and anal fins to produce relatively thin caudal peduncle. Caudal peduncle maintains same thin depth from origin to roughly around half its length, gradually widens into two symmetrical lobes. Body covered with transparent membrane that becomes increasingly translucent to opaque towards head, dorsal and ventral surfaces in preserved specimens. Head without scales; the rest of the body is covered with small cycloid scales with reticulated striae. Lateral line originates approximately above pectoral fin and forms a straight line to the caudal peduncle. There are 78*–85 (84) scales along the lateral line, 19–*24 (22) of them along caudal peduncle and 16 around caudal peduncle. Urogenital opening situated adjacent to origin of anal fin.
Rounded pectoral fin with 10 rays, extending just beyond pelvic‐fin origin. Short pelvic fin with 6 rays. The first two dorsal‐ and anal‐fin rays are unbranched, unsegmented and small. These two unbranched and unsegmented rays are followed by a single long unbranched and segmented ray, followed by numerous branched, segmented rays. The last dorsal‐ and anal‐fin ray is usually branched all the way to its base. Anal and dorsal fins set towards the posterior of the body. Anal‐fin origin is 5–6 (5) fin rays anterior to the dorsal‐fin origin. Dorsal‐fin rays: 22*–24 (23); anal‐fin rays: 28–*30 (30). In both dorsal and anal fins, the anterior fin rays increase in length up to fourth and fifth rays, then subsequent rays get progressively shorter. Caudal fin deeply forked with rounded lobes with bases covered in scales; distance from caudal‐fin flexion point to caudal‐fin tips is roughly equal to caudal peduncle length.
See Figure 12. Live colour: body colour ranges from silver to grey. Dorsal surface usually darker than the ventral surface. A series of thin curved vertical bars more conspicuous in the anterior portion of flank. Dark vertical bar originates from origin of dorsal fin. Dark blotch presents near the flexion point of the caudal fin and a dark vertical bar just anterior to this blotch. Fins grey, a shade lighter than the body. Preserved in ethanol: body colour light brown. Dorsal surface usually darker than ventral surface. Dark vertical bar originates from origin of dorsal fin. Dark blotch presents near the flexion point of the caudal fin and a dark vertical bar just anterior to this blotch. A series of clearly visible thin curved vertical bars more conspicuous in the anterior portion of flank, lateral line similarly darkened as well. Lightly coloured fins.
Total vertebrae: 46*–47 (46), pre‐caudal vertebrae: 20–21 (20), caudal vertebrae: 26–27 (27), vertebrae at first dorsal radial: 22–24 (22), vertebrae at first anal radial: 21–*22 (21).
Determining the sex of adults can be done externally by examining their anal‐fin base. In males, the body wall is dorsally indented, giving the dorsal margin of the anal fin sigmoid curvature. In females, the body wall appears almost straight. All the specimens of H. angusticaudata examined in this study had a straight body wall at their anal‐fin base. Because no verification of the sex of the specimens was carried out (e.g., by dissection of gonads), it was not possible to confirm whether this species is sexually dimorphic. It is possible that there were no sexually mature males among the examined specimens.
Distribution
4.4.6
The species was collected in the upper and middle sections of the Kwanza River mainstem; see Figure 9.
Etymology
4.4.7
The species name means narrow‐tailed, and it is a compound word composed of the Latin words angustus, meaning narrow, and cauda, meaning tail. This is a reference to the very thin caudal peduncle of this species.
H. xanekweorum sp. nov
4.5
https://zoobank.org/NomenclaturalActs/e59930ae-52a7-4adf-9fee-6a6bd16d18dc; Figure 13.
(a) Holotype of Heteromormyrus xanekweorum sp. nov. (SAIAB 186792), male 66.7 mm standard length (SL), from the Okavango River, southern Africa; (b) paratype of H. xanekweorum sp. nov. (SAIAB 186792), female 99 mm SL, from the Okavango River, southern Africa; (c) live picture of H. xanekweorum sp. nov.; (d) X‐ray radiograph of holotype. Scale bar =1 cm.
Hippopotamyrus sp. ‘OK’ in Mutizwa et al. (2021).
Heteromormyrus sp. ‘OK’ in Sullivan et al. (2022).
Holotype
4.5.1
SAIAB 246300, 84.5 mm SL, just south of Chitembo, Chitembo River, 13° 31′ 41.1″ S, 16° 45′ 31.4″ E, Okavango River system, Angola, R. Bills, P. Skelton, F. de Almeida and M. Domingos, 16 May 2012, BOLD genseq‐1 COI: ANGFW139‐12.
Paratypes
4.5.2
SAIAB 186792, 9 specimens 37.0*–*118.1 mm SL, just south of Chitembo, Chitembo River, 13° 31′ 41.1″ S, 16° 45′ 31.4″ E, Okavango River system, Angola, R. Bills, P. Skelton, F. de Almeida and M. Domingos, 16 May 2012, BOLD and GenBank genseq‐2 COI: ANGFW138‐12, MW600872, MW600873, GenBank genseq‐2 cyt b: MW600903, S7 intron: MW756320, MW756321.
Non‐type specimens
4.5.3
SAIAB 186693, 4 specimens 32.0–113.0 mm SL, bridge of 1000 Mines north of Mambue, ‘Bie’, Cacuchi River, 13° 35′ 39.6″ S, 16° 52′ 49.8″ E, Okavango River, Angola, R. Bills, P. Skelton, F. de Almeida and M. Domingos, 11 May 2012, BOLD genseq‐3 COI: ANGFW053‐12; SAIAB 186894, 1 specimen 38.0 mm SL, Cuanavale River just above confluence, 15° 08′ 08.5″ S, 19° 11′ 44.6″ E, Okavango River, Angola, R. Bills, P. Skelton, F. de Almeida and M. Domingos, 19 May 2012, GenBank genseq‐3 COI: MW600877, cyt b: MW600898; SAIAB 186651, 3 specimens 39–41 mm SL, Square Ponglesia‐Cubango, Quebe River, 14° 56′ 22.9″ S, 17° 43′ 07.7″ E, Okavango River system, Angola, R. Bills, P. Skelton, F. de Almeida, 9 May 2012, BOLD genseq‐3 COI: ANGFW019‐12, GenBank genseq‐3 cyt b: MW600899; SAIAB 202504, 1 specimen 84.0 mm SL, fish eagle camp, Cuito River, 13° 46′ 17.00″ S, 18° 34′ 48.00″ E, Okavango River system, Angola, A. Costa, 20 June 2015, GenBank genseq‐3 COI: MW600876; SAIAB 204769, 1 specimens 72.0 mm SL, Rapid 1, Cubango River, 13° 02′ 41.12″ S, 16° 22′ 29.93″ E, Okavango River system, Angola, P. Skelton and B. van der Waal, 8 May 2017, GenBank genseq‐3 COI: MW600870, cyt b: MW600893; SAIAB 203980, 1 specimen 51.0 mm SL, below bridge on Cuchi River, 14° 42′ 09.8″ S, 17° 22′ 43.1″ E, Okavango River system, Angola, B. van de Waal and N. Mazungula, 15 October 2016, GenBank genseq‐3 COI: MW600869; SAIAB 187022, 1 specimen 39.0 mm SL, Popa, Okavango River, 18° 07′ 18.2″ S, 21° 35′ 00.2″ E, Okavango River system, Namibia, R. Bills; P. Skelton, 24 May 2012, BOLD genseq‐3 COI: ANGFW227‐12, GenBank genseq‐3 cyt b: MW600900; SAIAB 204843, 3 specimens 48.0–61.0 mm SL, Kavango mission rapids camp, Cubango River, 13° 19′ 40.14″ S, 16° 24′ 39.82″ E, Okavango River system, Angola, P Skelton and B van der Waal on 11 May 2017, GenBank genseq‐3 COI: MW600871, cyt b: MW600897; SAIAB 186726, 3 specimens 28.0–92.0 mm SL, Cacuchi tributary (culvet), Mumbue, ‘Bie’, Watana River, 13° 16′ 46.7″ S, 16° 44′ 46.1″ E, Okavango River system, Angola, R. Bills; P. H. Skelton; M. Domingos; F. de Almeida on 13 May 2012, BOLD genseq‐3 COI: ANGFW090‐12, GenBank genseq‐3 cyt b: MW600894; SAIAB 202483, 1 specimen 66.0 mm SL, Hippo camp, 13° 08′ 29.00″ S, 18° 29′ 08.00″ E, Okavango River system, Angola, A. Costa on 11 June 2015, GenBank genseq‐3 COI: MW600874, cyt b: MW600895, S7 intron: MW756362, MW756363; SAIAB 203161, 1 specimen 57.0 mm SL, Kalva and Cuito confluence, Cuito River, 12.74878° S, 18.354334° E, Okavango River system, Angola, A. Costa on 25 Feb 2016, GenBank genseq‐3 COI: MW600875, cyt b: MW600896.
Diagnosis
4.5.4
The number of anal‐fin rays between the leading ray and the one directly below the leading dorsal‐fin ray distinguishes H. xanekweorum sp. nov. (4–5) from H. pappenheimi (6–9), H. pauciradiatus (6–9), H. ansorgii (6–9), H. tangwenai sp. nov. (6–7) and H. chilembwei sp. nov. (6–7). A lower number of anal‐fin rays further separate H. xanekweorum sp. nov. (22*–24) from H. angusticaudata (28–30), H. dolichorhynchus (26–29) and H. pappenheimi (26–28). A higher number of scales around caudal peduncle separate H. xanekweorum sp. nov. (16–20) from H. pappenheimi (12–14). A lower number of caudal vertebrae differentiate H. xanekweorum sp. nov. (22–24) from H. angusticaudata (26–27). The number of vertebrae before first dorsal radial further distinguishes H. xanekweorum sp. nov. (20–21) from H. angusticaudata (22–24). The vertebrae before first anal radial further separate H. xanekweorum sp. nov. (19–20) from H. pappenheimi (18). H. xanekweorum sp. nov. possesses a narrower caudal peduncle depth (5.0%–6.8%SL) that distinguishes it from H. longilateralis (7.0%–8.2%SL), H. tavernei (6.9%–8.6%SL) and H. pauciradiatus (7.3%–9.2%SL). H. xanekweorum sp. nov. (19.8%–*22.8%SL) has a narrower body depth compared to H. pauciradiatus (23.1%–27.9%SL) and H. szaboi (22.4%–26.3%SL). A shallower caudal peduncle depth further separates H. xanekweorum sp. nov. (5.0%–6.8%SL) from H. szaboi (6.4%–10.4%SL). A dark blotch near flexion point of the caudal fin distinguishes H. xanekweorum sp. nov. from H. dolichorhynchus, H. chilembwei sp. nov. and H. ndauorum sp. nov. that do not have any clearly visible marks on caudal peduncle; H. ansorgii, H. pauciradiatus and H. angusticaudata have only a clearly visible dark blotch present near flexion point of the caudal fin and a dark vertical bar on caudal peduncle.
Description
4.5.5
Proportional measurements and meristics are summarised in Table 5. Holotype meristic counts are presented in parentheses.
Rounded blunt snout, below eye level. Straight dorsal head profile forms obtuse angle with the gently inclined dorsal body profile. Small sub‐terminal mouth occurring below the level of pectoral‐fin base. Small rounded chin swelling transitions into deep concave ventral profile of head. Round orbit. Anterior and posterior nostrils laterally positioned, closer to snout tip than opercular opening, anterior to and arranged horizontally in line with orbit. Anterior nostril positioned slightly higher than posterior nostril. Small gill opening with soft skin cover adjacent to pectoral‐fin base. Bicuspid teeth: 6*–7 (7) in upper jaw and 6–*10 (8) in lower jaw.
Laterally compressed body with greatest breadth occurring across gill covers. Fusiform body with greatest depth occurring between origin of dorsal and anal fins. Body tapers gently towards head, while tapering sharply from origins of both dorsal and anal fins to produce a relatively thin caudal peduncle. Caudal peduncle maintains same thin depth from its origin to roughly around half its length, then it gradually widens into two symmetrical lobes. Body covered with transparent membrane that becomes increasingly translucent to opaque towards head, dorsal and ventral surfaces in preserved specimens. Head without scales, the rest of the body is covered with small cycloid scales with reticulated striae. Lateral line originates approximately above pectoral fin and forms a straight line to caudal peduncle. There are 64*–86 (76) scales along lateral line, 16–*23 (20) along caudal peduncle and 16–20 (20) around caudal peduncle. Urogenital opening situated adjacent to origin of anal fin.
Rounded pectoral fin with 10 rays, extends just beyond origin of pelvic fin. Short pelvic fin with 6 rays. The first two dorsal and anal‐fin rays are unbranched, unsegmented and small. These two unbranched and unsegmented rays are followed by a single long unbranched and segmented ray, followed by numerous branched, segmented rays. The last dorsal‐ and anal‐fin ray is usually branched all the way to its base. Anal and dorsal fins set towards the posterior of body. Anal‐fin origin is 4–5 (5) fin rays anterior to the dorsal‐fin origin. Dorsal‐fin rays: 18*–20 (18); anal‐fin rays: 22–*24 (22). In both dorsal and anal fins, the anterior fin rays increase in length up to the fourth and fifth rays, then subsequent rays get progressively shorter. Caudal fin deeply forked with rounded lobes with bases covered in scales; distance from caudal‐fin flexion point to caudal‐fin tips is roughly equal to caudal peduncle length.
Live colour: body colour ranges from light to dark brown. Dorsal surface usually darker than the ventral surface. Thin curved vertical bars more conspicuous in the anterior portion of flank. Dark vertical bar originates from origin of dorsal fin, barely visible in darker specimens. A barely visible dark blotch is present near flexion point of the caudal fin. Fins brown. Preserved in ethanol: body colour ranges from light to dark brown. Dorsal surface usually darker than ventral surface. Dark vertical bar originates from origin of dorsal fin, barely visible in darker specimens. A barely visible dark blotch is present near flexion point of the caudal fin. Thin curved vertical bars conspicuous in the anterior portion of flank. Fins brown.
Total vertebrae: 41*–42 (41), pre‐caudal vertebrae: 18–19 (19), caudal vertebrae: 22–24 (22), vertebrae at first dorsal radial: 20–21 (20), vertebrae at first anal radial: 19–*20 (19).
Determining the sex of adults can be done externally by examining their anal‐fin base. In males, the body wall is dorsally indented, giving the dorsal margin of the anal fin a sigmoid curvature. In females, the body wall appears almost straight.
Distribution
4.5.6
This species is found throughout the Okavango River, Angola and Namibia; see Figure 9.
Etymology
4.5.7
The specific epithet is a noun in the genitive case of the Xanekwe people of Okavango River. The Xanekwe are one of the groups of people that have historically lived along the Okavango River and its delta. They traditionally were hunter gatherers and fishermen with close ties to the riverine environment.
Remarks
4.5.8
H. xanekweorum and H. tangwenai have very similar morphology that makes these species difficult to distinguish.
H. chilembwei sp. nov
4.6
https://zoobank.org/NomenclaturalActs/2ba0ea02-8cdb-430b-99cb-0f2dac1a5cca; Figure 14.
(a) Holotype of Heteromormyrus chilembwei sp. nov. (SAIAB 119004), 65.0 mm standard length (SL), from the Ruo River, lower Zambezi River system, Malawi, southern Africa; (b) paratype of H. chilembwei sp. nov. (SAIAB 119004), 69.0 mm SL, from the Ruo River, lower Zambezi River system, Malawi, southern Africa; (c) X‐ray radiograph of holotype. Scale bar =1 cm.
Hippopotamyrus sp. ‘Ruo’ in Chakona et al. (2018).
Hippopotamyrus sp. ‘Ruo’ in Mutizwa et al. (2021).
Heteromormyrus sp. ‘Ruo’ in Sullivan et al. (2022).
Holotype
4.6.1
SAIAB 246299, 65.1 mm SL, Lujeri tea estate, 16° 02.19′ S, 35° 39.80′ E, Ruo River, lower Zambezi River system, Malawi, D. Tweddle and T. Mäkinen, 21 May 2011, BOLD genseq‐1 COI: MAFW025‐11, GenBank genseq‐1 cyt b: MW600888, S7 intron: MW756346.
Paratypes
4.6.2
SAIAB 119004, 3 specimens 48.0*–69.0 mm SL, Lujeri tea estate, 16° 02.19′ S, 35° 39.80′ E, Ruo River, lower Zambezi River system, Malawi, D. Tweddle and T. Mäkinen, 21 May 2011, GenBank genseq‐2 S7 intron: MW756347; SAIAB 119013, 3 specimens 33.9–*47.9 mm SL, Lujeri tea estate, Muluzi stream, lower Zambezi River system, Malawi, 16° 03.26′ S, 35° 40.27′ E, D. Tweddle and T. Mäkinen, 21 May 2011, BOLD genseq‐2 COI: MAFW039‐11, GenBank genseq‐2 cyt b: MW600889, S7 intron: MW756332, MW756333.
Diagnosis
4.6.3
H. chilembwei sp. nov. possesses 16 scales around its caudal peduncle that further distinguish it from H. tangwenai sp. nov. (18*–20), H. tavernei (20–22) and H. longilateralis (20). A high number of scales along the length of the caudal peduncle differentiate H. chilembwei sp. nov. (21–23) from H. ansorgii (16–19), H. pauciradiatus (14–18), H. dolichorhynchus (17–20) and H. ndauorum sp. nov. (16–20). A low number of anal‐fin rays separate H. chilembwei sp. nov. (23–25) from H. angusticaudata (28–30), H. dolichorhynchus (26–29) and H. pappenheimi (26–28). A low number of caudal vertebrae differentiated H. chilembwei sp. nov. (24–25) from H. angusticaudata (26–27). The high number of vertebrae before the first dorsal‐fin radial further distinguishes H. chilembwei sp. nov. (21–22) from H. pappenheimi (19). A higher number of vertebrae before the first anal‐fin radial separate H. chilembwei sp. nov. (20) from H. pauciradiatus (18–19) and H. pappenheimi (18). The number of anal‐fin rays between the leading ray and the one directly below the leading dorsal‐fin ray distinguishes H. chilembwei sp. nov. (6–7) from H. xanekweorum (4–5), H. longilateralis (3–5), H. szaboi (3–4) and H. ndauorum sp. nov. (3–4) with more anteriorly set dorsal fins. A narrow caudal peduncle distinguishes H. chilembwei sp. nov. (5.9%–*6.2%SL) from H. pauciradiatus (7.3%–9.2%SL), H. tavernei (6.9%–8.6%SL), H. longilateralis (7.0%–8.2%SL), H. szaboi (6.4%–10.4%SL), H. tangwenai sp. nov. (6.3%–7.9%SL) and H. ndauorum sp. nov. (6.7%–7.5%SL). The snout length distinguishes H. chilembwei sp. nov. (42.6%–50.2%HL) from H. ansorgii (34.6%–41.4%HL), H. pauciradiatus (29.7%–41.0%HL), H. tavernei (24.7%–30.0%HL), H. longilateralis (38.3%–42.5%HL) and H. angusticaudata (39.5%–42.3%HL). The interorbital width distinguishes H. chilembwei sp. nov. (46.4%–51.6%HL) from H. ansorgii (28.6%–45.4%HL), H. pauciradiatus (29.7%–41.0%HL), H. tavernei (24.7%–30.0%HL), H. longilateralis (35.5%–43.7%HL), H. dolichorhynchus (34.8%–43.4%HL) and H. angusticaudata (38.1%–43.8%HL). H. chilembwei sp. nov. is readily distinguished from its congeners by its deep, rounded dorsal head profile and dark pectoral fins. The lack of clearly visible marks in the caudal peduncle further distinguishes H. chilembwei from H. ansorgii, H. pauciradiatus and H. angusticaudata that have a clearly visible dark blotch present near the flexion point of the caudal fin and a dark vertical bar in the caudal peduncle; H. longilateralis, H. szaboi, H. tangwenai and H. xanekweorum have only a dark blotch present near the flexion point of the caudal fin.
Description
4.6.4
Proportional measurements and meristics are summarised in Table 5. Holotype meristic counts are presented in parentheses.
Rounded blunt snout, below eye level. Deep, rounded dorsal head profile that gradually becomes gently inclined like the rest of dorsal profile of the body. Small sub‐terminal mouth occurring below the level of pectoral‐fin base. Small rounded chin swelling transitions into concave ventral profile of the head. Round orbit. Anterior and posterior nostrils laterally positioned, closer to snout tip than opercular opening, anterior to and arranged horizontally in line with orbit. Anterior nostril positioned slightly higher than posterior nostril. Small gill opening with soft skin cover adjacent to pectoral‐fin base. Bicuspid teeth: 7*–8 (7) in upper jaw and 8–*9 (9) in lower jaw.
Laterally compressed body with greatest breadth occurring across gill covers. Fusiform body with greatest depth occurring roughly midway between the pectoral‐fin and pelvic‐fin origins. Body tapers gently towards head, while tapering sharply from origins of both dorsal and anal fins to produce a relatively thin caudal peduncle. Caudal peduncle maintains the same depth from its origin to roughly around half its length, then it gradually widens into two symmetrical lobes. Body covered with transparent membrane that becomes increasingly translucent to opaque towards head, dorsal and ventral surfaces in preserved specimens. Head without scales; rest of the body is covered in small cycloid scales with reticulated striae. Lateral line originates approximately above pectoral fin and forms a straight line to caudal peduncle. There are 82*–84 (82) scales along the lateral line, 21–*23 (22) along caudal peduncle and 16 around caudal peduncle. Urogenital opening situated adjacent to the origin of the anal fin.
Rounded pectoral fin with 10 rays, extends just beyond the origin of the pelvic fin. Short pelvic fin with 6 rays. The first two dorsal and anal‐fin rays are unbranched, unsegmented and small. These two unbranched and unsegmented rays are followed by a single long unbranched and segmented ray, followed by numerous branched, segmented rays. The last dorsal‐ and anal‐fin ray is usually branched all the way to its base. Anal and dorsal fins set towards the posterior of the body. Anal‐fin origin is 6–7 (7) fin rays anterior to the dorsal‐fin origin. Dorsal‐fin rays: 18*–20 (20); anal‐fin rays: 23–*25 (25). In both dorsal and anal fins, anterior fin rays increase in length up to the 4th and 5th rays then subsequent rays get progressively shorter. Caudal fin deeply forked with rounded lobes that have scaled bases; distance from caudal‐fin flexion point to caudal‐fin tips roughly equal to caudal peduncle length.
Body colour ranges from light to dark brown in specimens preserved in ethanol. Dorsal surface usually darker than ventral surface. Dark vertical bar originates from origin of dorsal fin, and it is barely visible in darker specimens. A barely visible dark blotch is present near flexion point of the caudal fin. Dark brown pectoral and pelvic fins. Dark caudal peduncle without any additional markings.
Total vertebrae: 43 (43), pre‐caudal vertebrae: 18–19 (18), caudal vertebrae: 24–25 (25), vertebrae at first dorsal radial: 21*–*22 (22), vertebrae at first anal radial: 20.
All H. chilembwei specimens examined in this study had a straight body wall at their anal‐fin base. Because no verification of the specimen's sex was carried out (e.g., by dissection of gonads), it is not possible to confirm whether these species were not sexually dimorphic. It is also possible that there were no sexually mature males among the examined specimens.
Distribution
4.6.5
This species was collected from the Ruo River, a tributary of the Shire River, which flows into the lower Zambezi River system. The Ruo is a small montane river that flows off the Mulanje Massif, which is situated in the southeast corner of Malawi close to the Mozambique border (see Figure 9).
Etymology
4.6.6
The specific epithet is a noun in the genitive case that honours the Reverend John Chilembwe, who led the first nationalist uprising in Malawi from 1914 to 1915 protesting against forced labour and African involvement in World War I.
H. tangwenai sp. nov
4.7
https://zoobank.org/NomenclaturalActs/21a96ccb-2ff7-476b-996b-d9d6859819b4; Figure 15.
(a) Holotype of Heteromormyrus tangwenai sp. nov. male (SAIAB 200926), 102.2 mm standard length (SL), from the Nyamukombe River, Pungwe River system, Zimbabwe; (b) paratype of H. tangwenai sp. nov. female (SAIAB 200926), 69.0 mm SL, from the Nyamukombe River, Pungwe River system, Zimbabwe; (c) X‐ray radiograph of holotype; (d, e) live pictures of H. tangwenai sp. nov. Scale bar =1 cm.
Hippopotamyrus sp. ‘Pungwe’ in Chakona et al. (2018).
Hippopotamyrus sp. ‘Pungwe’ in Mutizwa et al. (2021).
Heteromormyrus sp. ‘Pungwe’ in Sullivan et al. (2022).
Holotype
4.7.1
SAIAB 246298, 102.2 mm SL, Nyamukombe River bridge in Sagambe, 18° 23′ 45.0″ S, 32° 58′ 14.6″ E, Pungwe River system, Zimbabwe, A. Chakona, W. Kadye and T. Bere, 11 December 2014, GenBank genseq‐1 COI: MH432084, cyt b: MW600887, S7 intron: MW756348.
Paratypes
4.7.2
SAIAB 200926, 3 specimens 60.7–80.1 mm SL, Nyamukombe River bridge in Sagambe,18° 23′ 45.0″ S, 32° 58′ 14.6″ E, Pungwe River system, Zimbabwe, A. Chakona, W. Kadye and T. Bere, 11 December 2014, GenBank genseq‐2 COI: MH432085, S7 intron: MW756349.
Non‐type specimens
4.7.3
SAIAB 201082, 2 specimens 26.2–69.0 mm SL, Nyamukwarara River next to the Nyamukwarara Clinic 18° 41′ 30.5″ S, 32° 55′ 25.0″ E, Pungwe River system, Zimbabwe, A. Chakona, W. Kadye and T. Bere, 17 December 2014, GenBank genseq‐3 COI: MH432078, MH432081; SAIAB 201091, 6 specimens 49.2–88.3 mm SL, Chiyengwa River just before confluence with Nyamukwarara River, 18° 41′ 16.0″ S, 32° 55′ 19.2″ E, Pungwe River system, Zimbabwe, A. Chakona, W. Kadye and T. Bere, 17 December 2014, GenBank genseq‐3 COI: MH432086, MH432083; SAIAB 201071, 5 specimens 22.8*–*66.5 mm SL, Makanga bridge on the road to Hauna, 18° 32′ 37.8″ S, 32° 48′ 04.6″ E, Pungwe River system, Zimbabwe, A. Chakona, W. Kadye and T. Bere, 16 December 2014, GenBank genseq‐3 COI: MH432079; SAIAB 201030, 5 specimens, 24.8–83.9 mm SL, Rwera River through Marupeti shops at Zimbabwe and Mozambique border, 18° 32′ 36.4″ S, 32° 48′ 5.8″ E, Pungwe River system, Zimbabwe, A. Chakona, W. Kadye and T. Bere, 14 December 2014, GenBank genseq‐3 COI: MH432077, cyt b: MW600885, S7 intron: MW756328, MW756329; SAIAB 201074, 1 specimen 78.1 mm SL, Pungwe River mainstem by the abandoned church, 18° 26′ 29.1″ S, 32° 53′ 14.9″ E, Pungwe River system, Zimbabwe, A. Chakona, W. Kadye and T. Bere, 16 December 2014, GenBank genseq‐3 COI: MH432080; SAIAB 201044, 1 specimen 86.9 mm SL, Nyamukombe River bridge between Chisuko School and Katiyo, 18° 22′ 55.7″ S, 33° 01′ 57.8″ E, Pungwe River system, Zimbabwe, A. Chakona, W. Kadye and T. Bere, 14 December 2014, GenBank genseq‐3 COI: MH432076, cyt b: MW600884, S7 intron: MW756350, MW756351; SAIAB 205084, 1 specimen 62.0 mm SL, from bridge on road to Gatsi, below falls, 18° 31.945′ S, 032° 48.452′ E, Pungwe River system, Zimbabwe, A. Chakona, T. Bere and M. Machingura, 13 December 2013, GenBank genseq‐3 COI: MH432071; SAIAB 201021, 4 specimens 55.9–104.9 mm SL, bridge on road to Honde mission, 18° 32′ 37.5″ S, 32° 48′ 15.8″ E, Pungwe River system, Zimbabwe, A. Chakona, W. Kadye, T. Bere, M. Machingura, 13 December 2013, GenBank genseq‐3 COI: MH432082.
Diagnosis
4.7.4
H. tangwenai sp. nov. possesses a higher number of scales around its caudal peduncle (18*–20), differentiating it from H. ndauorum sp. nov. (16), H. chilembwei (16), H. ansorgii (14–16), H. dolichorhynchus (12–16) and H. pappenheimi (12–14). A lower number of anal‐fin rays further separate H. tangwenai sp. nov. (20–25) from H. angusticaudata (28–30) and H. dolichorhynchus (26–29). The number of anal‐fin rays between the leading ray and the one directly below the leading dorsal‐fin ray in H. tangwenai sp. nov. (6–7) distinguishes it from H. xanekweorum (4–5), H. longilateralis (3–5), H. szaboi (3–4) and H. ndauorum sp. nov. (3–4). H. tangwenai sp. nov. (21.0%–23.9%SL) has a narrower body depth compared to H. pauciradiatus (23.9%–27.9%SL). H. tangwenai sp. nov. is distinguished from H. tavernei by interorbital width (41.7%–54.4% vs. 24.7%–30.0%HL), snout length (41.1%–53.5% vs. 24.7%–30.0%HL) and head length (21.6%–24.9% vs. 25.8%–29.2%SL). H. tangwenai sp. nov. has fewer vertebrae (41–42) compared to H. chilembwei (43) and H. angusticaudata (46–47). H. tangwenai sp. nov. is further differentiated from H. angusticaudata by a low number of caudal vertebrae (22–25 vs. 26–27), fewer vertebrae before the first dorsal radial (19–21 vs. 22–24) and fewer vertebrae before the first anal‐fin radial (18–20 vs. 21–*22). H. tangwenai sp. nov. has a rounded head profile that distinguishes it from H. xanekweorum, H. szaboi and H. longilateralis that have a flat head profile that forms an obtuse angle with the rest of the body profile. H. tangwenai sp. nov. has a dark blotch near flexion point of the caudal fin which distinguishes it from H. dolichorhynchus, H. chilembwei sp. nov. and H. ndauorum sp. nov. that do not have any clearly visible marks in the caudal peduncle; H. ansorgii, H. pauciradiatus and H. angusticaudata have a clearly visible dark blotch present near the flexion point of the caudal fin and a dark vertical bar in the caudal peduncle.
Description
4.7.5
Proportional measurements and meristics are summarised in Table 5. Holotype meristic counts are presented in parentheses.
Rounded blunt snout, below eye level. Deep, rounded dorsal head profile that gradually becomes gently inclined like the rest of the dorsal profile of the body. Small sub‐terminal mouth occurring below the level of pectoral‐fin base. Small rounded chin swelling transitions into concave ventral profile of head. Round orbit. Anterior and posterior nostrils laterally positioned, closer to snout tip than opercular opening, anterior to and arranged horizontally in line with orbit. Anterior nostril positioned slightly higher than posterior nostril. Small gill opening with soft skin cover adjacent to pectoral fin‐base. Bicuspid teeth: 7*–8 (7) in upper jaw and 8–*9 (8) in lower jaw.
Laterally compressed body with greatest breadth occurring across gill covers. Fusiform body with greatest depth occurring between the origin of dorsal and anal fins. Body tapers gently towards head, while tapering sharply from origins of both dorsal and anal fins to produce a relatively thin caudal peduncle. Caudal peduncle maintains the same thin depth from its origin to roughly around half its length, then it gradually widens into two symmetrical lobes. Body covered with transparent membrane that becomes increasingly translucent to opaque towards head, dorsal and ventral surfaces in preserved specimens. Head without scales; rest of the body is covered with small cycloid scales with reticulated striae. Lateral line originates approximately above pectoral fin and forms a straight line to the caudal peduncle. There are 71*–*83 (83) scales along the lateral line, 18–22 (21) along caudal peduncle and 18–20 (20) around caudal peduncle. Urogenital opening situated adjacent to origin of anal fin.
Rounded pectoral fin with 10 rays, extends just beyond the origin of the pelvic fin. Short pelvic fin with 6 rays. The first two dorsal‐ and anal‐fin rays are unbranched, unsegmented and small. These two unbranched and unsegmented rays are followed by a single long unbranched and segmented ray, followed by numerous branched, segmented rays. The last dorsal‐ and anal‐fin ray is usually branched all the way to its base. Anal and dorsal fins set towards the posterior of the body. Anal‐fin origin is 6–7 (6) fin rays anterior to the dorsal‐fin origin. Dorsal‐fin rays: 17*–20 (18); anal‐fin rays: 20–*25 (22). In both dorsal and anal fins, the anterior fin rays increase in length up to the fourth and fifth rays, then subsequent rays get progressively shorter. Caudal fin deeply forked with rounded lobes with bases covered in scales; distance from caudal‐fin flexion point to caudal‐fin tips is roughly equal to caudal peduncle length.
Live colour: body colour ranges from light to dark brown. Body surface with irregular patches of light and dark scales giving a mottled pattern in some specimens, whereas others lack this pattern. Dorsal surface usually darker than the ventral surface. Dark vertical bar originates from origin of dorsal fin, barely visible in darker specimens. Dark vertical bar originates from origin of dorsal fin, barely visible in darker specimens. Barely visible dark blotch is present near flexion point of the caudal fin. Fins brown usually a lighter shade than the body. Preserved in ethanol: body colour ranges from light to dark brown. Body surface with irregular patches of light and dark scales giving a mottled pattern. Dorsal surface usually darker than ventral surface. Dark vertical bar originates from origin of dorsal fin, barely visible in darker specimens. Barely visible dark blotch is present near flexion point of the caudal fin. Fins brown, usually a lighter shade than the body.
Total vertebrae: 41*–42 (41), pre‐caudal vertebrae: 17–19 (19), caudal vertebrae: 22–25 (24), vertebrae at first dorsal radial: 19–21 (20), vertebrae at first anal radial: 18–*20 (18).
Determining the sex of adults can be done externally by examining their anal‐fin base. In males, the body wall is dorsally indented, giving the dorsal margin of the anal fin a sigmoid curvature. In females, the body wall appears almost straight.
Distribution
4.7.6
This species was collected in the headwater streams of the Pungwe River, Zimbabwe; see Figure 9.
Etymology
4.7.7
The specific epithet is a noun in the genitive case honouring Chief Rekayi Tangwena (1910–June 1984), a traditional chief of the Tangwena people of Nyanga in the Manicaland Province of Zimbabwe. Chief Tangwena fiercely resisted the unlawful eviction of his people from their ancestral lands by the white minority settler government. He remains a celebrated national hero for his efforts in the liberation of Zimbabwe.
H. ndauorum sp. nov
4.8
https://zoobank.org/NomenclaturalActs/af945b25-a685-4089-b168-711b77e7c7c4; Figure 16.
(a) Holotype of Heteromormyrus ndauorum sp. nov. (SAIAB 67420), 149.0 mm standard length (SL), from the Mudzira River, Buzi River system, Zimbabwe and Mozambique; (b) paratype of H. ndauorum sp. nov. (SAIAB 67420), 135.0 mm SL, from the Mudzira River, Buzi River system, Zimbabwe and Mozambique; (c) X‐ray radiograph of holotype; (d, e) live pictures of H. ndauorum sp. nov. Scale bar =1 cm.
Hippopotamyrus sp. ‘Buzi’ in Chakona et al. (2018).
Hippopotamyrus sp. ‘Buzi’ in Mutizwa et al. (2021).
Heteromormyrus sp. ‘Buzi’ in Sullivan et al. (2022).
Holotype
4.8.1
SAIAB 246297, 149.0 mm SL, Mudzira River below Muhate, 19.6750° S, 33.1910° E, Buzi River system, Mozambique, R. Bills, S. Chimela, A. Chivindzi, 20 September 2002, BOLD genseq‐1 COI: SAFW886‐14.
Paratypes
4.8.2
SAIAB 67420, 2 specimens 135.0–149.0 mm SL, Mudzira River below Muhate, 19.6750° S, 33.1910° E, Buzi River system, Mozambique, R. Bills, S. Chimela and A. Chivindzi on 20 September 2002; SAIAB 205113, 4 specimens 30.0–62.0 mm SL, Rusitu River bridge, 20° 03.519′ S, 32° 51.315′ E, Buzi River system, Zimbabwe, A. Chakona, W. Kadye, T. Bere, M. Machingura,17 December 2013, GenBank genseq‐2 COI: MH432072, MH432074; cyt b: MW600901, MW600902, S7 intron: MW756340, MW756341, MW756338, MW756339.
Diagnosis
4.8.3
The number of anal‐fin rays between the leading ray and the one directly below the leading dorsal‐fin ray in H. ndauorum sp. nov. (3–4) distinguishes it from H. pappenheimi (6–9), H. pauciradiatus (6–9), H. ansorgii (6–9), H. dolichorhynchus (5–7), H. angusticaudata (5–6), H. tangwenai sp. nov. (6–7) and H. chilembwei sp. nov. (6–7). H. ndauorum sp. nov. possesses 16 scales around its caudal peduncle that distinguish it from H. tangwenai (18*–20), H. tavernei (20–22) and H. longilateralis (20). A lower number of anal‐fin rays separate H. ndauorum sp. nov. (23–24) from H. angusticaudata (28–30), H. dolichorhynchus (26–29) and H. pappenheimi (26–28). A higher number of vertebrae differentiate H. ndauorum sp. nov. (42) from H. szaboi (40–41), H. chilembwei (43) and H. angusticaudata (46–47). H. ndauorum sp. nov. is further separated from H. chilembwei by having fewer scales along its lateral line (72–78 vs. 82–84), as well as fewer scales along the length of the caudal peduncle (16–20 vs. 21–*23). Lack of clearly visible marks on caudal peduncle distinguishes H. ndauorum sp. nov. from H. ansorgii, H. pauciradiatus and H. angusticaudata that have a clearly visible dark blotch near the flexion point of the caudal fin and a dark vertical bar in the caudal peduncle; H. longilateralis, H. szaboi, H. tangwenai and H. xanekweorum have a dark blotch present near the flexion point of the caudal fin.
Description
4.8.4
Proportional measurements and meristics are summarised in Table 5. Holotype meristic counts are presented in parentheses.
Rounded blunt snout, below eye level. Straight dorsal head profile forms obtuse angle with the gently inclined dorsal body profile. Small sub‐terminal mouth occurring below the level of the pectoral‐fin base. Small rounded chin swelling transitions into concave ventral profile of head. Round orbit. Anterior and posterior nostrils laterally positioned, closer to snout tip than opercular opening, anterior to and arranged horizontally in line with orbit. Anterior nostril positioned slightly higher than posterior nostril. Small gill opening with soft skin cover adjacent to pectoral‐fin base. Bicuspid teeth: 7*–9 (7) in upper jaw and 8–*9 (8) in lower jaw.
Laterally compressed body with greatest breadth occurring across gill covers. Fusiform body with greatest depth occurring between origin of dorsal and anal fins. Body tapers gently towards head while tapering sharply from origins of both dorsal and anal fins to produce a relatively thin caudal peduncle. Caudal peduncle maintains the same thin depth from its origin to roughly around half its length, then it gradually widens into two symmetrical lobes. Body covered with transparent membrane that becomes increasingly translucent to opaque towards head, dorsal and ventral surfaces in preserved specimens. Head without scales; rest of the body is covered with small cycloid scales with reticulated striae. Lateral line originates approximately above pectoral fin and forms a straight line to the caudal peduncle. There are 72*–78 (78) scales along the lateral line, 16–*20 (16) along caudal peduncle length and 16 around caudal peduncle. Urogenital opening is situated adjacent to the origin of anal fin.
Rounded pectoral fin with 10 rays, extends just beyond the origin of the pelvic fin. Short pelvic fin with 6 rays. The first two dorsal‐ and anal‐fin rays are unbranched, unsegmented and small. These two unbranched and unsegmented rays are followed by a single long unbranched and segmented ray, followed by numerous branched, segmented rays. The last dorsal‐ and anal‐fin ray is usually branched all the way to its base. Anal and dorsal fins set towards the posterior of the body. Anal‐fin origin is 3–4 (4) fin rays anterior to the dorsal‐fin origin. Dorsal‐fin rays: 19*–20 (20); anal‐fin rays: 23–*24 (23). In both dorsal and anal fins, the anterior fin rays increase in length up to fourth and fifth rays, then subsequent rays get progressively shorter. Caudal fin deeply forked with rounded lobes that have scaled bases; distance from caudal‐fin flexion point to caudal‐fin tips is roughly equal to caudal peduncle length.
Live colour: body colour ranges from light to dark brown. Body surface with irregular patches of light and dark scales giving a mottled pattern in some specimens, whereas others lack this pattern. Dorsal surface usually darker than the ventral surface. Dark vertical bar originates from the origin of dorsal fin, barely visible in darker specimens. Dark vertical bar originates from the origin of dorsal fin, barely visible in darker specimens. Barely visible dark blotch is present near flexion point of the caudal fin. Fins brown, usually a lighter shade than the body. Preserved in ethanol: body colour dark brown. Dorsal surface usually darker than ventral surface. Dark vertical bar originates from the origin of dorsal fin, and it is barely visible in darker specimens. Barely any visible markings in the caudal peduncle.
Total vertebrae: 41–43 (42), pre‐caudal vertebrae: 17–18 (18), caudal vertebrae: 24–25 (24), vertebrae at first dorsal radial: 19–20 (20), vertebrae at first anal radial: 18–19 (18).
All the specimens of H. ndauorum examined in this study had a straight body wall at their anal‐fin base. Because no verification of the sex of the specimens was carried out (e.g., by dissection of gonads), it is not possible to confirm whether this species is not sexually dimorphic. It is also possible that there was no sexually mature males among the examined specimens.
Distribution
4.8.5
This species was collected in the head water streams of the Buzi River, Mozambique and Zimbabwe; see Figure 9.
Etymology
4.8.6
The specific epithet is a noun in the genitive case that honours the Ndau people, who have historically lived in the Chimanimani district of Zimbabwe, where the species was collected.
Identification key for southern African Heteromormyrus species based on external characters
4.9
1a. Nostrils aligned horizontally below the level of the orbit H. dolichorhynchus sp. nov. (Figure 6a).
1b. Anterior nostril was always positioned higher than the posterior nostril and both nostrils horizontally in line with the orbit (Figure 6b,c) 2
2a. Clearly visible dark blotch is present near the flexion point of the caudal fin (Figure 7b,c) 3
2b. Barely visible blotch in the caudal peduncle (Figure 7a) 9
3a. Presence of dark vertical bar just anterior to the blotch near the flexion point of the caudal fin (Figure 7c) 4
3b. Absence of a dark vertical bar just anterior to the blotch near the flexion point of the caudal fin 6
4a. Presence of a series of thin slightly curved vertical bars more conspicuous in the anterior portion of flank (Figure 7d) H. angusticaudata sp. nov.
4b. Absence of a series of thin slightly curved vertical bars (Figure 7b,c) 5
5a. Deep caudal peduncle (7.3%–9.2%SL) H. pauciradiatus
5b. Slender caudal peduncle (6.0%–7.2%SL) H. ansorgii
6a. Number of anal‐fin rays between the leading ray and the one directly below the leading dorsal‐fin ray 6–7 H. tangwenai sp. nov.
6b. Number of anal‐fin rays between the leading ray and the one directly below the leading dorsal‐fin ray 3–5 7
7a. Head profile rounded (Figure 6c) H. szaboi
7b. Head profile forming an obtuse angle with the rest of the dorsal body surface (Figure 6a) 8
8a. Slender caudal peduncle (5.0%–6.7%SL) H. xanekweorum sp. nov.
8b. Deep caudal peduncle (7.2%–8.2%SL) H. longilateralis
9a. 12–14 scales around the caudal peduncle H. pappenheimi
9b 16–18 scales around the caudal peduncle 10
10a. Slender caudal peduncle (5.9%–6.2%SL) H. chilembwei sp. nov.
10b. Deep caudal peduncle (6.9%–7.5%SL) H. ndauorum sp. nov.
DISCUSSION
5
Assignment of lineages to available names
5.1
Prior to this study, the genus Heteromormyrus was represented by three species in the Kwanza River system: H. ansorgii, H. pauciradiatus and H. pappenheimi. Sullivan et al. (2022) provided molecular evidence, showing that Heteromormyrus sp. ‘K4’, identified by Mutizwa et al. (2021), is conspecific with H. pauciradiatus. However, there were no genetic data available from the type material of H. ansorgii and H. pappenheimi. Therefore, the morphological analysis conducted in the present study was crucial for determining lineages that are likely to be conspecific with these two species. Specimens belonging to Heteromormyrus sp. ‘K3’ were morphologically comparable to the syntypes of H. ansorgii. We therefore concluded that this lineage likely represents H. ansorgii s.s, further supporting the previous determination by Mutizwa et al. (2021) that the Kwanza River system is the most likely type system for this species. We compared the morphological characteristics for the remaining lineages from the Kwanza River system with the original description and the syntypes of H. pappenheimi to determine if any of these lineages could be conspecific with this species. None of these lineages were morphologically comparable to H. pappenheimi, indicating that fresh material for this species was not included in the molecular study by Mutizwa et al. (2021). H. pappenheimi was originally described based on specimens that were collected at Cunga in the lower Kwanza River. Future surveys targeting the lower Kwanza River, a system that remains poorly sampled, are likely to reveal additional diversity within the genus Heteromormyrus and possibly find topotypes of H. pappenheimi. Future studies that will successfully extract and sequence DNA from the syntypes of H. ansorgii and H. pappenheimi, following the approach used by Sullivan et al. (2022), may offer a way to link these historical specimens to freshly collected specimens and provide insights on the evolutionary relationships of these species.
Mormyrid diversity in the Kwanza River basin
5.2
The ichthyofauna of the Kwanza River system remains poorly documented because exploration was curtailed by 27 years of civil unrest. However, this has changed over the past decade, where international support for environmental conservation and research has grown (Huntley & Ferrand, 2019). This growth is reflected by collaborative research efforts in the country soon after the end of the civil war and the subsequent signing of memoranda of understanding between the Angolan Ministry of Environment, the Instituto Superior de Ciências da Educação (ISCED) and the South African National Biodiversity Institute (SANBI), in 2009, which paved the way for further cooperative biodiversity projects, including training exercises and rapid biodiversity assessments (Huntley & Ferrand, 2019; Rejmánek et al., 2017). This has also facilitated important collaborative expeditions, such as Kwanza River project (2007–2009), supported by Instituto Nacional de Investigação Pesqueira (INIP) in Angola, the NRF‐SAIAB and, more recently, the National Geographic Okavango Wilderness Project (NGOWP 2018). A number of new species have been described based on these collections, with representatives in the Amphilidae, Amphilius pagei Thomson & Swartz, 2018, Cichlidae, Serranochromis cuanza Stauffer et al., 2021, Serranochromis swartzi Stauffer et al., 2021; Procatopodidae, ‘Lacustricola’ pygmaeus Bragança et al., 2021; and the Anabantidae, Microctenopoma steveboyesi Skelton et al., 2021. The present study has increased the number of mormyrid species in the Kwanza River from seven to nine. This number is likely to increase as more surveys are undertaken and other mormyrid groups examined, as evidenced by findings of more species in adjacent basins that have been relatively well studied compared to the Kwanza River system (e.g., Kramer et al., 2004, 2016; Kramer & Swartz, 2010; Kramer & van der Bank, 2011). This recent discovery of new fish species in the Kwanza River basin indicates that it could possibly be a previously overlooked biodiversity and endemic hotspot.
Diversity and distribution of Heteromormyrus in southern Africa
5.3
The species from the genus Heteromormyrus are found in the Congo, Kwanza, Kunene, Okavango, Zambezi, Pungwe and Buzi River basins. The distribution of these species across multiple rivers suggests historical riverine connections, with subsequent isolation resulting in allopatric speciation. The earliest common ancestor of the Mormyridae is estimated to have between 82.92 and 67.53 million years old (Ma) with the clade, including Heteromormyrus diverging around 15.1*–*11 Ma (Peterson et al., 2022). The clade that contained Heteromormyrus is thought to have rapidly diversified primarily in the Congo River basin (Peterson et al., 2022). The relatively close proximity of some of the southern tributaries of the Congo River to the northern tributaries of the Kwanza River, such as the Lucala River, as well as the shared fish fauna between these river systems, suggests an exchange of species between these rivers, probably by headwater capture (Skelton, 2019). The early exchange of the fish species between the Kwanza and Congo rivers, as well as subsequent speciation, may explain the high number of Heteromormyrus species in the Kwanza River, as well as the more basal lineages represented by some of these species (e.g., Heteromormyrus sp. ‘Inkisi River’, H. dolichorhynchus and H. pauciradiatus) (Mutizwa et al., 2021; Sullivan et al., 2022).
Some of the species in the Kwanza River system, for instance, H dolichorhynchus, H. pauciradiatus and H. ansorgii s.s. occur in sympatry. The EODs produced by mormyrids have been described as a key innovation that allows them to communicate with relatively fewer constraints from the environment, predators and competing signallers (Carlson & Arnegard, 2011). This results in communication with reduced distortion from the environment unlike other means of communication, for example, acoustic communication. Thus, mormyrids may be more sensitive to even small differences in pulse waveforms, for instance, between males and females or between sister species. This unique communication facilitates sexual selection and has been shown to evolve at a much faster rate compared to other traits, such as morphological and trophic ecological traits, making it an important early driver of species divergence even among sympatric species (Arnegard et al., 2005; Carlson & Arnegard, 2011; Feulner et al., 2007; Lavoué et al., 2008). The development of unique EODs may have driven the divergence of the multiple sympatric species within the Kwanza River. Although this study does not provide EOD evidence to test this hypothesis, evidence from H. szaboi in the upper Zambezi River shows that it consists of at least three morphologically indistinguishable lineages with relatively low genetic divergence, but with distinct EODs that are thought to be in the early stages of species divergence (Kramer et al., 2004; Sullivan et al., 2022). Further investigations into the factors shaping the diversity and distribution patterns of Heteromormyrus will need to integrate EODs, morphological and trophic ecological traits.
Conservation implications
5.4
Prior to this study, the taxonomy of the Heteromormyrus in southern Africa was poorly known, as there were only five valid species recognised in this region (H. pauciradiatus, H. ansorgii, H. szaboi, H. longilateralis and H. pappenheimi). H. ansorgii acted as catch‐all species name previously assigned to specimens of this genus collected from southern Africa. Consequently, this species was thought to have a distribution spreading across several southern African river systems, which resulted in it being classified as least concern on the International Union for Conservation of Nature (IUCN) Red list of threatened species (Bills et al., 2018). H. pappenheimi and H. szaboi are considered to be of least concern due to presumed wide distribution ranges in a habitat considered to be healthy, whereas H. pauciradiatus is classified as data deficient due to the species being known from a single site prior to this study (Da Costa, 2007; Tweddle, 2007). This illustrates the critical gap in knowledge of many of the freshwater fish species inhabiting this river system. This study described six new species that were previously included under a single species, H. ansorgii, indicating that the conservation statuses of these species will need to be determined.
Because species in the genus Heteromormyrus are associated with lotic habitats in perennial streams and rivers (Ford & Albert, 2022), they are likely to face similar threats associated with these habitats. In southern Africa, increased anthropogenic pressure from impoundments and water abstraction altering the natural river flow (e.g., Kwanza and Kunene River), alteration of the riparian zone and chemical inputs from the farming activities (e.g., Ruo River), illegal mining activities, deforestation and introduction of non‐native fish species (e.g., Kwanza, Pungwe and Buzi Rivers) have been identified as some of the major threats to the aquatic fauna (Chakona et al., 2018; de Moor et al., 2000; Kadye & Chakona, 2013; Skelton, 2019). The ecology of the species identified in this study remains as a gap that future studies need to address to provide an understanding of how these threats impact the species and to assist with formulating effective conservation recommendations.
CONCLUSION
6
This study provided formal descriptions for six new species from the genus Heteromormyrus based on the genetic evidence provided by Mutizwa et al. (2021) and the morphological data from the present study. Two of the new species H. dolichorhynchus and H. angusticaudata were described from the Kwanza River, whereas H. xanekweorum, H. chilembwei, H. tangwenai and H. ndauorum were each described from the Okavango, Ruo, Pungwe and Buzi rivers, respectively. The description of six new species from the H. ansorgii species complex shows that the previous wide distribution pattern attributed to this species was due to incomplete taxonomic knowledge. The formal recognition of these morphologically similar species is a fundamental first step for further research and conservation efforts involving these species. Additional taxonomic work within this genus remains essential, as the limited number of samples prevented further exploration into the taxonomic statuses of other undescribed species of the genus Heteromormyrus (e.g., H. sp. ‘K2’ from the Kwanza, H. sp. ‘UZ1’ and ‘HaZ’ from the Zambezi River, ‘HaK’ from the Kwando River and H. sp. ‘Insiki River’ from the Insiki River) identified in other studies of this group (Kramer et al., 2004; Mutizwa et al., 2021; Sullivan et al., 2022). H. pappenheimi remains a valid species from the Kwanza River; however, there is a need for collection of additional voucher and genetic material for this species to define its phylogenetic relationships with its congeners, as well as providing an updated species description. Despite the limitations in addressing some of the known lineages, this study increased the number of described mormyrid species within southern Africa, as well as reduced some of the ambiguity surrounding the overall diversity of freshwater fish diversity in this region.
AUTHOR CONTRIBUTIONS
Project conceptualisation: Albert Chakona and Wilbert T. Kadye; data generation: Tadiwa I. Mutizwa and Pedro H. N. Bragança; data analysis: Tadiwa I. Mutizwa; manuscript preparation: Albert Chakona, Pedro H. N. Bragança, Tadiwa I. Mutizwa and Wilbert T. Kadye; funding: Wilbert T. Kadye and Albert Chakona.
FUNDING INFORMATION
This research was supported by the Rhodes University Sandisa Imbewu Grant, the NRF‐Research Development Grant (CSRP190416431023), NRF‐SAIAB Refresh project (FBIP‐211006643719) and NRF‐SAIAB topotypes project (IBIP‐BS 13100251309).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arnegard, M. E. , Bogdanowicz, S. M. , & Hopkins, C. D. (2005). Multiple cases of striking genetic similarity between alternate electric fish signal morphs in sympatry. Evolution, 59, 324–343.15807419 · pubmed ↗
- 2Arnegard, M. E. , & Hopkins, C. D. (2003). Electric signal variation among seven blunt‐snouted Brienomyrus species (Teleostei: Mormyridae) from a riverine species flock in Gabon, Central Africa. Environmental Biology of Fishes, 67, 321–339.
- 3Arnegard, M. E. , Mc Intyre, P. B. , Harmon, L. J. , Zelditch, M. L. , Crampton, W. G. R. , Davis, J. K. , Sullivan, J. P. , Lavoué, S. , & Hopkins, C. D. (2010). Sexual signal evolution outpaces ecological divergence during electric fish species radiation. The American Naturalist, 176, 335–356.10.1086/65522120653442 · doi ↗ · pubmed ↗
- 4Bell‐Cross, G. , & Minshull, J. L. (1988). The fishes of Zimbabwe. National Museums and Monuments of Zimbabwe.
- 5Bills, R. , Kazembe, J. , Marshall, B. E. , Moelants, T. , Tweddle, D. , & Vreven, E. J. W. M. N. (2018). Hippopotamyrus ansorgii. The IUCN Red List of Threatened Species, 8235.
- 6Boulenger, G. A. (1905). Descriptions of four new freshwater fishes discovered by Dr. WJ Ansorge in Angola. The Annals and Magazine of Natural History Including Zoology, Botany and Geology, 15, 457–459.
- 7Boulenger, G. A. (1910). On a large collection of fishes made by Dr. WJ Ansorge in the Quanza and Bengo Rivers, Angola. Annals and Magazine of Natural History, 6, 537–561.
- 8Bragança, P. H. N. , Skelton, P. H. , Bills, R. , Tweddle, D. , & Chakona, A. (2021). Revalidation and redescription of “Lacustricola” chobensis (fowler, 1935) and description of a new miniature species of “Lacustricola” from southern Africa (Cyprinodontiformes: Procatopodidae). Ichthyology & Herpetology, 109, 123–137.