Microbial biodiversity in Tunisian olive grove soils: a reservoir of phytopathogenic fungi and potential beneficial microorganisms
Meryam Belhedi, Besma Sghaier-Hammami, Sofienne B. M. Hammami, Samar Ben Slema, Palmira De Bellis, Stefania Somma, Haithem Nafati, Khaled Hibar, Charlie Abi Saad, Antonio Moretti, Mario Masiello

TL;DR
This study explores the diversity of fungi and bacteria in Tunisian olive grove soils, identifying harmful fungi and beneficial bacteria that could help protect plants.
Contribution
The study provides new insights into the microbial biodiversity of olive grove soils and identifies potential biocontrol agents against phytopathogenic fungi.
Findings
Fusarium species dominated the fungal community, with varying virulence across different intercropping systems.
Bacillus species and Priestia megaterium were the most common bacteria, showing antagonistic activity against pathogenic fungi.
Bacillus amyloliquefaciens inhibited fungal growth through biofilm formation and hyphal alteration.
Abstract
Intercropping in olive orchards increases the risk of soil-borne fungal infections, particularly when associated crops are susceptible to the same pathogens. This study aimed to identify soil-borne microorganisms colonizing the roots and rhizosphere of olive trees in Tunisia intercropped with Solanaceae plants and to evaluate co-occurring bacterial communities for their potential to mitigate wilt disease and promote plant health. Endophytic fungi and bacteria were isolated from olive soils and roots collected from three olive orchards subjected to different intercropping systems. Fungal strains were molecularly identified at the species level using Internal Transcribed Spacer (ITS) and translation elongation factor 1-α (TEF1) gene sequencing, while bacterial strains were characterized by rep-PCR profiling and 16S rDNA sequencing. The pathogenicity of selected Fusarium strains was…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
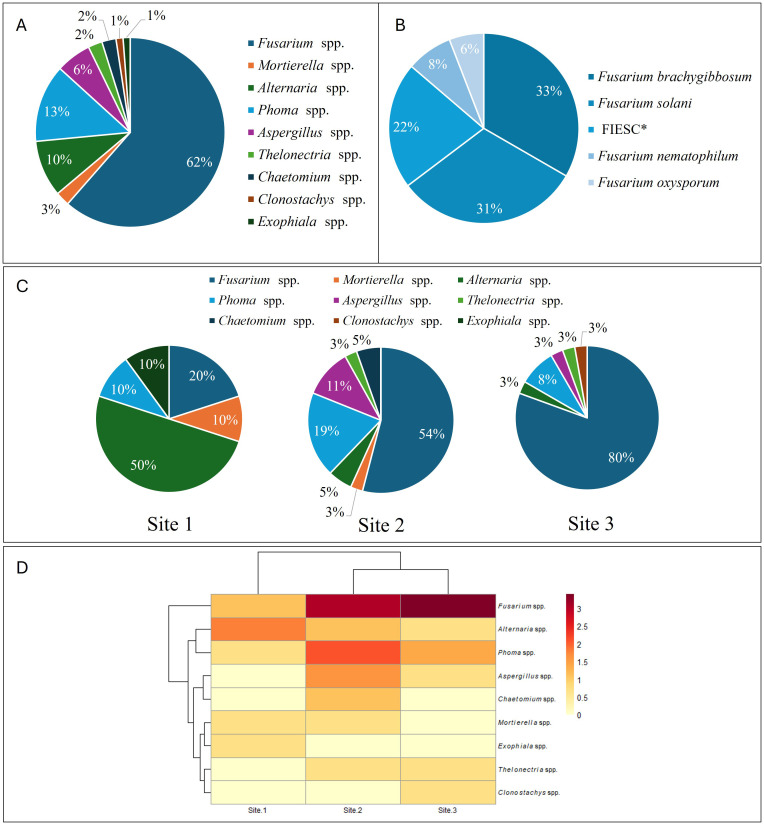 Figure 1
Figure 1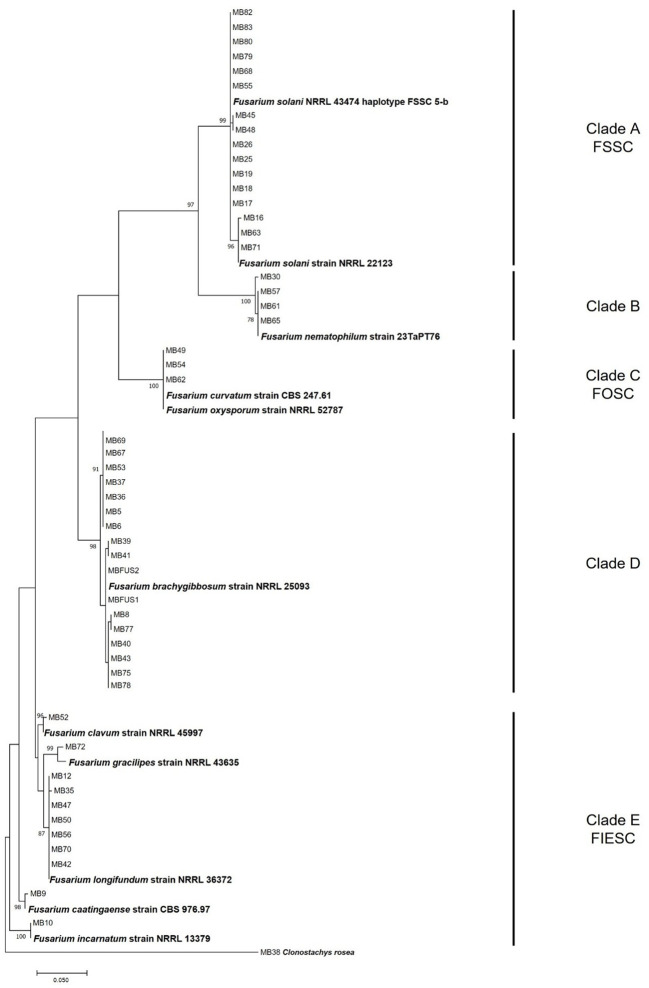 Figure 2
Figure 2 Figure 3
Figure 3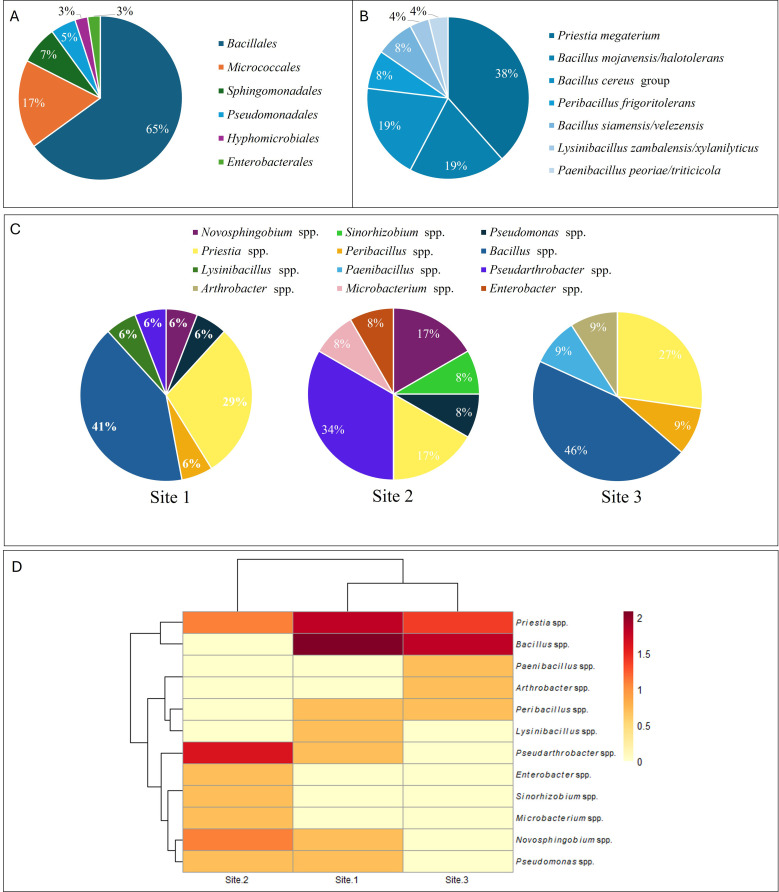 Figure 4
Figure 4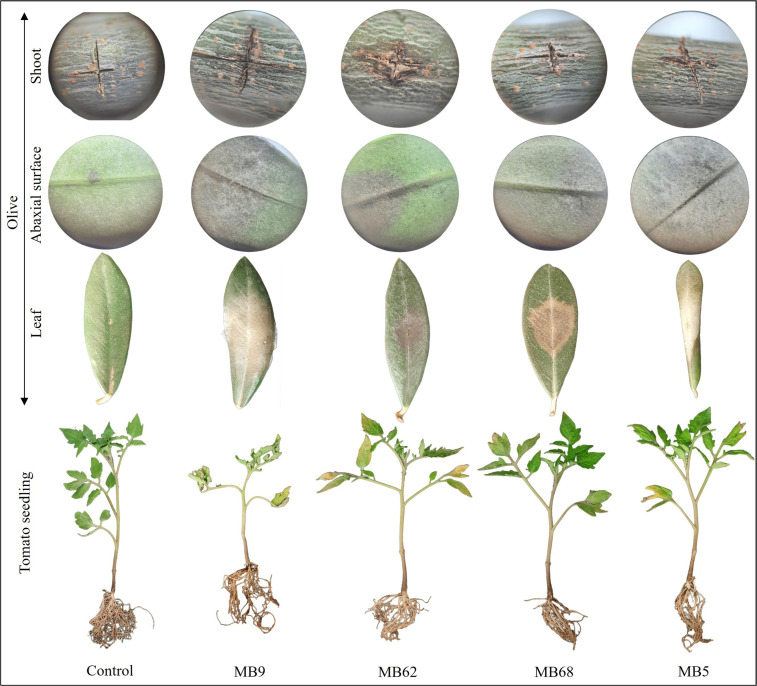 Figure 5
Figure 5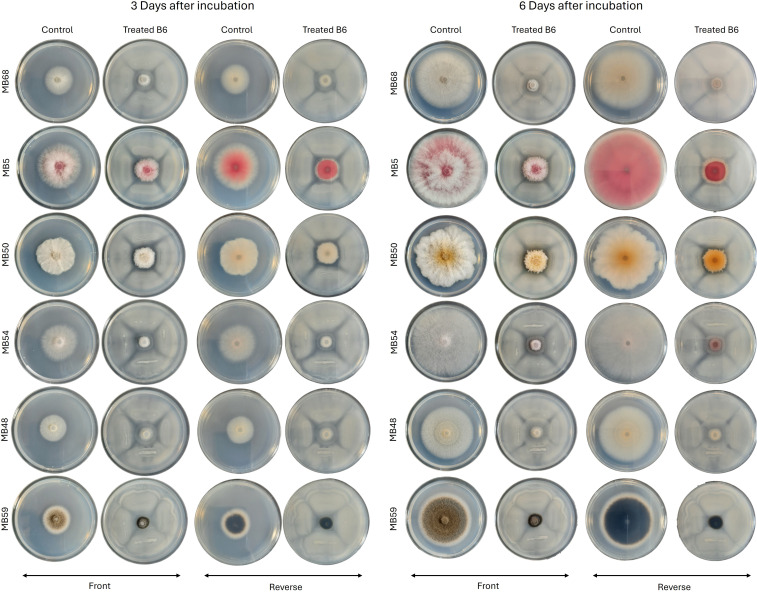 Figure 6
Figure 6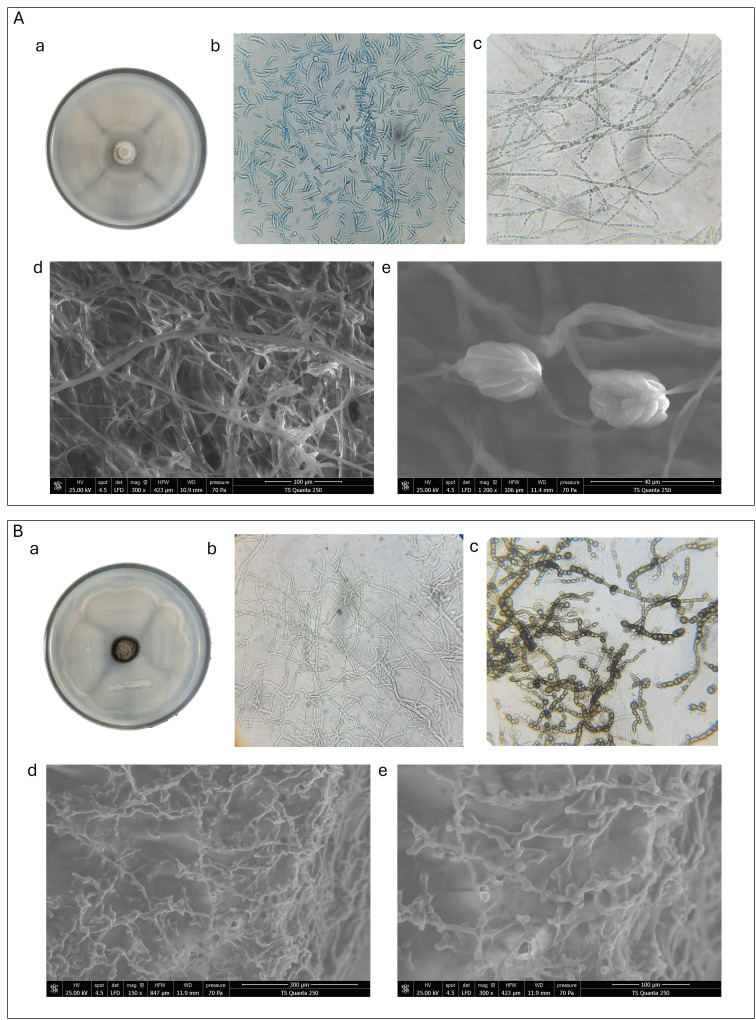 Figure 7
Figure 7| Strain | Fungal species | Tomato | Olive | |||
|---|---|---|---|---|---|---|
| DSI (%) | % Infected leaves | DSI (%) | % Infected leaves | % Infected stems | ||
| MB50 |
| 24.4 ± 5.9 cde | 67 | 3.6 ± 1.9gh | 20 | 50 |
| MB56 | 20.9 ± 3.1 ef | 50 | 14.8 ± 0.8e | 40 | 50 | |
| MB72 |
| 27.1 ± 0.4 e | 67 | 11.8 ± 1.0ef | 60 | 50 |
| MB9 |
| 80.0 ± 0.0 a | 100 | 28.2 ± 1.0c | 60 | 50 |
| MB54 |
| 11.3 ± 0.7 fg | 50 | 23.0 ± 1.5d | 100 | 100 |
| MB62 | 51.0 ± 2.1 b | 100 | 20.0 ± 0.7d | 80 | 75 | |
| MB5 |
| 23.0 ± 1.5 de | 50 | 51.8 ± 1.0a | 60 | 75 |
| MB41 | 0.0 ± 0.0 h | 0 | 8.2 ± 1.0fg | 40 | 75 | |
| MB43 | 0.0 ± 0.0 h | 0 | 45.2 ± 0.8b | 40 | 100 | |
| MB37 | 5.2 ± 0.8 gh | 33 | 5.2 ± 0.8g | 0 | 25 | |
| MB25 |
| 34.0 ± 1.0 c | 50 | 14.8 ± 0.8e | 60 | 100 |
| MB48 | 20.0 ± 2.9 ef | 33 | 28.3 ± 0.9c | 60 | 100 | |
| MB63 | 33.3 ± 2.7 cd | 60 | 20.0 ± 0.8d | 60 | 0 | |
| MB68 | 33.0 ± 2.5 cd | 50 | 20.0 ± 1.0d | 80 | 0 | |
| MB57 |
| 0.0 ± 0.0h | 0 | 0 ± 0h | 0 | 0 |
| MB20 | 0.0 ± 0.0 h | 0 | 13.4 ± 0.9e | 60 | 0 | |
| MB59 | 20.3 ± 2.6 ef | 25 | 8.3 ± 0.9fg | 40 | 75 | |
| MB24 | 0.0 ± 0.0 h | 0 | 3.6 ± 1.0gh | 20 | 50 | |
| MB86 | 0.0 ± 0.0h | 0 | 0 ± 0h | 0 | 0 | |
| Bacterial strain | Bacterial species | Mycelial growth inhibition (%) of | |||
|---|---|---|---|---|---|
| Antagonism | Indirect (VOCs) | Antagonism | Indirect (VOCs) | ||
| S1C1 |
| 38.7 ± 3.3 a | 38.8 ± 4.4 a | 41.2 ± 2.6 b | 52.6 ± 3.4 a |
| S1C12 |
| 36.0 ± 2.5 ab | 27.3 ± 5.5 ab | 28.2 ± 0.0 cde | 46.8 ± 1.4 a |
| S3C12 |
| 41.2 ± 2.5 a | 25.8 ± 2.7 ab | 29.8 ± 3.3 bcd | 29.6 ± 2.9 b |
| S3C1 |
| 37.9 ± 1.1 ab | 0.0 ± 0.0 c | 29.79 ± 3.6 bcd | 26.3 ± 2.9 bc |
| S1C17 | 29.0 ± 2.1 bc | 0.0 ± 0.0 c | 21.0 ± 0.8 cdef | 22.4 ± 3.1 bc | |
| S3C4 |
| 29.6 ± 2.6 bc | 14.3 ± 1.2 b | 18.6 ± 2.0 defg | 15.2 ± 0.5 c |
| B6 |
| nd | nd | 67.5 ± 0.5 a | 0 ± 0 d |
| S1C19 |
| 21.5 ± 0.5 d | nd | 13.6 ± 3.3 fg | nd |
| S1C3 |
| 19.6 ± 1.0 d | nd | 31.6 ± 5.5 bc | nd |
| S1C5 |
| 20.6 ± 0.9 cd | nd | 16.2 ± 3.2 efg | nd |
| S3C2 |
| 37.5 ± 0.0 ab | nd | 31.9 ± 2.5 bc | nd |
| S3C3 |
| 24.4 ± 0.6 cd | nd | 8.2 ± 1.6 g | nd |
| S2C17 | 26.2 ± 0.6 cd | nd | 13.8 ± 1.7 fg | nd | |
| S2C7 |
| 21.9 ± 1.1 cd | nd | 14.1 ± 0.3 fg | nd |
| S1C6 |
| 23.3 ± 0.5 cd | nd | 15.7 ± 2.1 efg | nd |
| S2C20 |
| 22.7 ± 1.2 cd | nd | 6.1 ± 2.8 g | nd |
| S3C8 |
| 19.6 ± 1.3 d | nd | 7.7 ± 0.7 g | nd |
| S1C8 |
| 37.1 ± 2.7 ab | nd | 17.0 ± 1.6 efg | nd |
| S2C12 |
| 22.1 ± 0.5 cd | nd | 18.6 ± 0.5 defg | nd |
| S3C6 | 38.7 ± 0.6 a | nd | 27.7 ± 1.2 cde | nd | |
| Incubation period | MB50 | MB5 | MB59 | MB68 | MB48 |
|---|---|---|---|---|---|
| 3 DAI | 57.5 ± 2.65a | 53.8 ± 0.593a | 48.9 ± 1.84ab | 29.9 ± 0.941bc | 19.8 ± 1.10c |
| 6 DAI | 74.7 ± 1.20a | 68.9 ± 0.486a | 69.0 ± 0.617c | 50.8 ± 0.869d | 46.3 ± 0.333e |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAgronomic Practices and Intercropping Systems · Plant-Microbe Interactions and Immunity · Mycorrhizal Fungi and Plant Interactions
Introduction
1
Olive (Olea europaea L.), one of the oldest domesticated fruit trees, represents an important crop in the Mediterranean Basin, accounting for almost 90% of the olive cultivation area worldwide. Tunisia alone cultivates about 1.9 million hectares and contributes around 7% of global olive oil production (FAOSTAT, 2024). Olive groves represent a relatively stable agroecosystem due to the ability of olive trees to adapt to a wide range of environmental conditions. Their resilience is also supported by the wide occurrence of beneficial microorganisms in the soil where they grow, which enhance tolerance to biotic and abiotic stresses, especially those occurring in the rhizosphere (Fernández-González et al., 2019). However, the rhizosphere of olive groves is threatened by several soil-borne pathogens, among which Verticillium dahliae is the most frequently reported and is considered the most important fungal pathogen affecting olive trees (Jiménez-Díaz et al., 2012; Keykhasaber et al., 2018; López-Escudero and Mercado-Blanco, 2011). In recent years, Fusarium species have emerged as additional pathogens associated with olive dieback in several countries worldwide (Nicoletti et al., 2020), including Morocco (Chliyeh et al., 2014), Spain (Hernández et al., 1998), Greece (Markakis et al., 2021), and Tunisia (Belhedi et al., 2025; Gharbi et al., 2020; Trabelsi et al., 2017). This rise can be attributed to changes in olive cultivation methods, the expansion of new plantations into Fusarium-infected areas, and the use of infected planting material. Indeed, the growing global demand for olive oil is leading to planting new olive orchards in the Mediterranean regions and a shift from traditional rainfed agriculture to high-density system (Cardoni and Mercado-Blanco, 2023). The new agronomic practices, together with climate changes, could drive the spread of different soil-borne pathogens, the incidence and the severity of olive diseases (Benítez-Cabello et al., 2023; Rallo et al., 2013). In addition, the cultivation of Solanaceae plants (e.g., tomato, pepper, eggplant) alongside olive has been reported to enhance the persistence of soil-borne fungi by providing alternative susceptible hosts (Gharbi et al., 2020; Nigro et al., 2005). Such intercropping practices may facilitate pathogen adaptation and transfer from annual to perennial hosts, thereby intensifying disease pressure in olive orchards. In Tunisia, however, this interaction remains poorly investigated, and the role of intercropped Solanaceae plants on olive tree diseases is still unclear.
Olive disease management relies mainly on agronomic measures, including the use of certified planting material, appropriate pruning, and improved soil and irrigation management (Fernández-Escobar et al., 2013). Yet, intensive systems and heavy agrochemical use can disrupt the natural soil microbiome (Bizos et al., 2020; Montes-Osuna and Mercado-Blanco, 2020; Zipori et al., 2021). Beneficial microbial species, belonging to Bacillus, Pseudomonas, Paenibacillus, Trichoderma, and Glomus genera, play crucial roles in plant health by promoting growth, enhancing nutrient uptake, and suppressing pathogens (Aloo et al., 2019; Mercado-Blanco and Lugtenberg, 2014). For instance, Bacillus velezensis and some Pseudomonas species isolated from olive rhizospheres inhibit Fusarium spp. and V. dahliae, through biofilm formation and production of siderophores (Cheffi et al., 2019; Mercado-Blanco et al., 2004).
A comprehensive knowledge of microbial communities associated to the olive root system, including the root endosphere and the rhizosphere soil, as well as knowledge on the influence of biotic and abiotic factors on these communities is crucial for selecting specific fungi and bacteria that enhance olive traits under specific conditions. This study aimed to provide a picture of the microbial communities of olive groves intercropped with Solanaceae plants in Tunisia, to characterize fungal pathogens isolated from the roots of olive trees showing wilt symptoms, and to select beneficial microorganisms with potential biocontrol properties.
Materials and methods
2
Origin of the samples and microorganisms’ isolation
2.1
Three different olive groves (Chemlali cultivar), located in two different regions and characterized by different intercropping systems, were considered. Site 1, located in Kairouan region, included forty-year-old olive trees intercropped with potato plants (Solanum tuberosum L.), continuously cultivated in this field for seven crop seasons. Site 2, located in Hergla, in Sousse region, consisted of ten-year-old olive trees intercropped with potato plants, cultivated in this field for thirty consecutive years. Site 3, located in Sidi Bou Ali, in Sousse region, included young olive trees intercropped with tomato (Solanum lycopersicum L.) and pepper (Capsicum annuum L.). At each site, four parcels were selected, and five trees were sampled per parcel (20 trees per site), with each tree considered as one biological replicate. Sampling was conducted after the harvest of the intercropped crops. For each tree, approximately 1 L of soil containing roots was collected at the projection of the canopy edge (drip line), which corresponds to the main rhizosphere interaction zone between olive tree roots and the associated intercropped vegetation. This zone was deliberately chosen to characterize microbial communities shaped by intercropping practices and root–root interactions. Soil samples were collected from four equidistant points around each tree and pooled to generate one composite sample per tree. Roots were subsequently separated from the soil by sieving. Sampling followed a completely randomized design arranged along a Z-shaped transect within each parcel.
After washing with running tap water, portions of roots were surface disinfected by dipping in 2% sodium hypochlorite solution for 2 minutes, followed by three rinses in sterile distilled water. Small pieces of root of about 3 mm diameter were transferred on Potato Dextrose Agar (PDA) and Malt Extract Agar (MEA), both amended with streptomycin sulphate (100 mg L^-1^) and neomycin sulphate (50 mg L^-1^). For each soil sample, 1.5 g was transferred to a sterile microbiological tube containing 13.5 mL of sterile distilled water and shaken for 30 min at 150 rpm in an orbital shaker. Aliquots (100 µL) of three serial dilutions were spread on Plate Count Agar (PCA). All plates were incubated at 25 ± 1°C for 5 days under an alternating 12-hour light/dark cycle. After incubation, fungal and bacterial colonies, selected on the basis of their morphological characteristics, were purified for further analysis. Each fungal colony was purified by spreading conidial suspension on Water Agar (WA) and, after 18 hours of incubation at 25 ± 1°C, a single germinated spore was collected under dissecting microscope and transferred on new PDA plate. To obtain pure bacterial cultures, single colonies were picked up and purified by repeated streaking on PCA medium.
Molecular identification of isolated microorganisms
2.2
Fungal species identification
2.2.1
Mycelium of 3-day-old colonies, grown on sterilized cellophane disks overlaid on PDA plates, was collected by scraping. Genomic DNA was extracted and purified from powdered lyophilized mycelia (each sample, 10–15 mg) using the Wizard Magnetic DNA Purification System for Food kit (Promega, Madison, WI, USA), according to the manufacture’s protocol. The quantity and integrity of DNA were checked by comparison with a standard 1 kb DNA Ladder (Fermentas GmbH, St. Leon-Rot, Germany) on 0.8% agarose gel, after electrophoretic separation.
For each selected strain, an informative fragment of the Internal Transcribed Spacer (ITS) was amplified and sequenced using the primer pair ITS-4/ITS-5 (White et al., 1990). In addition, all fungal strains belonging to Fusarium genus, were molecularly characterized by sequencing the translation elongation factor 1-α (TEF1) gene, commonly reported as the most informative gene for Fusarium identification to species level (Geiser et al., 2004), using the primer pair EF1/EF2 (O’Donnell et al., 1998). The Polymerase Chain Reaction mixture (15 μL) contained 30 ng of genomic DNA, 0.45 μL of each primer (10 µM), 1.2 μL of dNTPs (10mM), 1.5 µL of 10x buffer and 0.125 μL of Hot Start Taq DNA Polymerase (Fisher Molecular Biology, 1 U μL^-1^, Rome, Italy). The amplifications were carried out in a Mastercycler EP Gradient thermal cycler (Eppendorf, Hamburg, Germany). ITS region was amplified using the following conditions: initial denaturation of 50 sec at 72°C, followed by 40 cycles of 30 sec at 95°C, 30 sec at 52°C, 50 sec at 72°C, and a final extension of 5 min at 72°C. The TEF1 fragment gene was amplified using the following parameters: initial denaturation of 2 min at 95°C, followed by 35 cycles of 50 sec at 95°C, 50 sec at 59°C, 1 min at 72°C, and a final extension of 7 min at 72°C.
The PCR products, stained with GelRed^®^ (Biotium Inc., Fremont, CA, USA), were checked after electrophoretic separation on 1.5% agarose gel in 1X TAE buffer under UV light by comparison with a 100 bp DNA Ladder (ThermoFisher Scientific, Waltham, Massachusetts, USA). Before sequencing, the PCR products were purified with an enzymatic EXO/FastAP mixture (Exonuclease I and FastAP thermosensitive alkaline phosphatase, ThermoFisher Scientific, Waltham, Massachusetts, USA). Sequence reactions were performed for both strands using a BigDye Terminator v3.1 Cycle Sequencing Ready Reaction Kit (Applied Biosystems, Foster City, CA, USA) according to the manufacturer’s recommendations. The labeled products were purified by filtration through Sephadex G-50 (5%) (Sigma-Aldrich, Saint Louis, MO, USA) and analyzed using an ABI PRISM 3730 Genetic Analyzer (ThermoFisher Scientific, Waltham, Massachusetts, USA). The FASTA sequences obtained were analyzed and assembled using BioNumerics v. 5.1 software (Applied Maths, Kortrijk, Belgium). For molecular identification at the genus or species level, the ITS obtained sequences were compared with the sequences available in the NCBI Database through the BLASTN program.
Nucleotide sequences of TEF1 gene, joined to sequences of species reference strains retrieved from GenBank, were aligned using the ClustalW algorithm (Thompson et al., 1994). Phylogenetic relationships were studied using the maximum likelihood method with MEGA software version 7 (Kumar et al., 2016). The bootstrap analyses (Felsenstein, 1985) were conducted to determine the confidence of internal nodes using a heuristic search with 1000 replicates, removing gaps. The field strain MB38, identified as Clonostachys rosea, was used as outgroup taxon.
Bacterial species identification
2.2.2
Genomic DNA was extracted using the Wizard Genomic DNA Purification Kit (Promega, Madison, WI, USA) following the manufacturer’s protocol. All isolated bacterial strains were molecularly characterized through rep-PCR assay by using the primer pair REP-1R-Dt (5 –IIINCGNCGNCATCNGGC–3) and REP-2R-Dt (5 –NCGNCTTATCNGGCCTAC– 3) (Hyytiä-Trees et al., 1999). The PCR reactions were carried out in a total volume of 25 μl, containing 23 μL of Mega Mix (Microzone Ltd., United Kingdom), 0.5 μL of each primer (10 mM) and 1 μl of genomic DNA (15 ng). Amplifications were performed in a Mastercycler EP Gradient thermal cycler (Eppendorf, Hamburg, Germany) as reported by Zicca et al. (2020). Amplicons were separated by microfluidic electrophoresis using the DNA7500 LabChip kit with the Agilent 2100 Bioanalyzer (Agilent Technologies, Waldbronn, Germany), according to the manufacturer’s instructions. The rep-PCR profiles were analyzed and compared using the software provided by the same company.
All bacterial strains were grouped on the basis of their rep-PCR profile and for each different profile, a representative strain was identified by 16S rRNA gene sequencing using the universal bacterial primers 27f-YM and 1492r (Frank et al., 2008). PCR reactions were carried out in a 50 μl mixture containing 5 μl of 10× AccuPrime™ Pfx Reaction Mix, 1.25 U of AccuPrime™ Pfx DNA polymerase (Thermo Fisher Scientific, Waltham, Massachusetts, USA), 0.3 μM of each primer, and 1 μl of genomic DNA. The amplification program consisted of an initial denaturation at 95°C for 2 min, followed by 35 cycles of denaturation at 95°C for 30 s, annealing at 60°C for 5 cycles, 55°C for 5 cycles, and 48°C for 25 cycles, with extension at 68°C for 3 min, and a final extension at 68°C for 10 min. PCR products were sequenced using the BigDye™ Terminator cycle sequencing kit on an ABI Prism 3730xl DNA Analyzer (Thermo Fisher Scientific, Waltham, Massachusetts, USA). Sequences were assembled with BioNumerics v. 5.1 (Applied Maths, Austin, Texas, USA) and compared to type strain sequences available in the NCBI database (BLASTN, “Sequences from type material” option). Bacterial strains were tentatively assigned to a bacterial species on the basis of the highest score of the alignment and the percentage of identity between their 16S rRNA gene sequences and those of type strains in the database.
In vitro pathogenicity test
2.3
To assess the potential pathogenicity of the isolated fungal species on olive tree and Solanaceae plants, conidial suspensions of selected fungal strains, chosen based on molecular identification, were used to inoculate detached leaves and stems of olive tree seedlings, as well as one-month-old tomato seedlings. Olive leaves were placed in rectangular Petri dishes on moistened sterile paper. Aliquots of 20 µl of conidial suspension, adjusted to 10^6^ spores mL^-^¹, were injected into light incision made in the middle of each leaf. For olive stems, portions of about 110 mm and 25 mm in diameters were placed in Petri dishes. An incision, 10 mm deep, was made in the bark of each stem and inoculated with the same conidial suspension used for the leaves. A negative control, leaves and stems injected with sterile distilled water, was also considered. To assess the pathogenicity on Solanaceae plants, tomato seedlings were uprooted, their roots washed with tap water and then immersed for 4 days in the previously prepared conidial suspensions. Tomato seedlings immersed in sterile distilled water were used as negative control. All experiments were performed on five replicates.
After seven days of incubation at room temperature with a photoperiod of 12h, the number of olive leaves and stems and tomato seedlings showing symptoms on the total number of inoculated ones was recorded. Disease severity on olive and tomato leaves was visually assessed based on the extent of leaf yellowing and wilting, following the empirical scale: 0 = no visible symptoms; 1 = yellowing or wilting affecting up to 1/3 of the leaf; 2 = yellowing or wilting affecting 1/2 of the leaf; 3 = yellowing or wilting affecting 2/3 of the leaf; 4 = yellowing or wilting affecting the whole leaf; and 5 = complete necrosis or rolling of the leaf or leaf drop.
Disease Severity Index (DSI) was calculated using the formula:
where n is the number of leaves in each severity category, v is the severity rating, N is the total number of assessed leaves, and V is the maximum severity rating. Pathogenicity on the stems was assessed by counting the number of infected stems on the total ones. A stem was considered infected when a visibly hollow surrounding the point of conidial suspension injection was observed.
In vitro antagonist assay using bacteria
2.4
The antagonistic activity of identified bacterial strains was evaluated against two Fusarium strains, MB1 (F. brachygibbosum) and MB54 (F. oxysporum). The capability of these bacteria to inhibit mycelial growth was assessed by a dual culture assay on PCA medium (Zhao et al., 2014). In detail, a 6-mm mycelial plug collected from the margin of actively growing colony of each Fusarium strain was placed at the center of the plate, and two drops (5 µL each) of the bacterial suspension (10^9^ CFU/mL) were inoculated 1 cm from the fungal plug. Control plates consisted of fungal plug without bacteria. All treatments were replicated three times and incubated for seven days at 25°C, under a photoperiod of 12h.
To evaluate the antagonistic activity, fungal colony diameters were measured every two days and used to calculate the percentage of mycelial growth inhibition.
The mycelial growth inhibition, expressed as percentage, was calculated using the following formula:
Where Dc is the mean value of the two orthogonal diameters of the control colony and Dt is the mean value of the two orthogonal diameters of the fungal colonies’ growth in presence of bacterial strains.
Bacterial–fungal interactions via volatile organic compounds (VOCs)
2.5
Bacterial strains were used to assess their capacity to produce VOCs, capable of inhibiting mycelial growth. According to the method reported by Wu et al. (2019), with some modifications, an overnight bacterial culture in TSB broth was spread onto a PDA plate. The lid was replaced by a base plate of PDA containing a 4 mm diameter agar plug from an actively growing fungal mycelium of Fusarium. The two PDA base plates were sealed together with Parafilm. Control sets were prepared without bacteria in the bottom plate. The diameters of the fungal colonies, on three replicates, were measured after 7 days incubation at 25°C and used to calculate the mycelial growth inhibition as reported above.
Microscopic and SEM observations
2.6
After seven days of incubation, the effect of the Bacillus bacteria on the mycelium of the tested fungal strains was examined under an optical microscope. For high-resolution imaging, a Thermo Scientific™ Quanta™ 250 FEI scanning electron microscope (SEM) equipped with an Energy-Dispersive X-ray Spectroscopy (EDX) system was used (Thermo Fisher Scientific Inc., Waltham, MA, USA). Scanning electron microscopy was used to visualize sample surfaces at micro- to nanometer resolution. The electron beam interacts with the material to generate signals that reveal surface morphology, while the integrated EDX enabled qualitative and quantitative elemental analysis. Analyses were carried out under low-vacuum conditions to preserve the integrity of biological and non-conductive samples. Imaging was performed at different magnifications and in low-voltage mode to reduce damage and improve penetration.
Statistical analysis
2.7
The data were analyzed using R version 4.3.2. A one-way analysis of variance (ANOVA) was performed to evaluate the effects of bacterial strains on fungal mycelial growth and separately, the effect of fungal strains on plant material in pathogenicity assays. Differences between means were assessed using Tukey’s test with a significance threshold of p< 0.05. Results were presented in tables as means ± standard error, with homogeneous groups indicated. For clustering and heatmap visualization, abundance data were log transformed to reduce the influence of highly abundant taxa, using the ‘pheatmap’ package in R.
Results
3
Fungal species occurring in olive roots and rhizospheres
3.1
Molecular identification of 83 fungal strains, based on ITS sequence analyses, revealed a great biodiversity of fungal species occurring in olive roots and rhizosphere, in Tunisia (Supplementary Table 1; Figure 1A). The majority of the strains (62%) belonged to Fusarium genus, followed by Phoma (13%) and Alternaria (10%) genera. At lesser extent, few strains identified as Aspergillus, Chaetomium, Exophiala, Mortierella, and Thelonectria were also identified (Supplementary Table 1, Figure 1A).
(A) Distribution of fungal genera, identified by ITS region sequencing from olive root–associated isolates pooled across all sampled olive orchards in Tunisia; (B) Distribution of Fusarium species, molecularly identified by ITS and TEF1 gene sequencing on olive roots in Tunisia, representing the Fusarium spp. fraction shown in panel (A), FIESC, Fusarium incarnatum equiseti Species Complex: Fusarium longifundum, Fusarium clavum, Fusarium gracilipes, Fusarium caatingaense; (C) Distribution of fungal isolates according to sampling site, site 1 (Kairouan), site 2 (Hergla, Sousse) and 3 (Sidi Bou Ali, Sousse). (D) Heat map of fungal strains abundance according to sampling site. The rows represent the isolated fungal strains, and the columns represent the sampling site. Color scale represents log-transformed abundance values.
Molecular identification of Fusarium species strains, based on ITS and TEF1 gene sequencing, revealed the presence of several Fusarium species on olive groves, in Tunisia, including F. solani, F. nematophilum, F. oxysporum, F. brachygibbosum and members of Fusarium incarnatum-equiseti Species Complex (FIESC), namely F. caatingaense, F. clavum, F. gracilipes. F. incarnatum, and F. longifundum (Figure 1B).
The fungal species distribution varied across the three sampling sites. In Site 1 (Kairouan), Alternaria genus was the most dominant (50% of the total isolated strains) followed by Fusarium (20%) and an even distribution of Phoma, Exophiala and Mortierella (10% each; Figure 1C). In Site 2 (Hergla, Sousse), Fusarium was the predominant genus (54%), followed by Phoma (19%) and Aspergillus (11%). In Site 3 (Sidi Bou Ali, Sousse), Fusarium was overwhelmingly dominant (80%), followed by Phoma (8%), whereas other fungal genera were detected to a very less extent (Figure 1C). Clustering analysis revealed closer similarities between the fungal composition of Hergla and Sidi Bou Ali compared with Kairouan, which showed a distinct profile (Figure 1D).
The phylogenetic analysis including the TEF1 sequences of 51 Fusarium strains isolated from olive groves, 11 Fusarium reference sequences, and the sequence of Clonostachys rosea (MB38) used as an outgroup taxon, was shown in Figure 2. The phylogenetic tree was resolved in 5 well‐separated clades, supported by high bootstrap values. In clade A, sixteen strains are clustered together with the F. solani reference strains NRRL 22123 and NRRL 43474. In particular, three strains MB16, MB63 and MB71 showed high homology with F. solani NRRL 22123, while the other 13 Fusarium strains, showing 100% homology among them, grouped with F. solani NRRL 43474 haplotype FSSC 5-b. The strains MB30, MB57, MB61, and MB65 were identified as F. nematophilum, grouping with the reference strain 23TaPT76 (clade B). Only three strains, MB49, MB54 and MB62, grouping with the reference strains F. oxysporum NRRL 52787 and F. curvatum CBS 247.61, were identified as members of Fusarium oxysporum Species Complex (clade C, Figure 2).
Phylogenetic tree generated by maximum likelihood method (bootstrap 1000 replicates) of TEF1 gene sequences of 51 Fusarium strains isolated from olive roots, in Tunisia. Numbers on branches are bootstrap values (>70) based on 1000 replicates. FSSC: Fusarium solani Species Complex; FOSC: Fusarium oxysporum Species Complex; FIESC: Fusarium incarnatum equiseti Species Complex.
The most abundant species, accounting for 17 strains out of 51, grouped together with F. brachygibbosum NRRL25093 reference stain (Clade D).
A great genetic biodiversity was observed in the clade E, including the members of FIESC (Figure 2). In this clade, a single strain, MB52, showed high homology with F. clavum NRRL 45997 reference strain; the strain MB72 grouped with F. gracilipes NRRL 43635 reference strain; seven strain showed 100% homology among them and with F. longifundum NRRL 36372 reference strain; two strains, MB9 and MB10, clustered together with the reference strains F. caatingaense CBS 976.97 and F. incarnatum NRRL 13379, respectively (Figure 2).
Diversity of bacterial communities in olive rhizosphere
3.2
A total of 40 bacterial strains (Supplementary Table 2) were isolated from the rhizosphere samples and molecularly characterized by rep-PCR to reveal clonal relationships between isolates. Results from these assays highlighted 29 different rep-PCR profiles (Figure 3), indicating the genetic diversity of the bacterial isolates even if obtained from the same rhizosphere soil sample and belonging to the same species. A single strain for each rep-PCR profile was identified by 16S rDNA sequencing. For each isolated bacterial strain, the species was assigned on the basis of the comparison of 16S rRNA gene sequence with the sequences of type strains deposited in NCBI database. Three strains were assigned to Novosphingobium genus, showing 99.4% of identity with both N. lindaniclasticum and N. panipatense. A single strain was identified as belonging to Sinorhizobium genus (99.85% of identity with S. kummerowiae and S. meliloti type strains). Two strains, S1C8 and S2C12, were identified as Pseudomonas frederiksbergensis and Pseudomonas iranica, respectively. Ten out of 40 bacterial strains, were identified as Priestia megaterium, followed by 5 strains included in the Bacillus cereus group, 5 strains identified as Bacillus mojavensis/halotolerans and 4 strains identified as Pseudarthobacter siccitolerans (Supplementary Table 2). At lesser extent, two Peribacillus frigoritolerans strains, a single Lysinibacillus zambalensis/xylanilyticus strain, two Bacillus siamensis/velezensis strains, one Paenibacillus peoriae/triticicola strain, one Arthrobacter globiformis strain, one Pseudarthobacter aurescens strain, one Microbacterium phyllosphaerae/foliorum strain, and one Enterobacter spp. strain were also identified (Supplementary Table 2). The results showed that 65% of the isolated strains belonged to the order Bacillales (Figures 4A, B).
Gel-like image of rep-PCR profiles of 30 bacteria isolated from olive rhizosphere, in Tunisia.
(A) Distribution of bacterial Order, identified by 16S rRNA gene sequencing, on olive rhizosphere, in Tunisia; (B) distribution of strains belonging to Bacillales Order. (C) Distribution of bacterial strains according to sampling site, site 1 (Kairouan), site 2 (Hergla, Sousse) and 3 (Sidi Bou Ali, Sousse). (D) Heat map of bacterial strains abundance according to sampling site. The rows represent the isolated bacterial strains, and the columns represent the sampling site. Color scale represents log-transformed abundance values.
The distribution of these bacteria varied significantly among the sampling sites (Figure 4C). At Site 1 (Kairouan) and Site 3 (Sidi Bou Ali, Sousse), Bacillus species was overwhelmingly dominant, representing 41 and 46% of the strains, respectively which conferred them a similar profile (Figure 4D). In contrast, Site 2 (Hergla, Sousse) showed no presence of Bacillus species, and it exhibited a more balanced bacterial community, with Pseudarthrobacter (34%), P. megaterium (17%), and Novosphingobium (17%, Figure 4C).
Assessment of fungal pathogenicity on olive and tomato seedlings
3.3
Seven days after inoculation, necrotic lesions developed on olive leaves (Figure 5; Supplementary Figure 1). The symptoms varied from increasing necrotic areas (MB62, MB68, MB9 and the rest of the tested strains) to the necrosis of the leaf leading to its rolling in case of some strains (F. brachygibbosum MB5 strain). In tomato seedlings, visible differences in vigor and root health were observed. The control plants remained healthy, with fully expanded green leaves and a well-developed root system. In contrast, seedlings inoculated with F. caatingaense MB9 strain showed severe stunting, chlorosis, and root deterioration, indicating strong pathogenicity. Thirty-three percent of the tested strains caused growth reduction and moderate chlorosis, while 60% had a relatively mild impact, with slight yellowing and reduced root mass to no pathogenicity effect on tomato seedlings.
Pathogenicity test on detached olive leaves and stems and tomato seedling inoculated with strains MB9 (Fusarium caatingaense), MB62 (Fusarium oxysporum), MB68 (Fusarium solani) and MB5 (Fusarium brachygibbosum).
The pathogenicity test on olive leaves and stems and on tomato seedlings showed that all the selected fungal strains, with the exception of the F. nematophilum MB57 strain and MB86 strain, belonging to Alternaria genus, were able to cause symptoms on olive organs and/or tomatoes seedling (Table 1). On olive, all tested strains were able to cause symptoms with a rate of infection ranging from 20 to 100%. On tomato seedlings, the strains MB41, MB43 MB57, MB20 and MB24 did not cause symptoms, while the other strains were pathogenic, with infection rates ranging between 25 and 100%. Members of FIESC, namely F. longifundum, F. gracilipes, and F. caatingaense, were pathogenic on olive (DSI values ranging between 3.57 and 28.23%) and tomato seedlings with DSI values ranging from 20 to 80%. In particular, the F. caatingaense MB9 strain induced the highest symptoms both on olive and tomato (Table 1).
The four F. brachygibbosum strains (MB5, MB41, MB43, and MB37) showed a great diversity in causing symptoms, with DSI up to 23%, and between 5.2 and 51.8% on tomato and olive, respectively. In particular, MB5 and MB43 F. brachygibbosum strains were the most pathogenic on olive leaves, with DSI of 51.8 and 45.2%, respectively. All F. solani strains tested were pathogenic on both tomato and olive, with values ranging between 20 and 34% (Table 1). The two strains MB20 and MB59 belonging to Phoma genus, and the two strains MB24 and MB86, belonging to Alternaria genus, showed no or low pathogenicity on both tomato seedlings and olive leaves (Table 1).
Biocontrol potential of olive soil–derived bacteria against fungal pathogens
3.4
Twenty bacterial strains, isolated from olive rizosphere, representative of all identified bacterial species and of the different rep-PCR profiles, were tested against F. brachygibbosum and F. oxysporum species. All strains were used to evaluate their antagonistic activity by the dual culture method and six of them, selected among the most active against Fusarium strains were also used for studying the bacterial–fungal interactions via VOCs (Table 2).
All strains inhibited mycelial growth, with variable values ranging from 19.6 (S1C3 and S3C8) to 41.2% (S3C12) and from 6.1 (S2C20) to 67.5% (B6) when tested against MB1 F. brachygibbosum and MB54 F. oxysporum, respectively (Table 2; Supplementary Figure 2).
In particular, three strains identified as B. mojavensis/halotolerans (S1C1, S1C12 and S3C12), one strain identified as B. siamensis/velezensis (S3C1) and a single P. megaterium strain (S3C2) were the most effective against F. brachygibbosum, inhibiting mycelial growth more than 35%. These strains, together with the strain S1C3 identified as Lysinibacillus zambalensis/xylanilyticus (inhibition value of 31.6%), were also effective in inhibiting mycelial growth of F. oxysporum (Table 2).
With regard to mycelial growth inhibition via VOCs, all six considered strains inhibited F. oxysporum mycelial growth, with values ranging between 15.2 (S3C4, Paenibacillus peoriae/triticicola) and 52.6% (S1C1, B. mojavensis/halotolerans). On the contrary, the two strains S3C1 and S1C17, showing effective antagonistic activity, did not inhibit F. brachygibbosum growth via VOCs (Table 2). Interestingly, although B6 showed the highest antagonistic activity against F. oxysporum strain (inhibition value of 67.5%), it had no effect via VOCs.
Since B. amyloliquefaciens B6 strain exhibited the highest inhibition value against F. oxysporum, it was further tested against other Fusarium strains (MB5, MB50, MB48, MB68) and MB59 strain, belonging to Phoma genus. To evaluate its antagonistic activity, the mycelial growth inhibition was assessed after 3 and 6 days of inoculation (Table 3). Fungal growth was inhibited after three days, ranging from 19.8 (MB48) to 57.5% (MB50). After six days, inhibition values increased further, reaching values between 46.3 and 74.7%, with the strongest effect recorded against F. longifundum MB50 strain (Table 3).
In this experiment, control plates exhibited normal colony development with typical morphology, whereas colonies co-cultured with B6 strain displayed markedly reduced growth (Figure 6). Growth inhibition was particularly evident in MB68, MB54, and MB48 strains, with mycelium appearing less dense and compact. After six days of incubation, differences between control and treated plates became even more evident. In particular, MB5, MB50, and MB54 strains cultured in presence of B6 strain remained more compact and in displayed altered pigmentation (Figure 6).
In vitro interaction between Bacillus strain B6 and MB68, MB5, MB50, MB54, MB48 and MB59 in dual culture on PDA plate at 3rd and 6th day after incubation at 25°C in darkness.
Visualization of Bacillus interactions with Fusarium and Phoma, using light and SEM microscopy
3.5
To investigate the mechanism underlying the antagonistic effect of B6 B. amyloliquefaciens strain, both light microscopy and SEM analyses of the interaction zone between bacterium and F. solani and Phoma strain were performed (Figure 7). After 7 days of co-cultivation, light microscopy revealed clear morphological changes in the hyphae and spores of both fungi compared to control. In F. solani, hyphae appeared irregular and distorted, with evident aggregation of conidia reflected by their reduced density (Figures 7A–a, c), while control samples displayed healthy, elongated hyphae with abundant, well-formed macroconidia (Figure 7A–b). Similarly, Phoma strain showed disrupted hyphal networks and dense clusters of spores (Figures 7B–a, c), contrasting with the more dispersed and intact hyphae in the control (Figure 7B–b). SEM analysis confirmed these observations, showing a dense extracellular matrix surrounding and embedding fungal hyphae and spores in the interaction zones (Figures 7A–d, e; B–d, e). This matrix was consistent with biofilm secretion by B6 bacterial strain, suggesting that it inhibited fungal development by producing a biofilm that physically entrapped and coagulated spores, thereby interfering with sporulation and fungal growth.
Light microscopic (x40) and SEM observations of (A)Fusarium solani. Aa. fungal morphology of Fusarium co-culture with Bacillus. Ab. Light microscopic view of hyphae in control treatment without Bacillus. Ac, Light microscopic view of hyphae in the confrontation zone between Fusarium and Bacillus. Ad and Ae, SEM view of hyphae in the confrontation zone between Phoma and Bacillus. (B)Phoma spp., Ba. fungal morphology of Phoma spp. co-culture with Bacillus. Bb. Light microscopic view of hyphae in control treatment without Bacillus. Bc. Light microscopic view of hyphae in the confrontation zone between Phoma and Bacillus. Bd and Be. SEM view of hyphae in the confrontation zone between Phoma and Bacillus. Images were captured on the 7th day after incubation at 25°C.
Discussion
4
The agricultural soils represent a reservoir of phytopathogenic microorganisms as well as of beneficial microorganisms able to play a key role in the plant-soil system. Indeed, these microorganisms can fix and transform nutrients, vital both to the plant and to other microorganisms, and have direct impact on the plant health. Microbial communities of agricultural soils can also contribute to control pathogenic microorganisms inducing systemic resistance in the plant, and producing antibiotics, siderophore compounds, and cell wall degrading enzymes (Fasusi et al., 2023; Silva et al., 2023).
The study of the microbial community colonizing the agricultural soil is of great importance to predict both their potential impact on plant health and the spread of pathogenic species. Indeed, the presence of pathogenic fungi in the soil represents not only a direct threat to the host crop but also a risk of transfer to neighboring or intercropped species, thereby amplifying disease pressure. Understanding this diversity is therefore essential to predict plant health problems and to develop sustainable management strategies.
Dominance of Fusarium in olive rhizospheres and the influence of intercropping
4.1
Molecular characterization of isolated fungal strains, based on ITS and TEF1 gene sequencing, revealed that the fungal community associated with olive soils and roots was dominated by species belonging to Fusarium and Phoma genera. In particular, Fusarium was by far the most prevalent, confirming its global significance as a soilborne pathogen of numerous crops (Nikitin et al., 2023). Several Fusarium species, especially members of Fusarium oxysporum Species Complex, Fusarium solani Species Complex, and Fusarium Sambucinum Species Complex are responsible for devastating diseases such as vascular wilts and root rots, leading to substantial agricultural losses worldwide (Dean et al., 2012; Zhang et al., 2013).
In Tunisia, increasing reports of olive wilt and decline have been linked to Fusarium infections, although historically the focus has been on V. dahliae (Trabelsi et al., 2017). For decades, many Fusarium species were considered opportunists, mainly attacking olive trees already weakened by abiotic stress such as drought, poor soils, or insect infestation (Palmer and Kommedahl, 1969). However, the recent changes in cropping systems, including the new olive plantations on soils previously hosting Solanaceae plants and the expansion of olive groves intercropped with vegetables, have likely created ecological niches that favor the establishment and the spread of pathogenic Fusarium species. A similar trajectory has been observed for V. dahliae, which shifted from cotton to olive following intercropping or rotation with infected cotton plants (Jiménez-Ruiz et al., 2019; Zhang et al., 2019). These parallels highlight the role of agricultural practices and environmental factors to drive the microbial community dynamics, including the spread of soilborne fungi.
In 2017, Trabelsi et al. reported a high presence of Fusarium species, including F. solani and F. oxysporum, in Tunisian olive groves, where 81 out of 104 Fusarium strains were found to be nonpathogenic. In contrast, the present work provides evidence that links the emergence of pathogenic Fusarium populations to intercropping practices, suggesting that cropping systems may act as a key driver in the transition of these species from minor microbiome components to significant olive pathogens. Pathogenicity was assessed using detached olive leaves and stems, while root infection assays were not conducted due to experimental limitations. As Fusarium species are primarily soil-borne pathogens, symptoms observed on detached tissues may not fully represent the systemic nature of Fusarium as a vascular pathogen. Previous studies have shown that Fusarium culture filtrates and purified mycotoxins can induce necrosis and chlorosis on detached plant tissues independently of fungal colonization, highlighting the ability of Fusarium secondary metabolites to cause tissue damage in vitro (Perincherry et al., 2019; Xu et al., 2021). Therefore, the necrotic symptoms observed in this study may reflect not only the ability of these Fusarium strains to colonize olive plant tissues but also their potential toxigenicity.
Fusarium brachygibbosum represented nearly one-third of all Fusarium isolates in our samples. Although previously reported in olive by (Trabelsi et al., 2017), its relatively high frequency and detection across all sites highlight its emerging importance in Tunisian olive groves and its role in olive wilt decline. Given its wide host range and recent designation as a potential quarantine pest in the European Union (EFSA Panel on Plant et al., 2021), this species deserves particular attention. In addition, members of FSSC were the second most represented group (≈31%) in agreement with earlier findings (Trabelsi et al., 2017).
Meanwhile, members of FIESC accounted for approximately 22% of the isolated Fusarium species. Species of this complex are increasingly associated with tree hosts, including Aleppo Pine in Algeria (Lazreg et al., 2014), Eucommia ulmoides in China (Fang et al., 2020), citrus tree in Lebanon (Abi Saad et al., 2022) and date-palms in the south of Tunisia (Rabaaoui et al., 2021), suggesting that they produce enzymatic repertoires adapted for colonizing lignified tissues. Notably, infections by multiple Fusarium species have been shown to exacerbate disease severity compared to single-species infections (Zhang et al., 2022), which may explain the severity of olive decline observed in some groves. By contrast, F. nematophilum was rarely isolated, and consistent with recent reports, appears to act more as a beneficial endophyte or growth promoter rather than as pathogen (Yan et al., 2024, 2025).
Other fungal genera, although less abundant, may still influence olive health under certain conditions. Alternaria and Phoma include both pathogenic and saprophytic species. Alternaria species are known for producing phytotoxins and have been implicated in leaf spot diseases of olive (Basım et al., 2017; Tziros et al., 2021). However, they are frequently isolated from stressed or Xylella-infected olive trees, suggesting opportunistic colonization (Giampetruzzi et al., 2020; Lamendola et al., 2024). Phoma species have been associated to branch dieback in Tunisian olives (Rhouma et al., 2010), often in conjunction with environmental stressors. Ubiquitous genera such as Aspergillus and Penicillium were also recovered, reflecting the complex microbial community of olive groves (Gharsallah et al., 2023).
Altogether, these findings highlight the dominance of Fusarium in Tunisian olive rhizospheres and underscore the role of intercropping practices in shaping microbial communities. Although plants intercropping can introduce beneficial diversity, it may also favor the spread of soil-borne pathogens, posing risks for both olive and associated crops. The distribution of Fusarium species varied markedly among the surveyed sites, reflecting the influence of intercropping and site-specific management. In Site 1 (Kairouan), where olives were intercropped with potato for seven crop seasons, Fusarium accounted for only 25% of isolates, while Alternaria and other fungi dominated. In contrast, Sites 2 and 3 (Hergla and Sidi Bou Ali), with less diverse and long-term intercropping systems with Solanaceae plants, showed Fusarium as the predominant genus, reaching 54% and 80% of the strains, respectively. More generally, intercropping systems that increase host continuity or include highly susceptible crops may promote the persistence and accumulation of soil-borne fungi, whereas diversified crop rotations can disrupt pathogen life cycles and reduce overall fungal dominance (Huss et al., 2022). This was particularly observed in Site 3, where Fusarium species were the most dominant in the isolated fungal strains, likely due to growing tomato crops between olive rows, which is considered a susceptible host to Fusarium. Similar findings have been reported in other cropping systems where plant diversification was associated with lower incidence of soil-borne pathogens, likely due to changes in root exudate composition, increased microbial antagonism, and improved soil structure. In olives, intercropping with annual crops has been shown to influence both soil nutrient dynamics and microbial communities, with potential consequences for disease pressure (Trabelsi et al., 2017). Additionally, the absence of Verticillium strains among the isolated strains was notable. Similar findings were previously reported where typical Verticillium wilt symptoms were observed but the pathogen was not consistently re-isolated from symptomatic plants (ElDesouki-Arafat et al., 2021; Serrano et al., 2021). These studies emphasize the slow growth of Verticillium on general media, allowing faster-growing fungal taxa to dominate.
Dominance and distribution of bacterial species in olive rhizospheres
4.2
The molecular identification of bacteria isolated from olive rhizosphere samples revealed that the bacterial community was largely composed by Firmicutes, Proteobacteria, and Actinobacteria, consistent with previous studies in olive trees (Dias et al., 2024; Gharsallah et al., 2023; Melloni and Cardoso, 2023). Among the 40 isolated strains, Bacillus was the most frequently identified genus, as observed in previous studies carried out in different arid areas of Tunisia, including Sfax region (Gharsallah et al., 2023). The high presence of Bacillus species confirms the great adaptability of these bacteria to environmental conditions, including the tolerance to desiccation and UV radiation (Gharsallah et al., 2023).
Site-specific variations were observed, since in Site 2, Bacillus was not detected, probably due to specific soil attributes, as previously suggested for other olive orchards (Thenappan et al., 2024). Only one P. megaterium strain was recovered from this site, supporting the influence of local environmental factors on microbial recovery. Interestingly, despite Proteobacteria typically being dominant in olive root microbiomes (Giampetruzzi et al., 2020; Vergine et al., 2020), no Proteobacteria were isolated from Site 3, further illustrating site-specific variability. The presence of Enterobacter in this site may reflect shifts in agronomic practices, such as increased irrigation and potential use of lower-quality water sources (Baudišová et al., 2022).
The distribution patterns of bacterial and fungal communities across the three sites suggest possible ecological interactions that may influence pathogen dynamics in olive orchards. Sites 2 and 3, where Fusarium was overwhelmingly dominant, showed different bacterial species profiles. In Site 2, Bacillus was absent and only one P. megaterium strain was recovered, while Proteobacteria were present. In Site 3, Bacillus was dominant, but no Proteobacteria were isolated. Conversely, in intercropped Site 1, Fusarium abundance was lower and Bacillus co-occurred with other bacterial taxa, suggesting that bacterial diversity may contribute to suppressing fungal dominance. Disease suppressiveness is recognized as an emergent property of complex microbial community interactions (Todorović et al., 2023). Previous studies also indicate that although Bacillus spp. can effectively inhibit Fusarium through the production of antifungal metabolites and competitive interactions under controlled conditions (Yang et al., 2024), such antagonistic effects are highly strain-dependent and strongly influenced by environmental factors. Therefore, simple abundance patterns do not necessarily reflect functional antagonism in situ (Sun et al., 2022).
These observations align with the well-documented antagonistic potential of Firmicutes, particularly Bacillus species, against soilborne pathogens such as Fusarium (Han et al., 2019). Bacillus can inhibit fungal growth through multiple mechanisms, including production of antibiotics, volatile organic compounds, and biofilm formation, creating physical and chemical barriers to pathogen establishment. Therefore, the co-occurrence of diverse bacterial communities in Site 1 may partially explain the reduced dominance of Fusarium, whereas the absence or low abundance of antagonistic bacteria in Sites 2 could have facilitated pathogen proliferation and the observed emergence of pathogenic strains.
These findings suggest a complex interplay between bacterial and fungal communities, where site-specific environmental conditions and cropping practices modulate microbial interactions, ultimately influencing pathogen dynamics and disease risk in olive groves. This highlights the importance of integrating microbial community management, including promotion of beneficial bacteria, as a strategy for mitigating soil-borne pathogen threats.
Native bacterial antagonists as biocontrol agents in olive orchards
4.3
The bacterial strains recovered from olive soils exhibited variable in vitro antagonistic activity against Fusarium and Phoma species, highlighting the potential of soil-derived microbes as biocontrol agents. In dual culture assays, B. amyloliquefaciens B6 strain displayed the strongest inhibition activity, followed by other Bacillus isolates showing moderate inhibition activity. Other genera, including Bacillus cereus group, Paenibacillus, Priestia, Peribacillus, Pseudarthrobacter, Enterobacter, Sinorhizobium, and Pseudomonas, exhibited intermediate to low inhibition. These results indicate that the genus Bacillus is particularly effective in suppressing fungal growth, in line with previous reports of its broad-spectrum antifungal properties (Chen et al., 2009).
Interestingly, VOCs assays revealed that the mechanisms of antagonism varied among strains and species. Although B6 showed the highest direct inhibition in dual culture, it exhibited no effect via VOCs. In contrast, Bacillus strains S1C1 and S1C12 demonstrated strong VOCs-mediated inhibition, suggesting that different strains employ distinct antifungal strategies, including production of VOCs that can inhibit fungal growth (Raaijmakers et al., 2010; Sharifi and Ryu, 2018).
Further temporal analysis of B6 against highly pathogenic Fusarium strains showed that inhibition was evident as early as 3 days after inoculation and increased by 6 days up to 74.7%. The rapid onset of growth suppression highlights the strong and consistent antagonistic potential of B6, capable of acting quickly to limit pathogen development. Colonies exposed to B6 exhibited reduced hyphal density, compact morphology, and altered pigmentation, reflecting physiological stress and growth inhibition. These observations are based on in vitro assays and require further validation in planta to confirm their relevance under whole-plant conditions.
Light microscopy and SEM further elucidated the mechanism of antagonism. At the interaction zones, hyphae of F. solani and Phoma appeared distorted and aggregated, and spores clustered densely compared to controls. SEM images revealed a dense extracellular matrix embedding fungal hyphae and spores, consistent with biofilm production by B6. This suggests a biofilm-mediated mechanism in which the bacterium physically traps and coagulates fungal structures, interfering with sporulation and mycelial expansion. Such biofilm-mediated inhibition has been reported for other Bacillus strains, providing both physical and chemical barriers against phytopathogens (Bais et al., 2004; Ongena and Jacques, 2008).
Overall, these findings demonstrate that Bacillus species activate multiple strategies to interact with other microorganisms including antagonism, direct mycelial inhibition, VOCs production, and biofilm-mediated entrapment. Their activity is likely influenced by both strain-specific traits and environmental conditions, highlighting the importance of selecting robust native isolates for biocontrol applications. The integration of such bacterial biocontrol agents in olive and vegetable cropping systems could reduce Fusarium and Phoma incidence, complementing cultural practices such as intercropping and promoting sustainable disease management in Mediterranean orchards (Raaijmakers et al., 2010).
Conclusion
5
This study provides a comprehensive overview of the microbial communities in Tunisian olive soils and roots, highlighting interactions between fungi and bacteria and the influence of cropping practices. Fusarium was the dominant fungal genus, with its abundance varying across sites. The highest prevalence and severity were observed in Sidi Bou Ali, where the presence of tomato a highly susceptible crop likely contributed to the proliferation of virulent Fusarium strains. Notably, compared to earlier reports, the pathogenicity of Fusarium in these olive orchards has increased over the past seven years, indicating a rise in disease risk. The in vitro assay showed that bacterial communities, particularly Bacillus species, exhibited strong antagonistic potential against Fusarium and Phoma, employing diverse mechanisms including direct mycelial inhibition, VOCs, and biofilm-mediated entrapment which should further be validated through in planta assays. Overall, these findings emphasize the importance of considering site-specific cropping systems, host crop susceptibility, and native bacterial communities in managing soilborne pathogens. Leveraging indigenous biocontrol bacteria alongside informed crop management offers a promising strategy to reduce disease risk and support sustainable olive cultivation in Mediterranean agroecosystems.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abi Saad C. Masiello M. Habib W. Gerges E. Sanzani S. M. Logrieco A. F. . (2022). Diversity of fusarium species isolated from symptomatic plants belonging to a wide range of agri-food and ornamental crops in Lebanon. J. Fungi 8, 897. doi: 10.3390/jof 8090897, PMID: 36135622 PMC 9502176 · doi ↗ · pubmed ↗
- 2Aloo B. N. Makumba B. A. Mbega E. R. (2019). The potential of Bacilli rhizobacteria for sustainable crop production and environmental sustainability. Microbiological Res. 219, 26–39. doi: 10.1016/j.micres.2018.10.011, PMID: 30642464 · doi ↗ · pubmed ↗
- 3Bais H. P. Fall R. Vivanco J. M. (2004). Biocontrol of Bacillus subtilis against Infection of Arabidopsis Roots by Pseudomonas syringae Is Facilitated by Biofilm Formation and Surfactin Production. Plant Physiol. 134, 307–319. doi: 10.1104/pp.103.028712, PMID: 14684838 PMC 316310 · doi ↗ · pubmed ↗
- 4Basım E. Basım H. Abdulai M. Baki D. Öztürk N. (2017). Identification and characterization of Alternaria alternata causing leaf spot of olive tree (Olea europaea) in Turkey. Crop Prot. 92, 79–88. doi: 10.1016/j.cropro.2016.10.013, PMID: 41813336 · doi ↗
- 5BaudišováD. BobkováŠ. JakubůV. JeligováH. Kožíšek F. (2022). Bacteria from the family Enterobacteriaceae in reused water and their antibiotic susceptibility. Vodohospodářské Technicko-Ekonomické Informace 64, 4–9. doi: 10.46555/VTEI.2022.05.002 · doi ↗
- 6Belhedi M. Sghaier-Hammami B. Masiello M. Nafati H. Somma S. Gambacorta L. . (2025). Silicon dioxide (Si O 2) nanoparticles affect the morphology, sporulation, mycotoxin production, and pathogenicity of Fusarium brachygibbosum infecting olive trees. Front. Nanotechnology 7. doi: 10.3389/fnano.2025.1569453, PMID: 41810299 · doi ↗
- 7Benítez-Cabello A. Delgado A. M. Quintas C. (2023). Main challenges expected from the impact of climate change on microbial biodiversity of table olives: current status and trends. Foods 12, 3712. doi: 10.3390/foods 12193712, PMID: 37835365 PMC 10572816 · doi ↗ · pubmed ↗
- 8Bizos G. Papatheodorou E. M. Chatzistathis T. Ntalli N. Aschonitis V. G. Monokrousos N. (2020). The role of microbial inoculants on plant protection, growth stimulation, and crop productivity of the olive tree (Olea europea L.). Plants 9, 743. doi: 10.3390/plants 9060743, PMID: 32545638 PMC 7356289 · doi ↗ · pubmed ↗