CBLN2 promoter enables genetic access to wide-field neurons of the tree shrew superior colliculus
Arda Kipcak, Alev Erisir

TL;DR
Researchers developed a genetic tool using the CBLN2 promoter to target wide-field neurons in tree shrews and mice, revealing distinct input patterns in these neurons.
Contribution
The first viral tool derived from the tree shrew genome for cell-type-specific targeting across species.
Findings
CBLN2 is validated as a molecular marker for tree shrew wide-field neurons.
AAVs based on the CBLN2 promoter enable selective targeting of wide-field neurons across species.
Tree shrew wide-field neurons receive distinct excitatory inputs based on dendritic location.
Abstract
Wide-field (WF) neurons of the tectopulvinar pathway integrate retinal and cortical inputs via large dendritic arbors crucial for rapid visual motion detection. Previous studies identified potential marker genes for mouse WF neurons. Here, we validate CBLN2 as a molecular marker of the tree shrew WF neurons and construct AAVs that exploit CBLN2 promoter to selectively target WF neurons across species. Using intersectional genetics in the tree shrew, we show that WF neuron dendrites receive a distinct pattern of VGluT1+ and VGluT2+ inputs based on their distance from the cell body in the dorsoventral axis of the superior colliculus (SC). This represents the first example of a viral tool derived from the tree shrew genome for cell-type-specific targeting across species. Our results provide a foundation for studying SC circuitry in higher-order mammals and for extending this approach to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
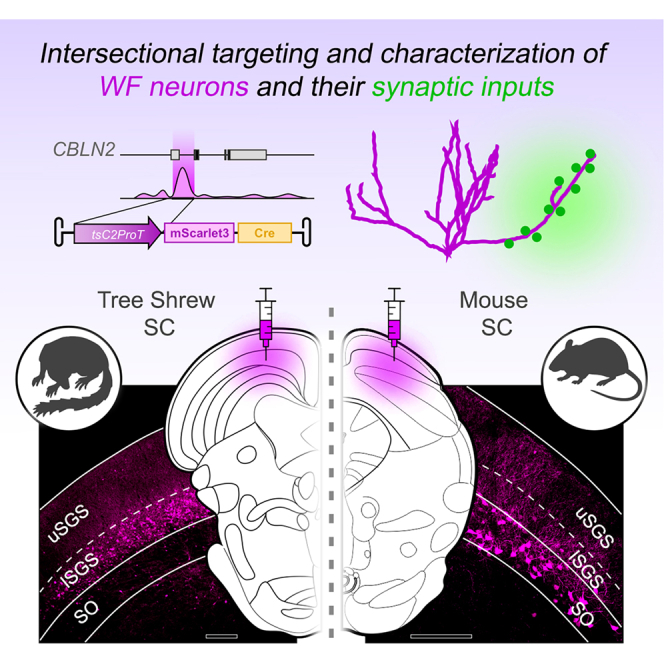 Figure 1
Figure 1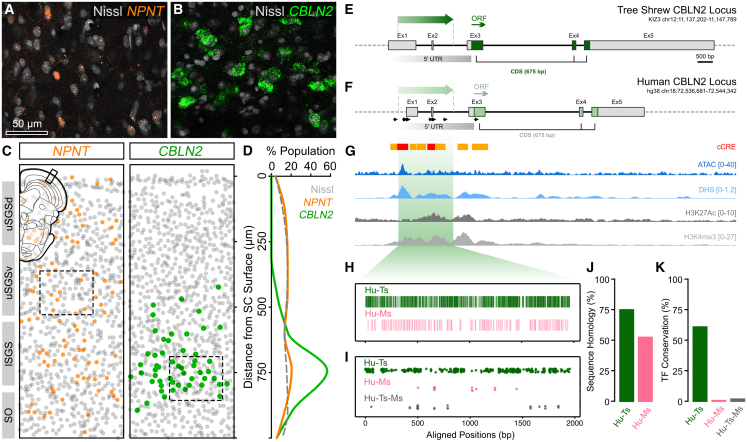 Figure 2
Figure 2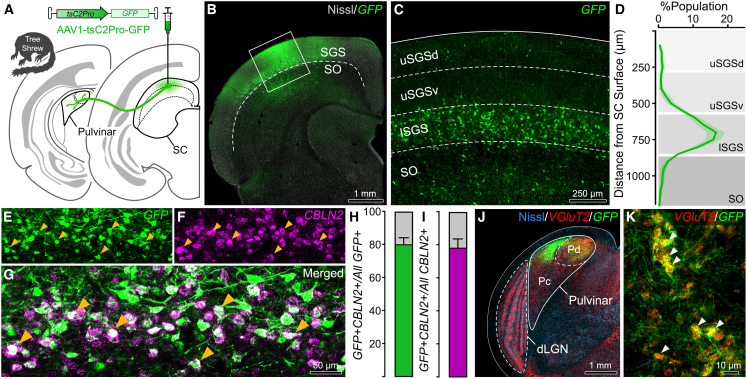 Figure 3
Figure 3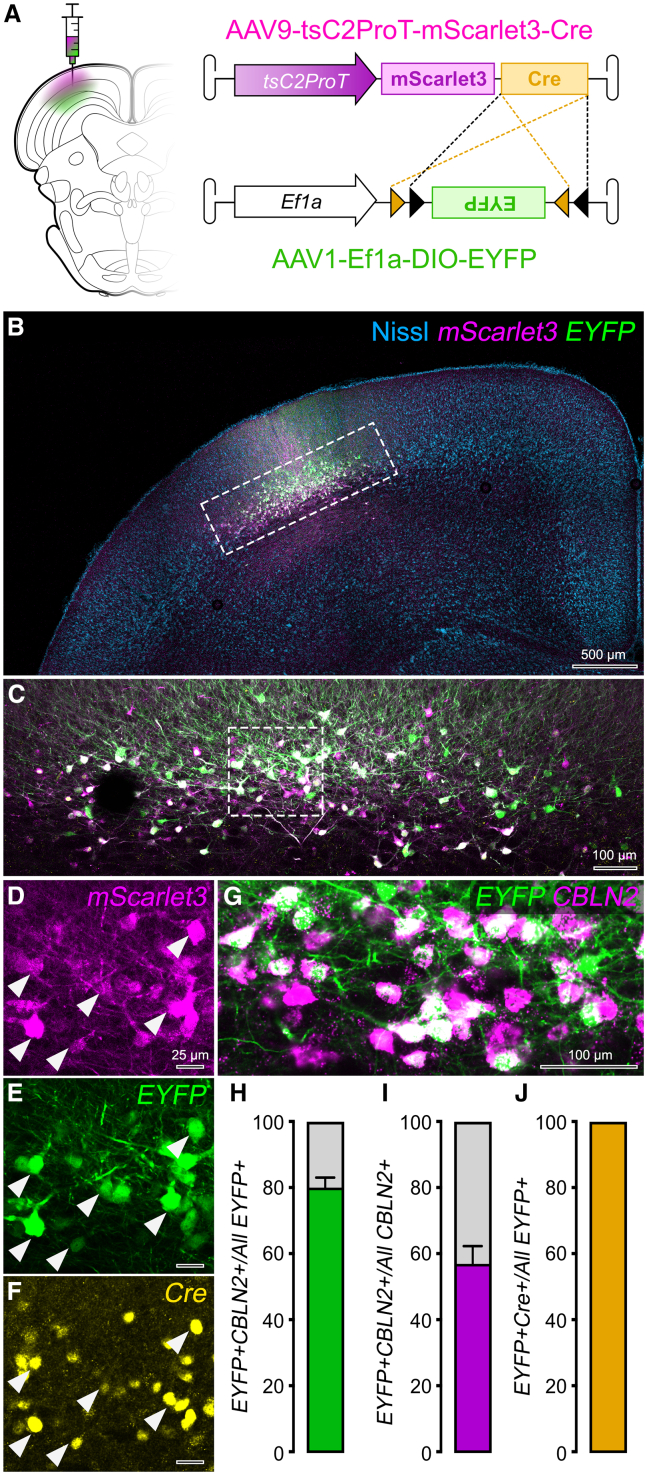 Figure 4
Figure 4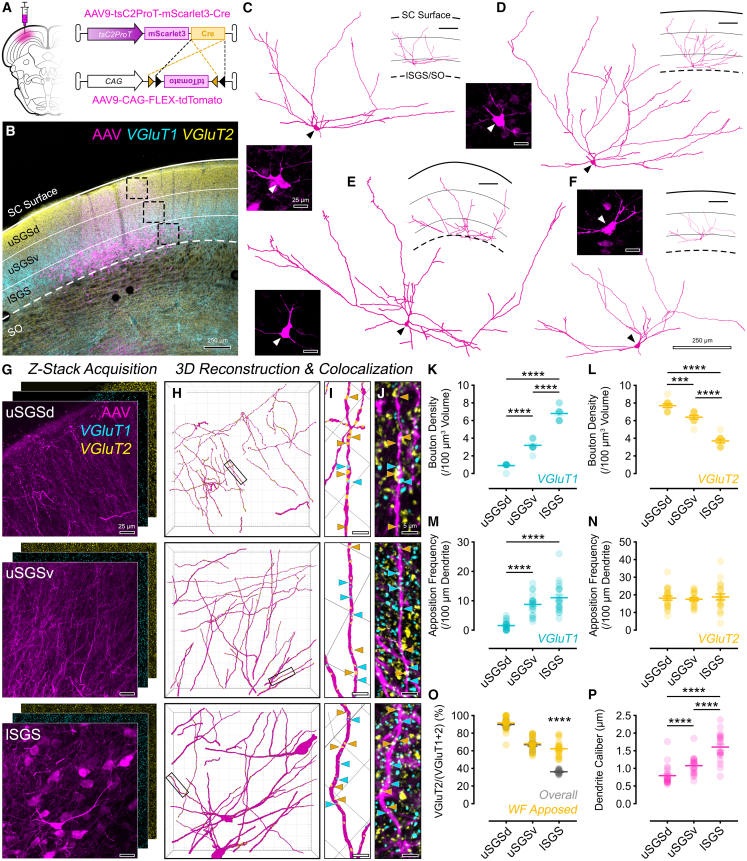 Figure 5
Figure 5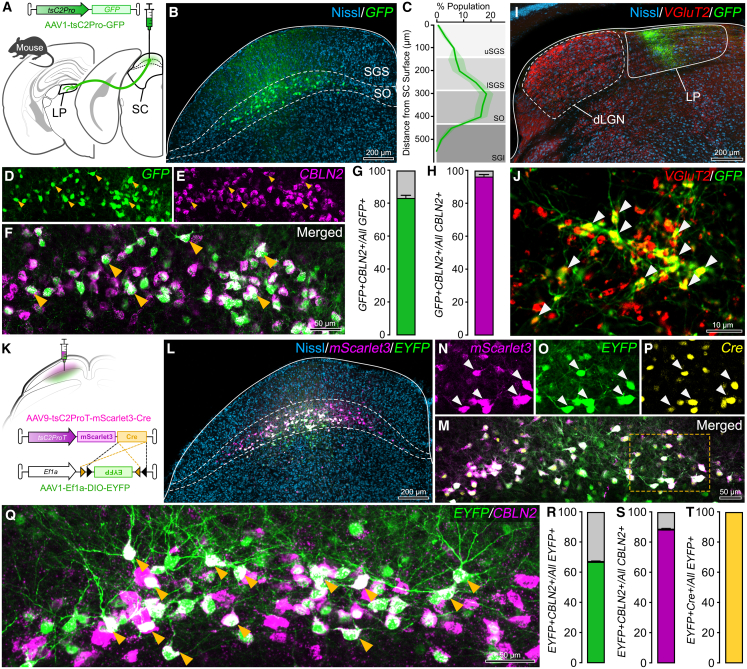 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHearing, Cochlea, Tinnitus, Genetics · Developmental Biology and Gene Regulation · Animal Vocal Communication and Behavior
Introduction
The superior colliculus (SC) is a topographically organized midbrain center critical for visually guided behaviors.1 SC cell types have been classified using morphological, physiological, and, more recently, transcriptomic approaches,2 with most insights derived from mice due to available genetic tools. In particular, Cre-recombinase-expressing transgenic lines have enabled the targeting of four principal superficial SC (sSC) cell types—stellate (RORB),3^,^4 horizontal (GAD2),5^,^6^,^7 narrow-field (GRP),6^,^7^,^8^,^9 and wide-field (NTSR1) neurons5^,^7^,^8^,^9^,^10—but were not systematically generated to target these cell types specifically, often resulting in off-target labeling of additional neuronal populations.7^,^9^,^11^,^12 Moreover, such genetic accessibility in mice is not readily transferable to higher-order mammals. Recent advances in viral vector technologies, including novel capsid variants13^,^14 and gene regulatory elements such as promoters and enhancers,15^,^16^,^17^,^18^,^19^,^20 offer new opportunities for cell-type-specific targeting, yet current tools remain largely limited to forebrain cell types, leaving SC unexplored.
Among the main sSC cell types, wide-field (WF) neuron somata are located at the lower stratum griseum superficiale (lSGS) and stratum opticum (SO), with expansive dendritic arbors and projections to the pulvinar nucleus (lateral posterior [LP] in rodents) of the thalamus.21 Unlike the geniculostriate pathway22^,^23^,^24 that is more specialized in object recognition, tectopulvinar pathway25^,^26^,^27^,^28 is associated with object detection and motion localization in the visual field,21 functions critical for survival. WF neurons, found across a wide range of species, including amphibians,29^,^30 avians,31^,^32 rodents,3^,^5^,^6^,^8^,^10 prosimians,26^,^33^,^34^,^35^,^36 and non-human primates,37^,^38^,^39^,^40^,^41^,^42 are thought to extract motion saliency by integrating retinal and cortical inputs, thereby guiding behaviors such as predator avoidance and prey capture.5^,^6^,^8^,^12^,^43^,^44^,^45^,^46^,^47^,^48 Accessing WF neurons outside mice typically requires retrograde labeling from the pulvinar, a complex hub interconnected with multiple brain regions,49^,^50^,^51^,^52 often necessitating dual adeno-associated virus (AAV) strategies such as combining a Cre-expressing retrograde AAV in the pulvinar with a Cre-dependent AAV in the SC to selectively label WF neurons. This approach further demands high stereotaxic precision to align the topographic maps in both structures to ensure co-expression of the transgene in the same cells via both retrograde and anterograde routes. These challenges underscore the need for simpler, locally restricted, and cross-species compatible tools to access WF neurons.
The tree shrew (Tupaia belangeri) is increasingly recognized as a model organism due to its close evolutionary relationship to primates and its highly developed visual system, which exhibits behavioral and synaptic features more akin to those of primates.53^,^54 Moreover, its relative accessibility compared to primates makes it an ideal bridge species for the development of cross-species neurobiological tools, aligning with the principles of refinement and reduction in animal research.
Building off of recent transcriptomic studies identifying Cerebellin 2 (CBLN2) as a WF neuron marker in mice,7^,^9^,^10^,^55 in the present study, we validated CBLN2 as a conserved molecular signature for WF neurons in the tree shrew. We then identified the tree shrew CBLN2 promoter and used it to engineer cell-type-specific AAVs to target WF neurons across species. For proof of principle, we used these AAVs to map the spatial distribution of excitatory inputs on WF neurons in the tree shrew SC, revealing unique connectivity patterns of putative retinal and cortical inputs on WF dendrites. Together, our findings establish CBLN2 as a conserved marker of WF neurons and demonstrate the utility of CBLN2-promoter-driven AAVs as effective tools for genetic access to this cell type beyond rodents. By enabling precise targeting of WF neurons in the tree shrew, this study lays the groundwork for dissecting the structural and functional architecture of the tectopulvinar circuit.
Results
Identification of tree shrew CBLN2 promoter
Recent molecular and transcriptomic studies of the mouse superior colliculus identified a number of genes including Nephronectin (NPNT) and CBLN2 as potential molecular markers of WF neurons.9^,^10^,^44^,^55^,^56^,^57^,^58^,^59 We asked whether these markers are conserved in tree shrew and could enable genetic access to WF neurons. Using immunohistochemistry and single-molecule RNA fluorescent in situ hybridization (smRNA-FISH), we examined NPNT and CBLN2 expression in tree shrew SC. NPNT+ cells were broadly distributed across SGS, without any clear sublayer selectivity (Figures 1A and 1C), whereas CBLN2+ cells were primarily localized to the lSGS (Figures 1B and 1D), where the tree shrew WF neurons reside35 (Figure S1G). This led us to focus on CBLN2 as a candidate marker.Figure 1CBLN2 locus shares conserved features across species(A and B) Confocal images of Nephronectin (NPNT) IHC (A) and Cerebellin 2 (CBLN2) FISH (B) in tree shrew sSC.(C and D) Spatial and relative distributions of NPNT+ (orange) and CBLN2+ (green) cells across sSC depth, revealing clustering of CBLN2+ cells at the lSGS/SO border. Gray, Nissl-stained cells.(E) Tree shrew CBLN2 locus showing the human-aligned promoter (dark green arrow, tsC2Pro), annotated exons (gray), and protein-coding sequence (green). Genomic coordinates are indicated in the upper-right subtext. ORF, open reading frame; 5′ UTR, 5′ untranslated region.(F) Human CBLN2 locus with the light green region identified as the promoter (huC2Pro) and the mapped transcription start sites (black arrows) from 5′CAGE data (FANTOM5).(G) Human cCRE, ATAC, DNAse hypersensitivity (DHS), H3K27Ac, and H3K4Me3 peaks aligned to the CBLN2 locus. Scale range is shown in brackets at the right.(H) Multiple sequence alignment of tree shrew (Ts) and mouse (Ms) homologs to the human promoter showing pairwise sequence identity between human and tree shrew (Hu-Ts, green) and human and mouse (Hu-Ms, pink). Each bar represents a conserved base.(I) Transcription factor (TF) enrichment plot showing the aligned position of human TF binding sites that are present in tree shrew (green), mouse (pink), or both (gray).(J and K) Quantification of overall DNA sequence homology between the human promoter and aligned tree shrew (green) or mouse (pink) sequences (J) and shared TF-binding sites (K). 100% represents the complete human sequence.See also Data S4.
The tree shrew CBLN2 coding sequence is highly conserved between human (mRNA: 93%, protein: 96%) and mouse (mRNA: 89%, protein: 95%) (see method details for Transcript IDs; see also Data S1). To identify transcriptional cis-regulatory elements, we compared the 5′ UTR, upstream to the coding sequence across tree shrew and human loci, and noted a 3.7 kb conserved region in the vicinity of exon 1/transcription start site. To further delineate this region, we used publicly available human brain chromatin accessibility (ATAC-seq and DNAse-seq) and histone ChIP-seq data (see key resources table for Study IDs). Alignment of the epigenomic data to CBLN2 locus revealed that this region was enriched for histone markers H3K4Me3 and H3K27Ac as well as ATAC and DNase hypersensitivity peaks, which are known to be positioned at active promoter/enhancer regions60^,^61^,^62 (Figures 1F and 1G). Similarly, ENCODE cCRE database indicated a number of candidate enhancer and promoter-like elements concentrated in this region (Figure 1G). Integrating this information with cross-species alignment, we defined the boundaries, designated a 1.8 kb sequence as the putative CBLN2 promoter, and mapped this sequence to the tree shrew genome; this sequence is referred to as tsC2Pro from here on (Figure 1E; see also Data S2). Further analysis of tsC2Pro using JASPAR database (see method details) revealed a high density of transcription-factor-binding motifs shared between the tree shrew and the human (Figures 1I and 1K; see also Data S4), suggesting that transcriptional regulation of CBLN2 is conserved in higher mammals. Accordingly, cross-species comparison of the putative promoters showed 76% homology between human and tree shrew and 53% homology between human and mouse (Figure 1J). In light of these, we posited that tsC2Pro may house the core promoter and potentially other cis-enhancers of the locus and thus can be used to gain genetic access to WF neurons.
CBLN2 promoter directs selective reporter expression in tree shrew WF neurons
To test the hypotheses that (1) tsC2Pro drives CBLN2-specific transgene expression in vivo and (2) the tsC2Pro-driven virus-labeled cells are WF neurons, an AAV vector encoding tsC2Pro upstream of a GFP cassette (AAV1-tsC2Pro-GFP) was stereotaxically injected into adult tree shrew SC (Figure 2A). Three to four weeks following the surgery, coronal brain sections were prepared for confocal imaging. The AAV injection site showed a distinct band of GFP+ cells localized to lSGS at approximately 750 μm depth from the SC surface (Figures 2B, 2C, and S1). Colocalization analysis using smRNA-FISH with probes against tree shrew CBLN2 transcript revealed a high degree of specificity of AAV1-tsC2Pro-GFP: 80% ± 2.16 of the GFP+ cells were CBLN2+. Similarly, 78% ± 2.96 of CBLN2+ cells in analyzed areas expressed GFP, suggesting a high degree of sensitivity of AAV1-tsC2Pro-GFP for CBLN2+ cells (Figure 2E), indicating that tsC2Pro drives CBLN2-specific transgene expression in vivo. Furthermore, the pulvinar nucleus, which receives WF neuron projections, contained GFP-labeled fibers appearing as distinct clustered boutons in both central pulvinar (Pc) and dorsal pulvinar (Pd), as well as diffuse unitary boutons only in Pd (Figures 2F and S4). GFP-labeled terminals in the pulvinar nucleus contained vesicular glutamate transporter type 2 (VGluT2; Figures 2J and 2K), which is characteristic for the WF boutons.28 Thus, the CBLN2-promoter-driven AAV-labeled cells are WF neurons. Together, these results demonstrate that AAV1-tsC2Pro-GFP effectively and selectively targets CBLN2+ WF neurons in tree shrew SC.Figure 2CBLN2-promoter-driven AAV selectively labels WF neurons of the tree shrew SC(A) Experimental scheme for injecting AAV-tsC2Pro-GFP into tree shrew SC. Labeled WF neurons project to the pulvinar.(B) Confocal image of the injection site with AAV-expressed GFP (green) and Nissl (gray) labels. Dashed line = lSGS/SO border.(C) Higher magnification view showing GFP-expressing cells localized to lSGS.(D) Distribution of the GFP+ cell labeling across superficial SC (% of total GFP+ cells).(E–G) Higher magnification of GFP (E) and CBLN2 RNA-FISH (F) labeling in lSGS, showing a high degree of colocalization (G).(H) AAV specificity: GFP+CBLN2+ cells as % of all GFP+ cells (mean ± SEM; 2,466 cells from three tree shrews).(I) AAV sensitivity: GFP+CBLN2+ cells as % of CBLN2+ cells (same dataset as H).(J) Confocal image of a thalamic section showing selective tectopulvinar axonal labeling (green) in pulvinar. Nissl (blue) and VGluT2 (red) IHC mark intra- and internuclear borders of dorsal pulvinar (Pd), central pulvinar (Pc), and dorsolateral geniculate nucleus (dLGN).(K) Higher magnification view of pulvinar showing GFP-labeled tectopulvinar terminals that are colocalized with VGluT2. Note the “grape-wine” appearance of clustered terminals, a characteristic of “specific” tree shrew WF projections (see also Figure S4).
CBLN2-promoter-driven Cre expression allows intersectional targeting of WF neurons
After confirming that AAV1-tsC2Pro-GFP drives reporter expression in CBLN2+ WF cells, we optimized our design to enable intersectional genetics. Conventional AAV serotypes have a 4–5 kb packaging limit,63 restricting payload to a single function, in this case, fluorescent labeling. To expand its utility, we truncated the promoter from 1.8—1.2 kb (tsC2ProT, see method details) and replaced GFP with the brighter, red-shifted reporter mScarlet364 and inserted Cre recombinase. We incorporated these elements into a bicistronic construct expressing mScarlet3 and Cre separated by a P2A element, packaged as AAV9 (AAV9-tsC2ProT-mScarlet3-Cre). To validate Cre function, we injected AAV9-tsC2ProT-mScarlet3-Cre along with a Cre-dependent EYFP expression vector under the control of the ubiquitous promoter Ef1a (AAV1-EF1a-DIO-EYFP, 1:1) into tree shrew SC (Figure 3A). Both reporters were expressed, and EYFP colocalized with mScarlet3 in lSGS, confirming Cre compatibility for intersectional approaches (Figures 3B and 3C).Figure 3tsC2ProT-driven Cre expression allows efficient intersectional labeling of the tree shrew WF neurons(A) Experimental scheme for co-injection of mScarlet3+Cre-expressing truncated CBLN2-promoter-driven AAV and Cre-dependent EYFP-expressing Ef1a-driven AAV into tree shrew SC.(B and C) Low- (B) and high-magnification (C; rectangle in B) confocal images of the injection site showing mScarlet3 and EYFP expression. Note the oblique soma morphology typical of WF neurons.(D–F) Higher magnification view of area marked in (C), showing endogenous expression of mScarlet3 (D), colocalized EYFP (E), and Cre IHC (F).(G) Confocal image showing colocalization of EYFP in CBLN2+ cells.(H) AAV specificity: EYFP+CBLN2+ cells as a percentage of all EYFP+ cells. Total of 1,817 cells from three images of one tree shrew.(I) AAV sensitivity: EYFP+CBLN2+ cells as percentage of all CBLN2+ cells. Same dataset as in (H).(J) Cre-driven recombination specificity: EYFP+Cre+ cells as percentage of all EYFP+ cells. The dataset contained 762 cells from three images.
To rule out non-specific recombination events resulting in spontaneous expression of Cre-dependent transgene in the absence of Cre,65^,^66^,^67 we quantified the percentage of Cre+ cells among all EYFP+ cells using Cre IHC; virtually all EYFP+ cells were also Cre+, confirming that EYFP expression was driven by Cre-specific recombination and not an artifact (Figures 3E and 3F). Similarly, although bi-cistronic systems can show expression bias toward the first gene over the second in the open reading frame,68^,^69 mScarlet3 and Cre were consistently co-expressed in our dataset (Figures 3D and 3F). Finally, specificity and sensitivity remained high despite promoter truncation (80% ± 1.67; 57% ± 3.07) (Figures 3G–3J). Thus, AAV9-tsC2ProT-mScarlet3-Cre enables intersectional strategies for functional and structural analyses of WF neurons.
Tree shrew WF neurons receive retinal and cortical excitatory inputs preferentially at distal or proximal dendrites
Tree shrew SC cells receive glutamatergic input from contralateral retinal ganglion cell axons25^,^70^,^71 and from a variety of cortical areas.72 While the WF neurons receive monosynaptic input from both these sources,9^,^43^,^59^,^73^,^74^,^75 the spatial organization of these inputs as well as any pattern of selectivity to WF dendrites remain unexplored. As a proof-of-concept experiment, we used our intersectional AAV strategy (co-injection of AAV9-tsC2ProT-mScarlet3-Cre and AAV9-CAG-FLEX-tdTomato), which led to ubiquitous expression of red fluorescent proteins selectively in WF neurons. In addition, we used VGluT2 and VGluT1 IHC to visualize putative retinal and cortical axonal terminals in 100–200-μm-thick coronal sections (Figures 4A and 4B). WF neurons, located in lSGS, have large somata (200–300 μm^2^) with lateral and apical dendrites that span over 1 mm lateromedially along the entire depth of the sSC (Figures 4C and 4D). As WF dendrites ramify along the dorsoventral axis, the dendrite caliber is thickest in lSGS and thinnest in the upper SGS dorsal (uSGSd) (Figure 4P). While dendritic arbor reconstructions were constrained by tissue thickness and sectioning plane, numerous dendrite branches originating from somata extended into uSGSd, forming characteristic tufts of this cell type; additional dendritic terminations were observed as tufted endings within the upper SGS ventral (uSGSv).Figure 4. Tree shrew WF neuron dendrites are differentially innervated by putative retinal and cortical inputs based on distance from somata(A) Experimental scheme for co-injection of mScarlet3+Cre-expressing truncated CBLN2-promoter-driven AAV and Cre-dependent tdTomato-expressing CAG-driven AAV into tree shrew SC.(B) Confocal image of the injection site showing mScarlet3/tdTomato expression (magenta), VGluT1 (cyan), and VGluT2 IHC (yellow). Note the reciprocal staining patterns: VGluT1 enriched in lSGS and VGluT2 in uSGSd.(C–F) Tracings of AAV-labeled WF neuron dendritic arbors in 80–100-μm-thick coronal sections. Insets show the somata and dendrites of traced neurons in SC sublayers. Scale bar in (F) applies to (C)–(F) and reveals that WF dendritic arbors may span over a millimeter along the lateromedial plane.(G) Confocal z stacks (0.06 × 0.06 × 0.3 μm voxels) of AAV (magenta), VGluT1, and VGluT2 labeling in uSGSd, uSGSv, and lSGS. The z stacks from each area cover approximately 250 × 250 × 50 μm volume.(H) 3D reconstruction of AAV-labeled WF dendrites across SC sublayers.(I and J) 3D reconstructions (I) based on confocal volumes (J) of representative dendrite segments (rectangles in H) depicting their apposition sites (arrows) with VGluT1+ (cyan) and VGluT2+ (yellow) boutons (not depicted).(K and L) Overall (global) VGluT1 (K) and VGluT2 (L) bouton density in each sublayer confirm the differential innervation of SGS by cortical and retinal axon boutons. N = 10 ROIs/lamina.(M) Frequency of the VGluT1 appositions (putative synapses) along 100 μm length of the WF dendrite reveals that cortical input at distal dendrites is sparse yet robust at proximal segments. N = 20≈ dendrites/lamina.(N) Frequency of the VGluT2 appositions (putative synapses) along 100 μm length of the WF dendrite reveals that retinal axons densely and indiscriminately innervate dendrites in all sublayers. Same dataset as in (M).(O) Relative density index (% ratio of VGluT2 input among all excitatory inputs) on WF dendrites (yellow) and overall (gray), showing that although VGluT1 is the predominant overall input type in lSGS, WF-VGluT2 appositions are significantly more prevalent.(P) Calibers of WF dendrites in SGS sublayers.
Consistent with the previous work,76 the pattern of VGluT2 and VGluT1 staining displayed reciprocal dorsoventral gradients, with VGluT2 enriched in uSGSd and VGluT1 in lSGS (Figures 4B and S1A–S1C). Quantitatively, VGluT1 bouton density increased with depth and differed significantly across sublamina (Figure 4K; uSGSd: 1 ± 0.1, uSGSv: 3 ± 0.2, lSGS: 7 ± 0.25 boutons/100 μm^3^; one-way ANOVA with Tukey’s, p < 0.0001 for all comparisons). Similarly, VGluT2 bouton density was highest in uSGSd and decreased ventrally (Figure 4L; uSGSd: 8 ± 0.21, uSGSv: 6 ± 0.22, lSGS: 6 ± 0.21 boutons/100 μm^3^; one-way ANOVA with Tukey’s, p < 0.0001, for all comparisons).
To map putative synaptic appositions on WF dendrites, high-resolution confocal z stacks were acquired from uSGSd, uSGSv, and lSGS (Figure 4G), capturing distal tufts, middle dendritic arbor, and proximal dendrites, respectively. Individual AAV-labeled dendrite segments in each sublamina were reconstructed and appositions by VGluT1 and VGluT2 (i.e., putative cortical and retinal synapses) on WF dendrites were identified (Figures 4H–4J and S2; see method details for the pipeline). VGluT1 appositions were rare in the uSGSd, mirroring the low overall density of boutons there (Figure 4M). However, disproportionate to the overall bouton density, VGluT1 boutons formed appositions on dendrites with high frequency in uSGSv and lSGS, suggesting VGluT1 inputs are situated to exert a selective and strong influence on WF excitability via synapses on proximal dendrites (uSGSd: 1.57 ± 0.31, uSGSv: 8.76 ± 0.76, lSGS: 11.05 ± 1.09 VGluT1/100 μm dendrite; one-way ANOVA with Tukey’s, p < 0.0001 for uSGSd vs. uSGSv and uSGSd vs. lSGS; p = 0.1013 for uSGSv vs. lSGS). A similar mismatch was also evident for VGluT2 inputs: VGluT2 apposition frequency was uniform across all three sublamina, despite an overall bouton density gradient of VGluT2 (Figure 4N; uSGSd: 18 ± 1.17, uSGSv: 18 ± 0.87, lSGS: 19 ± 1.74 VGluT2/100 μm^3^; one-way ANOVA with Tukey’s, uSGSd vs. uSGSv p = 0.9333; uSGSd vs. lSGS p = 0.9266; uSGSv vs. lSGS p = 0.7585). That is, despite the dorsoventral decrease in VGluT2 boutons across SGS sublamina, VGluT2 appositions are as frequent on proximal dendrites (i.e., in lSGS) as they are on distal tufts (i.e., in uSGSd), suggesting that retinal terminals are situated to influence WF cell excitability via inputs throughout its dendritic arbor.
To assess relative cortical vs. retinal innervation of WF dendrite segments, we used a ratiometric index, plotting VGluT2 as a percentage of all glutamatergic inputs (VGluT1+VGluT2) for both the WF appositions and the overall terminals (Figure 4O). While retinal appositions dominated in the uSGSd, cortical inputs nearly matched the prevalence of retinal inputs in uSGSv and lSGS, suggesting strong influences by both cortex and retina via synapses close to soma, and a selective role for retinal inputs in determining WF neuron’s large receptive fields through synapses on the fanned-out distal dendrites. In addition, only in lSGS, relative innervation of WF neurons by cortical vs. retinal boutons could not be predicted by overall relative presence of these boutons (WF-apposed uSGSd: 91.6% ± 1.65, uSGSv: 68.1% ± 1.77, lSGS: 62.3% ± 1.83; overall uSGSd: 90% ± 1.1, uSGSv: 66.94% ± 1.84, lSGS: 36.1% ± 0.75; two-way ANOVA with Sidak’s, uSGSd p = 0.9070; uSGSv p = 0.9680; lSGS p < 0.0001), providing further evidence for selectivity of retinal axons for WF neurons over other possible targets in lSGS.
Tree shrew CBLN2 promoter also labels WF neurons of the mouse
Given the high sequence similarity of the CBLN2 loci, and prior evidence that CBLN2 marks the WF neurons of the mouse,10^,^55^,^58^,^77 we tested whether tree shrew CBLN2 promoter can also label mouse WF neurons by injecting AAV1-tsC2Pro-GFP into mouse SC (Figure 5A). Three to four weeks following surgery, SC showed GFP+ cells concentrated in the SO/lSGS (Figures 5B and 3C), known location of mouse WF neurons.7^,^10^,^58 Colocalization of GFP+ and CBLN2+ cells using smRNA-FISH probes against mouse CBLN2 transcript (Figure 5D) verified similarly high specificity (83% ± 0.80) and even greater sensitivity (97% ± 0.56), likely due to the lower density of WF cells in mouse SC, affording higher coverage in a smaller volume. We verified the selective axonal labeling of WF projections in the mouse LP nucleus (pulvinar homolog) (Figure 5F). Similarly, as in the tree shrew, these boutons were VGluT2+. Lastly, we also tested the intersectional strategy by injecting the mouse SC with AAV9-tsC2ProT-mScarlet3-Cre and AAV1-Efa1-DIO-EYFP (1:1) and performed the same analysis (Figure 5K). Both AAVs were expressed as visualized by mScarlet3 and EYFP fluorescence (Figure 5L). We also verified Cre co-expression in mScarlet3+EYFP+ cells (Figures 5M–5P and 5T). Colocalization analysis with CBLN2 RNA showed 67% ± 0.55 specificity and 89% ± 0.66 sensitivity. Collectively, these data show that tree shrew CBLN2 promoter can be used to target mouse WF neurons.Figure 5. Tree shrew CBLN2-promoter-driven AAVs robustly target mouse WF neurons(A) Experimental scheme for injection of AAV-tsC2Pro-GFP into mouse SC. Labeled WF neurons project to the lateral posterior nucleus (LP, pulvinar homolog) of the thalamus.(B) Confocal image of the injection site with AAV-expressed GFP labeling (green) and Nissl staining (blue). Dashed lines indicate sSC borders.(C) Distribution of the GFP+ cells across sSC (% of all GFP+ cells).(D–F) Higher magnification views of GFP, CBLN2 RNA-FISH, and merged colocalization.(G and H) AAV specificity and sensitivity: GFP+CBLN2+ cells as a % of all GFP+ (G) or CBLN2+ (H) cells. N = 6 images from three mice, yielding 4,252 cells.(I) Confocal image of a thalamus section, showing selective WF axon labeling (green) in LP. Nissl and VGluT2 IHC are used to highlight intra- and internuclear borders.(J) Higher magnification of the LP showing colocalization (arrows) of GFP-labeled WF terminals with VGluT2.(K) Experimental scheme for the co-injection of mScarlet3+Cre-expressing truncated CBLN2-promoter-driven AAV and Cre-dependent EYFP-expressing Ef1a-driven AAV into mouse SC.(L) Confocal image of the injection site showing mScarlet3 and EYFP expression.(M) High-magnification image of the injection site showing mScarlet3+EYFP+ cells.(N–P) Higher magnification view of the area marked in (M), showing endogenous expression of mScarlet3 (N), colocalized EYFP (O), and Cre IHC (P).(Q) Confocal image showing colocalized EYFP expression with CBLN2+ cells.(R) AAV specificity. N = 9 images yielding 4,196 cells from three mice.(S) AAV sensitivity. N = 6 images yielding 4,252 cells from three mice.(T) Cre-driven recombination specificity. N = 4 images yielding 490 cells from two mice.
Discussion
In this study, we validate CBLN2 as a WF neuron marker, identify its promoter, and characterize viral tools using this promoter to access WF neurons of the tree shrew and mouse. AAVs described here enable local genetic access to WF neurons, offering new means to assay the structure and function of these cells. Additionally, these AAVs represent the first viral tools derived from the tree shrew genome that affords cell-type-specific genetic access to a conserved SC neuron population across species. With these advancements, several outstanding questions remain to be answered.
Role of CBLN2 in WF neurons
CBLN2 is a secretory signaling protein expressed widely across the brain.78^,^79^,^80^,^81^,^82 Acting as a transsynaptic adapter, CBLN2 connects the presynaptic neurexins (NRXN1/3) with postsynaptic delta-type glutamate receptors (GRID1/2).79^,^82 Shibata et al. (2021) reported a developmental role for CBLN2, where elevated CBLN2 expression promoted spine formation in mouse PFC.83 Mechanistically, Nrxn1^SS4+^-CBLN2 complexes enhance NMDAR-mediated transmission in mouse PFC, and conditional CBLN2 deletion markedly reduces NMDAR-EPSCs,79 suggesting that CBLN2 is critical for efficient synaptic signaling. Given the role of tectopulvinar circuitry in rapid detection of predators and preys, CBLN2 may not only serve as a selective marker but also contribute functionally to WF neuron’s excitability as a presynaptic and/or postsynaptic agent.
Influence of retinal and cortical inputs on WF dendritic integration
Our input-mapping results indicate that although VGluT2 terminals are less abundant than VGluT1 in the lSGS, WF dendrites there receive numerous VGluT2 contacts, often surpassing VGluT1. The strong VGluT1 presence in lSGS and uSGSv aligns with earlier reports that WF neurons are more likely than other SC cell types (narrow-field, stellate, and horizontal) to receive V1 input.74 Our results further reveal that retinal contacts are positioned to provide excitation along the entire dorsoventral span of WF dendrites. Consistent with this, Kühn et al. (2025) confirmed the strong excitatory role of retinal inputs, while suggesting that the V1 input may play a minimal role in the dendritic integration.43 However, it is important to note that Kühn et al. study was conducted in V1-excised animals. Notably, no study has exhaustively mapped the cortical inputs beyond V1 that innervate WF neurons in the tree shrew. Thus, the tools described should enable future transsynaptic mapping of WF afferents. Also to note, VGluT2 served as a proxy for all retinal inputs, without distinguishing RGC subtypes. Previous work in mice shows SC-projecting RGC subtypes vary in their laminar targeting—some innervate the entire superficial SC (ZO+SGS+SO), others project selectively to SGS, lower SGS/SO, or to a combination of these layers.9^,^43^,^59^,^84 In our datasets (not quantified here), we observed diverse VGluT2 bouton morphologies across sSC, likely reflecting distinct RGC subtypes.24 Further studies are needed to define the molecular, structural, and functional diversity of retinotectal RGCs and their targeting preferences toward WF vs. non-WF neurons.
Distinct tectopulvinar circuitries and WF neuron subtypes
Tree shrew WF circuitry exhibits features distinct from rodents: first, as shown in our study (Figure S3), tree shrew tectopulvinar axons are exclusively ipsilateral. Second, unlike rodents where bilateral and ipsilateral projections map to Pcm (tree shrew Pd) and Pl (tree shrew Pc), respectively, tree shrew tectopulvinar axons differ by innervation mode rather than subdivision. Both Pd and Pc receive topographically organized or “specific” WF inputs, but Pd subdivision additionally receives non-topographic or “diffuse” inputs.28^,^33^,^35 Luppino et al.33 first described this pattern, noting that Pd was always diffusely labeled regardless of SC injection site, whereas clustered bouton patches reflected retinotopy, leading to the current terminology.33 This pattern suggests the presence of two WF subtypes, each with distinct termination types.85 Using CBLN2-promoter-driven AAVs (Figure S4), we replicated these findings: central SC injection produced both “diffuse” Pd labeling (Figure S4B) and “specific” patches in Pc and Pd (Figures S4C–S4H). Furthermore, labeled terminals in each subdivision exhibited synapses and distinct circuitry properties, confirming that diffuse appearance of labeling in Pd reflects genuine connectivity rather than axons of passage (Figures S4I and S4J). In contrast, mouse SC injections yielded only topographic boutons in LP without clear diffuse/specific patterns, consistent with prior tracer studies.10^,^57
Because our AAV strategy relies on CBLN2 enrichment in WF neurons, an important question is whether CBLN2 marks all WF neurons or only a subset. If the latter, our vectors would preferentially label CBLN2+ subtypes. While this cannot be fully excluded, a recent study reported that a vast majority of retrogradely identified WF neurons in both mouse and tree shrew were CBLN2+,55 supporting its suitability as a molecular target for accessing WF neurons. Taken together, CBLN2-targeted AAVs access WF neurons contributing to both projection types.
Two WF subtypes have been described in the ground squirrel: WF-I projecting bilaterally to caudal pulvinar (tree shrew Pd) and WF-II projecting ipsilaterally to rostral pulvinar (tree shrew Pc).86 WF-I neurons occupy the lSGS with dendrites reaching the collicular surface, whereas WF-II neurons lie near the SGS/SO border with tufts in uSGSv,86 a morphology that may constrain dendritic spread and receptive field size. The presence of bilateral and ipsilateral WF inputs to Pcm (tree shrew Pd) and Pl (tree shrew Pc) is also evident in the mouse57 but not in primates or tree shrews. In primates, early lesion studies suggested potential contralateral tectal projections to inferior pulvinar,87^,^88 but subsequent work reported strictly ipsilateral projections.38^,^39^,^40^,^42^,^72^,^89 Consistent with primates, tree shrew SC exhibited only ipsilateral tectopulvinar projections in both anterograde and retrograde tracing experiments (Figure S3). Thus, tree shrew may represent an early stage in proto-primate evolution where cross-hemispheric tectopulvinar projections have largely disappeared.
Limitations of the study
We acknowledge a number of technical limitations of this study. First, the epigenomic datasets used were derived from human. In the absence of tree shrew data—and given its closer phylogenetic relationship to primates than rodents90^,^91^,^92^,^93—we used human epigenomic datasets assuming shared chromatin accessibility profiles. We also compared the mouse CBLN2 locus to assess cross-species conservation—a hallmark of promoters/enhancers and transcription-factor-binding sites.94^,^95^,^96 The human brain epigenetic data used to help identify cis-regulatory elements are derived from cortex, rather than SC; ideally, tree shrew SC multi-omics datasets would be preferable but were unavailable. We therefore relied on human cortical data, assuming promoter use would be global and vary less by brain region compared to enhancers.83 Notably, a previous study identified a developmentally restricted PFC-specific CBLN2 enhancer (E2), which was active only during mid-fetal cortical development,83 whereas the putative promoter identified in the same study—which partially overlaps with the promoter identified in our study—exhibits constitutive DNA accessibility in the mouse, and H3K27Ac marks in the human, indicating consistent CBLN2 promoter activity across developmental time points. Our study focused on adult SC and did not assess viral expression in other regions or at earlier time points.
Second, the endogenous GFP signal from AAV1-tsC2Pro-GFP was weak and required IHC amplification, likely reflecting the strength of the promoter or a serotype-dependent effect in the tree shrew. In contrast, AAV9-tsC2ProT-mScarlet3-Cre exhibited robust endogenous mScarlet3 expression without amplification.
Third, WF neuron tracings were limited by tissue thickness; we optimized tissue thickness to include a large axial volume without compromising high-magnification confocal imaging using a 40× objective with a travel range of ∼180 μm (z stack volume). Initial light-sheet imaging of cleared intact SC at 20× lacked sufficient resolution to capture thin dendrites in upper SGS, prompting a switch to confocal imaging. While fluorescent labeling enabled multiplexed AAV and input marker visualization for a proof-of-concept approach, input mapping was limited to structural imaging; future work should include larger volume datasets and physiological validation.
Finally, we acknowledge the possibility of off-target AAV expression in non-WF populations. Previous work with enhancer-driven AAVs indicates a large range of achieved maximal specificity and sensitivity, particularly with Cre-expressing vectors.20 Thus, we recognize that the identity of labeled neurons is putative, underscoring the need for future physiological characterization of visual responses and associated behaviors. Additionally, low-level CBLN2 expression in non-WF cells cannot be excluded; indeed, we detected, although less robust, CBLN2 transcripts outside lSGS (data not shown), suggesting that some labeled cells represent on-target, non-WF CBLN2+ populations. Nevertheless, using lower viral titers (10^9−10^ vg/mL) minimized off-target labeling. The AAV delivery routes tested were limited to stereotaxic injections directly to brain parenchyma; future work should explore systemic routes (e.g., retroorbital or tail vein) and broader range of titers.
Resource availability
Lead contact
Requests for further information reported in this paper should be directed to the lead contact, Alev Erisir ([email protected]).
Materials availability
Newly generated AAV vectors in this paper are made publicly available as of the publication date as DNA intermediates through the Addgene IDs listed in the key resources table. Further inquiries regarding AAVs generated in this paper will be fulfilled upon reasonable request by the lead contact.
Data and code availability
- •All genetic data are available in this paper’s supplemental information. Existing, publicly available genomic and epigenomic data used in this paper are accessible at respective repositories reported in the key resources table.
- •All original code is deposited to UVA DataVerse and is publicly available at https://doi.org/10.18130/V3/NEGJWP as of the publication date.
- •Any additional information regarding the data and analysis reported in this paper is available from the lead contact upon request.
Acknowledgments
We thank Francesca Sciaccotta, Christopher Turner, and Rebecca Roberts (UVA, Psychology) for assistance with tree shrew surgeries and handling and Yuanming Liu (UVA, Biology) and Chen Chen (UVA, Psychology) for help with smRNA-FISH and mouse surgeries. We also thank Jessica Connely, PhD (UVA, Psychology) for smRNA-FISH equipment access and Jianhua Cang, PhD (UVA, Biology and Psychology) and John Campbell, PhD (UVA, Biology) for valuable feedback. This work was supported by the 10.13039/100000002NIH-10.13039/100000065NINDS U01NS122040 grant.
Author contributions
A.K. and A.E. designed the study. A.K. designed the viral vectors and conducted the experiments. A.K. and A.E. analyzed the data, interpreted the results, and designed the figures. The manuscript was drafted by A.K. and edited by A.E. A.E. secured the funding. Both authors read and approved the final version of the manuscript.
Declaration of interests
The authors declare no competing interests.
Declaration of generative AI and AI-assisted technologies in the writing process
During the preparation of this work, the authors used GPT4 in order to compress and improve the organization of the R code used for transcription factor analysis. After using this tool, the authors reviewed and edited the content as needed and take full responsibility for the content of the publication.
STAR★Methods
Key resources table
REAGENT or RESOURCESOURCEIDENTIFIERAntibodiesRabbit Anti-NPNT Antibody (1:500)NovusCat# NBP1-83990Rabbit Anti-CBLN2 Antibody (1:500)InvitrogenCat# PA5-101514;RRID: AB_2850949Rabbit Anti-Cre (1:1000)Sigma-AldrichCat# 69050Rabbit Anti-GFP (1:1000)Sigma-AldrichCat# AB3080Rabbit Anti-GABA (1:250)Sigma-AldrichCat# A2052Mouse Anti-PV (1:200)Sigma-AldrichCat# P3088Guinea Pig Anti-VGluT2 (1:1000)Sigma-AldrichCat# AB2251-IMouse Anti-VGluT2 (1:1000)ChemiconCat# MAB5504Mouse Anti-VGluT1 (1:1000)ChemiconCat# MAB5502Rabbit Anti-VGluT1 (1:1000)Synaptic SystemsCat# 135 303Rabbit Anti-PSD95 (1:100)Sigma-AldrichCat# S1-6900Rabbit Anti-SP (1:1000)ChemiconCat# AB1566Goat Anti-ChAT (1:100)ChemiconCat# AB144PRabbit Anti-CALB2 (1:250)SwantCat# CR7697Donkey Anti-Goat Cy3Jackson ImmunoResearchCat# 705-165-003RRID: AB_2340411Goat Anti-Guinea Pig AF488 (1:250)AbcamCat# ab150185Donkey Anti-Guinea Pig Cy5 (1:250)Jackson ImmunoResearchCat# 706-175- 14;RRID: AB_2340462Goat Anti-Guinea Pig AF594 (1:250)InvitrogenCat# A-11076Donkey Anti-Rabbit AF555 (1:250)InvitrogenCat# A-31572Donkey Anti-Rabbit AF488 (1:250)InvitrogenCat# A-21206;RRID: AB_2535792Donkey Anti-Rabbit Cy5 (1:250)Jackson ImmunoResearchCat# 711-175-152RRID: AB_2340607Donkey Anti-Mouse TRITC (1:250)Jackson ImmunoResearchCat# 715-025-151RRID: AB_2340767Donkey Anti-Mouse AF647 (1:250)InvitrogenCat# A-31571Donkey Anti-Mouse AF488 (1:250)InvitrogenCat# A-21202RRID: AB_141607Goat Anti-Rabbit Biotinylated (1:200)Vector LaboratoriesCat# BA-1000Bacterial and virus strainsAAV1-tsC2Pro-GFPThis paperAddgene, Cat# 248521AAV9-tsC2ProT-mScarlet3-CreThis paperAddgene, Cat# 248522AAV1-Ef1a-DIO-EYFPKarl Deisseroth LabAddgene, Cat# 27056AAV9-CAG-FLEX-tdTomatoEdward Boyden LabAddgene, Cat# 28306AAVrg-CAG-GFPEdward Boyden LabAddgene, Cat# 37825Chemicals, peptides, and recombinant proteinsBSAThermo ScientificCat# AAJ6410036Triton X-100MP BiomedicalsCat# 194854Sodium AzideMP BiomedicalsCat# 102891GlycerolMallinckrodtN/AEthylene GlycolSupelcoCat# EX0565NeuroTrace Nissl StainInvitrogenCat# N21479TSA Vivid Fluorophore 650 (1:1000)ACDCat# 323273Vectashield Plus DAPIVector LaboratoriesCat# H-2000Critical commercial assaysRNAscope™ Multiplex v2ACDCat# 323270VECTASTAIN® ABC-HRP Kit, PeroxidaseVector LaboratoriesCat# PK-4000Deposited dataTF binding motif dataThis paperData S4AAV vector maps and sequencesThis paperData S3Cross-species CBLN2 promoter sequence annotationsThis paperData S2Cross-species CBLN2 CDS annotationsThis paperData S1FANTOM5 TSS dataLizio et al.97https://genome.ucsc.edu/cgi-bin/hgTrackUi?db=hg38&g=fantom5BOCA ATAC-Seq dataFullard et al.98https://labs.icahn.mssm.edu/roussos-lab/boca/; GEO: [GSE96949](GSE96949)ENCODE DNase-Seq dataJohn StamatoyannopoulosENCSR880CUBENCODE H3K4Me3 ChIP-Seq dataBradley BernsteinENCSR257VEOENCODE H3K27Ac ChIP-Seq dataBradley BernsteinENCSR004HIEExperimental models: Organisms/strainsT. belangeri Tree ShrewThis paperUVA Tree shrew ColonyM. musculus MouseThis paperC57BL/6J; RRID: IMSR_JAX:000664OligonucleotidesTree Shrew CBLN2 FISH ProbeACDCat# 428551Mouse CBLN2 FISH ProbeACDCat# 1265181-C1Recombinant DNAAAV-tsC2Pro-GFPThis paperAddgene, Cat# 248521AAV-tsC2ProT-mScarlet3-CreThis paperAddgene, Cat# 248522Software and algorithmsImaris (10.2.0)Oxford Instrumentshttps://imaris.oxinst.com/ImageJ/FIJI (1.54p)Schindelin et al.99https://imagej.net/software/fiji/SNT (4.3.0)Arshadi et al.100https://imagej.net/plugins/snt/GraphPad Prism (10.5.0)Dotmaticshttp://www.graphpad.comSnapGene (8.1.1)Dotmaticshttp://www.snapgene.comCIIDERGearing et al.101https://ciiider.erc.moash.eduNCBI BLASTNIH/NLMhttps://blast.ncbi.nlm.nih.gov/Blast.cgiTree shrew DatabaseYe et al.91http://www.treeshrewdb.org/UCSC Genome BrowserPerez et al.102https://genome.ucsc.edu/ENCODE PortalLuo et al.103https://www.encodeproject.org/R (4.4.1)R Core Team104https://cran.rstudio.com/RStudio (2025.05.0 + 496)Posit team105https://posit.co/download/rstudio-desktop/msa (R library)Bodenhofer et al.106https://doi.org/10.18129/B9.bioc.msaBiostrings (R library)Pagès et al.107https://doi.org/10.18129/B9.bioc.Biostringsdplyr (R library)Wickham et al.108https://dplyr.tidyverse.org/tibble (R library)Müller and Wickham109https://tibble.tidyverse.org/readr (R library)Wickham et al.110https://readr.tidyverse.org/stringr (R library)Wickham111https://stringr.tidyverse.orgggplot2 (R library)Wickham112https://ggplot2.tidyverse.org/tidyr (R library)Wickham113https://tidyr.tidyverse.org/Grid (R library)Murrell114https://stat.ethz.ch/R-manual/R-devel/library/grid/html/grid-package.html
Experimental model and study participant details
Data for this study was collected from the brains of 10 adult tree shrews (Tupaia belangeri) and 6 adult mice. Further Information related to the animals used in this study are provided in the table below. All procedures were approved by the University of Virginia Institutional Animal Care and Use Committee (IACUC) and were conducted in accordance with the National Institutes of Health guidelines.SpeciesAnimal IDSexAge (months)T. belangeri1M82T. belangeri2M35T. belangeri3M9T. belangeri4F15T. belangeri5M19T. belangeri6M32T. belangeri7F33T. belangeri8M10T. belangeri9F12T. belangeri10F17M. musculus1M7M. musculus2M7M. musculus3M9M. musculus4M3M. musculus5M3M. musculus6M3
Method details
Stereotaxic surgeries
The animals were anesthetized with 4% isoflurane (SomnoFlo; Kent Scientific, Torrington, CT) and placed on a stereotaxic apparatus. An incision was made along the scalp, and a craniotomy was performed above the SC. The stereotaxic coordinates used for tree shrew SC injections were AP: −6.0-7.5, ML: 2.0–2.5, DV: 3.5–4.0 mm; and for mice SC injections were AP: −3.5–4, ML: 0.6, DV: 1.2 mm. A NanoFil 10 μL syringe equipped with 34g beveled needle (WPI, Sarasota, FL) was used to deliver ≈500nL/site of AAV in saline (see key resources table) at an infusion rate of 1 nL/s using Legato 130 Nanosystem (KD Scientific). The final titers used to dose SC with AAVs were as follows: AAV1-tsC2Pro-GFP (5 × 10^11^ GC/mL), AAV9-tsC2ProT-mScarlet3-Cre (5 × 10^11^ GC/mL), AAV1-Ef1A-DIO-EYFP (2.5 × 10^12^ GC/mL), AAV9-CAG-FLEX-tdTomato (1.2 × 10^12^ GC/mL), AAVrg-CAG-GFP (2 × 10^13^ GC/mL). In the experiments where two AAVs were co-injected, the aforementioned final titers were reached through a 1:1 mix of the two AAVs before injection. The needle was left at target for an additional 5 min at the end of the injection cycle to minimize backflow before it was retracted. The craniotomy was sealed with bone wax, the scalp was sutured, and the animals were placed on a heating pad until mobile. After the surgery, animals were monitored for three days to ensure proper wound healing and were observed for any behaviors indicative of pain or discomfort.
AAV design and preparation
Custom AAVs described in the paper were designed using SnapGene software (GSL Biotech, Boston, USA). Borders of the full-length tree shrew promoter (tsC2Pro) were determined based on the sequence conservation to and predicted regulatory elements of the aligned human promoter. Briefly, human CBLN2 locus ±50 kb flanking the first and the last exon (UCSC Genome102: hg38, chr18:72,536,681-72,544,342) was aligned to tree shrew genome (Tree shrew Database, KIZ391) using NCBI Basic Local Alignment Search Tool (BLASTn), which yielded A 3.7kb conserved block in the human locus spanning from 0.3 kb upstream to the first exon to 0.9 kb downstream to the third exon. 5′ end of this sequence was marked as the start of the promoter. Within this block, there was an Open Reading Frame (chr18:72,542,352-72,542,639) in the (+)strand, 0.2 kb upstream to the coding sequence (CDS) of the canonical transcript (NM182511.4), which may interfere with the expression of the reporter to be cloned. For this reason, that region of the block was omitted and the 3′ end of the rest of the blasted region was marked as the end of the promoter. In addition, the incorporated region also neatly aligned with the predicted cis-regulatory elements with promoter- and enhancer-like signatures (Figures 1F and 1G). Thus, the coordinates of the human CBLN2 promoter (huC2Pro) were designated as chr18:72,542,815-72,544,612. Finally, this region was aligned to the tree shrew (tsC2Pro: KIZ3 chr12:11145679-1114749) and mouse (mm39 chr18:86728752-86729905) loci reported in the Figure 1. Truncation of the promoter (tsC2ProT) was performed at the 3′ end of the tree shrew-aligned predicted promoter-like element (Encode cCRE ID: EH38E1925959, chr18:72543433-72543672). Both the promoter (Data S2) and the complete AAV vector sequences (Data S3) are provided in the supplemental information. Custom Gateway cloning and AAV packaging was outsourced commercially. For AAV-tsC2Pro-GFP, tsC2Pro was inserted upstream of an eGFP-WPRE cassette sandwiched between two AAV2 ITR sequences and packaged as AAV1. For AAV-tsC2ProT-mScarlet3-Cre, tsC2ProT was inserted upstream of a mScarlet3-P2A-Cre-WPRE cassette sandwiched between two AAV2 ITR sequences and packaged as AAV9. Both AAVs were produced with HEK293T triple-transfection, ultrapurified with cesium chloride (CsCl) gradient ultracentrifugation and titrated with qPCR targeting AAV2 ITRs to stock concentration of >10^13^ GC/mL. The necessary quality control documentation is available from the lead contact. All sequences and plasmid maps are made available at Addgene IDs listed in the key resources table.
Epigenomic data visualization
All human brain epigenomic data was imported from the respective web servers listed in the key resources table directly to UCSC Genome Browser (hg38) as custom tracks. Tracks height and Y axis upper limit were set to maxima within the browser for the reported region in Figure 1 to enable better visualization. The final browser panel with all tracks including CBLN2 RefSeq were downloaded as vector graphics (pdf).
Tissue preparation
Three to six weeks following injections, the animals were deeply anesthetized with an overdose of Euthasol (excess of 0.25 mL/kg i.p.) and transcardially perfused with Tyrode’s solution (137 mM NaCl, 2 mm KCl, 0.9 mM CaCl2, 1.2 mM MgCl2, 11.9 mM NaHCO3, 0.4 mM NaH2PO4, 5.5 mM glucose, 281 mOsm, pH 7.4) for 1–2 min, followed by 300mL of a fixative solution containing 4% paraformaldehyde (PFA) in 0.1 M phosphate buffer (pH 7.4). Subsequently, brains were blocked and sectioned coronally or sagittaly at 50–200 μm on a Leica VT 1000 S vibratome (Leica Biosystems). Sections used for electron microscopy were incubated in 1% sodium borohydride and rinsed in 0.01M PBS. Sections were then stored in 0.05% sodium azide (NaN3) in 0.01 M PBS at 4°C prior to immunohistochemistry and/or in cryoprotectant buffer (20% Glycerol, 30% Ethylene Glycol, 50% 0.1M PB) at −20°C for smRNA-FISH.
Immunohistochemistry (IHC)
Antibodies and their dilutions used in the current study are detailed in the key resources table. For confocal microscopy, sections were first blocked in 0.01M in PBS containing 1% bovine serum albumin (BSA), 0.5% Triton X- and 0.05% NaN3 for 1 h. Following rinsing with PBS, the sections were placed in the primary antibodies diluted in the same buffer for 18-36h at room temperature on an orbital shaker with slight agitation. To terminate the incubation, sections were rinsed in 0.01 M PBS and incubated in fluorophore-conjugated secondary antibodies for 4-12h. sSC Lamina borders were determined by the expression of different markers including VGluT1, VGluT2, Nissl, DAPI, Myelin and anatomical landmarks (Figures S1A–S1F) as previously noted.1^,^26^,^71^,^76^,^115 For electron microscopy, sections were blocked with 0.01M in PBS containing 1% BSA, 0.1% Triton X- and 0.05% NaN3 for 30 min and then rinsed in PBS. Sections were then incubated in primary antibody diluted in 1% BSA and 0.05% NaN3 for 12-18h at room temperature on an orbital shaker with slight agitation. To terminate the incubation, sections were rinsed in 0.01 M PBS and incubated in biotin-conjugated secondary antibody for 4-6h.
RNA fluorescent in situ hybridization (smRNA-FISH)
RNAscope Multiplex v2 (ACD, Cat #323270) was used to detect CBLN2 transcripts. Protocol consisted of pre-treatment, FISH and IHC (where required) steps. For pre-treatment, 50 μm-thick fresh/cryopreserved tree shrew or mouse SC sections were treated with 0.3% Hydrogen Peroxide for 10 min at room temperature in a 24 well-plate. Sections were then rinsed in 0.01 M PBS and mounted on a SuperFrost+ slide (Fisher Scientific; Cat #12-550-15) and air-dried. The slide was then baked in an oven at 40°C for 30 min, followed by incubation in 4% PFA at 4°C for 15 min and dehydration in ethanol gradient (50%, 70%, 100%) at room temperature (5 min each). The slide was then air-dried and boiled first in distilled water for 10 s and then in target retrieval buffer for 5 min. After target retrieval, the slide was rinsed in distilled water and dehydrated in 100% ethanol for 3 min at room temperature, air-dried and kept in a closed container until the next steps. For FISH, sections were outlined in a hydrophobic barrier, briefly rehydrated with distilled water at room temperature and then digested with Protease IV (tree shrew) or Protease III (Mouse) at 40°C for 40 min in HybEZ II Hybridization System (ACD, Cat# 321711). Following protease treatment, sections were hybridized with tree shrew (ACD, Cat#1265181-C1) or mouse (ACD, Cat#428551) CBLN2 probes at 40°C for 2h. After probe hybridization, the sections were then treated with amplification reagents (AMP 1–3) at 40°C for 30-30-15 min respectively. This was followed by treatment with HRP (15 min) and TSA Vivid Dye (30 min, 1:1000) at 40°C for fluorescence visualization. Finally, the slide was washed in 0.01 M PBS and either air-dried for coverslipping with Aqua-Poly/Mount Mounting medium (Polysciences, Cat #18606) or treated with blocking buffer for regular on-slide IHC as described in the respective methods subsection.
Confocal microscopy
Tissue sections were mounted on Superfrost Plus Slides (Fisher Scientific, Cat#22-037-246), air-dried, and cover slipped with Aqua-Poly/Mount Mounting medium. Images were subsequently collected using Leica Stellaris 5 laser scanning confocal microscope. Low magnification representative images were collected using HC PL FLUOTAR 5×/0.15 NA and HC PL FLUOTAR 10×/0.40 NA. High magnification images used for RNA-FISH were collected using HC PL APO 20×/0.75 NA, HC PL APO 40×/1.3 NA or HC PL APO 63×/1.4 NA. 3D datasets used for WF dendritic arbor tracing in Figures 4C–4F were acquired using HC PL APO 40×/1.3 at 4K or 6K resolution yielding a final voxel size of 0.1 × 0.1 × 0.3–1 μm. 3D datasets used for synaptic apposition analysis in Figures 4G–4J were acquired using HC PL APO 63×/1.4 NA at 4K resolution yielding a 246 × 246 μm a field of view of with final voxel size of 0.06 × 0.06 × 0.3 μm.
WF neuron tracing
Dendritic arbor tracings in Figures 4C–4F was performed in SNT Plugin for ImageJ.100 Briefly, tiled Z-Stacks encompassing the SGS depth at the injection site (FOV Height: ≈0. 8 mm from the SC surface, FOV width: ≈2–4 mm on the lateromedial axis) were imported to SNT. First, each visualized primary dendrite was traced (as individual “paths”) from soma until the branching point (if present), and merged in a single root, representing the soma. Then, from every branching point (often bifurcation) all secondary, tertiary and higher-order paths were traced as daughters to the path above (the parent dendrite), preserving the path order info. For visualization purposes, paths were filled using the “fill” command individually for each path and the fill was exported as a binarized mask. This masked stack was Z-projected (Maximum) and the original soma shape outlined using free-hand selection was added to the image. The final binary trace was pseudo colored to pink and the background was subtracted in Adobe Photoshop 2025. Insets showing location of the cells were generated by superimposing the trace to the original image with the SC borders.
Electron microscopy
Vibratome-cut 60 μm pulvinar sections from a tree shrew injected with AAV9-tsC2ProT-mScarlet3-Cre and AAV1-Ef1a-DIO-EYFP in the SC (Figure 3), were incubated in anti-GFP as described above, followed by biotinylated secondary antibody treatment to visualize AAV-expressed EYFP labeling in the pulvinar. Electron microscopy compatible staining was achieved via an incubation in Avidin-Biotin-Complex (VECTASTAIN ABC-HRP Kit, Peroxidase) solution for 2h, followed by incubation in a solution of 0.02% hydrogen peroxide (H2O2) and 0.05% diaminobenzidine (DAB) for 3–8 min. For EM resin embedding, the sections were postfixed in 1% osmium tetroxide (OsO_4_) prepared in 0.1M phosphate buffer for 1 h. They were subsequently stained with 4% uranyl acetate in 70% ethanol for 1 h, followed by dehydration in 90% and 100% ethanol series and acetones, and infiltration in a 1:1 acetone-resin mixture overnight. The next day, sections were transferred to 100% resin (EMBED 812; EMS, Hatfield, PA) and incubated overnight. Sections were then flat-embedded between two Aclar sheets (EMS, Hatfield, PA) and polymerized in a 60°C oven overnight. Brightfield images from flat-embedded sections were acquired to identify the pulvinar subdivision designated for electron microscopy. Relevant areas incorporating both Pd and Pc were excised and placed into BEEM capsules (EMS, Hatfield, PA), filled with resin, and cured at 60°C for 48 h until fully polymerized. The region of interest including anatomical landmarks such as medio-lateral boundaries and capillaries was reconstructed using a camera lucida. Embedded tissue was then trimmed into a 1 × 2 mm trapezoid encompassing the sample regions of the Pd+Pc. Ultrathin sections (60 nm) were cut using an ultramicrotome (Ultracut UCT7; Leica, Buffalo Grove, IL) and collected on 400-mesh copper grids (Ted Pella, Redding, CA). Pulvinar subdivisions in the ultrathin sections were identified based on trapezoid orientation within the capsule-embedded tissue and the presence of defining landmarks. Ultrathin sections on copper grids were examined on a JEOL1010 electron microscope equipped with a 16-megapixel CCD camera (SIA). Immuno-labeled terminal images were acquired using 8,000–15,000× magnification, yielding a pixel size of 0.8–1.7 nm at the lipid bilayer resolution.
Quantification and statistical analysis
All data are presented as mean ± SEM unless otherwise stated. Group comparisons were performed using one-way or two-way ANOVA with Tukey’s post-hoc tests unless otherwise indicated. Sample sizes are reported in figure legends. All analyses were performed using GraphPad Prism 10 and R.
Sequence alignment and transcription factor binding site analysis
Promoter sequences described above (in the section titled “AAV design and preparation”) from human, tree shrew, and mouse were imported in FASTA format and aligned using the MUSCLE algorithm via the msa package in R. The alignment was processed using Biostrings to calculate per-species coverage and pairwise percent identity, based on ungapped aligned positions. Sequence names were standardized, and alignment statistics were computed using dplyr, tibble, and base R functions. A curated list of human TFBSs was obtained using CIIDER tool.101 Briefly, HuC2Pro (FASTA) was scanned using default settings for JASPAR CORE vertebrates (2020) matrix and the resulting promoter panel (CSV) was imported to R using readr. Each TFBS was mapped to the aligned human sequence by identifying corresponding ungapped positions, preserving strand orientation and using Biostrings for reverse complementation as needed. Aligned sequences from tree shrew and mouse were extracted at the mapped TFBS positions and compared to the human-aligned TFBS using a character-wise similarity score. A 70% sequence similarity threshold was used to classify TFBSs as human-specific or conserved with one or both non-human species. Classification and comparisons were performed using dplyr and stringr. TFBS conservation was visualized using ggplot2. A scatterplot was used to display the distribution of TFBSs along the aligned promoter, colored by conservation class. A complementary rug plot was used to show basewise identity between human and the tree shrew or mouse across the full alignment. Visual layouts were refined using patchwork, tidyr, and grid. Access to all data and code used here is publicly available and provided in the data and code availability section.
Calculation of AAV specificity and sensitivity
AAV specificity and sensitivity were calculated with the formulas below. Plotting and hypothesis testing were performed in GraphPad Prism (Boston, Massachusetts USA).
Dendrite reconstruction and synaptic apposition analysis
Pipeline for the dendrite reconstructions and synaptic apposition analysis was carried out in ImageJ and Imaris with four main steps. First step (Pre-processing): the multi-channel confocal z-stacks with tdTomato, VGluT1 and VGluT2 were imported into the Imaris for pre-processing (of the VGluT1 and VGluT2 signal) with the following filters: Layer Normalization, Background Subtraction (1 μm), Gaussian (0.061 μm). Second step (Segmentation): Pre-processed VGluT1 and VGluT2 channels were segmented using supervised machine learning (ML) function of the Imaris. Briefly, for each channel, “surface” objects were generated with the default settings. Using the interactive ML interface, foreground (the punctated VGluT signal) and background (the regions where VGluT signal is not expected, soma core) was marked iteratively to train the algorithm until prediction matched the ground truth (raw image). Then, the generated surface objects were used as 3D masks to binarize the signal by “Mask All” function, setting the intensity (8-bit, range: 0–255) inside of the objects to 255, and the outside to 0. The binarized VGluT channels were saved as multi-channel TIFs. Images were then imported into ImageJ, and the following operations were performed: Binarize (default), Fill Holes, Watershed, Erode, Median (Size = 1). Third Step (Tracing): SNT plugin was used to trace dendrites on the tdTomato channel as described above. Each dendrite and their daughter paths (if present) were manually traced and individually filled. The fill was then exported as a binarized mask (“Binary Fill”). Fourth Step (Colocalization): The channels for the binary VGluT1/VGluT2 signal and the binary dendrites were colocalized using the “AND” operator of the Image Calculator, yielding two separate image stacks where All VGluT1 or VGluT2 pixels contained within the dendrite i.e., synaptic appositions as described earlier.116^,^117
Additionally, we benchmarked the validity of this approach by dual-labeling VGlut2 (pre-synaptic) with PSD95 (post-synaptic) and carrying out the same pipeline. Among 438 VGluT2 appositions detected, 83.6% colocalized with PSD95 (Figure S3F). Analysis of the apposition volumes revealed that those that are PSD95- (16.4%) were the smallest volumes within the distribution, approximately 10× smaller than the PSD95- (PSD95+: 0.8647μm^3^ ± 0.057, PSD95-: 0.0881μm^3^ ± 0.016). Thus, we devised a volume cutoff, below which the detected putative appositions would be considered false positive. We calculated %Prediction Accuracy and %True Positive Retention for a range of cutoff values between 0 and 0.5 μm^3^ with the following formulas:
By calculating the point at which regression of these two functions cross, we determined the volume cutoff at which our method obtains both the highest %Prediction Accuracy and the highest %True Positive Retention (0.0218 μm^3^, 90.86% N = 21 %Prediction Accuracy: R^2^ = 0.6755, p < 0.0001; %True Positive Retention: R^2^ = 0.9509, p < 0.0001). Thus, the appositions with volumes smaller than 0.0218 μm^3^ in the subsequent analysis reported in Figure 4 were considered false positive and excluded from the analysis. It is also worthy to note the previous work by others reporting PSD95- VGluT2 synapses or at least the failure of its detection thereof in the mouse cortex.118
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Basso M.A.Bickford M.E.Cang J.Unraveling circuits of visual perception and cognition through the superior colliculus Neuron 109202191893710.1016/j.neuron.2021.01.01333548173 PMC 7979487 · doi ↗ · pubmed ↗
- 2Cang J.Chen C.Li C.Liu Y.Genetically defined neuron types underlying visuomotor transformation in the superior colliculus Nat. Rev. Neurosci.25202472673910.1038/s 41583-024-00856-439333418 · doi ↗ · pubmed ↗
- 3Gale S.D.Murphy G.J.Distinct cell types in the superficial superior colliculus project to the dorsal lateral geniculate and lateral posterior thalamic nuclei J. Neurophysiol.12020181286129210.1152/jn.00248.201829897837 PMC 6171055 · doi ↗ · pubmed ↗
- 4Byun H.Lee H.-L.Liu H.Forrest D.Rudenko A.Kim I.-J.Rorβ regulates selective axon-target innervation in the mammalian midbrain Development 1462019 dev 17192610.1242/dev.171926 PMC 667936131332038 · doi ↗ · pubmed ↗
- 5Gale S.D.Murphy G.J.Distinct Representation and Distribution of Visual Information by Specific Cell Types in Mouse Superficial Superior Colliculus J. Neurosci.342014134581347110.1523/JNEUROSCI.2768-14.201425274823 PMC 4180477 · doi ↗ · pubmed ↗
- 6Gale S.D.Murphy G.J.Active Dendritic Properties and Local Inhibitory Input Enable Selectivity for Object Motion in Mouse Superior Colliculus Neurons J. Neurosci.3620169111912310.1523/JNEUROSCI.0645-16.201627581453 PMC 6601912 · doi ↗ · pubmed ↗
- 7Chen C.Liu Y.Cang J.Accessing genetically defined cell types in the superior colliculus with transgenic mouse linesi Science 28202511219410.1016/j.isci.2025.112194 PMC 1198248340212591 · doi ↗ · pubmed ↗
- 8Hoy J.L.Bishop H.I.Niell C.M.Defined Cell Types in Superior Colliculus Make Distinct Contributions to Prey Capture Behavior in the Mouse Curr. Biol.29201941304138.e 510.1016/j.cub.2019.10.01731761701 PMC 6925587 · doi ↗ · pubmed ↗