Development of Stereotypic Behaviors and Personality Traits of Captive Male Forest Musk Deer and Relationships with Musk Secretion
Xiaoping Lu, Yan Sheng, Hong Ye, Zisong Yang, Xiuxiang Meng

TL;DR
This study explores how personality traits and repetitive behaviors in captive male forest musk deer relate to musk quality, offering insights for better breeding and conservation strategies.
Contribution
The study identifies a novel link between exploratory personality traits and musk quality in forest musk deer.
Findings
Exploratory and active personalities correlate with higher muscone content in musk.
Stereotypic behaviors do not significantly affect musk production or quality.
Personality traits can inform breeding and conservation strategies for captive musk deer.
Abstract
Musk deer are shy and easily stressed, and thus struggle to adapt to artificial environments compared to other captive animals. This stress can lead to them performing unnatural, repetitive behaviors. Understanding their individual personalities could help us improve their welfare and manage them better. In this study, we watched the behavior of 25 male forest musk deer living on a farm in China. We recorded how often they performed repetitive actions and assessed their personalities, categorizing them as more exploratory and active or more cautious. We also measured the amount and quality of the musk secreted by the male musk deer, a valuable substance used in medicine. We found that musk deer with more exploratory and active personalities produced higher quality musk. The repetitive behaviors were generally mild and did not reduce their musk production. This suggests that an animal’s…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
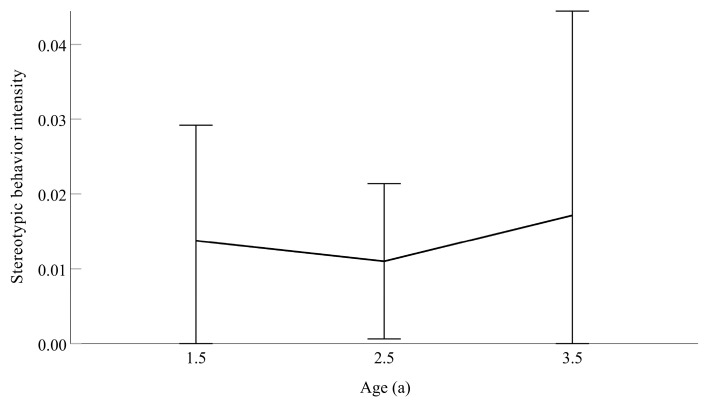 Figure 1
Figure 1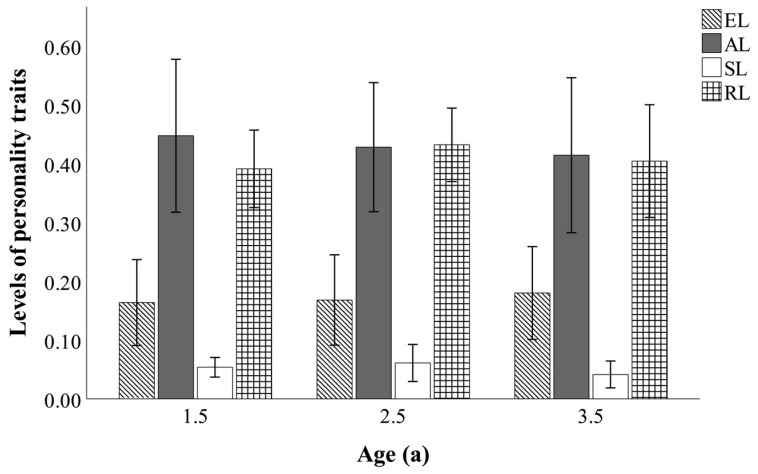 Figure 2
Figure 2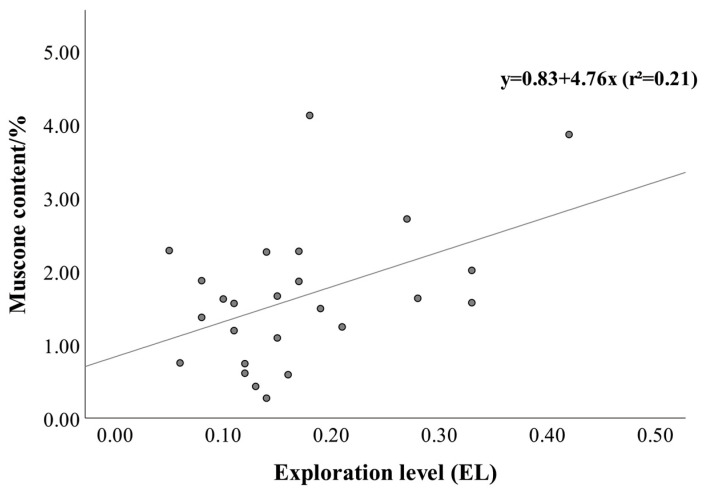 Figure 3
Figure 3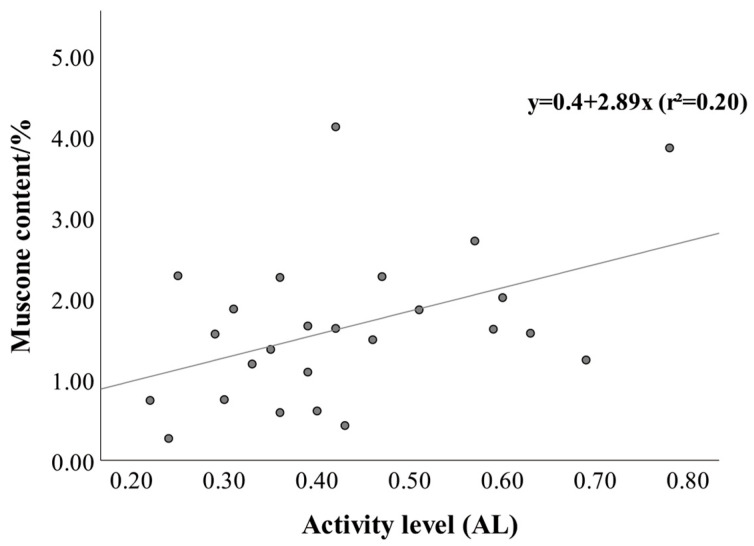 Figure 4
Figure 4- —National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsWildlife Ecology and Conservation · Primate Behavior and Ecology · Animal Behavior and Welfare Studies
1. Introduction
Musk deer (Moschus spp.), a group of small solitary forest ruminants (SSFRs), are endangered resource species endemic to Asia [1,2]. The musk secreted by male individuals is widely utilized in traditional Asian medicine and the perfume industry, rendering it a threatened resource species [3]. Due to historical overexploitation and habitat changes, wild populations of musk deer have become critically endangered [2]. Musk deer species are listed on the Red List of the International Union for Conservation of Nature [4] and in the Appendix of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) [1].
In situ strategies (e.g., habitat protection) have been insufficient due to ongoing poaching and habitat loss [5]. Ex situ management, particularly captive breeding, has been crucial for musk deer conservation and the sustainable use of musk [6,7]. China’s captive breeding programs have currently produced over 60,000 individuals and enabled reintroduction efforts [5,8]. The forest musk deer is the most numerous Moschus species in captivity, followed by the alpine musk deer (Moschus chrysogaster) [7]. The healthy development and sustainable production of captive musk deer populations have received widespread attention [8,9]. The musk production capacity can be reflected by indicators such as the secretion amount of musk and muscone content [10].
To enhance the scope of ex situ conservation and scale of musk production, forest musk deer have been introduced to Fujian Province in southeastern China, in addition to breeding efforts being undertaken in their natural habitats, such as in Sichuan and Shaanxi Provinces [11]. This initiative has contributed to the establishment of the easternmost ex situ conservation population of forest musk deer [7,12]. In male forest musk deer, musk secretion typically begins around 1.5 years of age and gradually increases during subsequent years, reaching relatively stable levels by 3.5 years [10]. Therefore, the 1.5–3.5-year age range represents a critical period from initial musk production to early maturity, making it suitable for examining the relationships between behaviors, personalities, and musk secretion [13]. Understanding stereotypic behaviors and personality traits is important because they affect animal welfare, reproductive success, and musk production in captivity [14]. Improving welfare and productivity supports sustainable ex situ populations, which in turn can provide individuals for reintroduction into the wild. However, there is a lack of behavioral and physiological research concerning captive forest musk deer in the eastern regions of China, particularly regarding the relationship between their stereotypic behaviors, personality traits, and musk secretion [12,13].
The environment in which captive animals are kept is inherently artificial, characterized by limited space and scheduled feeding, which can easily lead to the development of stereotypic behaviors [15,16,17]. Studies have shown that the manifestation of these behaviors can adversely impact both animal welfare and the productivity of captive animals (such as with regard to meat, eggs, milk production, etc.) [18]. However, some studies have shown that this correlation does not always exist [19]. For instance, cows (Bos taurus) exhibiting stereotypic behaviors may yield more milk than normal cows, as their activated energy metabolism mobilizes body nutrient reserves to support lactation, while their more stable lying and drinking behaviors and stronger heat tolerance collectively enhance lactation performance [20]. A study on captive alpine musk deer has also found no significant correlation between stereotypic behavior and musk secretion [14].
Furthermore, personality traits can represent stable behavioral trends in animals across time and situations among individuals, and are closely related to resource use, reproduction, and predation [21,22,23]. For example, male zebrafish (Danio rerio) with heightened levels of boldness and aggression tend to possess a competitive advantage during the breeding process [24]. Some studies have analyzed the personality of musk deer but have not associated it with musk secretion [25,26]. Another study focused on identifying boldness–shyness traits in forest musk deer using behavioral tests, but did not examine how these traits relate to musk production [25]. Moreover, one study explored the link between boldness and hormone levels in alpine musk deer, but did not extend their analysis to musk secretion [26]. Thus, although these studies advanced our understanding of personality in musk deer, they left a gap in evaluating whether personality traits influence musk yield or quality—an important consideration for integrating behavioral indicators into breeding and management strategies.
This study, based on the behavioral sampling of individuals under human care with the lowest latitude, investigated the stereotypic behaviors and personality traits of forest musk deer and their relationships with musk secretion. This study aims to (1) quantify the types and intensities of stereotypic behaviors in the captive male forest musk deer population; (2) assess the levels of personality traits, including exploration level, activity level, self-directed level, and redundancy level; (3) analyze the differences in the above indicators among different age groups; and (4) explore the relationships between the stereotypic behaviors and personality traits with the musk secretion of forest musk deer. And we hypothesized the following: (1) Stereotypic behavior intensity and personality traits differ among age groups. (2) There are differences in the musk secretion (amount of musk, muscone content) among individuals with different personality traits, e.g., individuals with a higher exploration level produce more musk and higher muscone content. (3) Stereotypic behaviors are associated with reduced musk secretion. The research findings can provide important references for the ex situ conservation, breeding, and musk production of this threatened species.
2. Materials and Methods
2.1. Research Area
This study was conducted from 1 September to 20 December 2023 at a musk deer farm located in Zherong County, Fujian Province, China. The musk deer farm is situated in the southeastern inland mountainous area of Fujian Province (27°05′ N27°19′ N, 119°43′ E120°04′ E), with an average altitude of about 850 m, an annual average temperature of 16 °C, and an annual average rainfall of 2100 mm. This population represents the easternmost and lowest-latitude captive forest musk deer population in China, established through introduction from traditional breeding areas in Sichuan and Shaanxi Provinces.
2.2. Research Population
This study involved 25 male captive forest musk deer, with an age range of 1.5–3.5 years. This age range was based on the actual breeding situation of the newly established musk deer farm, where the introduced individuals had a relatively young age, and no adult individuals over 3.5 years old were available at the time of the study. Notably, all individuals had reached the musk-secreting stage, which ensured the uniformity of the core research index (musk secretion) and the validity of the subsequent behavioral and physiological correlation analysis. The individuals were further divided into three age groups with clear sample sizes: 1.5-year-olds (N = 8), 2.5-year-olds (N = 10), and 3.5-year-olds (N = 7).
Each musk deer was kept alone in a 10 m^2^ enclosure and a 30 m^2^ activity area. Their health condition was good, and ear tags were used for individual recognition. The feed composition comprised concentrate (40% corn, 12% wheat bran, 25% soybeans, 15% soybean meal, and 8% whey powder) and fresh green leaves (such as leaves of Morus alba and Taraxacum mongolicum). The feed was provided twice daily (at 05:00 and 17:00), and the enclosures were cleaned during feeding, with no disturbances to the musk deer at other times.
2.3. Behavior Sampling
Following reference [1], focal sampling and all-occurrences recording were employed to record individual behavior categories and durations. Observations were carried out outside the enclosures using binoculars (10 × 50) to minimize disturbance, as recommended for behavioral studies on captive ungulates [9]. Each focal sampling session lasted 10 min per individual, conducted during the peak activity periods of forest musk deer (05:00–08:00 and 17:00–20:00). A total of 25 individuals were observed, and each individual was sampled repeatedly across different days throughout the study period to obtain representative behavioral data. If a focal animal moved out of sight during a session, sampling was paused and later rescheduled to ensure complete records [27].
Stereotypic behaviors observed in this population were defined based on the established ethogram for captive musk deer [9] (Table 1). The stereotypic behaviors can be categorized into oral stereotypic behaviors (e.g., eating non-food items and stereotyped licking and scraping) and locomotor stereotypic behaviors (e.g., galloping, to-fro-walking, platform standing, wall jumping, and stereotypic gazing).
The exploration level of personality traits is reflected by the duration of forest musk deers’ exploratory behaviors, which include environmental sniffing and anogenital sniffing. The activity level is reflected by the duration of non-resting behaviors, which include moving (stepping, walking, trotting, running). The self-directed level is reflected by the duration of self-directed behaviors, which include self-grooming and self-scratching [9].
The duration proportion of stereotypic behaviors (SBs) was used to evaluate the stereotypic behaviors intensity (SBI) [9]. Similarly, the duration proportions of exploratory behaviors (EB), active behaviors (AB), and self-directed behaviors (SDB) were used to evaluate personality traits: exploration level (EL), activity level (AL), and self-directed level (SL), respectively [28]. The behavioral redundancy level (RL) was calculated as the complement of the behavioral diversity index (BDI) [29]. Thus, the stereotypic behavior intensity (SBI) and four personality traits (EL, AL, SL, RL) are calculated in this section. The formulas are as follows: , and .
2.4. Musk Collection and Muscone Measurement
A process table was drafted before musk collection, and the necessary tools were prepared, such as a sterile spoon and tray, the electronic scale, and sealable sampling bottles. After each musk deer was manually restrained by experienced keepers, the spoon was used to gently probe into the musk sac and extract the musk via rotation. The duration of the procedure was approximately 2–3 min per animal, and no distress could be observed, only mild struggling and an increased heart rate; this has been proven to be a non-invasive and sustainable harvesting procedure [30,31]. Then, the collected musk was placed on a tray for weighing and then stored in a sealed bag. The secretion amount of musk (SAM) collected from an individual musk deer was accurately recorded.
The muscone content (MC) was determined using the gas chromatography method [32]. A Shimadzu Nexis GC-2030 gas chromatograph (Shimadzu Corporation, Kyoto, Japan) equipped with an SH-Rtx-5 capillary column (30 m × 0.25 mm × 0.25 μm) was used. The test solution was prepared as follows: Take approximately 0.05 g of the dried sample, weigh it accurately, add 2 mL of anhydrous ethanol, seal it, and sonicate it for half an hour. Let it stand for 1 h, and then filter it to obtain the solution. The reference solution was prepared as follows: Take an appropriate amount of muscone reference substance, weigh it accurately, and dissolve it in anhydrous ethanol to prepare a solution of 1.5 mg·mL^−1^. The injection volume is 1.0 μL, and the solvent and sample are rinsed once and twice, respectively, before injection [33]. The temperature is set at 25 °C, the carrier gas is nitrogen, the pressure is 114.0 kPa, the total flow rate is 50.0 mL·min^−1^, the column flow rate is 1.91 mL·min^−1^, the equilibrium time is 3.0 min, and the tailing flow rate is 24 mL·min^−1^.
2.5. Statistical Analysis
The Shapiro–Wilk test was used to assess the normality of the data for all measured variables: stereotypic behavior intensity (SBI), exploration level (EL), activity level (AL), self-directed level (SL), behavioral redundancy level (RL), secretion amount of musk (SAM), and muscone content (MC). For comparisons among the three age groups (1.5, 2.5, and 3.5 years), one-way ANOVA was used for normally distributed variables (AL, SL, RL), while the Kruskal–Wallis test was applied to non-normally distributed variables (SBI, EL, SAM, MC). For pairwise comparisons between stereotypic and non-stereotypic groups within each age category, independent t-tests (for normally distributed data) or Mann–Whitney U tests (for non-normally distributed data) were used.
Pearson’s correlation analysis was used to evaluate correlations among normally distributed variables (AL, SL, RL), while Spearman’s correlation analysis was applied to non-normally distributed variables (EL, SBI, SAM, MC), based on the Shapiro–Wilk test results. The significance level was set at p = 0.05, and all data analyses were conducted with SPSS 26.0 [13].
3. Results
3.1. Stereotypic Behaviors of Captive Forest Musk Deer
Behavioral sampling results indicate that the forest musk deer population in this study developed stereotypic behaviors such as stereotyped licking and scraping, to-fro-walking, platform standing, wall jumping, and stereotypic gazing, and each type is described in Table 1.
3.2. Intensities of Stereotypic Behaviors
Analysis of stereotypic behaviors revealed that there is no statistically significant difference in the stereotypic behavior intensity across the three age groups (1.5, 2.5, and 3.5 years) of forest musk deer (p > 0.05) (Figure 1).
3.3. The Relationship Between Stereotypic Behaviors and Musk Secretion
Not all individuals displayed stereotypic behaviors. The results show that the SAM in the stereotypic behavior group (16.47 ± 6.12 g, N = 11) tended to be higher than that in the non-stereotypic behavior group (12.14 ± 4.96 g, N = 14), but the difference did not reach statistical significance (p > 0.05). Similarly, no significant differences were found in the muscone content between individuals that exhibited stereotypic behaviors (1.75 ± 0.88%, N = 11) and those that did not (1.56 ± 1.02%, N = 14) (p > 0.05) (Table 2).
To further explore the age-specific distribution of stereotypic behaviors, we stratified the samples by age groups and quantified the number of individuals with and without stereotypic behaviors in each group (Table 2). Statistical analysis revealed no significant differences in musk secretion between the stereotypic and non-stereotypic subgroups within each age group (all p > 0.05).
3.4. Personality Traits of Captive Forest Musk Deer
The correlation among the four personality traits is shown in Table 3. A significant positive correlation exists between EL and AL (Spearman’s ρ = 0.780, p < 0.05), while the other correlations do not reach significance (p > 0.05). In the analysis of the relationship between SB and the four personality traits, SBI exhibited a significant negative correlation with RL (Pearson’s r = −0.399, p < 0.05), while other correlations remained non-significant (p > 0.05).
The personality traits across various age groups, ranging from 1.5 to 3.5 years, are shown in Figure 2. No significant differences were found among age groups in exploration level, activity level, self-directed level, or behavioral redundancy level (all p > 0.05).
3.5. Relationship Between Personality Traits and Musk Secretion
Analysis of the relationship between personality traits and musk secretion revealed that EL (Spearman’s ρ = 0.262, p > 0.05), AL (Pearson’s r = 0.128, p > 0.05), and SL (Pearson’s r = 0.224, p > 0.05) showed positive but non-significant correlations with SAM. Similarly, RL demonstrated a negative but non-significant correlation with the SAM (Pearson’s r = −0.016, p > 0.05) (Table 4).
In the context of muscone content, EL was found to have a significant positive correlation (Spearman’s ρ = 0.461, p < 0.05), which can be expressed through the regression equation y = 0.83 + 4.76x (r^2^ = 0.21) (Figure 3). Similarly, AL exhibited a significant positive correlation with MC (Pearson’s r = 0.443, p < 0.05), represented by the regression equation y = 0.4 + 2.89x (r^2^ = 0.20) (Figure 4). Conversely, SL (Pearson’s r = −0.288, p > 0.05) and RL (Pearson’s r = −0.104, p > 0.05) were associated with negative but non-significant correlations with MC (Table 4).
4. Discussion
Captive environments designed for ex situ conservation often restrict natural ranging and foraging, predisposing small ungulates to stereotypic behaviors [34]. This study found that captive forest musk deer in Fujian Province displayed a higher frequency of head stereotypic behaviors, such as stereotypic rubbing. Notably, such head stereotypic behaviors have not been reported in previous studies of captive alpine musk deer, which primarily reported oral stereotypic behaviors (such as eating non-food items, stereotypic licking and scraping) [35]. Given that forest musk deer are smaller in size compared to alpine musk deer, it is speculated that they are more inclined to regulate energy metabolism and release stress through head movements under captive conditions, while alpine musk deer have evolved more oral stereotypic behaviors in restricted activities. This difference may be attributed to species-specific coping styles: smaller-bodied forest musk deer may rely more on locomotor and head movements to dissipate stress, whereas larger alpine musk deer may exhibit more oral stereotypies as a form of displacement activity [36].
In this study, although individuals exhibiting stereotypic behaviors showed a trend of higher secretion amount of musk and muscone content, this difference was not significant. Compared to in previous studies on captive forest musk deer [12,37], the average musk secretion in our study is within the normal range, indicating that the population is not impaired in musk production. This contrasts with some previous research conclusions [37,38] suggesting that the higher intensity of stereotypic behaviors developed in forest musk deer may reduce the behavioral diversity, thereby affecting animal welfare and musk production. In this study, the overall expression intensity of stereotypic behaviors in forest musk deer was relatively low, and the diversity index of individuals exhibiting stereotypic behaviors showed little difference compared to those not exhibiting such behaviors, thus not causing negative impacts on production efficiency.
Some studies have also indicated that, in some instances, low-intensity stereotypic behaviors may have a positive impact on animal welfare and productivity, serving as a form of stress regulation similar to self-enrichment or meditation [18]. Additionally, low-intensity stereotypic behaviors may be a coping mechanism that redirects metabolic resources and energy to key physiological functions, such as musk synthesis [20,39]. This redirection may be mediated by neuroendocrine changes, such as altered cortisol levels, which prioritize energy allocation towards reproduction or secretion [40]. For example, in this musk deer farm, individuals with relatively high stereotypic intensity (0.1) had a musk secretion of 19.4 g, but it should be noted that long-term or increased intensity of stereotypic behaviors may pose health risks. This energy reallocation might occur through reduced investment in non-essential activities (e.g., excessive vigilance) and optimized metabolic pathways [20]. Therefore, in practical management, a tiered strategy should be developed: For individuals with low-intensity stereotypic behaviors (<0.08), appropriate environmental enrichment can be used to maintain stress adaptation, and excessive intervention should be avoided to prevent the disruption of balance. For individuals with high-intensity stereotypic behaviors (≥0.10), priority should be given to more environmental enrichment and welfare enhancement measures [41,42].
This study found no significant differences in personality traits across different age groups of forest musk deer. We divided the individuals into three age groups (1.5, 2.5, and 3.5 years) to capture potential developmental changes during early maturity, as personality traits may stabilize after sexual maturity [21]. The grouping criterion was based on actual age (in years) recorded for each individual, ensuring clear separation of developmental stages. The lack of age-related differences indicates that personality development is largely completed by 1.5 years of age and remains relatively stable thereafter. The presentation of such findings may also be attributed to the limited age range of the sample, which did not cover the complete developmental stages (≥3.5 years), resulting in the age effect not being apparent. Future research should expand the age range to analyze the potential impact of developmental stages on behavior and personality.
This study demonstrated that forest musk deer with higher exploration and activity levels tend to have higher muscone content, indicating better musk quality. Additionally, personality traits of exploration and activity significantly promote the muscone content. This suggests that individuals with high EL and AL are better adapted to the captive environment, resulting in higher musk quantity and quality, which is consistent with previous findings [28]. The high-exploration and high-activity traits of forest musk deer may activate lipid metabolism pathways through frequent exposure to new stimuli, thereby facilitating the muscone synthesis. There is also a general tendency of explorative individuals to travel more and possess large home ranges or its core areas, which is good for their health and welfare [43,44,45]. Therefore, personality traits can serve as predictive indicators for musk secretion in forest musk deer. Thus, in breeding selection, individuals with higher musk quantity and quality can be identified based on their exploratory and activity performance. In the captive environment for forest musk deer, practical measures such as updating feeding devices and environmental enrichment can stimulate exploratory behavior and maintain optimal activity levels [25].
5. Conclusions
In summary, the low-intensity stereotypic behaviors observed in this study did not cause damage to the health and musk production of captive musk deer, which may represent an adaptative response to the captive environment. Therefore, one of the key implications of this research is that excessive intervention should be avoided to prevent disrupting this adaptive balance. The intensity of exploratory and active behaviors are important factors in enhancing musk quantity and quality. These two personality traits can be used as predictive indicators for musk secretion in breeding practices. Future research should integrate endocrine indicators, such as cortisol and sex hormones, to elucidate the molecular mechanisms affecting musk secretion. Furthermore, the implementation of a synergistic management model that prioritizes welfare production, based on behaviors and personality traits, could facilitate the development of specialized enclosures, thereby promoting the sustainable breeding of forest musk deer and sustainable musk production. Additionally, future research should include longer-term longitudinal studies to assess cumulative behavioral effects on population dynamics, and comparative studies across different latitudes to understand how environmental factors influence behavioral expression and musk production in captive musk deer.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yang Q.S. Meng X.X. Xia L. Feng Z.J. Conservation status and causes of decline of musk deer (Moschus spp.) in China Biol. Conserv.200310933334210.1016/S 0006-3207(02)00159-3 · doi ↗
- 2Jiang F. Zhang J.J. Gao H.M. Cai Z.Y. Zhou X.W. Li S.Q. Zhang T.Z. Musk deer (Moschus spp.) face redistribution to higher elevations and latitudes under climate change in China Sci. Total Environ.202070413533510.1016/j.scitotenv.2019.13533531784177 · doi ↗ · pubmed ↗
- 3Fan Z.X. Li W.J. Jin J.Z. Cui K. Yan C.C. Peng C.J. Jian Z.Y. Bu P. Price M. Zhang X.Y. The draft genome sequence of forest musk deer (Moschus berezovskii)Giga Science 20187 giy 03810.1093/gigascience/giy 03829635287 PMC 5906906 · doi ↗ · pubmed ↗
- 4IUCN The IUCN Red List of Threatened Species Available online: https://www.iucnredlist.org(accessed on 1 March 2026)
- 5Lu X.P. Geng S. Yang J.Y. Wu J.Y. Feng C.T. Li B.Q. Li R.X. An Z.X. Aryal A. Meng X.X. Transboundary conservation of musk deer (Moschus spp.): Global synthesis of populations, distributions, threats, and recovery strategies J. Nat. Conserv.20258712699110.1016/j.jnc.2025.126991 · doi ↗
- 6Meng X.X. Cody N. Gong B.C. Xiang L.L. Stable fighting strategies to maintain social ranks in captive male Alpine musk deer (Moschus sifanicus)Anim. Sci. J.20128361762210.1111/j.1740-0929.2011.01007.x 22862933 · doi ↗ · pubmed ↗
- 7Feng H. Wang L. Cao F.J. Ma J. Tang J. Feng C.L. Su Z.J. Forest musk deer (Moschus berezovskii) in China: Research and protection J. Vertebr. Biol.2023722206710.25225/jvb.22067 · doi ↗
- 8Zheng C.L. Zhao R.H. Meng Z.B. Hu D.F. Chen S.K. Liu B.Q. Wang J.M. Current situation and future perspectives of musk deer farming in China Mod. Chin. Med.20224416841692(In Chinese)