Fine Mapping of the Co-12 Anthracnose Resistance Gene in the Andean Common Bean Cultivar in Brazil
Jaqueline Bezerra da Silva, Maria Celeste Gonçalves-Vidigal, Pedro Soares Vidigal Filho, Giselly Figueiredo Lacanallo, Mariana Vaz Bisneta, Giseli Valentini, Larissa Fernanda Sega Xavier

TL;DR
Researchers identified the location of the Co-12 gene in common beans, which helps resist anthracnose disease, and found new candidate genes involved in this resistance.
Contribution
The study fine-maps the Co-12 resistance gene and identifies new candidate genes, including lipid transfer proteins, linked to anthracnose resistance in common beans.
Findings
The Co-12 gene was mapped to chromosome Pv04 within a 41 kb interval.
Three candidate genes, including two lipid transfer proteins (LTP2), were identified in the Co-12 region.
LTP2 genes suggest a new resistance mechanism against anthracnose in common beans.
Abstract
The common bean (Phaseolus vulgaris L.) cultivar Jalo Vermelho carries the Co-12 gene, which confers resistance to both Andean and Mesoamerican races of Colletotrichum lindemuthianum. Despite its importance for breeding programs, the genomic location and candidate genes underlying this resistance remain poorly defined. The Co-12 locus was fine-mapped using a biparental population derived from the cross Jalo Vermelho × Crioulo 159. A total of 172 F2 plants were used to generate 172 F2:3 families, which were phenotyped after inoculation with race 1545 of C. lindemuthianum. Segregation analysis confirmed a 1:2:1 Mendelian ratio, consistent with a single dominant resistance gene. Genotyping of resistant and susceptible plants using the BARBean6K_3 Illumina BeadChip (5398 SNP markers) mapped Co-12 to chromosome Pv04, between 1695 bp (ss715649768) and 9,651,954 bp (ss715646644). Subsequent…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1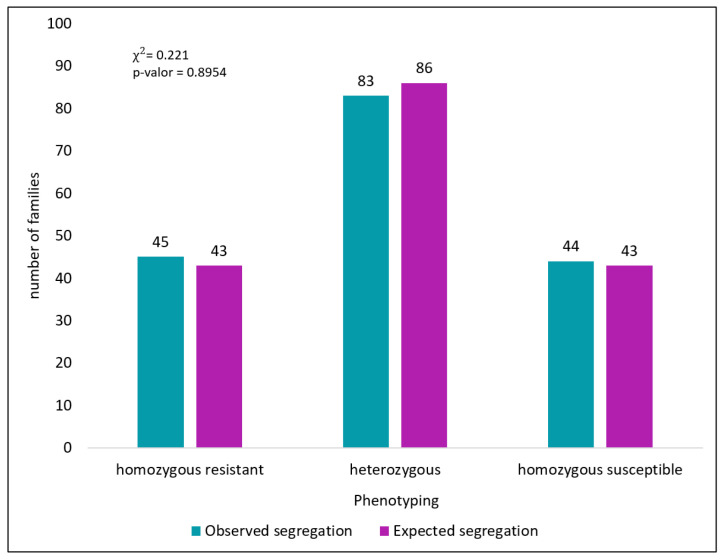 Figure 2
Figure 2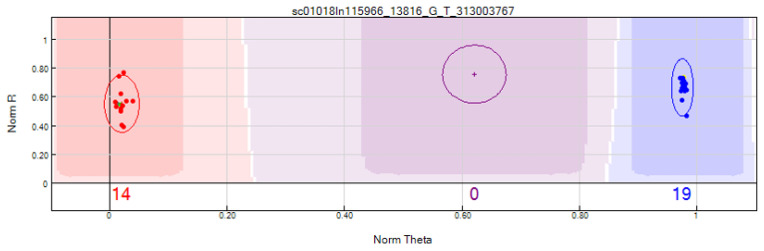 Figure 3
Figure 3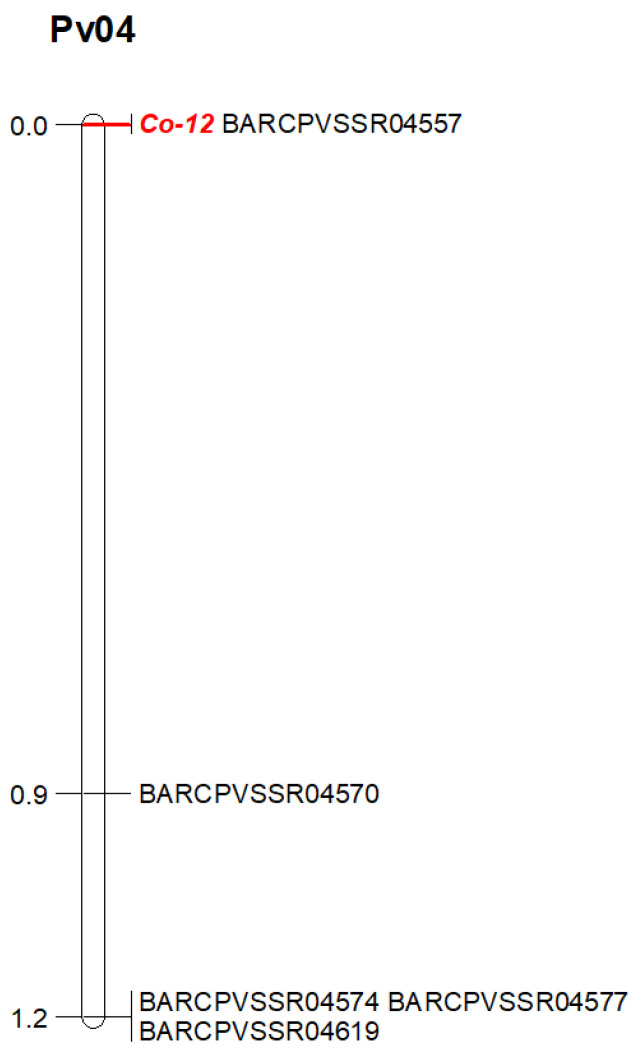 Figure 4
Figure 4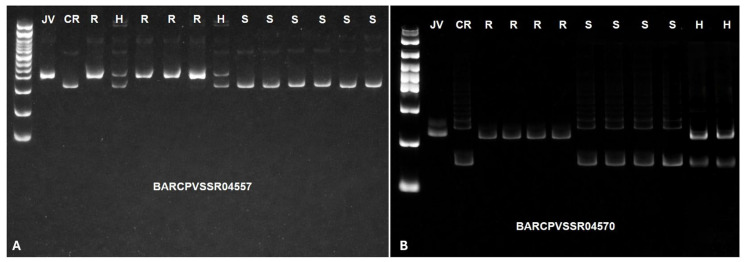 Figure 5
Figure 5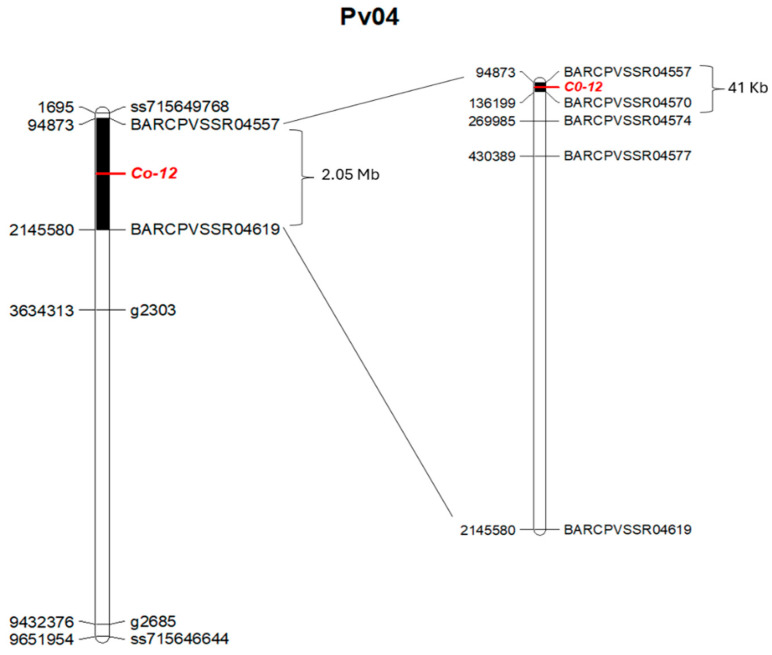 Figure 6
Figure 6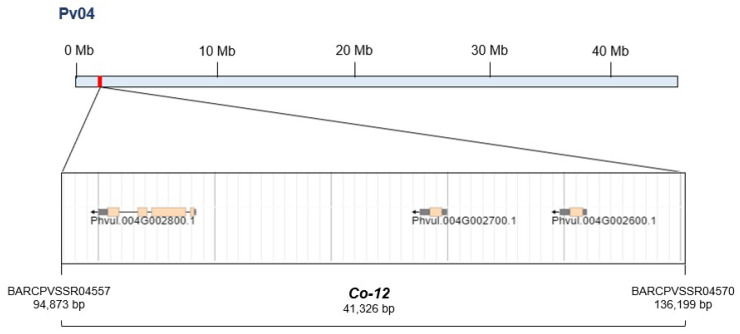 Figure 7
Figure 7- —National Council for Scientific and Technological Development (CNPq)
- —Coordination for the Improvement of Higher Education Personnel (Capes)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant pathogens and resistance mechanisms · Plant Pathogens and Fungal Diseases · Plant Pathogenic Bacteria Studies
1. Introduction
The genus Phaseolus comprises approximately 76 species, of which five have been domesticated: P. vulgaris (common bean), P. lunatus (lima bean), P. coccineus (ayocote bean), P. acutifolius (tepari bean), and P. dumosus [1,2]. Among these, the common bean (P. vulgaris L.) is the most widely cultivated and economically important species, serving as a major source of proteins, micronutrients, carbohydrates, dietary fiber, antioxidant molecules, phenolic compounds, and unsaturated fatty acids. Consequently, it constitutes a fundamental component of human nutrition for millions of people worldwide [3,4,5]. India, Myanmar, and Brazil are currently the leading producers of the common bean. In Brazil, total grain production for the 2025/2026 harvest is estimated at approximately 3.0 million tons [6], underscoring the strategic importance of the common bean for national food security and agribusiness.
Despite its agronomic importance, common bean production is constrained by several abiotic and biotic stresses. Among biotic stresses, anthracnose—caused by the fungus Colletotrichum lindemuthianum (Saccardo and Magnus) Briosi and Cavara—stands out as one of the most destructive diseases affecting this crop. Seeds represent the primary source of inoculum and pathogen dissemination, and disease damage is most severe during the early stages of plant development. Consequently, anthracnose can cause severe reductions in yield and grain quality [7,8,9].
To date, approximately 298 physiological races of C. lindemuthianum have been reported worldwide [10]. This high level of genetic and pathogenic variability greatly complicates disease management, requiring integrated strategies that include the use of certified seeds, chemical control, crop rotation, and, most importantly, resistant cultivars. Among these measures, the deployment of resistant cultivars is considered the most efficient, economical, and environmentally sustainable approach for anthracnose control [11,12]. Accordingly, common bean breeding programs play a central role in identifying resistance sources to diverse pathogen races and in developing cultivars with broad and durable resistance through gene pyramiding strategies [13].
C. lindemuthianum is traditionally classified into two major groups based on its virulence patterns: Mesoamerican and Andean. This classification reflects the genetic structure of the common bean, which is composed of two primary gene pools, Middle American and Andean. Andean races of the pathogen predominantly infect Andean bean genotypes, whereas Mesoamerican races exhibit a broader virulence spectrum, infecting both gene pools, with preferential virulence toward Middle American genotypes [14].
The anthracnose resistance genes are denoted by the symbol “Co” followed by a number or letter [13,15]. Several genes and allelic series have been described in both the Andean and Middle American gene pools of the common bean [16]. A total of 11 dominant genes and allelic series have been described in the Middle American gene pool: Co-2 (and allelic series Co-2^A252^ and Co-2^AB136^) [17,18,19,20]; Co-3 (and allelic series Co-3^2^, Co-3^3^, Co-3^4^, Co-3^5^, and Co-3^A252^) [19,21,22,23,24]; Co-4 (and allelic series Co-4^2^ and Co-4^3^) [25,26,27,28,29,30,31,32]; Co-5 (and allelic series Co-5^2^) [33,34,35]; Co-6 [16,36]; Co-11 [16]; Co-16 [37]; Co-17 [38]; Co-u [39]; Co-v [40]; and CoPv09cC [41].
In the Andean gene pool, 14 dominant genes and allelic series have been described: Co-1 (and allelic series Co-1^2^, Co-1^3^, Co-1^4^, Co-1^5^, Co-1^HY^, and Co-1^x^) [16,23,42,43]; CoPv01^CDRK^ [16]; and allelic series Co-3^K^ [44], Co-3^M^ [20,33], Co-3^W^ [21], Co-3^cX^ [41], Co-12 [45], Co-13 [16], Co-14 [16], Co-15 [46], Co-w [39], Co-x [47], Co-y [39,48], Co-z [39,48], Co-AC [16], Co-Pa [49], CoPv02cX [41], and Co-Bf [50,51].
Seeds of the common bean cultivar Jalo Vermelho were originally collected from small farms in the state of Paraná, southern Brazil, in 2001 [52]. This Andean cultivar represents an important source of resistance, exhibiting effective defense against thirteen races of C. lindemuthianum, including races 9, 23, 31, 55, 64, 65, 81, 83, 89, 95, 449, 453, and 1545 [45,49,52]. Genetic characterization of the Co-12 gene through inheritance and allelism tests demonstrated that Jalo Vermelho has a single dominant resistance gene located at a distinct locus, clearly different from previously described resistance genes [45]. Considering the relevance of this resistance source for common bean breeding programs, this study aimed to fine-map the Co-12 gene in Jalo Vermelho and to identify closely linked molecular markers to support marker-assisted selection for anthracnose resistance.
2. Results
2.1. Inheritance of Resistance to Race 1545 of Colletotrichum lindemuthianum
The Andean cultivar Jalo Vermelho showed complete resistance to race 1545 of Colletotrichum lindemuthianum, whereas the Mesoamerican cultivar Crioulo 159 was highly susceptible (Figure 1). The resistant control Widusa exhibited no disease symptoms, whereas the susceptible control Cornell 49-242 displayed typical anthracnose lesions, confirming the reliability of the inoculation procedure. Segregation analysis of the 172 F_2:3_ families derived from the Jalo Vermelho × Crioulo 159 cross fitted the expected Mendelian ratio of 1RR:2RS:1SS (χ^2^ = 0.221; p = 0.895) (Table 1; Figure 2), demonstrating that resistance to race 1545 in Jalo Vermelho is governed by a single dominant gene.
2.2. Mapping Using SNP Markers
Genotyping of 17 resistant (Table S1) and 18 susceptible (Table S2) F_2_ plants with the BeadChip platform revealed 150 SNP markers associated with resistance to race 1545 (Table S3). These SNPs delimited the Co-12 gene to a region spanning from 1695 bp (ss715649768) to 9,651,954 bp (ss715646644) on chromosome Pv04. Clear clustering of resistant and susceptible genotypes relative to the parental lines confirmed linkage between these markers and the resistance locus (Figure 3).
2.3. Mapping Using SSR Markers and a Linkage Map
A total of 22 SSR markers were developed for the region delimited by the SNP markers ss715649768 and ss715646644. All SSR markers were analyzed in the parental lines Jalo Vermelho and Crioulo 159, and the results revealed five polymorphic markers between the parents (Table 2). These markers were subsequently analyzed in the 172 F_2:3_ families (Table S4). The chromosomal region containing these five SSR markers was delimited by the SNP markers ss715649772 (33,578 bp) and ss715646246 (2,428,047 bp) based on the G19833 reference genome (v2.1), available at Phytozome: “https://phytozome-next.jgi.doe.gov/ (accessed on 11 December 2025)”.
The anthracnose resistance gene in Jalo Vermelho was closely linked to the marker BARCPVSSR04557 at 0.0 cM and located 0.9 cM from the marker BARCPVSSR04570 (Figure 4). The SSR banding patterns corresponding to homozygous resistant, homozygous susceptible, and heterozygous individuals are shown in the genotyping gels using the markers BARCPVSSR04557 and BARCPVSSR04570 (Figure 5). Based on the physical position of the SSR markers and the anthracnose reaction of the F_2:3_ families, four F_2_ plants exhibited recombination breakpoints between markers BARCPVSSR04557 (95 kb) and BARCPVSSR04570 (136 kb). These results delimit the anthracnose resistance gene in Jalo Vermelho to a 41 kb region on chromosome Pv04 (Figure 6; Table 3).
2.4. Candidate Genes and Associated Functions
The 41 kb genomic region harboring the Co-12 resistance locus, delimited by the markers BARCPVSSR04557 and BARCPVSSR04570 on chromosome Pv04, contains three annotated candidate genes in the common bean reference genome (Figure 7). These genes include Phvul.004G002800, which encodes a gamma-glutamyl-GABA protein, and Phvul.004G002700 and Phvul.004G002600, both of which encode lipid transfer proteins of the LTP2 family. The gene Phvul.004G002800 shows 71.9% sequence similarity to the Arabidopsis thaliana gene AT5G38200, which is associated with amino acid metabolism and cellular stress responses. The genes Phvul.004G002700.1 and Phvul.004G002600.1 exhibit 44.7% and 53.8% sequence similarity, respectively, with the A. thaliana gene AT3G18280.1, which encodes a lipid transfer protein involved in membrane stability, lipid transport, and defense-related processes (Table 4).
3. Discussion
Cultivars resistant to multiple races of Colletotrichum lindemuthianum are essential for breeding programs targeting durable anthracnose resistance. In this context, the Andean cultivar Jalo Vermelho represents a particularly valuable resistance source, as it is resistant to both Andean and Mesoamerican races, including races 9, 23, 31, 55, 64, 65, 81, 83, 89, 95, 449, 453, and 1545 [45,49,52]. Compared with other Andean cultivars, Jalo Vermelho shows a broader resistance spectrum, surpassing differential cultivars such as Michigan Dark Red Kidney and Perry Marrow, as well as Paloma, Jalo Listras Pretas, Pitanga, Corinthiano, Beija Flor, and Jalo EEP 558 (Table S5) [49]. In addition, its resistance to race 63-39 of Pseudocercospora griseola, the causal agent of angular leaf spot, further highlights its relevance as a multi-disease resistance source for breeding programs.
Race 1545 of C. lindemuthianum, used in this study, has been identified in Colombia, Costa Rica, Guatemala, Mexico, and Honduras, demonstrating its wide geographic distribution and epidemiological importance [53,54]. The presence of this race across diverse bean-producing regions reinforces the need to identify and characterize resistance genes effective against race 1545, as such genes are likely to contribute to broader and more durable disease control.
Previous evaluations of F_2:3_ families derived from the Jalo Vermelho × Crioulo 159 cross, inoculated with races 55 and 1545, demonstrated cosegregation of the Co-12 gene for both races [55]. In this study, the inoculation of 172 F_2:3_ families with race 1545 resulted in the expected Mendelian segregation ratio of 1:2:1, confirming the monogenic inheritance of Co-12. Furthermore, genotypic analysis revealed the precise localization of Co-12 on the upper arm of chromosome Pv04 within a 41 kb genomic interval flanked by BARCPVSSR04557 and BARCPVSSR04570. The tight linkage between Co-12 and BARCPVSSR04557 highlights its practical value for marker-assisted selection, facilitating the development of cultivars with durable anthracnose resistance. These markers could be used in marker-assisted selection; however, they have not yet been validated in independent populations.
The upper arm of chromosome Pv04 has been reported as a genomic region containing an important cluster of genes conferring resistance to multiple pathogens. This cluster includes genes for resistance to rust (Ur-5, Ur-14, Ur-Dorado-108, Ur-PI310762 and Ur-19833) [56,57,58,59], angular leaf spot [24,60,61], bean golden yellow mosaic virus (BGYMV) [62], and anthracnose. The presence of multiple resistance genes in this cluster highlights its potential as a key target for breeding programs aimed at developing cultivars with broad and durable resistance to different diseases.
Numerous anthracnose resistance genes from both Andean and Mesoamerican gene pools have been mapped to Pv04, including the Co-3 allelic series (Mexico 222, Mexico 227, PI 207262, BAT 93, Ouro Negro, G2333, MDRK, Kaboon, Widusa, A 252, and Xana) [19,21,22,23,24,33,41,44,63], as well as Co-y and Co-z in Jalo EEP 558 [48], Co-15 in Corinthiano [46], Co-16 in Crioulo 159 [37], and Co-Bf in Beija Flor [51]. In addition, GWASs have identified loci associated with resistance to races 2, 4, 7, 9, 65, 81, and 109 on Pv04 [16,42,64,65,66]. This dense accumulation of resistance loci confirms that Pv04 is a central genomic region for anthracnose resistance breeding.
Allelism tests performed between Jalo Vermelho and multiple resistant cultivars demonstrated that Co-12 is genetically independent from several resistance genes, including Co-1^3^, Co-2, Co-3, Co-3^3^, Co-3^M^, Co-3^K^, Co-3^W^, Co-3^4^, Co-4, Co-4^3^, Co-5, Co-6, Co-7, Co-9, Co-11, Co-13 [45], Co-15 [46], Co-16 [37], and Co-Bf [50], as all the crosses exhibited the expected 15:1 resistance–susceptible segregation ratio. This genetic independence reinforces the importance of Co-12 and supports its use in pyramiding strategies aimed at enhancing resistance durability.
Comparative physical mapping further supported the distinct position of Co-12. The Co-3^4^ allele, which cosegregates with the Phg-ON gene in Ouro Negro, is located approximately 3.2 Mb from BARCPVSSR04570 [24], whereas Co-15 in Corinthiano lies approximately 8.9 Mb from the Co-12 region [46]. The Co-Bf gene from Beija Flor is located downstream of Co-12, between markers SS333 and SS509 [51], representing the closest mapped resistance gene to Co-12. These distances confirm that Co-12 occupies a unique genomic position within the Pv04 resistance cluster.
Within the 41 kb Co-12 interval, three candidate genes were identified: one encoding a gamma-glutamyl-GABA protein and two encoding a lipid transfer protein (LTP2). Lipid transfer proteins are widely recognized as multifunctional components of plant defense and participate in cuticle formation, signal transduction, and direct antimicrobial activity. Several studies have demonstrated that LTPs can inhibit fungal growth, contribute to cell wall reinforcement, and modulate host–pathogen interactions. Therefore, the presence of two LTP2 genes within the Co-12 genomic interval strongly suggests a potential role in resistance to C. lindemuthianum. Most disease resistance genes in plants belong to the NBS-LRR or kinase families, which are highly abundant on chromosomes Pv04, Pv10, and Pv11 in the common bean [65,67,68]. In contrast, lipid transfer proteins are less commonly associated with resistance loci, although they have long been recognized for their antifungal and antibacterial activities [69,70]. LTP-mediated defense responses have been reported in Capsicum annuum [71], Brassica rapa [72], Triticum durum [73], Arabidopsis thaliana [74], and Gossypium hirsutum [75].
Similarity analysis against A. thaliana revealed identities ranging from 44.7% to 71.9%. The gene annotated as Gamma-glutamyl-gamma-aminobutyrate hydrolase (Gamma-glutamyl-GABA) in Phytozome showed 71.9% similarity, confirming its predicted function. Genes annotated as LTP2 in Phytozome displayed moderate similarity (44.7–53.8%) to Tracheary Element Differentiation-Related 4 in Arabidopsis. Previous studies in Zinnia mesophyll cells revealed differentiation-specific genes associated with tracheary element formation that exhibit typical LTP features [76]. Notably, in Phytozome, the LTP2 genes exhibit 98.1% sequence similarity with those in Phaseolus vulgaris and 83.7% with those in Phaseolus acutifolius, suggesting that despite evolutionary divergence and differences in nomenclature, conserved domains allow inference of the likely functions of these candidate genes in the common bean.
This study represents the first report linking LTP2 genes to anthracnose resistance in Phaseolus vulgaris. The identification of LTP2 genes within the Co-12 interval suggests that this resistance locus may involve a noncanonical defense mechanism, expanding the current understanding of resistance pathways in the common bean.
Although fine mapping narrowed the target region to a small genomic interval containing three candidate genes, further studies are needed to functionally validate these genes. Specifically, gene expression analyses and gene-editing approaches should be used to identify causal genes. In addition, comprehensive validation of the associated molecular markers across diverse genetic backgrounds and environments is necessary to confirm their stability and reliability for effective application in breeding programs.
4. Materials and Methods
4.1. Experimental Site and Plant Material
The experiments were conducted in a greenhouse at the Núcleo de Pesquisa Aplicada à Agricultura (Nupagri), Universidade Estadual de Maringá (UEM), Paraná, Brazil (23°26′7.744″ S, 51°53′43.288″ W), between July 2018 and December 2022. The seeds of all the cultivars used in this study were obtained from the Common Bean Germplasm Bank (BGF) of UEM.
The plant material was derived from a biparental cross between the Andean cultivar Jalo Vermelho and the Mesoamerican cultivar Crioulo 159. Crosses were performed in a greenhouse in 2018. In 2019, F_1_ plants were grown, and F_2_ seeds were subsequently harvested. In 2020, F_2_ seeds were sown to generate F_2:3_ families. When the trifoliate leaves of F_2_ plants had expanded, leaf tissue was collected for DNA extraction. The plants were maintained in the greenhouse to produce seeds, with each F_2_ plant harvested separately, generating 172 F_2:3_ families. In 2021, the F_2:3_ families were phenotyped, and F_2_ plants were genotyped using the BeadChip platform; in 2022, all F_2_ plants were genotyped with SSR markers.
To increase the number of seeds, seeds from the F_1_, F_2_, and parental plants were sown individually in plastic pots (8 × 8 × 12 cm) filled with biostabilized Pinus bark substrate (MecPlant—Registration EP PB 09549-4/MAPA Brasil, MEC PREC Ind. Com. Ltda., Telêmaco Borba, PR, Brazil) and maintained under greenhouse conditions until harvest.
4.2. Inoculum Preparation, Inoculation and Evaluation
The Colletotrichum lindemuthianum race 1545 isolate was cultured on Petri dishes containing potato-dextrose-agar medium. For inoculum production, test tubes containing bean pods (8–10 cm) partially immersed (1–2 cm) in water–agar medium were autoclaved at 120 °C for 40 min. Small fragments of the isolate race 1545 of C. lindemuthianum from the Petri dish were placed on the pods using a metal loop that was sterilized via flame sterilization; this procedure was carried out in a laminar flow hood. The tubes were then incubated in a growth chamber at 20 ± 2 °C for 14 days to promote sporulation [77].
After incubation, the pods containing spores were removed using sterile forceps and transferred to beakers containing autoclaved distilled water. The suspension was filtered through double-layer gauze to obtain a spore suspension. The spore concentration was adjusted to 1.2 × 10^6^ conidia mL^−1^ using a Neubauer hemocytometer, and one drop of Tween 20 was added per 100 mL of suspension [78].
For anthracnose evaluation, the F_2:3_ families were sown in plastic trays (48 × 30 × 11 cm) filled with MecPlant substrate. Each family consisted of approximately 8–10 seeds, depending on seed availability, resulting in a total of 1651 F_3_ plants evaluated phenotypically. Comparable family sizes have been successfully employed in previous studies analyzing resistance inheritance in F_2:3_ populations inoculated with C. lindemuthianum races, demonstrating that this sampling strategy is sufficient to detect segregation patterns and reliably assess resistance inheritance [51,58]. Six genotypes were sown per tray, with seeds spaced 8 cm apart between genotypes and 3 cm between individual seeds, including the parents, controls, and F_2:3_ families.
At the V3 growth stage, corresponding to the full expansion of the first trifoliate leaf, the trays were transferred to a fog chamber (20 ± 2 °C) for inoculation. The differential cultivars Widusa (resistant) and Cornell 49-242 (susceptible) were included as controls.
Each plant was individually inoculated with race 1545 of C. lindemuthianum and considered an independent biological replicate for assessing disease response. This experimental design enabled accurate estimation of the proportion of resistant and susceptible plants within each family and supported reliable genetic segregation analysis of the trait.
The plants were inoculated by spraying the suspension onto both the abaxial and adaxial surfaces of the trifoliate leaves and applying one spray per leaf on each surface. Trays were then incubated for 72 h at 20 ± 2 °C under controlled lighting (12 h light at 680 lux and 12 h darkness) and a relative humidity above 95%.
Anthracnose symptoms can be observed between 10 and 15 days after inoculation, depending on the virulence of the isolate under study. In this population, visual assessments were conducted 10 days after inoculation, as the virulence of race 1545 produced pronounced symptoms. The standard anthracnose severity scale was used, with the following scores: 1—Absence of symptoms; 2—Up to 1% of the veins show necrotic spots, noticeable only on the underside of the leaves; 3—Up to 3% of the veins show necrotic spots, noticeable only on the underside of the leaves; 4—Up to 1% of the veins show necrotic spots, noticeable on both sides of the leaves; 5—Up to 3% of the veins show necrotic spots, noticeable on both sides of the leaves; 6—Necrotic spots on the veins, noticeable on both sides of the leaves, with some lesions on stems, branches, and petioles; 7—Necrotic spots on most veins and a large portion of adjacent mesophyll tissue, which ruptured, with abundant lesions on stems, branches, and petioles; 8—Necrotic spots on almost all veins, very abundant on stems, branches, and petioles, causing ruptures, defoliation, and reduced plant growth; 9—Plant death. Scores from 1 to 3 indicate resistant plants, and scores from 4 to 9 indicate susceptible plants [79]. Families were classified as homozygous resistant when all the evaluated F_3_ plants were resistant, as susceptible when all the evaluated F_3_ plants were susceptible, and as heterozygous when the F_3_ plants showed a mix of resistant and susceptible individuals.
4.3. Inheritance Analysis
Previous inheritance studies have shown that Jalo Vermelho has a single dominant gene [45]. To confirm these results, the inheritance of resistance in the Jalo Vermelho cultivar was evaluated using the 172 F_2:3_ families inoculated with race 1545 of Colletotrichum lindemuthianum. Based on the phenotypic classification of resistant, susceptible and heterozygous families, segregation ratios were analyzed using the chi-square (χ^2^) test to verify conformity with the expected Mendelian segregation for a single dominant gene in the F_2_ population. A bar graph was constructed to better visualize the results of the chi-square test.
4.4. SNP Genotyping and Primary Mapping
Given that Jalo Vermelho carries a single gene, we decided to perform fine mapping by first genotyping a subset of the F_2_ plants with 5398 SNPs using the BeadChip to identify the chromosome and approximate region where the gene is located. SSR markers were subsequently designed within this specific region to genotype the entire population and further narrow the location of the gene [49,51,58,59].
Genomic DNA was extracted from 172 F_2_ plants and the parental cultivars using the PureLink Genomic Plant DNA Purification Kit (Thermo Fisher Scientific, Waltham, MA, USA), following the manufacturer’s instructions. Based on the phenotyping of the F_2:3_ families with race 1545 of Colletotrichum lindemuthianum, 17 resistant and 18 susceptible families were selected, and the DNA of the corresponding F_2_ progenies was sent to Soybean Genomics and Improvement Laboratory, USDA-ARS/BARC-W (Beltsville, MD, USA), for genotyping using the BARBean6K_3 Illumina BeadChip containing 5398 SNP markers on the Illumina Infinium^®^ HD Assay Ultra platform [80].
Fluorescence signals were captured with an Illumina BeadArray Reader (Illumina, Inc., San Diego, CA, USA), and allele calling was performed using GenomeStudio V2011.1 (Illumina, Inc.). All 5389 SNPs were visually inspected and manually curated when necessary. SNPs were distributed across all 11 chromosomes of the common bean; however, SNPs linked to the Co-12 gene were identified exclusively on chromosome Pv04. SNPs were considered linked to Co-12 when they were polymorphic between parents and when resistant F_2_ individuals clustered with the resistant parent, while susceptible individuals clustered with the susceptible parent.
4.5. SSR Marker Development and Genotyping
Genomic fragments containing SNPs from the BeadChip were used to develop SSR markers [58]. SNPs associated with the Co-12 region were selected, and the corresponding SSR markers were used to genotype the 172 F_2_ individuals.
PCRs were performed with 20 µL reaction mixtures containing 50 ng of genomic DNA, 0.25 µM of each primer, PCR buffer, 1.5 mM MgCl_2_, 0.2 mM of each dNTP, Taq DNA polymerase (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA), and ultrapure water. Amplified products were separated on 6% nondenaturing polyacrylamide gels and stained with 0.02% SYBR Safe. Bands were visualized under ultraviolet light, and images were captured using an L-PIX Image EX system (Loccus Biotecnologia, Cotia, SP, Brazil).
The physical positions of SSR markers were determined by BLAST analysis against the Phaseolus vulgaris v2.1 reference genome using the Phytozome database: “https://phytozome-next.jgi.doe.gov/ (accessed on 14 January 2026)”.
4.6. Linkage Map Construction
Genetic linkage analysis was performed using JoinMap 4.0 [81]. Marker ordering and distance estimation were carried out using the regression mapping algorithm with the Kosambi mapping function. Linkage groups were established using a minimum LOD threshold of 3.0 and a maximum recombination distance of 50 cM. The final linkage map was visualized using MapChart 2.3 [82].
4.7. Candidate Gene Identification
The physical position of Co-12 was determined based on the flanking SSR markers using BLAST analysis against the Phaseolus vulgaris v2.1 reference genome in Phytozome. All annotated genes located within the marker-defined interval were retrieved, and those with predicted functions related to plant defense or stress response were considered potential Co-12 candidate genes.
5. Conclusions
The Andean common bean cultivar Jalo Vermelho represents a highly valuable source of anthracnose resistance, conferring broad effectiveness against multiple races of Colletotrichum lindemuthianum. The Co-12 gene was precisely mapped to a 41 kb genomic region on chromosome Pv04, confirming its monogenic inheritance and genetic independence from previously described resistance loci within the same chromosomal cluster. The closely linked SSR markers BARCPVSSR04557 and BARCPVSSR04570 provide reliable tools for marker-assisted selection and will facilitate the efficient introgression of Co-12 into elite breeding lines. The identification of candidate genes encoding gamma-glutamyl-GABA proteins and lipid transfer proteins suggests that Co-12-mediated resistance may involve a noncanonical defense mechanism distinct from that of classical NBS-LRR pathways. These findings expand current knowledge of resistance mechanisms in Phaseolus vulgaris and highlight the diversity of host defense strategies against anthracnose. Overall, the high-resolution mapping of Co-12 and the development of closely linked molecular markers provide important genomic resources for common bean breeding programs and contribute directly to the development of cultivars with durable and broad-spectrum anthracnose resistance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Debouck D. Diversity in Phaseolus species in relation to the common bean Common Bean Improvement in the Twenty-First Century Singh S.P. Kluwer Dordrecht, The Netherlands 19992552
- 2Bitocchi E. Rau D. Bellucci E. Rodriguez M. Murgia M.L. Gioia T. Santo D. Nanni L. Attene G. Papa R. Beans (Phaseolus ssp.) as a model for understanding crop evolution Front. Plant Sci.2017872210.3389/fpls.2017.0072228533789 PMC 5420584 · doi ↗ · pubmed ↗
- 3Gepts P. Aragão F. Barros E. Blair M. Brondani R. Broughton W. Galasso I. Hernandez G. Kami J. Lariguet P. Genomics of Phaseolus beans, a major source of dietary protein and micronutrients in the tropics Genomics of Tropical Crop Plants Moore P.H. Ming R. Springer New York, NY, USA 2008113143
- 4Celmeli T. Sari H. Canci H. Sari D. Adak A. Eker T. Toker C. The nutritional content of common bean (Phaseolus vulgaris L.) landraces in comparison to modern varieties Agronomy 2018816610.3390/agronomy 8090166 · doi ↗
- 5Vieira N.M. Peghinelli V.V. Monte M.G. Costa N.A. Pereira A.G. Seki M.M. Azevedo P.S. Polegato B.F. Paiva S.A.R. Zornoff L.A.M. Beans consumption can contribute to the prevention of cardiovascular disease Clin. Nutr. ESPEN 202355738010.1016/j.clnesp.2023.01.00736963901 · doi ↗ · pubmed ↗
- 6Conab Companhia Nacional de Abastecimento Available online: www.conab.gov.br(accessed on 11 December 2025)
- 7Melotto M. Kelly J.D. Sciences S. Lansing E. Melotto M. Kelly J.D. An allelic series at the Co-1 locus conditioning resistance to anthracnose in common bean of Andean origin Euphytica 200011614314910.1023/A:1004005001049 · doi ↗
- 8Alzate-Marin A.L. Costa M.R. Menarim H. Moreira M.A. Barros E.G. Herança da resistência à antracnose na cultivar de feijoeiro comum Cornell 49-242Fitopatol. Bras.20032830230610.1590/S 0100-41582003000300013 · doi ↗