Genome-Wide Identification of CFEM Proteins in Sclerotinia sclerotiorum Reveals Effector Candidates with Cell Death Suppression Activity
Xihong Li, Yuting Wu, Linxuan Liu, Shuang Liu, Dan Zhang, Xianfeng Yi, Lele Wang, Shan Liu, Rongchao Jia, Jinpeng Shi, Stefan Olsson, Congcong Lu, Airong Wang, Ya Li

TL;DR
This study identifies and characterizes CFEM proteins in the fungus Sclerotinia sclerotiorum, revealing some as potential virulence factors that suppress cell death in plants.
Contribution
The first comprehensive identification and functional analysis of CFEM proteins in Sclerotinia sclerotiorum, including their roles in suppressing cell death.
Findings
Thirteen CFEM proteins were identified in Sclerotinia sclerotiorum with diverse properties and predicted functions.
SsCFEM4 and SsCFEM9 were shown to suppress cell death in Nicotiana benthamiana, indicating their role as effectors.
Transcriptomic data revealed upregulation of most SsCFEM genes during early plant infection.
Abstract
The CFEM (Common in Fungal Extracellular Membrane) domain defines a family of cysteine-rich proteins unique to fungi, playing pivotal roles in host–pathogen interactions. However, the repertoire and functions of CFEM proteins in the broad-host-range necrotrophic pathogen Sclerotinia sclerotiorum remain largely unexplored. Through genome-wide bioinformatic analysis, we identified 13 CFEM-containing proteins (SsCFEM1–13) in S. sclerotiorum. Characterization revealed substantial diversity in their physicochemical properties, domain architecture, and predicted subcellular localization. Ten proteins possess a secretion signal, with six predicted to be GPI-anchored and three classified as high-confidence effectors. Members lacking transmembrane domains were predicted to adopt the conserved CFEM “helical-basket” fold. Phylogenetic analysis grouped SsCFEMs into two distinct clades and indicated…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
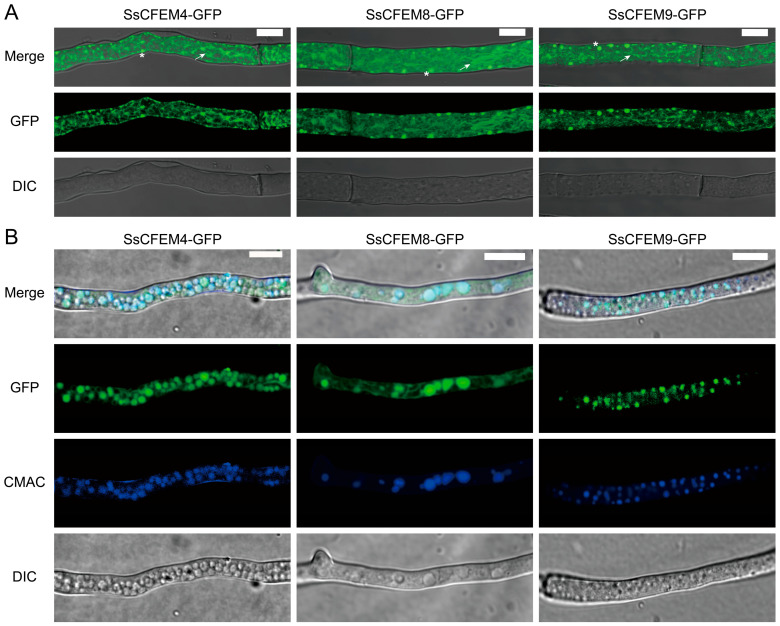 Figure 1
Figure 1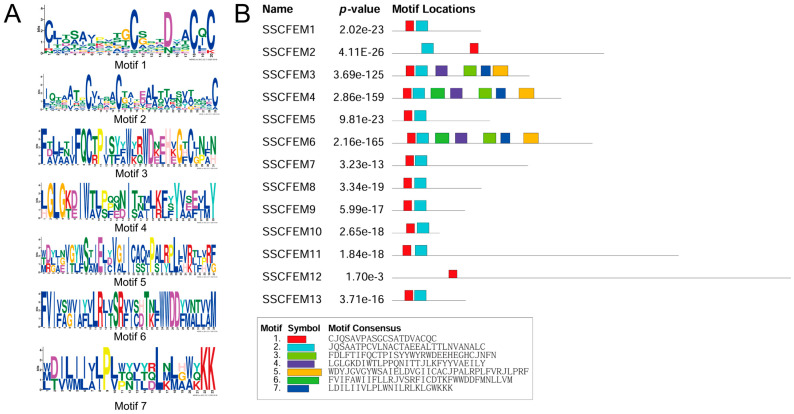 Figure 2
Figure 2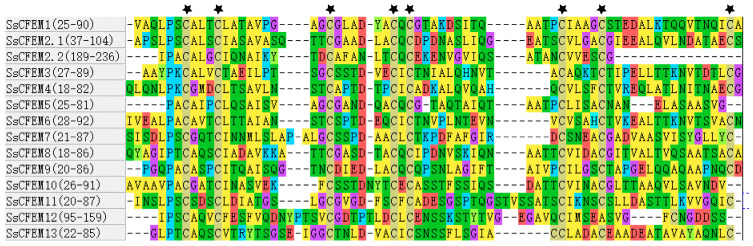 Figure 3
Figure 3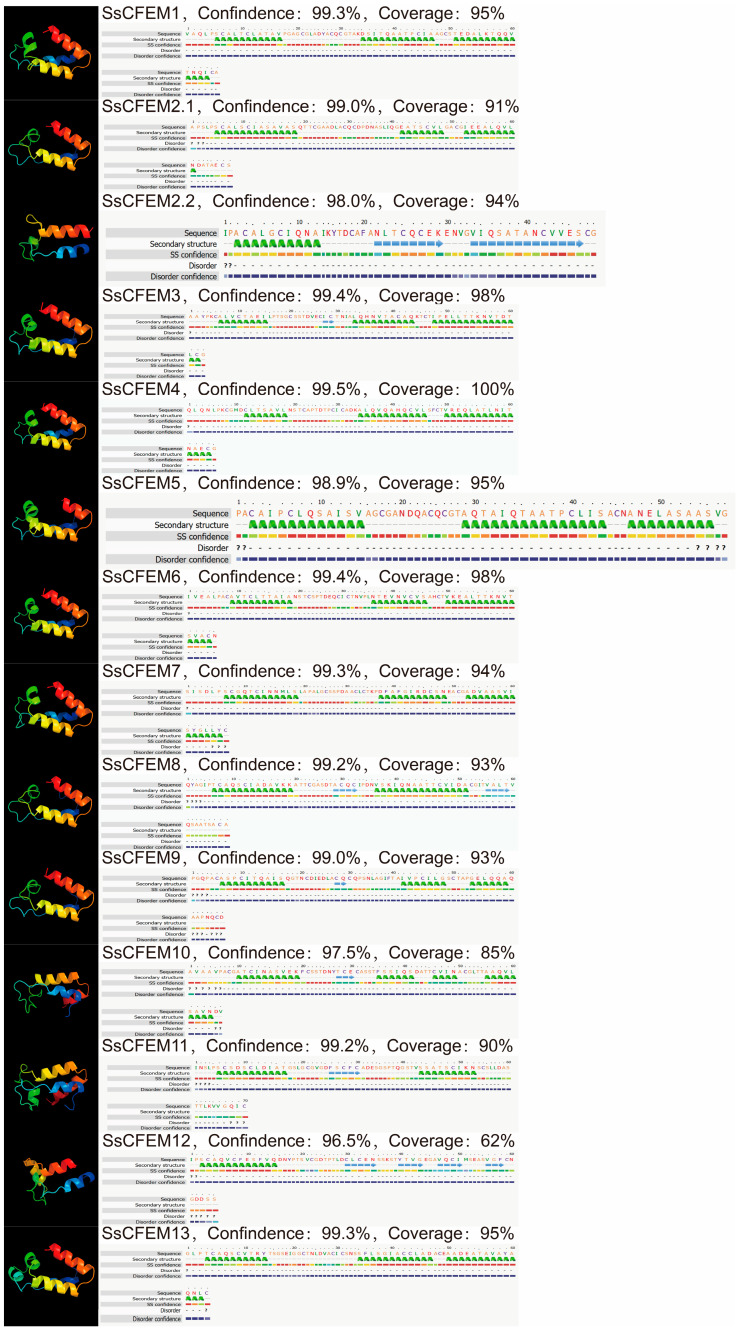 Figure 4
Figure 4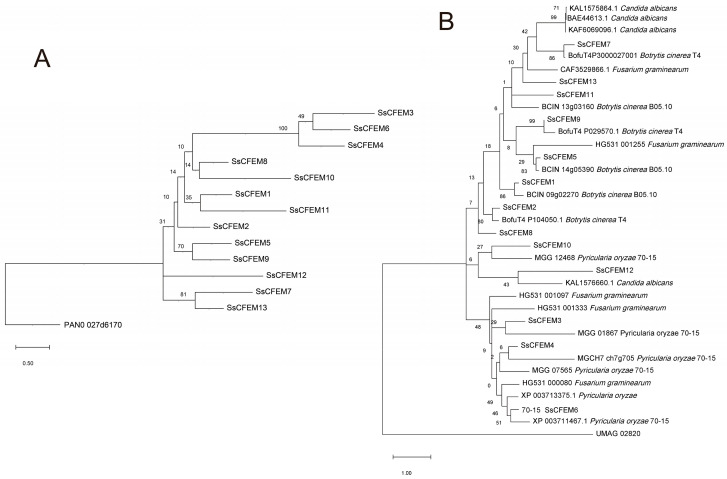 Figure 5
Figure 5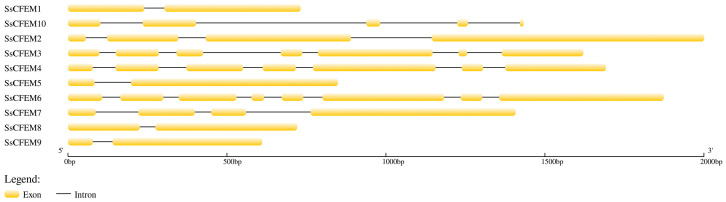 Figure 6
Figure 6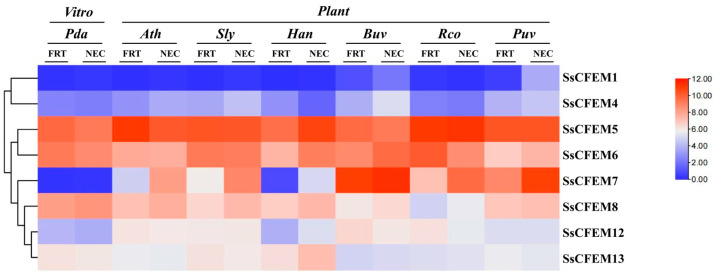 Figure 7
Figure 7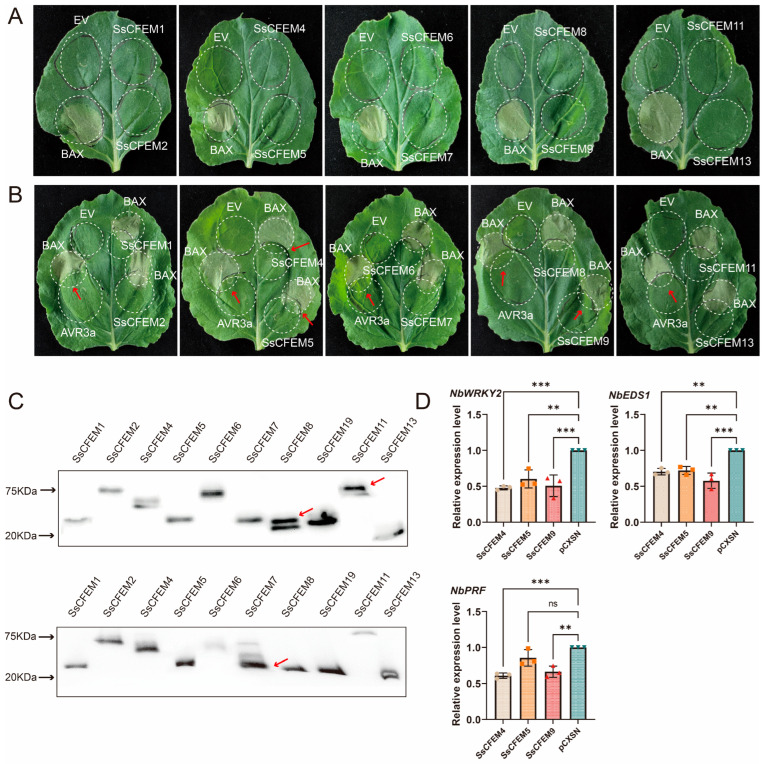 Figure 8
Figure 8- —Key Program of Science and Technology in Fujian Province, China
- —National Key Research and Development Program of China
- —National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant pathogens and resistance mechanisms · Fungal and yeast genetics research · Plant-Microbe Interactions and Immunity
1. Introduction
The CFEM (Common in Fungal Extracellular Membrane) domain is a fungi-specific cysteine-rich extracellular domain characterized by the presence of eight conserved cysteine residues that form a specific disulfide bond network, stabilizing its three-dimensional structure [1]. Initially identified through the analysis of over 25 fungal protein sequences, its structural features are distinct from other known cysteine-rich domains, suggesting a unique role in fungal biology [1]. Subsequent systematic phylogenomic analyses confirmed that the CFEM domain is indeed unique to fungi, with a single origin dating back to the most recent common ancestor of Ascomycota and Basidiomycota, rather than multiple independent origins [2]. The length and architecture of this domain are relatively conserved, but its copy number varies significantly among different fungal species. Notably, pathogenic fungi generally possess a larger number of CFEM domains, implying a potentially important role in host–pathogen interactions [2].
In human pathogenic fungi, CFEM proteins have been extensively characterized, especially in Candida albicans, where they are crucial for iron acquisition—a key virulence determinant. The GPI-anchored proteins Rbt5 and Pga7 bind heme and facilitate its utilization from hemoglobin, forming a relay network that delivers heme to the endocytic machinery [3,4]. Another CFEM protein, Csa2, also supports heme-iron uptake, particularly during hyphal growth [5]. Proteomic studies confirm that several CFEM proteins are upregulated under iron limitation, and their domains directly bind heme, underscoring their central role in iron metabolism [6]. This heme-acquisition system functionally cooperates with ferric reductase-related proteins (Frp1/Frp2), suggesting an evolutionary adaptation from ancestral reductases [7]. Beyond iron uptake, CFEM proteins in C. albicans contribute to biofilm integrity, cell surface architecture, and adhesion, with mutants showing increased sensitivity to wall stressors and altered hydrophobicity [8,9]. Their expression is induced by hypoxia and sterol inhibitors, regulated by transcription factors Upc2 and Bcr1 [10]. Functional conservation and divergence are observed in other pathogenic yeasts: While C. parapsilosis CFEM proteins retain roles in iron acquisition, their involvement in biofilm formation differs [11], and in C. glabrata, the CFEM protein CgCcw14 is vital for iron homeostasis, host cell adherence, and virulence [12].
In plant pathogenic fungi, CFEM proteins are established key virulence factors with multifunctional roles in host recognition, infection structure development, immune suppression, and nutrient acquisition. In necrotrophic pathogens such as Botrytis cinerea, CFEM proteins like BcCFEM1 are upregulated during early infection and contribute to pathogenicity, conidiation, and stress tolerance [13]. A non-GPCR membrane-bound CFEM protein in the same fungus also influences lesion progression and conidial germination [14]. Comparative secretome analyses further suggest that CFEM-containing effectors may differ among necrotrophs, reflecting pathogen-specific adaptation [15]. In biotrophic/hemibiotrophic pathogens, CFEM proteins exhibit even greater functional diversity. In Magnaporthe oryzae, CFEM-containing GPCRs such as Pth11 and WISH are essential for surface sensing, appressorium differentiation, and pathogenicity [16,17,18,19]. Genome-wide studies confirm that CFEM-GPCRs are expanded in phytopathogens and induced during infection [20]. In Verticillium dahliae, secreted CFEM effectors localize to the host plasma membrane and broadly suppress immunity; they can be classified into Asp- or Asn-types based on iron-binding residues, potentially separating iron acquisition from immune suppression roles [21]. Another V. dahliae CFEM effector, VdTRP, relies on membrane targeting for cytotoxicity, which can be neutralized by host apoplastic chitinase-like 1 [22]. Even in obligate biotrophic rust fungi, CFEM-containing candidates have been identified, such as in Puccinia triticina [23], and the stripe rust effector PstCFEM1 suppresses immunity and is required for full virulence [24]. Beyond these systems, CFEM proteins have been implicated in virulence and host immunity modulation across a broad range of other pathogens, including species of Colletotrichum, Fusarium, Alternaria, Neofusicoccum, Marssonina, and Neostagonosporella, highlighting their conserved yet adaptable role in fungal pathogenicity [25,26,27,28,29,30,31]. In particular, CFEM-containing effectors from various phytopathogens have been demonstrated to actively manipulate host programmed cell death (PCD), either eliciting or suppressing it, to facilitate infection [21,26,27].
The three-dimensional structure of CFEM proteins has been pivotal in understanding their functional mechanisms. The first solved CFEM domain, from C. albicans Csa2, adopts a unique helical-basket fold stabilized by four disulfide bonds from its eight characteristic cysteines. Heme is bound within this structure via a specific aspartate residue that coordinates ferric (Fe^3+^) iron, a feature likely critical for its acquisition role [32]. CFEM proteins often function as effectors, secreted to manipulate host immunity. They typically carry conserved motifs, such as CFEM, LysM, or RXLR, that facilitate their function or delivery [33]. These effectors can target key immune components: CfEC12 from Colletotrichum fructicola competitively binds apple MdNIMIN2 to disrupt salicylic acid signaling [34]; a group of F. graminearum CFEM proteins interacts with the maize receptor ZmWAK17 to suppress immune-triggered cell death [35]; and MaCFEM85 from the insect pathogen Metarhizium anisopliae activates jasmonic acid defenses by interacting with alfalfa MsWAK16 [36]. Evolutionarily, the CFEM domain has expanded in pathogenic fungi through gene duplication and is often under positive selection, reflecting adaptation to hosts [2,37]. In the F. solani species complex, CFEM genes are notably expanded and upregulated during infection, underscoring their role in cross-kingdom pathogenicity [38].
S. sclerotiorum is a devastating necrotrophic fungal pathogen with an exceptionally broad host range, causing stem rot or white mold on hundreds of dicotyledonous plants, resulting in significant economic losses worldwide. As a model necrotroph, it rapidly kills host tissue via a combination of lytic enzymes, oxalic acid, and effector proteins, facilitating nutrient acquisition from dead cells. Comparative secretome analyses between S. sclerotiorum and the related necrotroph B. cinerea have revealed important insights into their infection strategies. Notably, a key distinction was observed: while the predicted secretome of B. cinerea lacks proteins containing CFEM or LysM domains, that of S. sclerotiorum includes CFEM-containing proteins [15]. This finding suggests that CFEM proteins may represent a unique component of S. sclerotiorum’s virulence arsenal, potentially playing roles absent in B. cinerea. Despite the identification of CFEM candidates in its secretome and their implied importance, a comprehensive genome-wide identification and characterization of the CFEM protein family in S. sclerotiorum is entirely lacking.
To bridge this knowledge gap, this study aims to perform the first systematic and functionally oriented analysis of the CFEM protein family in S. sclerotiorum. We conducted a genome-wide identification of all SsCFEM genes, followed by a comprehensive in silico characterization of their sequence and structural features, phylogenetic relationships, and expression patterns during key life cycle stages. Importantly, to move beyond prediction and assign potential biological roles, we experimentally characterized the function of the candidate effectors using a heterologous plant expression system, specifically testing their ability to modulate host cell death. This integrated approach provides not only a foundational resource but also direct functional insights into the SsCFEM family, pinpointing key candidates involved in pathogenicity and host interaction for future mechanistic studies.
2. Results
2.1. Genome-Wide Identification and General Characteristics of the SsCFEM Protein Family
A comprehensive search of the S. sclerotiorum strain 1980 UF-70 genome, combining BLASTP+ 2.17.0 with the CFEM domain HMM profile and subsequent SMART validation, identified thirteen genes encoding proteins containing a bona fide CFEM domain. These were designated SsCFEM1 to SsCFEM13. Analysis of their primary sequences (Table S1) revealed substantial heterogeneity in key physicochemical properties (Table 1). The proteins varied considerably in size, with amino acid lengths ranging from 119 (SsCFEM10) to 995 (SsCFEM12), corresponding to predicted molecular weights between 12.25 kDa and 91.69 kDa. Their theoretical isoelectric points (pI) spanned a broad spectrum from acidic (pI 3.45 for SsCFEM2) to alkaline (pI 9.42 for SsCFEM12), suggesting diverse electrostatic properties that may influence subcellular localization and interaction partners. Stability predictions classified five proteins (SsCFEM4, SsCFEM5, SsCFEM6, SsCFEM9, and SsCFEM13) as stable (instability index below 40) and eight as unstable. Furthermore, the grand average of hydropathicity (GRAVY) calculations indicated that ten members are predominantly hydrophobic (GRAVY > 0), while SsCFEM2, SsCFEM11, and SsCFEM12 are hydrophilic (GRAVY < 0), hinting at potential differences in solubility and membrane association.
2.2. Analysis of Subcellular Localization and Effector Potential of SsCFEM Proteins
Bioinformatic tools were employed to predict the cellular fate and potential functional roles of the SsCFEM proteins (Table S1). Signal peptide prediction using SignalP-4.1 indicated that ten proteins possess a cleavable N-terminal signal peptide, targeting them to the secretory pathway (Table 2). Transmembrane helix analysis with TMHMM-2.0 revealed that eight of these secreted candidates lacked transmembrane domains and were thus classified as putative secreted proteins. The remaining five proteins were predicted to contain one or more transmembrane helices, suggesting they may be membrane-associated. Among the candidate secreted proteins, six were predicted to contain a glycosylphosphatidylinositol (GPI) anchor modification site via the big-PI Fungal Predictor, which could facilitate their attachment to the plasma membrane or cell wall. Notably, effector prediction using EffectorP 2.0 identified three of the secreted candidates—SsCFEM9, SsCFEM10, and SsCFEM13—as high-confidence fungal effectors, strongly implicating them in host–pathogen interactions.
To experimentally validate these predictions, we examined the subcellular localization of SsCFEM4, SsCFEM8, and SsCFEM9 via GFP fusion in S. sclerotiorum. Fluorescence microscopy revealed that all three proteins localized to the cytoplasm, exhibiting a composite pattern of a fine reticular network alongside numerous punctate structures (Figure 1A). Morphologically, this pattern is suggestive of an association with the endoplasmic reticulum (reticular network) and vesicles/lysosomes/vacuoles (puncta). To further confirm their subcellular localization, we stained the strain with the vacuole-specific dye CMAC and the mitochondrial marker CMXR. All three CFEM proteins were found to co-localize with CMAC, verifying their vacuolar localization (Figure 1B). In contrast, no co-localization was detected with the mitochondrial marker CMXR, suggesting that these proteins do not localize to mitochondria (Figure S1). Notably, the observed intracellular distribution for SsCFEM8 and SsCFEM9 contrasts with their primary bioinformatic prediction as extracellular proteins (Table 2), indicating potential retention within the secretory pathway or additional intracellular functions. For SsCFEM4, the observed pattern partially aligns with its predicted localization to the membrane and endomembrane compartments.
2.3. Conserved Motif Distribution and Cysteine Pattern Within the CFEM Domain
To uncover conserved sequence elements beyond the canonical domain, a motif analysis of the full-length SsCFEM proteins (Table S1) was conducted using the MEME Suite. This identified seven conserved motifs (Motifs 1–7) (Figure 2A). Their distribution across the family revealed distinct patterns (Figure 2B). Motif 1 was present in all members and corresponds to the core region of the CFEM domain. Motif 2 was conserved in all members except SsCFEM12, suggesting it may represent an auxiliary functional element critical for most family members. Specific subgroups of proteins, notably SsCFEM3, SsCFEM4, and SsCFEM6, shared a common set of additional motifs (Motifs 3–7), implying closer functional relatedness or cooperation. A detailed multiple sequence alignment of the isolated CFEM domains highlighted the hallmark cysteine pattern (Figure 3). Eight cysteine residues were strictly conserved at fixed positions in the majority of members, which are known to form disulfide bonds stabilizing the domain structure. However, deviations were observed: three domains—the second CFEM domain of SsCFEM2 (SsCFEM2.2), along with SsCFEM5 and SsCFEM10—lacked one conserved cysteine at the C-terminal region, while SsCFEM12 lacked two. These cysteine deficiencies suggest potential structural and functional diversification within the family.
2.4. Predicted Tertiary Structure and Phylogenetic Relationships
Homology modeling using Phyre2, with the C. albicans Csa2 (PDB: 4Y7S) structure as a template, generated high-confidence (>95%) three-dimensional models for all SsCFEM proteins (Figure 4). All models adopted the characteristic “helix-basket” fold definitive of the CFEM domain. The core structural network, stabilized by four pairs of conserved cysteine-derived disulfide bonds as seen in Csa2, was largely recapitulated in the SsCFEM models. The model for SsCFEM12 showed notably lower sequence coverage (62%) and a less ordered C-terminal region, consistent with its absence of two key cysteines identified in the alignment. The topology of the rooted tree indicates that the CFEM protein family of S. sclerotiorum forms a distinct monophyletic group, which exhibits a sister-group relationship with the CFEM protein PAND_027d6170 from Ustilago maydis (Figure 5A). This suggests an early evolutionary divergence following their speciation from a common ancestor. Within the ingroup, CFEM proteins did not cluster into a single, highly supported clade but were distributed across multiple fine-scale branches. Notably, conserved motif analysis revealed that members harboring multiple motifs (SsCFEM3, SsCFEM4, and SsCFEM6) clustered together into a compact, highly supported subclade (Figure 2B), implying potential functional similarity among these proteins. A broader interspecific phylogeny including CFEMs from other fungi revealed that SsCFEM proteins are polyphyletic, dispersing across different evolutionary clusters rather than forming a single monophyletic group (Figure 5B). A subset of SsCFEMs clustered with strong support (>80% bootstrap) alongside CFEMs from the related necrotroph B. cinerea, indicating a shared ancestral origin potentially linked to pathogenicity. The dispersal of other SsCFEM members suggests that the family has undergone multiple independent duplications and lineage-specific expansion events in S. sclerotiorum.
2.5. Genomic Architecture
Analysis of gene structure using GSDS 2.0 revealed a significant correlation between exon-intron organization and phylogenetic clustering as well as conserved motifs (Figure 6). Notably, several intron-containing members (SsCFEM3, SsCFEM4, and SsCFEM6) formed a cluster in the phylogenetic tree (Figure 5A), constituting a well-supported monophyletic clade. This distinct genomic structural feature provides further support for the evolutionary relationships inferred from the phylogeny.
2.6. Expression Dynamics During In Vitro Growth and Early Host Colonization
Transcriptomic analysis of publicly available RNA-Seq data (GSE159792) (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE159792 (accessed on 9 December 2025)) elucidated the expression patterns of eight detectable SsCFEM genes across different growth and infection contexts (Figure 7). As the gene annotation for the GSE159792 dataset contains only eight of the thirteen SsCFEM genes—excluding SsCFEM2, 3, 9, 10, and 11—our expression analysis was restricted to those eight detectable members. During in vitro mycelial growth, several genes displayed spatial regulation, with significantly higher expression in the actively growing apex compared to the colony center; this pattern was most pronounced for SsCFEM8. During the early colonization stage across six different host plants, the majority of detected genes (5 out of 8) were upregulated in planta compared to in vitro conditions, consistent with a role during infection. SsCFEM8 exhibited the most robust and consistent induction, being upregulated in all tested hosts, with peak expression observed during infection of sugar beet (Beta vulgaris). Conversely, SsCFEM13 was generally downregulated during plant infection. The expression profiles of SsCFEM genes also showed variation depending on the host plant species, indicating that S. sclerotiorum modulates its CFEM effector repertoire in a host-specific manner. The combined expression data highlight SsCFEM8 as a prime candidate for a key virulence factor involved in host penetration and/or early colonization.
2.7. Functional Validation of SsCFEM Proteins as Cell Death Suppressors
To experimentally assess their functional role, we performed transient expression assays in Nicotiana benthamiana. Ten of the thirteen SsCFEM genes were successfully cloned and expressed. Initial screens showed that none induced cell death when expressed alone (Figure 8A). However, in a suppression assay co-expressing each with the pro-apoptotic protein BAX, SsCFEM4 and SsCFEM9 potently inhibited BAX-induced cell death, while SsCFEM5 showed weaker activity (Figure 8B). The remaining seven members had no significant effect. Concurrently, Western blot analysis using an anti-HA antibody was performed to detect the expression of SsCFEM proteins in all experimental groups, thereby further validating the reliability of the cell death induction and suppression assays mediated by SsCFEM from S. sclerotiorum. The results demonstrated that SsCFEM proteins were successfully detected in all treatment groups (Figure 8C), confirming the credibility of the cell death induction and suppression experiments.
To verify the accuracy of the cell death suppression assay, RNA was extracted from N. benthamiana leaves infiltrated with Agrobacterium tumefaciens cultures harboring SsCFEM4, SsCFEM5, SsCFEM9, or the empty vector pCXSN. Following reverse transcription, quantitative real-time PCR (qRT-PCR) was performed to assess the expression of PTI pathway marker genes. The results showed that the transcript levels of NbWRKY2 (GeneID: 107780258), NbEDS1 (GeneID: 107782626), and NbPRF (GeneID: 107782528) were significantly downregulated in leaves expressing SsCFEM4, SsCFEM5, or SsCFEM9 compared to the pCXSN control. Notably, SsCFEM4 and SsCFEM9 induced a more pronounced downregulation of these PTI marker genes than SsCFEM5, a trend consistent with the observed suppression of cell death.
This functional validation directly supports and extends our bioinformatic predictions: SsCFEM9, a predicted effector (Table 2), is confirmed as a cell death suppressor. Interestingly, SsCFEM4, not predicted as an effector, also exhibits this activity.
3. Discussion
This study provides the first genome-wide analysis of the CFEM protein family in the broad-host-range necrotroph S. sclerotiorum. We identified 13 SsCFEM proteins exhibiting substantial diversity in sequence, structure, and predicted subcellular localization. Their complex phylogenetic and expression patterns suggest nuanced regulatory mechanisms and potential adaptation to host infection. Notably, functional screening revealed that specific members, such as SsCFEM4 and SsCFEM9, act as suppressors of plant cell death, highlighting their likely role in modulating host immunity during pathogenesis.
3.1. Diversity and Predicted Functional Roles of SsCFEM Proteins
The identification of 13 CFEM proteins in S. sclerotiorum places it among fungi with a moderate number of such domains, consistent with patterns observed in other phytopathogens where CFEM copy number can correlate with pathogenic lifestyle [2]. The high sequence and physicochemical heterogeneity within the family suggest functional diversification (Table 1). Our prediction that a subset of SsCFEMs are canonical secreted proteins lacking transmembrane domains aligns with the well-established role of CFEM proteins as extracellular effectors in other fungi. For instance, proteins such as Rbt5 and Pga7 in C. albicans function as hemophores involved in nutrient acquisition [4], whereas other CFEM family members, including certain proteins in V. dahliae, have been characterized as effectors that suppress host immunity [21]. The prediction of GPI-anchor modification sites in several secreted SsCFEMs is particularly noteworthy (Table 2). In C. albicans, GPI-anchored CFEM proteins like Rbt5 are integral to cell wall architecture and heme-iron acquisition [6]. Similarly, in B. cinerea, the GPI-anchored CFEM protein BcCFEM1 contributes to virulence [13]. This suggests that GPI-anchoring may be a conserved mechanism for localizing CFEM domains to the fungal cell surface to mediate interactions with the host environment, potentially including nutrient scavenging or adhesion.
The identification of three SsCFEMs (SsCFEM9, 10, and 13) as high-confidence effectors by EffectorP further supports their potential involvement in manipulating host physiology (Table 2). This is reminiscent of CFEM effectors in other plant pathogens, such as PstCFEM1 from wheat stripe rust, which suppresses host immunity [24], and a suite of CFEM effectors in Fusarium species that inhibit plant cell death [26,27]. The presence of proteins with multiple transmembrane helices (e.g., SsCFEM11, 12) defines another subfamily, possibly functioning as membrane-bound receptors or transporters (Table 2). The CFEM-containing GPCR Pth11 in M. oryzae is a key precedent, essential for surface sensing and pathogenicity [16,17]. Although our predictions did not strongly indicate GPCR domains for these transmembrane SsCFEMs, they may represent a divergent class of membrane-integral CFEM proteins with undiscovered signaling or transport functions.
3.2. Structural Insights and Evolutionary Trajectory
The conservation of the core “helical-basket” fold, as predicted by AlphaFold2 and confirmed by high model confidence (Figure 4), underscores the structural unity of the CFEM domain across kingdoms, from human pathogens like C. albicans [32] to plant pathogens like S. sclerotiorum. However, variations in the canonical eight-cysteine pattern, observed in several SsCFEMs (Figure 3), indicate potential structural plasticity. Such variations have been noted in CFEM families of other fungi, like F. graminearum [41], and may lead to altered disulfide bonding, affecting ligand binding specificity or protein stability. The markedly lower structural coverage for SsCFEM12, which lacks two cysteines, strongly suggests a distinct structural conformation, possibly reflecting a specialized function (Figure 4). Phylogenetic analysis provides crucial insights into the evolutionary history of this family. The topology of the rooted tree reveals that the CFEM protein family in S. sclerotiorum does not exhibit deep phylogenetic divergence, but instead forms multiple shallow, poorly supported branches radiating from a common ancestral node (Figure 5A). This pattern of limited diversification suggests that the family may have undergone strong functional constraints during subsequent evolution, leading to conserved sequence and functional characteristics. The non-monophyletic distribution of SsCFEMs in the interspecific tree suggests a complex evolutionary history (Figure 5B). The clustering of some SsCFEMs with CFEMs from the necrotroph B. cinerea with high support implies shared ancestry and possibly conserved functions related to necrotrophy, such as host cell death manipulation or detoxification of plant defenses. The scattering of other SsCFEMs across the tree likely results from lineage-specific gene duplication and expansion, a driving force in the evolution of pathogen effector repertoires [37]. This pattern of both conservation and lineage-specific innovation is common in fungal pathogenicity factor families and allows for adaptation to specific hosts or niches.
3.3. Expression Dynamics Suggest Context-Specific Functions
The expression profiling during plant infection offers the most direct functional clues (Figure 7). The widespread upregulation of SsCFEM genes in planta, especially the strong and consistent induction of SsCFEM8 across all hosts, strongly implicates these genes in virulence. This pattern mirrors the induction of CFEM effector genes during infection in other pathosystems, such as FsCFEMs in F. sacchari [26] and PstCFEM1 in wheat stripe rust [24]. The specific high expression of SsCFEM8 in the mycelial apex, both in vitro and in planta, points to a role in hyphal tip growth, host surface sensing, or initial penetration—functions analogous to the CFEM-GPCR WISH in M. oryzae [17]. Conversely, the downregulation of SsCFEM13 during infection suggests its primary role may be in saprophytic growth or development, highlighting functional specialization within the family. The host-specific variation in expression profiles further suggests that S. sclerotiorum employs a flexible strategy, modulating its CFEM arsenal to optimize interaction with different plant species.
3.4. Experimental Validation of Cell Death Suppression Activity
Our transient expression assays provide the first experimental evidence for the function of SsCFEM proteins as modulators of plant cell death, a cornerstone of plant defense and necrotrophic pathogenesis (Figure 8). The potent suppression of BAX-induced PCD by SsCFEM4 and SsCFEM9 aligns with the emerging paradigm of necrotrophic effectors that actively inhibit host immunity to facilitate colonization [39]. This finding is particularly significant for SsCFEM9, as it confirms its bioinformatic classification as a high-confidence effector (Table 2). The activity of SsCFEM4, which was not predicted as an effector by computational tools, underscores the complementary value of experimental screening in identifying functional candidates that may be missed by current algorithms.
The functional divergence observed—where only a subset of tested SsCFEMs showed suppressive activity—parallels their sequence, structural, and phylogenetic diversity (Figure 2, Figure 3 and Figure 5). It is plausible that cell death suppression is a specialized function acquired by specific members within Clades I and II, possibly through distinct molecular mechanisms. The failure to clone SsCFEM3, 10, and 12 under standard laboratory growth conditions is intriguing. Their functional characterization remains an important goal for future work.
The lack of cell death-inducing activity for any SsCFEM is noteworthy (Figure 8A). Unlike some necrotrophic effectors that directly elicit necrosis, these SsCFEM suppressors may function early during infection to dampen pattern-triggered immunity (PTI), creating a permissive environment for the fungus before the onset of widespread tissue necrosis driven by lytic enzymes and oxalic acid. This “stealth” strategy would be consistent with the early upregulation of several SsCFEM genes, including SsCFEM8, during host colonization (Figure 7).
3.5. Limitations and Future Perspectives
This study provides a robust predictive framework and initial functional validation for the SsCFEM family. Key limitations include reliance on a single reference strain, which may not capture the full allelic diversity of SsCFEMs across populations, and the use of transcriptomic data from early infection stages, potentially missing roles during later necrotrophic growth or sclerotial development. Furthermore, our functional assays are based on heterologous expression in a model plant. While informative, confirming these activities during actual infection by S. sclerotiorum through gene knockout/complementation is essential.
Future research should prioritize elucidating the molecular mechanisms by which SsCFEM4 and SsCFEM9 suppress cell death (Figure 8B) and determining the in planta role of the high-expression candidate SsCFEM8 (Figure 7). Gene knockout and complementation studies in S. sclerotiorum are essential to confirm their roles in virulence, host range, and specific pathogenic processes like oxalate production or tissue maceration. For transmembrane members like SsCFEM11/12, investigating their role in sensing or transport will be crucial (Table 2). An interesting direction for future research is to explore whether SsCFEM proteins, beyond their immune-modulating functions, may also participate in iron acquisition. In C. albicans, the CFEM proteins Rbt5 and Pga7 bind heme via a conserved aspartic acid (Asp) residue essential for iron scavenging [4]. Recent work in V. dahliae has shown that mutation of this residue can drive functional divergence from iron acquisition toward immune suppression [21]. Whether SsCFEM proteins possess this conserved residue and retain heme-binding capability remains to be determined. Biochemical studies are needed to determine if SsCFEMs bind heme or other ligands, as seen in Candida [3,6], which would link them to iron acquisition—a critical virulence factor even for necrotrophs. Furthermore, identifying host targets of the predicted effector SsCFEMs, following examples like the interaction between F. graminearum CFEMs and maize ZmWAK17 [35] or C. fructicola CfEC12 and apple MdNIMIN2 [34], will unravel the molecular mechanisms of immune suppression. For the uncloned members (SsCFEM3, 10, and 12), alternative strategies such as using induced RNA-seq guided cDNA synthesis from infected tissue should be employed to capture their transcripts and elucidate their functions.
4. Materials and Methods
4.1. Bacterial Strains, Plant Materials, Vectors, and Primers
The fungal strain S. sclerotiorum 1980 was used in this study. N. benthamiana plants were employed as the plant material. The vectors used included the plant expression vector pCXSN and the pNAH-ONG vector, which carries the Olic promoter and a green fluorescent protein (GFP) tag. All primers used are listed in Supplementary Table S6.
4.2. Genome-Wide Identification and Physicochemical Characterization
The genome sequence and annotation of S. sclerotiorum strain 1980 UF-70 were used as the basis for this study. CFEM domain-containing proteins were initially identified by performing a local BLASTP search against the S. sclerotiorum proteome using the hidden Markov model (HMM) profile of the CFEM domain (Pfam: PF05730) as the query, with an E-value cutoff of 1 × 10^−5^. All candidate protein sequences were subsequently validated for domain integrity using the SMART online database (https://smart.embl.de/smart/change_mode.cgi) (accessed on 15 November 2025). Proteins confirmed to contain a bona fide CFEM domain were designated SsCFEM1 to SsCFEM13. Their basic physicochemical properties, including amino acid number, molecular weight, theoretical isoelectric point (pI), instability index, and grand average of hydropathicity (GRAVY), were computed using the ExPASy ProtParam tool (https://web.expasy.org/protparam/) (accessed on 15 November 2025).
4.3. Sequence Alignment, Conserved Motif, and Gene Structure Analysis
Multiple sequence alignment of the CFEM domains extracted from the 13 SsCFEM proteins was performed using the ClustalW algorithm implemented in MEGA12 software (Table S2). For the discovery of conserved motifs across the full-length protein sequences, the MEME Suite (https://meme-suite.org/meme/doc/download.html) (accessed on 15 November 2025) was employed with the following parameters: maximum number of motifs to find = 10, minimum motif width = 6, and maximum motif width = 50 amino acids.
The exon–intron structures of the SsCFEM genes were illustrated using the Gene Structure Display Server 2.0 (https://gsds.gao-lab.org/Gsds_help.php) (accessed on 15 November 2025). Genomic DNA sequences and their corresponding coding sequences (CDSs) were obtained from the genome annotation file to generate schematic diagrams depicting the number, length, and arrangement of exons (represented as yellow rectangles) and introns (represented as black lines) (Tables S4 and S5). Since the coding sequences (CDSs) of SsCFEM11 to SsCFEM13 were only available in the EMBL (https://www.ebi.ac.uk/) (accessed on 15 November 2025) database, and their non-coding regions could not be retrieved, the gene structure analysis in this study was limited to SsCFEM1 to SsCFEM10.
4.4. Prediction of Subcellular Localization and Effector Potential
The presence and cleavage site of N-terminal signal peptides were predicted using SignalP-4.1 (https://services.healthtech.dtu.dk/services/SignalP-4.1/) (accessed on 15 November 2025) with default settings for eukaryotes. Transmembrane helices were predicted using TMHMM-2.0 (https://services.healthtech.dtu.dk/services/TMHMM-2.0/) (accessed on 15 November 2025). Mitochondrial targeting peptides were predicted using TargetP 2.0 (https://services.healthtech.dtu.dk/services/TargetP-2.0/) (accessed on 15 November 2025). Potential glycosylphosphatidylinositol (GPI) anchor modification sites were predicted using the big-PI Fungal Predictor (https://mendel.imp.ac.at/gpi/fungi_server.html) (accessed on 15 November 2025). The potential of SsCFEM proteins to function as fungal effectors was assessed using EffectorP 3.0 (https://effectorp.csiro.au/) (accessed on 15 November 2025).
4.5. Phylogenetic Analysis
Two phylogenetic trees were constructed using the maximum likelihood (ML) method in MEGA12 software. First, a phylogenetic tree was constructed using the full-length sequences of the 13 SsCFEM proteins (Table S1), with the CFEM protein PAND_027d6170 from U. maydis as the outgroup (Table S3). Second, an interspecific tree was constructed by aligning the SsCFEM sequences with previously characterized CFEM protein sequences from representative fungi (M. oryzae, B. cinerea, F. graminearum, and C. albicans), which were retrieved from the NCBI database. For the phylogenetic tree of multiple species, we selected the UMAG_02820 protein from U. maydis as the outgroup. For both analyses, the best-fit amino acid substitution model (JTT+G for the intraspecific tree; WAG+G for the interspecific tree) was selected based on the lowest Bayesian Information Criterion (BIC) score. The robustness of tree topology was assessed by bootstrap analysis with 1000 replicates.
4.6. Three-Dimensional Structure Modeling
The three-dimensional (3D) structures of the SsCFEM domains were predicted using the protein homology modeling server Phyre2 (https://www.sbg.bio.ic.ac.uk/~phyre2/html/page.cgi?id=index) (accessed on 15 November 2025) in the intensive modeling mode. The crystal structure of the C. albicans Csa2 CFEM domain (PDB: 4Y7S) was used as the primary template. Models with confidence scores > 95% were selected for visualization. The final models were rendered to highlight secondary structure elements (α-helices as green cylinders, β-strands as blue arrows) and amino acid properties colored by type (small/polar: yellow; hydrophobic: green; charged: red; aromatic and cysteine: purple).
4.7. In Silico Expression Profiling
Publicly available RNA-Seq data under NCBI accession GSE159792 [42] were utilized to analyze SsCFEM gene expression. This dataset includes transcriptomes from S. sclerotiorum during in vitro mycelial growth (colony center and apex) and during early infection stages on six different host plants (Arabidopsis thaliana, Beta vulgaris, Ricinus communis, Phaseolus vulgaris, Helianthus annuus, and Solanum lycopersicum). Fragments Per Kilobase of transcript per Million mapped reads (FPKM) values for the SsCFEM genes present in the dataset were extracted. A hierarchical clustered heatmap was generated using TBtools-II to visualize expression dynamics across the different biological conditions.
4.8. Transient Expression Assays in N. benthamiana
The coding sequences of SsCFEM genes were cloned into the plant expression vector pCXSN. which provides an N-terminal HA tag under the control of the CaMV 35S promoter. The constructs were transformed into A. tumefaciens strain GV3101. Bacterial suspensions were prepared in infiltration buffer (10 mM MES, 20 g/L sucrose, 200 μM acetosyringone) and adjusted to OD600 = 1.0 for leaf infiltration in 4–5-week-old N. benthamiana plants.
To screen for cell death-inducing activity, leaves were infiltrated with A. tumefaciens suspensions carrying individual SsCFEM constructs. The empty vector served as a negative control, and BAX alone served as the positive control for cell death induction. For cell death suppression assays, leaves were first infiltrated with A. tumefaciens carrying an SsCFEM construct. After 24 h, an adjacent site was infiltrated with A. tumefaciens carrying BAX to create an overlapping zone, which was then assessed for suppression of BAX-induced cell death. Controls for this assay included the empty vector (negative control) and the co-infiltration of BAX with the known suppressor AVR3a (positive control for suppression). All infiltrated plants were kept under low-light conditions for 24 h and then transferred to normal light. Cell death symptoms were visually monitored daily for up to 5 days post-infiltration. All experiments included at least three biological replicates and were repeated twice independently. Total proteins were extracted from the infiltrated zones of tobacco leaves and analyzed by Western blot with an anti-HA antibody to determine the expression of HA-tagged SsCFEM proteins.
4.9. Analysis of PTI Marker Gene Expression
A. tumefaciens strains harboring the SsCFEM-pCXSN recombinant plasmid or the pCXSN empty vector were resuspended in infiltration buffer to an optical density and adjusted to OD600 = 1.0. The bacterial suspensions were then infiltrated into the abaxial side of the leaves of N. benthamiana. The empty vector strain served as a negative control. At least three biological replicates were performed for each treatment.
Leaf tissue samples from the infiltrated zones were collected at 24 h post-infiltration. Total RNA was extracted using a plant RNA extraction kit, and its concentration was determined. After DNase I digestion to remove genomic DNA contamination, 1 μg of total RNA was reverse-transcribed into first-strand cDNA using a reverse transcription kit.
Quantitative real-time PCR (qRT-PCR) was performed using SYBR Green Premix on a real-time PCR detection system, with the cDNA as template. The expression levels of the target PTI marker genes (PRF, EDS1, and WRKY2) were examined, and a tobacco housekeeping gene was used as an internal control. Three technical replicates were analyzed for each sample. Relative gene expression levels were calculated using the 2^−ΔΔCt^ method to compare expression differences between the treatment and control groups. All the primers used in this study are listed in Table S6. The reverse transcription kit (Cat. No. R333-C1, Version 24.1) and SYBR Green qPCR mix (Cat. No. Q312, Version 23.1) were purchased from Nanjing Vazymc, Nanjing, China
4.10. Subcellular Localization Analysis
To determine the subcellular localization of SsCFEM proteins, GFP-fusion constructs were expressed in S. sclerotiorum while retaining their native signal peptides. The coding sequence (CDS) of each SsCFEM gene was amplified from wild-type strain 1980 cDNA and cloned into the SpeI-digested pNAH-ONG vector using a seamless cloning kit, generating a C-terminal GFP fusion plasmid with a G418 resistance marker. After sequence verification, the recombinant plasmid was used as a template to amplify a linearized fragment with primers pmtF/pmtR. This linear DNA was then introduced into S. sclerotiorum protoplasts via PEG-mediated transformation. Following transformation, the mixture was incubated in antibiotic-free RM medium for 12–16 h at 22 °C, after which transformants were selected by overlaying with SRM medium containing 100 µg/mL G418. Positive transformants were subcultured 3–4 times on G418-containing PDA to ensure genetic stability.
For microscopy, mycelial plugs from stable strains were examined using a laser scanning confocal microscope. Fluorescence distribution was recorded from at least three independent fields per strain. The GFP signals were observed using 488 nm light. The vacuoles were stained with 10 mM CellTracker™ Blue CMAC (7-amino-4-chloromethylcoumarin, Invitrogen) for 45 min at room temperature in the dark and observed with the DAPI channel using 405 nm light. The mitochondria were stained with 1 mM MitoTracker™ Red CMXR (Chloromethyl-X-rosamine, Invitrogen, Source:Thermo Fisher Scientific Eugene USA) for 45 min at room temperature in the dark and observed with the RFP channel using 561 nm light.
5. Conclusions
In summary, our integrated study deciphers the CFEM protein repertoire of S. sclerotiorum and provides direct functional insights into its role in pathogenesis. The genome-wide analysis revealed a diverse family with features indicative of effector potential, including secretion signals, GPI anchors, and specific regulatory motifs. Crucially, transient expression assays validated the immunosuppressive function of key members, identifying SsCFEM4 and SsCFEM9 as potent suppressors of host cell death. This experimental confirmation establishes SsCFEM9 as a functional effector candidate and uncovers a previously unannotated function for SsCFEM4. Together with the distinct phylogenetic clades and infection-induced expression patterns, our findings portray the SsCFEM family as a dynamically regulated toolkit evolved for host manipulation. This work not only provides a foundational resource but also delivers prioritized, functionally characterized candidates for future mechanistic studies, advancing the quest for targeted control strategies against this devastating pathogen.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kulkarni R.D. Kelkar H.S. Dean R.A. An eight-cysteine-containing CFEM domain unique to a group of fungal membrane proteins Trends Biochem. Sci.20032811812110.1016/S 0968-0004(03)00025-212633989 · doi ↗ · pubmed ↗
- 2Zhang Z.N. Wu Q.Y. Zhang G.Z. Zhu Y.Y. Murphy R.W. Liu Z. Zou C.G. Systematic analyses reveal uniqueness and origin of the CFEM domain in fungi Sci. Rep.201551303210.1038/srep 1303226255557 PMC 4530338 · doi ↗ · pubmed ↗
- 3Weissman Z. Kornitzer D. A family of Candida cell surface haem-binding proteins involved in hemin and haemoglobin-iron utilization Mol. Microbiol.2004531209122010.1111/j.1365-2958.2004.04199.x 15306022 · doi ↗ · pubmed ↗
- 4Kuznets G. Vigonsky E. Weissman Z. Lalli D. Gildor T. Kauffman S.J. Turano P. Becker J. Lewinson O. Kornitzer D. A relay network of extracellular heme-binding proteins drives Candida albicans iron acquisition from hemoglobin P Lo S Pathog.201410 e 100440710.1371/journal.ppat.100440725275454 PMC 4183699 · doi ↗ · pubmed ↗
- 5Okamoto-Shibayama K. Kikuchi Y. Kokubu E. Sato Y. Ishihara K. Csa 2, a member of the Rbt 5 protein family, is involved in the utilization of iron from human hemoglobin during Candida albicans hyphal growth FEMS Yeast Res.20141467467710.1111/1567-1364.1216024796871 · doi ↗ · pubmed ↗
- 6Sorgo A.G. Brul S. de Koster C.G. de Koning L.J. Klis F.M. Iron restriction-induced adaptations in the wall proteome of Candida albicans Microbiology 20131591673168210.1099/mic.0.065599-023728625 · doi ↗ · pubmed ↗
- 7Roy U. Yaish S. Weissman Z. Pinsky M. Dey S. Horev G. Kornitzer D. Ferric reductase-related proteins mediate fungal heme acquisitione Life 202211 e 8060410.7554/e Life.8060436200752 PMC 9635878 · doi ↗ · pubmed ↗
- 8Pérez A. Pedrós B. Murgui A. Casanova M. López-Ribot J.L. Martínez J.P. Biofilm formation by Candida albicans mutants for genes coding fungal proteins exhibiting the eight-cysteine-containing CFEM domain FEMS Yeast Res.200661074108410.1111/j.1567-1364.2006.00131.x 17042757 · doi ↗ · pubmed ↗