Preparation and Immunogenicity Prediction of Brucella melitensis mRNA Vaccine Candidate Based on omp16 and omp19 Genes
Jingjie Zhang, Haiyan Borijihan, Yixuan Chen, Huricha Baigude, Lili Bao, Fu Quan, Dezhi Yang

TL;DR
This study explores the development of an mRNA vaccine candidate for brucellosis using the Brucella melitensis omp16 and omp19 genes, showing promising immunogenic properties.
Contribution
The novel contribution is the synthesis and evaluation of mRNA vaccines based on omp16 and omp19 genes for brucellosis.
Findings
Recombinant plasmids pIVTRup-omp16 and pIVTRup-omp19 were successfully constructed and used to synthesize mRNA.
Omp16 showed stronger immunogenicity than Omp19 and could activate both CD4+ and CD8+ T cells.
Epitope prediction identified seven and six epitopes for Omp16 and Omp19, respectively.
Abstract
Background: Brucella outer membrane proteins (Omps) are an important part of its cell wall and major virulence-related factors. Omp16 and Omp19 proteins are the advantageous antigens of Brucella and have been widely used in research on vaccines against brucellosis. As an emerging vaccine, the mRNA vaccine has unique advantages in the fight against intracellular parasitic bacteria. Methods: In this study, mRNA encoding the omp16 and omp19 genes of Brucella. melitensis (B. melitensis) was synthesized using in vitro transcription. The target mRNA was transfected into HEK 293T cells to evaluate protein expression levels and assess its immunogenicity. Finally, bioinformatic approaches were employed to analyze potential antigenic epitopes. Results: In this study, the successfully constructed recombinant plasmids pIVTRup-omp16 and pIVTRup-omp19 were utilized to synthesize omp16-mRNA and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
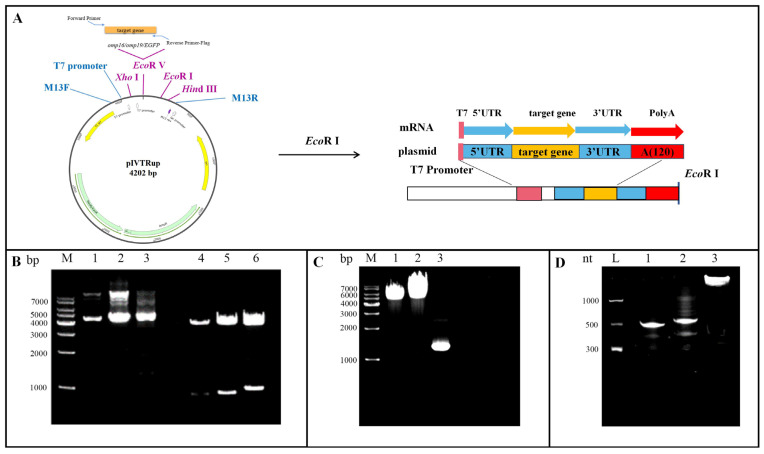 Figure 1
Figure 1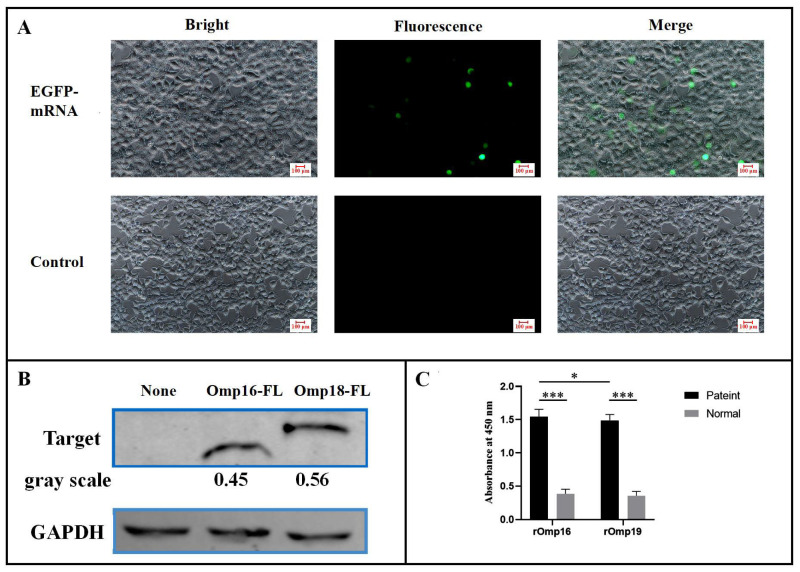 Figure 2
Figure 2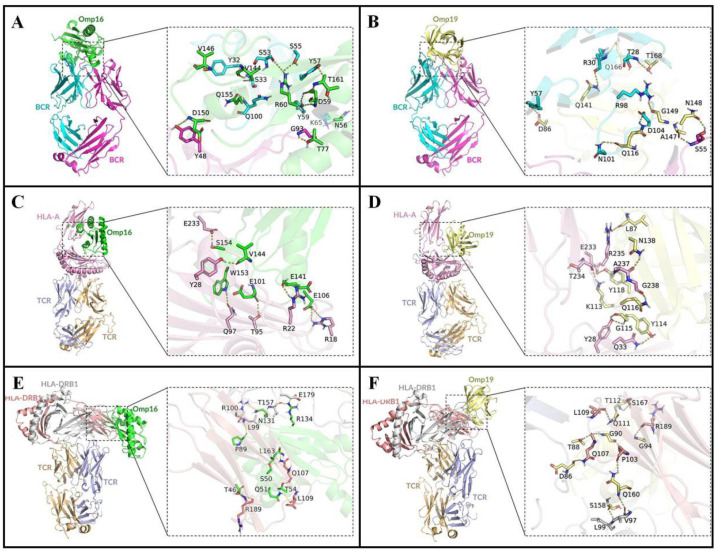 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5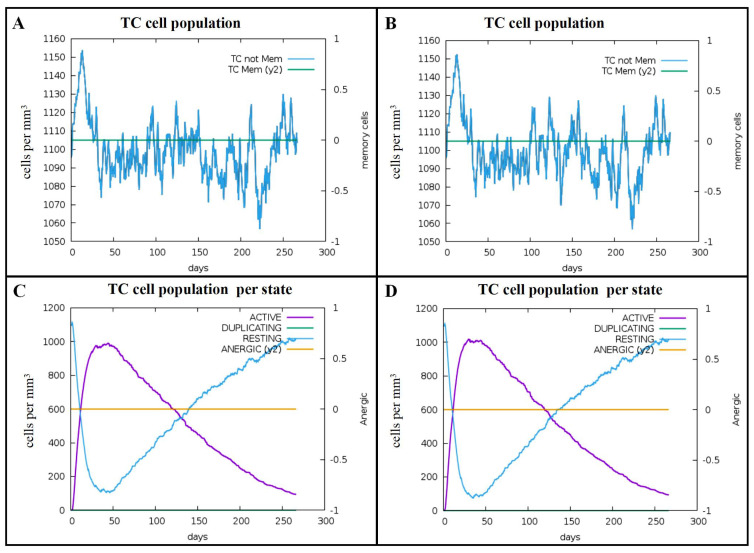 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8- —Natural Science Foundation of Inner Mongolia Autonomous Region
- —High-Level Clinical Specialty Development Science and Technology Project for Public Hospitals in the Capital Region
- —General Program of Inner Mongolia Medical University
- —Science and Technology Program of the Joint Fund of Scientific Research for the Public Hospitals of Inner Mongolia Academy of Medical Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBrucella: diagnosis, epidemiology, treatment · vaccines and immunoinformatics approaches · Toxoplasma gondii Research Studies
1. Introduction
Brucella is an intracellular parasitic, globular, Gram-negative bacterium that infects ruminants and humans, causing brucellosis [1]. Recent studies show that there are 16,000 to 2.1 million new infections each year, which has a huge impact on population health and animal husbandry development [2]. Brucella melitensis (B. melitensis) is the most common and most pathogenic species of Brucella [3]. Goats and sheep are considered the natural hosts for B. melitensis, and infection often leads to impaired fertility [4]. Humans typically become infected through contact with infected animals or consumption of contaminated animal products, resulting in a chronic debilitating febrile illness [5,6]. The incubation period for human brucellosis generally ranges from 5 to 60 days, with common clinical manifestations including fever, profuse sweating, fatigue, arthralgia, and generalized myalgia [7]. Vaccination remains the primary strategy for preventing and controlling its transmission. Attenuated live vaccines, such as S19, RB51, SR82, and Rev1, are the most common types developed for livestock [8]. This highlights an urgent need for novel, safe, and effective vaccine strategies.
With the advancement of precise molecular technologies and an improved understanding of brucellosis pathogenesis, novel genetically engineered vaccines have been developed to replace conventional ones for the prevention and control of brucellosis [9,10]. In recent years, nucleotide-modified messenger RNA (mRNA) vaccines have emerged as a leading platform with great potential against a wide range of infectious pathogens [11]. Unlike DNA vaccines, mRNA is translated directly in the host cytoplasm without the need for nuclear entry or genomic integration, and is eventually degraded, thereby reducing the risk of genetic dysregulation [12].
The selection of an appropriate antigen is paramount for vaccine efficacy. The outer membrane proteins (Omps) family maintains bacterial integrity while eliciting a strong immune response and is thought to be involved in Brucella survival and immune escape in macrophages [5,13]. Among these, Omp16 and Omp19 lipoproteins are highly conserved across major pathogenic Brucella species and are considered advantageous antigens for brucellosis vaccine development [14,15]. Omp16 has been shown to induce strong Th1-biased immune responses, while Omp19 plays a vital role in mucosal immunity by interacting with Toll-like receptor 2 (TLR2) [16,17,18,19]. Therefore, Omp16 and Omp19 represent ideal targets for preparing brucellosis vaccines.
This study aims to utilize the key antigens Omp16 and Omp19 from B. melitensis to construct an in vitro transcription vector for generating antigen-encoding mRNA. We will investigate their protein expression in eukaryotic cells and assess the immunogenicity of the expressed proteins. Furthermore, bioinformatic approaches will be employed to predict and analyze B-cell and T-cell epitopes of B. melitensis Omp16 and Omp19 proteins. Molecular docking with corresponding receptors and immune challenge simulations will be conducted to preliminarily evaluate the feasibility of an mRNA-based vaccine against Brucella, thereby providing a theoretical foundation for the development of novel brucellosis vaccines.
2. Materials and Methods
2.1. Plasmids, Strains and Cells
pIVTRup plasmid was purchased from Addgene Company (Cambridge, MA, USA). pCDNA3-EGFP plasmid was preserved by Innovation Mongolian Medicine Engineering Research Center of Inner Mongolia International Mongolian Medical Hospital (Hohhot, China). Competent cells E. coli TransT1 were purchased from Beijing Transgen Biotech (Beijing, China). Human embryonic kidney 293T cells (HEK 293T) (ATCC: CRL-3216) were cultured in DMEM medium containing 10% fetal bovine serum (Gibco, Waltham, MA, USA), 100× streptomycin mixture (Solarbio, Beijing, China) and cultured in a constant temperature incubator containing 5% CO_2_ at 37 °C.
2.2. Liposomes, Brucella Genome and Serum
Liposome Dogo 4 was provided by Professor Huricha Baigude, School of Chemical Engineering, Inner Mongolia University [20]. B. melitensis 6144 genome (Genbank: CP098767.1) and brucellosis patients (10 cases) and healthy human serum (10 cases) were from Clinical Laboratory Center, Affiliated Hospital of Inner Mongolia Medical University. Positive serum inclusion criteria: ① clinical diagnosis of brucellosis; ② Brucella serum antibody test positive, antibody titer >1:50; ③ fever, fatigue and lumbago and other obvious clinical symptoms. Negative serum was collected from the healthy check-up population, with the criterion: negative for Brucella antibody testing.
2.3. Construction of Plasmid pIVTRup-omp16/omp19/EGFP
Search Genbank for B. melitensis omp16 and omp19 gene sequences (omp16 GeneID: 29593101; omp19 GeneID: 29594621), and the EGFP gene sequence on pCDNA3-EGFP plasmid. Primer 5.0 software was used to design primers containing Flag tag sequences (Table 1). The omp16, omp19 and EGFP gene fragments purified by agarose gel were cloned into the in vitro transcription vector pIVTRup to obtain pIVTRup-omp16, pIVTRup-omp19 and pIVTRup-EGFP.
2.4. Synthesis of Target mRNA
2.4.1. DNA Template
pIVTRup-omp16 and pIVTRup-omp19 were digested with EcoR I (NEB, Ipswich, MA, USA) and linearized respectively. pIVTRup-EGFP was amplified by PCR to obtain DNA template of suitable fragment. EGFP-F and M13R were used as upstream and downstream primers to prepare amplification reaction system with volume of 50 μL. The reaction conditions: (95 °C, 30 s; 55 °C, 30 s; 72 °C, 1 min, 20 s; 35 cycles in total); 72 °C, 5 min; 4 °C. DNA template for in vitro transcription was purified as described above using Agarose Gel DNA Extraction Kit (TaKaRa Bio, Kusatsu, Japan).
2.4.2. In Vitro Transcription
HiScribe^®^ T7 ARCA mRNA in vitro transcription kit (NEB, Ipswich, MA, USA) was used to prepare transcription system in EP tube according to Table 2. 37 °C water baths for 30 min, 1 µL DNase I was added, and after mixing, 37 °C water bath for 15 min to remove DNA template. Target mRNA was purified with LiCl and ethanol according to kit instructions and stored at 80 °C.
2.4.3. Target mRNA Identification
A 5% denaturing PAGE gel was prepared and pre-electrophoresis was performed in 1 × TBE electrophoresis buffer at 200 V for 30 min. A 2 µL RNA sample and a 2 µL RNA Ladder were mixed with 2 µL loading buffer respectively and placed into PCR instrument at 95 °C for 5 min, then placed on ice rapidly for 5 min to prevent RNA renaturation. RNA sample was added into gel well and electrophoresis was performed at 80 V for 25 min until bromophenol blue reached the bottom of gel. The gel block was soaked in nucleic acid dye for 1 h and the results were observed under an ultraviolet lamp.
2.5. HEK 293T Cell Transfection
HEK 293T cells were inoculated into 10 mm cell culture dishes and cultured at 37 °C for 16 h in 5% CO_2_ until the cell fusion reached 80%. The cells were replaced with DMEM basal medium. 20 µL Dogo 4 liposome and 500 µL Opti-MEM (Gibco, Waltham, MA, USA) were added to each enzyme-free EP tube, mixed and allowed to stand at room temperature for 5 min to prepare solution A. In new enzyme-free EP tubes, 20 µL ddH_2_O, 20 μg omp 16-mRNA, omp19-mRNA and EGFP-mRNA were mixed with 500 µL Opti-MEM serum reduced medium to prepare solution B. Solution B was added to solution A and incubated at room temperature for 5 min. Solution AB was added slowly to the cells and cultured at 37 °C for 5 h in 5% CO_2_, then replaced with DMEM complete medium containing 10% fetal bovine serum and 1% streptomycin for 19 h. Green fluorescence of EGFP-mRNA group was observed under an fluorescence microscope 24 h after transfection.
2.6. Extraction and Purification of Protein
Twenty-four hours after transfection, omp16-mRNA, omp19-mRNA and EGFP-mRNA groups were collected into EP tubes with 1 × PBS, and the supernatant was discarded. 700 μL RIPA lysate (Beyotime, Shanghai, China) containing 1 mM PMSF (Beyotime, Shanghai, China) was added to each tube to lyse the cells. Purify the protein of interest using Pierce™ Anti-DYKDDKAffinity Resin (Thermo Fisher Scientific, Waltham, MA, USA), collect the first wash solution (containing the internal reference protein) into EP tubes, store the final eluate (containing the protein of interest), and freeze at 80 °C.
2.7. Western Blot Analysis of Protein Expression
A total amount of 30 µL of purified protein samples was mixed with 6 µL of 6× Protein Buffer, heated at 95 °C for 5 min, and placed on ice. Western blotting was performed according to the instructions of SDS-PAGE gel preparation kit (CWBio, Taizhou, China). Flag-Tag Mouse Monoclonal Antibody (Absin, Shanghai, China) was used to detect Omp16 and Omp19 proteins expressed by transfected cells. GAPDH Mouse Monoclonal Antibody (Absin, Shanghai, China) was used to detect internal reference proteins. FITC-conjugated Goat Anti-Mouse IgG (H + L) was used as secondary antibody. The results were observed with a two-color infrared laser imager (Odyssey CLX, 700 nm and 800 nm).
2.8. ELISA to Detect Protein Immunogenicity
To detect protein immunogenicity, 0.5 μg purified rOmp16 and rOmp19 protein coated 96 well plate respectively. PBST washed 3 times, each well added 200 µL PBST preparation of 1% skim milk powder blocked at room temperature for 1 h. PBST washed 3 times, each well added 100 µL diluted brucellosis patient serum, control group added healthy human serum, incubated at 37 °C for 1.5 h. PBST wash buffer 4 times, add 100 µL HRP-labeled rabbit anti-human IgG (1:5000) to each well, incubate at 37 °C for 1 h. PBST wash buffer 5 times. Soluble TMB kit (CWBio, Taizhou, China), adding 100 µL of stop solution per well (1 mol/L HCL), and after 15 min, the OD value at 450 nm was read using a microplate reader. The results of rOmp16 and rOmp19 reactions with serum were expressed as mean ± standard deviation of OD_450_. The experiment was repeated three times. The experimental data were plotted using GraphPad Prism 9.5 software. Statistical differences in serum immune response between patients and healthy subjects were analyzed by independent sample t-test, and statistical differences in immunogenicity between rOmp16 and rOmp19 were analyzed by paired sample t-test. p < 0.05 was considered a statistical difference.
2.9. Prediction of Antigenic Epitopes of Omp16 and Omp19 Proteins
2.9.1. Prediction of B Cell Epitopes for Omp16 and Omp19 Proteins
Comprehensive prediction of B-cell epitopes for Omp16 and Omp19 proteins was performed using the online prediction software IEDB (Bepipred Linear Epitope Prediction 2.0), ABCpred (https://webs.iiitd.edu.in/raghava/abcpred (accessed on 23 May 2025), and the SVMTriP database (http://sysbio.unl.edu/SVMTriP (accessed on 24 May 2025) [21,22].The response threshold criteria for the three databases were set at 0.5, 0.8, and 1.0, respectively.
2.9.2. Prediction of T Cell Epitopes for Omp16 and Omp19 Proteins
Integrated prediction of T-cell epitopes for Omp16 and Omp19 proteins was performed using the online prediction software IEDB and the SYFPEITHI database (https://syfpeithi.de/) (accessed on 25 May 2025) [23,24,25]. Major histocompatibility complex (MHC) molecules are categorized into class I and class II, which activate cytotoxic T lymphocytes (CTL) and helper T lymphocytes (HTL) in the human body, respectively. Based on a list of common and well-documented human leukocyte antigen (HLA) alleles in the Chinese population, four alleles encoding MHC class I molecules (HLA-A02:01, HLA-A11:01, HLA-A24:02, HLA-C07:02) and four MHC class II alleles (HLA-DRB103:01, HLA-DRB107:01, HLA-DRB109:01, HLA-DRB115:01) were selected [26].Peptides with an IC50 value < 500 nmol/L predicted by NetMHCpan 3.0 and an immunogenicity score > 0.2 for MHC-I were identified as CTL epitopes. HTL epitopes were screened using NetMHCIIpan 4.0 from the IEDB database by selecting peptides ranked within the top 0.2%. The SYFPEITHI database was applied to predict both CTL and HTL epitopes, and peptides with a score > 20 were selected.
2.10. Prediction of Epitope Allergenicity and Toxicity
The toxicity of each epitope was assessed using the ToxinPred server (https://webs.iiitd.edu.in/raghava/toxinpred/ (accessed on 29 May 2025), while allergenicity was evaluated via the AllergenFP server (https://ddg-pharmfac.net/AllergenFP/ (accessed on 29 May 2025) Candidate epitopes identified as non-toxic and non-allergenic were selected for further analysis [27,28].
2.11. Molecular Docking
Molecular docking is a computational technique that predicts receptor-ligand interactions by minimizing the binding energy between three-dimensional protein structures. The amino acid sequences of Omp16 and Omp19 were retrieved from the UniProt database, while the crystal structures of the receptor proteins used for docking were obtained from the RCSB PDB database (https://www.rcsb.org/ (accessed on 5 June 2025) [29]. The most prevalent MHC class I and II alleles in the Chinese population, HLA-A02:01 and HLA-DRB115:01, were selected to form complexes with their corresponding T-cell receptors (TCRs). Molecular docking was performed between Omp16 (ID: P0A3S7) and Omp19 (ID: P0A3P1) and the following receptors: ① B-cell receptor (BCR, PDB ID: 5IFH); ② HLA-A02:01 (PDB ID: 6TDS) complexed with a cytotoxic T lymphocyte TCR (PDB ID: 5YXN); and ③ HLA-DRB115:01 (PDB ID: 8TBP) complexed with a helper T lymphocyte TCR (PDB ID: 1YMM). Docking simulations were carried out using AutoDock Vina 1.1.2. Prior to docking, all receptor structures were preprocessed with PyMol 2.5.2. The results were visualized and analyzed to evaluate the binding affinity of Omp16 and Omp19 to immune receptors and MHC molecules.
2.12. Immune Simulation
Immune stimulation simulations were performed using the C-ImmSim server (https://150.146.2.1/C-IMMSIM/index.php (accessed on 14 June 2025) to predict the expression dynamics of T cells, B cells, and cytokines in response to Omp16 and Omp19 protein stimulation [30].Parameter configurations were set as follows: random seed and simulation volume were set to default values; HLA alleles were selected according to the server’s recommended settings. The simulation was run for 800-time steps. Three injections of Omp16 protein were administered at days 0, 28, and 42, respectively.
3. Results
3.1. Constructing Recombinant Plasmids and Synthesizing mRNA via in Vitro Transcription
The target genes omp16 (about 507 bp), omp19 (about 534 bp), and EGFP (about 1082 bp) were amplified by PCR. The resulting fragments were ligated into a vector and transformed into E. coli competent cells. Positive clones were screened and identified by double digestion with Xho I and Hind III. The results showed that the digested plasmid produced a band at 4200 bp, while the target genes appeared as bands below 1000 bp (Figure 1B). Sequencing of the recombinant plasmids and alignment with reference sequences from GenBank confirmed that the omp16 and omp19 gene sequences were 100% correct. The recombinant plasmids pIVTRup-omp16/omp19 were linearized by digestion with EcoR I, and the full-length EGFP functional region was obtained by amplification. As shown in Figure 1C, the product sizes were consistent with expectations. Using an in vitro transcription kit, linearized recombinant plasmids and the amplified EGFP product were used as templates to synthesize structurally intact and mature omp16-mRNA, omp19-mRNA, and EGFP-mRNA. Electrophoresis on a 5% denaturing PAGE gel revealed that the omp16-mRNA and omp19-mRNA were approximately 600 nt in length, and the EGFP-mRNA was approximately 1200 nt, which matched the predicted sizes (Figure 1D).
3.2. In Vitro Cell Experiments
Twenty-four hours after transfection of HEK 293T cells with in vitro transcribed mRNA, green fluorescent protein expression was observed under fluorescence microscopy in the EGFP-mRNA group, while no fluorescence was detected in the blank control group (Figure 2A). The transfection efficiency was estimated to be approximately 14.1% using ImageJ 1.54g. Using the EGFP-mRNA group as a reference, these results indicate that the mRNA was successfully translated into functionally intact protein in HEK 293T cells. The target proteins were purified using affinity resin conjugated with anti-Flag antibodies. Western blot analysis revealed protein bands at approximately 16 kDa for the omp16-mRNA group and 19 kDa for the omp19-mRNA group, consistent with expected sizes, while no obvious band was observed in the EGFP-mRNA group (Figure 2B). These findings demonstrate that both omp16-mRNA and omp19-mRNA were stably expressed in HEK 293T cells. To further investigate the immunogenicity of the expressed proteins, an indirect ELISA was performed. Wells were coated with 0.6 μg of purified recombinant protein antigen per well and incubated with diluted serum from patients with brucellosis, using healthy human serum as a control. The results showed that both rOmp16 and rOmp19 proteins specifically bound to antibodies in the patient sera (*** p <0.001), and the binding affinity of rOmp16 was significantly stronger than that of rOmp19 (* p <0.05) (Figure 2C).
3.3. Prediction of Antigenic Epitopes in Omp16 and Omp19 Proteins
3.3.1. Prediction of B-Cell Antigenic Epitopes in Omp16 and Omp19 Proteins
Based on hidden Markov models, artificial neural networks, and support vector machine algorithms, linear B-cell epitopes of Omp16 and Omp19 proteins were predicted using the online tools BepiPred 2.0, ABCPred, and SVMTriP. High-scoring sequences were identified (Table 3 and Table 4). Specifically, the IEDB BepiPred 2.0 database was employed to predict linear B-cell epitopes using a response threshold of 0.5. Ultimately, overlapping sequences predicted by multiple tools were selected as final linear B-cell epitopes. For Omp16, the chosen epitopes were L_27–38_ (SKKNLPNNAGDL), L_62–72_ (FFDLDSSLIRA) and L_100–110_ (DERGTREYNLA). For Omp19, the selected epitopes included L_74–89_ (TQVASLPPASAPDLTP), L_111–121_ (QTKYGQGYRAG) and L_160–171_ (QGRFDGQTTGGQ) (Table 5).
3.3.2. Prediction of T-Cell Antigenic Epitopes in Omp16 and Omp19 Proteins
Cytotoxic T lymphocyte (CTL) epitopes for Omp16 and Omp19 proteins were predicted using the IEDB (MHC-I Binding, TAP Transport, and MHC-I Immunogenicity) and SYFPEITHI databases. High-scoring overlapping sequences were selected as candidate epitopes (Table 6 and Table 7). The final selected CTL epitopes for Omp16 were L_9–17_ (RSPIAIALF), L_21–29_ (AVAGCASKK), L_87–95_ (YPQYSITI) and L_118–126_ (ATRDFLASR). For Omp19, the CTL epitopes chosen were L_10–19_ (SLAAAGIVL), L_99–107_ (SLGGQSCKI) and L_136–144_ (AVNGKQLVL) (Table 5). Helper T lymphocyte (HTL) epitopes were predicted using the IEDB (MHC-II Binding) and SYFPEITHI databases, with high-scoring overlapping sequences identified as candidate epitopes (Table 8 and Table 9). The final HTL epitopes selected for Omp16 included L_11–25_ (PIAIALFMSLAVAGC), L_58–72_ (GDRIFFDLDSSLIRA) and L_119–132_ (TRDFLASRGVPTN). For Omp19, the HTL epitopes chosen were L_57–71_ (PTQFPNAPSTDMSAQ) and L_140–151_ (KQLVLYDANGGTVAS) (Table 5).
3.3.3. Prediction of Allergenicity and Toxicity
Prior to the consideration of viable candidate vaccines, it is essential to analyze the allergenicity and toxicity profiles of the conserved epitopes identified in the previous stages. Subsequent evaluation using AllergenFP and ToxinPred confirmed the selection of non-allergenic and non-toxic epitopes (Table 10). For the Omp16 protein, the following epitopes were selected: two B-cell epitopes: L_62–72_ (FFDLDSSLIRA) and L_100–110_ (DERGTREYNLA); two CTL epitopes: L_9–17_ (RSPIAIALF) and L_118–126_ (ATRDFLASR); two HTL epitopes: L_11–25_ (PIAIALFMSLAVAGC) and L_58–72_ (GDRIFFDLDSSLIRA). For the Omp19 protein, the selected epitopes included: three B-cell epitopes: L_74–89_ (TQVASLPPASAPDLTP), L_111–121_ (QTKYGQGYRAG) and L_160–171_ (QGRFDGQTTGGQ); one CTL epitope: L_136–144_ (AVNGKQLVL); two HTL epitopes: L_57–71_ (PTQFPNAPSTDMSAQ) and L_140–151_ (KQLVLYDANGGTVAS).
3.4. Molecular Docking
The docking simulations revealed that both Omp16 and Omp19 form stable complexes with their respective target receptors (Figure 3). Specifically, the interaction between Omp16 and BCR involved 11 hydrogen bonds, while Omp19 bound to BCR through 9 hydrogen bonds (Figure 3A,B). In the ternary complexes with HLA-A02:01 and the CTL-surface TCR, Omp16 engaged in 8 hydrogen bonds, compared to 6 for Omp19 (Figure 3C,D). Both proteins formed interactions involving 9 hydrogen bonds within the complexes comprising HLA-DRB115:01 and the HTL-surface TCR (Figure 3E,F). These results indicate that Omp16 exhibits a superior binding stability relative to Omp19 in complexes with both BCR and MHC class I–TCR, suggesting its enhanced potential in activating humoral immune responses and cytotoxic T-cell mechanisms.
3.5. Immune Simulation
3.5.1. Immune Stimulation of Innate Immune Cells by Omp16 and Omp19 Proteins
The C-ImmSim server is a computational tool designed for simulating immune responses. Simulations performed using this platform allow for the observation of dynamic changes in human immune cells. Results from C-ImmSim indicated that both Omp16 and Omp19 proteins significantly activated macrophages (Figure 4A,B). Specifically, the population of active macrophages remained at approximately 150 cells/mm^3^ from the first to the third immunization, followed by a rapid decline to around 20 cells/mm^3^ by 60 days after the initial inoculation. Furthermore, dendritic cells (DCs), as the most potent antigen-presenting cells (APCs), play a critical role in presenting vaccine antigens to T cells. Our simulation showed that vaccination with Omp16 and Omp19 also induced significant activation of DCs (Figure 4C,D). Upon protein stimulation, the total number of DCs per state stabilized at approximately 100 cells/mm^3^, while the count of activated DCs increased rapidly at a rate of 20 cells/mm^3^ per state.
3.5.2. Immune Stimulation of Adaptive Immune Cells by Omp16 and Omp19 Proteins
Adaptive immune cells, primarily including CD4^+^ T cells, CD8^+^ T cells, and B cells, play a critical role in counteracting the pathogenesis of Brucella. Our results demonstrated that stimulation with Omp16 protein induced a significant increase in the populations of total T helper (TH) cells, non-memory TH cells, and memory TH cells, which peaked after the second stimulation (Figure 5A, reaching approximately 12,500 cells/mm^3^, 11,000 cells/mm^3^, and 1600 cells/mm^3^, respectively). Concurrently, the numbers of active and resting CD4^+^ T cells per state also reached their highest levels post-second stimulation, at 9500 cells/mm^3^ and 3500 cells/mm^3^, respectively (Figure 5C). In contrast, Omp19 protein stimulation resulted in a comparatively lower increase in CD4^+^ T cell quantities. These also peaked following the second stimulation, with total TH cells, non-memory TH cells, and memory TH cells reaching approximately 10,000 cells/mm^3^, 9000 cells/mm^3^, and 1100 cells/mm^3^, respectively (Figure 5B). After the second stimulation, the counts of active and resting CD4^+^ T cells per state peaked at 7000 cells/mm^3^ and 3000 cells/mm^3^, respectively (Figure 5D).
According to the C-ImmSim server results, two stimulations with Omp16 and Omp19 proteins induced a peak in the number of non-memory CTLs per state, reaching maximum values of 1150 cells/mm^3^ and 1155 cells/mm^3^, respectively (Figure 6A,B). Furthermore, around day 40 after the initial immunization, the population of active CTLs per state peaked at 1000 cells/mm^3^ for both proteins, while the number of inactive CTLs per state reached a trough of 100 cells/mm^3^ (Figure 6C,D).
We also predicted the B-cell population per state following immunization with Omp16 and Omp19 proteins. The results indicated that after the second stimulation with Omp16, the total B-cell count peaked at 700 cells/mm^3^ (Figure 7A). The number of active B cells reached a maximum of 700 cells/mm^3^ post-second stimulation, while preseting-2 cells peaked at 340 cells/mm^3^ after the first stimulation (Figure 7C). In contrast, immunization with Omp19 elicited a slightly lower B-cell response. The total B-cell count reached a peak of 600 cells/mm^3^ following the second stimulation (Figure 7B). Active B cells peaked at 600 cells/mm^3^ after the second stimulation, and preseting-2 cells reached a maximum of 350 cells/mm^3^ after the first stimulation (Figure 7D).
3.5.3. Humoral Immune Response Induced by Omp16 and Omp19 Proteins
In computer analysis, IgM antibody levels peaked at 85,000/mL and 50,000/mL, respectively, after the second injection of Omp16 and Omp19 proteins, and then gradually decreased (Figure 8). IgG1 + IgG2 antibody levels peaked at 350,000/mL and 300,000/mL, respectively, after the second immunization, and then gradually decreased. Therefore, Omp16 was able to produce higher levels of specific antibodies.
4. Discussion
Brucella, as an intracellular parasite, survives and replicates in macrophages and placental trophoblasts by disrupting the host immune response through immune evasion mechanisms [31,32]. Upon injection, mRNA vaccines encapsulated in lipid nanoparticles (LNPs) are taken up by activated monocytes and DCs. These cells synthesize the encoded antigenic proteins and present the processed antigens in lymphoid tissues, effectively driving adaptive immune responses and demonstrating strong efficacy in activating cellular immunity [33].
The selection of appropriate antigens is crucial for the development of mRNA vaccines. Omp16 and Omp19 are outer membrane lipoproteins that act as pathogen-associated molecular patterns of Brucella and can activate DCs and induce Th1 immune responses in vivo [34]. Extensive research has been conducted on DNA vaccines and recombinant protein vaccines targeting Omp16 and Omp19. The molecular weights of rOmp16 and rOmp19 proteins expressed and purified by prokaryotic expression are 16 kDa and 19 kDa respectively, which is consistent with Western blot results [35]. EGFP-mRNA was prepared with EGFP as reporter gene, and green fluorescence was observed 24 h after transfection into HEK 293T cells, which further confirmed the successful expression of EGFP-mRNA in eukaryotic expression system.
Most eukaryotic mRNAs require a 7-methylguanosine at the 5 ‘end We synthesized the complete mRNA using the ARCA cap analog in the in vitro transcription kit and the Poly (A) tail sequence of the template plasmid, but it can only be used for in vitro cell transfection experiments. For in vivo immunogenicity studies in animals, the incorporation of signal peptide sequences at the 5′ end is essential. This modification enables the translated antigen to be directed through the endoplasmic reticulum/Golgi pathway, facilitating its secretion into the extracellular space [36]. There are two major obstacles in the development of mRNA vaccines: mRNA instability and high inflammatory response [37,38]. Using ionizable lipid nanoparticle technology, intact mRNA can be efficiently delivered into the cytoplasm of cells and then translated into encoded antigenic proteins [39]. Dogo 4 is a less toxic, mannose functionalized liposome that efficiently delivers siRNA in vitro and in vivo [40]. In this study, Dogo 4 liposomes were used to encapsulate and transport mRNA into cells, greatly preserving mRNA integrity and improving transfection efficiency.
The purified rOmp16 and rOmp19 proteins were reacted with serum samples from patients infected with Brucella. Analysis revealed that both proteins specifically bound to IgG antibodies in the patient sera, indicating that the in vitro transcribed mRNA was successfully expressed in the eukaryotic system and retained immunogenicity. This finding indirectly validates the initial step of mRNA vaccine-induced immune activation in vivo. However, further animal studies are required to assess the in vivo expression efficiency of the mRNA and its ability to induce antibody production. Notably, we observed differential reactivity between rOmp16 and rOmp19 proteins expressed in mRNA-transfected cells when probed with patient sera, while no such difference was detected in the healthy control group. This discrepancy is particularly intriguing and warrants deeper exploration beyond simply attributing it to variations in immunogenicity. The stronger binding affinity of rOmp16 compared to rOmp19 (as shown in Figure 2C) suggests that rOmp16 may possess a higher density of accessible epitopes, or perhaps its epitopes are more conformationally stable and better recognized by the circulating antibodies in brucellosis patients. This could be due to structural differences between the two proteins, leading to varying degrees of antigen processing and presentation in vivo, or differential exposure of key antigenic determinants during natural infection. Future studies could involve detailed structural analysis of rOmp16 and rOmp19, as well as epitope mapping using patient sera, to elucidate the molecular basis for this observed difference in immunogenicity. Understanding these nuances is critical for optimizing antigen selection and design in multi-epitope mRNA vaccines.
Furthermore, this study predicted the dominant epitopes of Omp16 and Omp19 proteins and performed molecular docking with MHC molecules and surface receptors of immune cells to investigate their mechanisms of specific binding to antibodies in patient sera, as well as their ability to elicit humoral and cell-mediated immune responses. B-cell epitope prediction tools utilize the principle that complementarity-determining regions (CDRs) of antibodies recognize antigenic epitopes with high specificity. By analyzing antibody–antigen protein structures, these tools enable accurate prediction of epitopes across diverse antigens [41]. B-cell epitopes of Omp16 and Omp19 were systematically mapped with the ABCpred algorithm, mirroring the epitope-prediction workflow previously established for the L7/L12 ribosomal protein [25]. Two dominant epitopes were identified in Omp16 (L_62–72_ and L_100–110_), and three were identified in Omp19 (L_74–89_, L_111–121_, and L_160–171_). In contrast, predictions of CTL and HTL epitopes were based on the binding affinity between antigenic peptides and MHC class I and II molecules [42,43]. Through integrated use of IEDB and SYFPEITHI, we identified two CTL epitopes (L_9–17_ and L_118–126_) and two HTL epitopes (L_11–25_ and L_58–72_) for Omp16, while one CTL epitope (L_136–144_) and two HTL epitopes (L_57–71_ and L_140–151_) were predicted for Omp19. These dominant epitopes critically determine the immunogenicity of the antigens and play essential roles in the immune response against Brucella infection. Multi-epitope vaccine, constructed by linking and optimizing these epitopes, have been shown to induce high levels of both humoral and cellular immunity in mice [44,45]. The identification of these specific B-cell, CTL, and HTL epitopes provides a rational basis for the design of a multi-epitope mRNA vaccine. By focusing on these immunodominant regions, we can potentially enhance the breadth and magnitude of the immune response, leading to more effective protection. For instance, the presence of multiple CTL epitopes suggests that both Omp16 and Omp19 can effectively stimulate cytotoxic T-lymphocyte responses, which are crucial for clearing intracellular pathogens like Brucella. The distinct sets of epitopes identified for Omp16 and Omp19 also highlight the potential benefit of combining both antigens in a vaccine strategy to achieve a more comprehensive immune coverage against different strains or stages of infection.
In the immune response to Brucella infection, bacterial antigens are phagocytized by antigen presenting cells and processed into antigenic epitopes. These peptide fragments are present on the cell surface and can be recognized and bound by immune cells. Docking results of Omp16 and Omp19 with BCR, TCR and MHC molecules show that Omp16 and Omp19 can form stable complexes with them and form a large number of hydrogen bonds. Hydrogen bonds reduce the energy of the complex system and the binding tends to stabilize [46]. For example, aspartic acid in L_74–89_ and glycine in L_112–122_ can form hydrogen bonds with BCR, and asparagine in L_136–144_ can form hydrogen bonds with MHCI-TCR complexes. The molecular docking analysis provides crucial insights into the structural basis of antigen-receptor interactions. The observation that Omp16 forms more hydrogen bonds with BCR and MHC class I-TCR complexes compared to Omp19 (11 vs. 9 with BCR; 8 vs. 6 with MHC I-TCR) is significant. This suggests a potentially stronger and more stable binding affinity for Omp16 with these key immune receptors. A higher number of hydrogen bonds implies a more robust interaction, which could translate to more efficient antigen presentation and subsequent activation of B cells and cytotoxic T cells. This finding aligns with our ELISA results showing stronger immunogenicity for rOmp16 and provides a mechanistic explanation at the molecular level. For vaccine design, this suggests that Omp16 might be a more potent immunogen, or that specific modifications to Omp19 could be explored to enhance its binding stability and immunogenicity. Further computational and experimental validation of these binding energies and their correlation with immune activation would be valuable.
To further characterize the immunoinformatic profiles of Omp16 and Omp19, we utilized the C-IMMsim server to simulate cell-mediated immune responses induced by an MP3RT vaccine formulation. The simulation results demonstrated that following three successive immunizations, the number of activated macrophages remained stable at approximately 100 cells/mm^3^, while the total DCs count was maintained at around 150 cells/mm^3^, with actively secreting DCs stabilizing at 20 cells/mm^3^. These data suggest that both Omp16 and Omp19 can initiate innate immune responses by activating macrophages and DCs. The C-ImmSim results are highly encouraging, as they predict a robust activation of innate immune cells (macrophages and DCs) by both Omp16 and Omp19. The sustained levels of activated macrophages and DCs after multiple immunizations indicate that these antigens can effectively prime the immune system for adaptive responses. DCs, as professional antigen-presenting cells, are critical for initiating T cell responses. Their significant activation suggests that the mRNA vaccine encoding Omp16 and Omp19 would efficiently present antigens to T cells, thereby bridging innate and adaptive immunity. The stability of these cell populations over time also implies a sustained immune surveillance, which is desirable for long-term protection against Brucella.
Regarding adaptive immune responses, the simulation showed that Omp16 stimulation led to a higher peak in total T helper (TH) cells, non-memory TH cells, and memory TH cells compared to Omp19. Specifically, Omp16 induced approximately 12,500 cells/mm^3^ total TH cells, while Omp19 induced around 10,000 cells/mm^3^. This difference in TH cell activation is crucial because CD4^+^ TH cells play a central role in coordinating both humoral and cellular immunity. A stronger TH cell response, particularly the generation of memory TH cells, suggests a more potent and long-lasting adaptive immune response. Similarly, while both proteins induced comparable peaks in non-memory cytotoxic CTLs, the overall kinetics and magnitude of the CD4^+^ T cell response appear more favorable with Omp16. This reinforces the notion from ELISA and molecular docking that Omp16 might be a more potent component for inducing a comprehensive adaptive immune response. The predicted B-cell responses also showed Omp16 inducing a slightly higher total B-cell count and active B cells compared to Omp19 (700 vs. 600 cells/mm^3^). This further supports the stronger humoral response observed with Omp16. The higher IgM and IgG1 + IgG2 antibody levels predicted for Omp16 (85,000/mL IgM and 350,000/mL IgG for Omp16 vs. 50,000/mL IgM and 300,000/mL IgG for Omp19) directly correlate with its superior ability to elicit specific antibodies, which is a cornerstone of protective immunity against extracellular pathogens and toxins, and also contributes to intracellular pathogen clearance.
While these in silico modeling results provide valuable insights into the potential immune responses elicited by Omp16 and Omp19, it is crucial to acknowledge that computational predictions are not a substitute for experimental validation. The C-ImmSim server offers a powerful tool for hypothesis generation and preliminary assessment of immunogenicity profiles; however, the complex interplay of immune cells and molecules in a living organism cannot be fully replicated in a simulated environment. Therefore, the next critical steps in the development of this mRNA vaccine candidate must involve rigorous in vivo studies. These will include evaluating the actual immunogenicity of the mRNA vaccine in animal models (e.g., mice or guinea pigs), assessing the magnitude and quality of both humoral and cellular immune responses, and most importantly, conducting challenge studies to determine the protective efficacy against B. melitensis infection. Therefore, we further evaluated the immunogenicity by predicting epitopes for common mouse MHC alleles (See Supplementary Materials Tables S1–S5). Such in vivo experiments are indispensable for confirming the translational potential of our in silico and in vitro findings and moving closer to a viable human vaccine.
In summary, the integrated bioinformatic and in vitro experimental findings strongly support the potential of Omp16 and Omp19 as promising antigens for a B. melitensis mRNA vaccine. The observed differences in immunogenicity and receptor binding between Omp16 and Omp19 highlight the importance of careful antigen selection and potentially the synergistic benefits of a multi-antigen vaccine approach. While this study provides a preliminary exploration, the successful expression of immunogenic proteins in eukaryotic cells, coupled with robust immune simulation predictions, lays a solid theoretical foundation. Future research should focus on in vivo animal studies to validate these findings, assess the protective efficacy of the mRNA vaccine, and further optimize the vaccine formulation and delivery system. Additionally, investigating the precise mechanisms underlying the differential immunogenicity of Omp16 and Omp19 at a structural and cellular level will be crucial for the rational design of next-generation brucellosis vaccines.
5. Conclusions
In this study, mRNA sequences encoding the omp16 and omp19 genes of B. melitensis were synthesized via in vitro transcription. The resulting mRNAs mediated the expression of immunogenic proteins in mammalian cells. Furthermore, bioinformatic approaches were employed to identify dominant antigenic epitopes of Omp16 and Omp19 and to simulate immune response profiles following vaccination. These findings provide a foundational investigation for the development of an mRNA-based vaccine against B. melitensis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yu H. Gu X. Wang D. Wang Z. Brucella infection and Toll-like receptors Front. Cell. Infect. Microbiol.202414134268410.3389/fcimb.2024.134268438533384 PMC 10963510 · doi ↗ · pubmed ↗
- 2Laine C.G. Johnson V.E. Scott H.M. Arenas-Gamboa A.M. Global Estimate of Human Brucellosis Incidence Emerg. Infect. Dis.2023291789179710.3201/eid 2909.23005237610167 PMC 10461652 · doi ↗ · pubmed ↗
- 3KılıçS. Çelebi B. Turan M. Brucella melitensis and Brucella abortus genotyping via real-time PCR targeting 21 variable genome loci J. Microbiol. Methods 202118010612510.1016/j.mimet.2020.10612533333100 · doi ↗ · pubmed ↗
- 4Dorneles E.M. Sriranganathan N. Lage A.P. Recent advances in Brucella abortus vaccines Vet. Res.2015467610.1186/s 13567-015-0199-726155935 PMC 4495609 · doi ↗ · pubmed ↗
- 5Roop R.M.2nd Barton I.S. Hopersberger D. Martin D.W. Uncovering the Hidden Credentials of Brucella Virulence Microbiol. Mol. Biol. Rev.202185 e 00021-1910.1128/MMBR.00021-1933568459 PMC 8549849 · doi ↗ · pubmed ↗
- 6Chen S. Fu Q. Mandlaa M. Wang Q. Sheng D. Saijilahu S. Tana T. Yang D. Whole Genome Sequencing of Brucella melitensis Isolated from Patients in Hohhot, China Microbiol. Resour. Announc.202312 e 000322310.1128/mra.00032-2336972233 PMC 10112156 · doi ↗ · pubmed ↗
- 7Young E.J. Hasanjani Roushan M.R. Shafae S. Genta R.M. Taylor S.L. Liver histology of acute brucellosis caused by Brucella melitensis Hum. Pathol.2014452023202810.1016/j.humpath.2014.07.00725147098 · doi ↗ · pubmed ↗
- 8Masjedian Jezi F. Razavi S. Mirnejad R. Zamani K. Immunogenic and protective antigens of Brucella as vaccine candidates Comp. Immunol. Microbiol. Infect. Dis.201965293610.1016/j.cimid.2019.03.01531300122 · doi ↗ · pubmed ↗