Effects of NaCl Treatment on Flavonoid Biosynthesis and Antioxidant System During Buckwheat Germination
Miaoyao Yu, Meixia Hu, Dehcontee Diana Adams, Meilin Wang, Zhengfei Yang, Jiangyu Zhu, Yongqi Yin

TL;DR
This study shows how salt treatment affects flavonoid production and antioxidant activity in buckwheat sprouts.
Contribution
The study identifies specific enzyme activities and gene expressions linked to flavonoid biosynthesis under NaCl stress in buckwheat.
Findings
80 mM NaCl treatment maximized total flavonoid content in three-day-old buckwheat sprouts.
NaCl treatment activated the antioxidant system, increasing enzyme activity and gene expression.
Malondialdehyde and hydrogen peroxide levels rose, indicating growth inhibition in treated sprouts.
Abstract
Exposure to abiotic stresses commonly stimulates the production of secondary metabolites in plants, and flavonoids represent a major class of these bioactive compounds. NaCl effects on antioxidant system treatment and flavonoid production in buckwheat sprouts was examined in this study using buckwheat as the raw material. In order to clarify the regulatory function of NaCl in these physiological processes, the changes in pertinent indices of buckwheat sprouts exposed to the control and NaCl treatments were studied. The results indicated that at three days old, the sprouts subjected to 80 mM NaCl treatment exhibited the highest total flavonoid content. The significant increase in enzyme activity (cinnamate 4-hydroxylase and 4-coumaroyl-CoA ligase, etc.) responsible for flavonoid biosynthesis provides strong evidence for this conclusion. The antioxidant system in buckwheat was activated…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
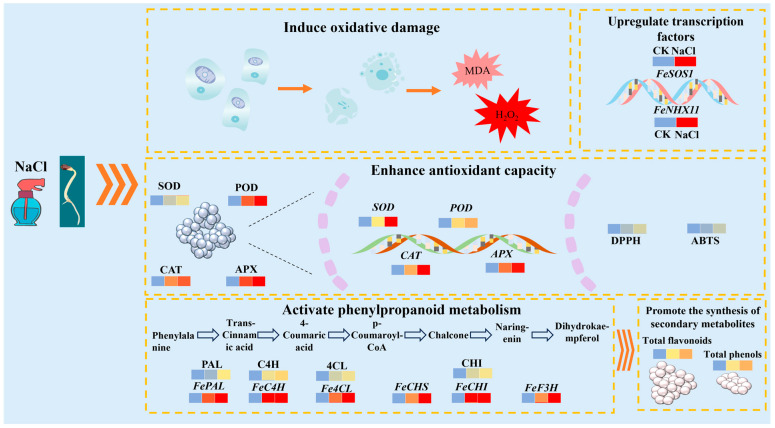 Figure 7
Figure 7- —Jiangsu Provincial Sci-Tech Deputy General Manager Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSeed and Plant Biochemistry · Plant Gene Expression Analysis · Phytochemicals and Antioxidant Activities
1. Introduction
As an important group of plant secondary metabolites, flavonoids function as key protective compounds that mediate plant responses to biotic and abiotic stresses. [1]. Salt stress, as a common abiotic stressor, has recently been acknowledged for its role in enhancing the accumulation of bioactive compounds in plants. Relevant research has demonstrated that NaCl application effectively stimulates the biosynthesis of secondary metabolites, such as flavonoids and phenolic compounds, during seed germination in crops, including soybean [2], maize [3], and millet [4]. Flavonoids exhibit multiple pharmacological activities, including antihypertensive, anticancer, antioxidant, and antiviral effects [5,6]. Research indicates that paclitaxel, a flavonoid derivative found in natural plants, inhibits tumor growth by modulating the signaling pathways [7]. Additionally, dauricine interferes with specific cyclins and activates caspases, thereby impeding tumor cell proliferation and promoting apoptosis [8,9]. Flavonoids can effectively prevent insulin resistance and oxidative stress initiated by high-fructose diets by reducing insulin concentrations and enhancing insulin signaling molecules [10]. Buckwheat, as an emerging functional food ingredient, is rich in flavonoid bioactive compounds [11], with numerous processing applications within the food, health supplement, and pharmaceutical sectors [12]. Due to the high flavonoid content in buckwheat and the close relationship between flavonoids and human health requirements, buckwheat has emerged as an essential raw ingredient in functional food formulation and phytopharmaceutical products.
To increase flavonoid levels in Fagopyrum esculentum, our previous findings indicate that germination enhances both the enzymatic activities and transcriptional expression of cinnamate 4-hydroxylase (C4H), chalcone isomerase (CHI), 4-coumaroyl-CoA ligase (4CL) and phenylalanine ammonia-lyase (PAL) in the flavonoid biosynthetic pathway, thereby facilitating flavonoid production and accumulation. [13]. The biosynthesis of flavonoids in plants originates from the common phenylpropanoid pathway and is subsequently synthesized through branch pathways of phenylpropanoid metabolism [14] and proceeds through branch-specific enzymatic reactions involving a series of complex enzyme-mediated steps, ultimately leading to the formation of flavonoid compounds [15]. Flavonoid accumulation in plants is associated with various factors; upon experiencing abiotic stress, plants activate their antioxidant systems, leading to flavonoid enrichment. Under UVB and extreme temperature stress, UVB irradiation enhances flavonoid accumulation through the upregulation of gene expression and increased activities of key enzymes involved in flavonoid biosynthesis, including chalcone synthase and phenylalanine ammonia-lyase, thereby enhancing flavonoid biosynthesis in soybean [16]. Tomato seedlings subjected to heat shock treatment exhibit a significant increase in flavonoid content, thereby enhancing their antioxidant capacity [17].
Under 60 mM NaCl exposure, pea sprouts displayed higher total phenolic levels, along with enhanced activities of PAL, C4H, and 4CL, thereby augmenting the antioxidant capacity of the sprouts [18]. Moreover, NaCl application stimulates essential enzyme activity and enhances the levels of transcriptionally associated genes, thereby promoting the biosynthesis of secondary metabolites. Relevant studies indicate that 150 mM NaCl stress enhances the biosynthesis and accumulation of resveratrol by upregulating 4CL activity and C4H, PAL, and 4CL relative levels of expression in the cotyledons, while concurrently increasing PAL and C4H activities and PAL and 4CL relative expression levels in non-cotyledonous tissues [19]. Significant upregulation of CHS (chalcone synthase) and CHI expression levels in mung bean sprouts was observed following treatment with 50 mM NaCl [20]. NaCl treatment can modulate transcription factors to enhance plant tolerance to stress. Previous studies have shown that NaCl stress significantly increases the relative transcription levels of the FtNHX1 and FtSOS1 genes. [21]. The aforementioned studies indicate that NaCl treatment promotes the enrichment of functional components in plants; however, they do not investigate or analyze the regulatory mechanisms of the antioxidant system in buckwheat under NaCl stress or the biosynthesis of flavonoid compounds. Our study not only investigates the antioxidant system regulatory mechanisms and the biosynthesis of flavonoid compounds under NaCl stress but also includes an analysis of the underlying molecular mechanisms.
Based on our previous research [13], this study selected “Pin Tian 2” buckwheat seeds as the material. Under NaCl treatment conditions, we examined how NaCl regulates flavonoid biosynthesis in buckwheat sprouts and explored the allied molecular mechanisms. This study also offers a theoretical basis for the processing of flavonoid-rich buckwheat sprouts as a raw food material.
2. Results
2.1. Growth Status, MDA Content, and H2O2 Levels
As shown in Figure 1I, NaCl significantly inhibited seedling growth in three-day-old sprouts. As Figure 1II demonstrates, in comparison to the CK, the fresh weight of three-day-old sprouts decreased by 29% under NaCl treatment (p < 0.05; one-way ANOVA), whereas the fresh weight of five-day-old sprouts decreased by 13.6%.
The levels of MDA and H_2_O_2_ serve as indicators of the degree of stress experienced by the plants. With prolonged germination, the contents of MDA and H_2_O_2_ in the sprouts initially increased and then declined (Figure 1III,IV). Compared to the CK, NaCl treatment significantly elevated the MDA and H_2_O_2_ levels in buckwheat sprouts at both three and five days. Under NaCl stress, the MDA and H_2_O_2_ contents in three-day-old sprouts increased by 35.29% and 52.94%, respectively, reaching their peak values (18.44 nM/g FW and 5.29 mM/g FW, respectively). NaCl stress led to a significant increase in MDA and H_2_O_2_ contents in three-day-old sprouts, with both reaching their peak levels during the early stages of stress. These findings suggest that NaCl treatment markedly elevates MDA and H_2_O_2_ contents in three-day-old sprouts.
2.2. Total Phenolic Content and Total Flavonoid Content
Figure 2 demonstrates that, under the CK condition, total phenolic and flavonoid levels peaked in three-day-old buckwheat buds at 10.48 mg GAE/g FW and 979.35 µg/g FW, correspondingly (p < 0.05; one-way ANOVA). Compared to CK, NaCl treatment significantly elevated the flavonoid and phenolic levels in both three- and five-day-old sprouts (Figure 2I). Specifically, NaCl application resulted in peak concentrations of 15.23 mg GAE/g FW for total phenolics and 1476.54 µg/g FW for total flavonoids in three-day-old sprouts. The flavonoid content was highly amplified, which is presumed to be an adaptive strategy of buckwheat sprouts to cope with salt stress. These findings suggest that NaCl treatment substantially promotes biosynthesis and phenolic compound accumulation as well as flavonoid concentration in buckwheat buds.
2.3. Antioxidant Capacity
As shown in Figure 3, the CK treatments reached their peak values at three days. Compared to CK, NaCl treatment highly improved the DPPH scavenging activity in three-day-old spouts (Figure 3II) (p < 0.05; one-way ANOVA). Under NaCl treatment, ABTS and DPPH scavenging activities in three-day-old buds were 60.26% and 63.28%, respectively. These results indicate that both CK and NaCl treatments achieved maximum rates of ABTS and DPPH scavenging at three days, with NaCl treatment further improving the antioxidant capacity of the shoots, meaning that exogenous stress successfully activated the antioxidant system in buckwheat sprouts.
2.4. Relative Activity of Antioxidant Enzymes and the Corresponding Gene Expression Levels
Figure 4 indicates that the highest value under CK treatment was observed on day 3. Compared to CK, NaCl treatment markedly enhanced the enzymatic activities of POD, CAT, and SOD in three-day-old buds (Figure 4I–III). Under NaCl treatment, CAT, POD, and SOD activities peaked at three days, reaching 454.98 U/g FW, 925.78 U/g FW, and 51.95 U/g FW (p < 0.05; one-way ANOVA), respectively. These values represent 1.40-fold, 1.08-fold, and 1.18-fold increases when compared to the CK, respectively. Concurrently, the POD, SOD, and CAT expression levels in three-day-old buckwheat shoots were markedly enlarged (Figure 4IV–VI). Relative to the CK, NaCl treatment markedly enhanced the expression of CAT, POD, and SOD in three-day-old buckwheat sprouts by 1.88-fold, 3.30-fold, and 5.7-fold, correspondingly (Figure 4IV–VI). These outcomes indicate that NaCl treatment meaningfully reinforces the antioxidant system in buds by increasing both antioxidant enzyme activities and the comparative levels of expression of related genes; notably, the expression levels of related genes may indicate a consistent upregulation trend with enzyme activity.
2.5. Relative Expression Levels of Key Flavonoid Biosynthetic Enzyme Activities and Their Corresponding Genes
As shown in Figure 5, the CK treatments reached their peak values at three days. Compared to the CK, the treatment of NaCl majorly increased the enzymatic C4H, PAL, 4CL, and CHI activities in three-day-old shoots (Figure 5I–IV) (p < 0.05; one-way ANOVA). Under NaCl treatment, the activities of PAL, C4H, 4CL, and CHI peaked at three days, reaching 4255.26 U/g FW, 2188.97 U/g FW, 4334.29 U/g FW, and 6454.33 U/g FW, respectively, which are evidently higher than those observed in the CK group. Additionally, the expression levels of genes related to flavonoid biosynthesis enzymes in three-day-old sprouts under NaCl treatment were significantly higher than at other germination stages (Figure 5V–X). Compared to the CK, NaCl treatment notably enhanced the expression levels of C4H, PAL, CHI, 4CL, and F3H in three-day-old shoots (Figure 5V–X). These results specify that NaCl treatment can enhance the action of key metabolic enzymes involved in flavonoid biosynthesis in buckwheat buds significantly. The upregulation amplitude of some gene expressions was higher than the increase in enzyme activity. It is speculated that there may be post-translational regulation, which requires further research for verification.
2.6. The Effect of NaCl Treatment on the Relative Expression Levels of Stress-Responsive Genetic Factor in Buckwheat Shoots
As demonstrated in Figure 6, compared to the control (CK), NaCl treatment meaningfully amplified the expression levels of stress-related genes, with the relative expression of SOS1 and NHX1 continuously upregulated during germination. On day 5, the relative expression of SOS1 in the NaCl-treated group was 19 times higher than that in the CK group, while the relative expression of NHX1 was 49.45 times higher than that in the CK group. The results obtained show that continuous upregulation of SOS1 and NHX1 gene expression suggests that buckwheat sprouts adapt to salt stress through molecular regulation, although the direct link between this mechanism and flavonoid biosynthesis requires further validation.
3. Discussion
Numerous studies have demonstrated that NaCl treatment promotes the biosynthesis of secondary metabolites, such as flavonoids and phenolic compounds, during the germination of plants including soybean [2], maize [3], and millet [4]. This was ascribed mostly to the fact that NaCl stimulated the plant’s antioxidant defense system, in which flavonoids, as essential antioxidant molecules, played a vital part, scavenging reactive oxygen species (ROS) and defending cells against oxidative impairment, thus enhancing their biosynthesis.
Plants’ phenylpropanoid pathway is regarded as one of the main pathways towards the accumulation of flavonoid secondary metabolites, and the activity level of the gene expression of major enzymes determines the combination and accumulation of the flavonoid secondary metabolites. Research has demonstrated that the activities of alpha-galactosidase and the total flavonoid content, CHI and PAL in buckwheat sprouts are positively correlated [22]. This research showed that the activities of CHI, C4L, PAL, and C4H in buckwheat buds were significantly enhanced by NaCl treatment in a comparison with non-treated samples. This improvement indicates that NaCl may stimulate the buildup of total flavonoids by adjusting the phenylpropanoid metabolic pathway. These results are in line with past reports which suggest that electromagnetic fields and weakly acidic treatments with electrolyzed water can cause alterations in the phenylpropanoid pathway in buckwheat [23]. This study focused on the Pin Tian 2 cultivar, examining germination under varying NaCl concentrations and revealing significant differences in total flavonoid accumulation. Previous studies have demonstrated that a NaCl concentration of 60 mM significantly enriches flavonoid compounds in soybeans [24], further confirming that different cultivars exhibit varying responses to NaCl stress. During germination under salt stress, flavonoid accumulation also varied among the buckwheat varieties. Initial investigations have revealed that buckwheat sprouts attain their highest flavonoid content at 10 days [25], 7 days [26] and 6 days [27], respectively. Our results have shown that the total flavonoid content tends to increase and then decrease, reaching its highest point at 3 germination days. These differences can be explained by the fact that the cultivars and germination conditions were different.
NaCl treatment in the present study had a significant effect on the activities of C4H, CHI, C4L, and PAL in buckwheat buds in comparison to the control samples. This amplification shows that NaCl enhances the amassment of total flavonoids by regulating the phenylpropanoid metabolic pathway. Besides enzyme action, the comparative expression levels of six main genes were also examined. NaCl treatment improved the expression of FeF3H, FeC4H, FeCHS, Fe4CL, FeCHI, and FePAL over three days of germination significantly. Other reports have also indicated that the expression of key genes in buckwheat sprouts is also significantly induced by germination [28], microwave exposure [29], and phenylalanine treatment [30]. It was discovered that microwave-treated buckwheat sprouts have different abundances of proteins involved in flavonoid biosynthesis and the phenylpropanoid pathway. Additional studies in the future involving transcription factor regulation as well as the use of new omics technologies like proteomics, metabolomics, and transcriptomics can give extra information on the processes that control flavonoid enrichment in buckwheat. In this study, it was noted that NaCl treatment suppressed buckwheat bud growth and caused a significant increase in MDA and H_2_O_2_, which showed that the accumulation of ROS and cell membrane destruction caused by lipid peroxidation occurred. Exposure to NaCl leads to the formation of osmotic stress and the removal of water in the cytoplasm, leading to cellular dehydration and subsequent loss of both vacuole and cytoplasmic volume. But over an extended period of evolution, buckwheat has acquired a strong self-defense system to overcome endogenous free radicals. The antioxidant defense system is also correspondingly adapted to endure under NaCl-induced stress as the levels of bioactive compounds like flavonoids and phenolics change. The findings show that NaCl treatment enormously improved the activity and expression of the major antioxidant enzymes of buckwheat sprouts such as SOD, POD, and CAT compared to normal germination conditions. This was likely due to the NaCl treatment activating the antioxidant system in buckwheat sprouts, as evidenced by a significant increase in both the activity of related antioxidant enzymes and their corresponding gene expression levels compared to the control group. The joint action of the elevated gene expression and enzyme activity of antioxidant enzymes helps the removal of reactive oxygen species, which subsequently leads to the protection of the plant cells against degrading effects. This observation is in line with earlier reports that indicated that soybean [31,32], buckwheat [33], wheat [34,35], and Elymus sibiricus [36] react to changes in the environment by improving the process of antioxidant system activation. This may indicate that NaCl treatment has a significant effect on enhancing the antioxidant ability of buckwheat sprouts including ABTS and DPPH radical scavenging activity. In the same manner, previous researchers have indicated that microwave irradiation [37,38], laser treatment [39] and trace element water treatment [40] can be used to increase the antioxidant capacity of buckwheat buds.
In conclusion, NaCl treatment produced the maximum quantity of total flavonoids and overall phenolics in buckwheat sprouts following three days of germination. NaCl treatment significantly enhanced the activities of key enzymes involved in the phenylpropanoid pathway, as well as the expression of associated genes. Also, NaCl treatment elevated the functions of antioxidant enzymes and their respective gene expressions in buckwheat buds. But NaCl treatment restrained the growth and development of buckwheat sprouts as indicated by the significant changes in the appearance of stress-associated genes and MDA and H_2_O_2_ levels (p < 0.05).
4. Materials and Methods
4.1. Germination Treatment and Experimental Design
Buckwheat seeds: The variety was Pin Tian 2, which was offered by the Crop Genetic Resources Institute of Jiangsu Academy of Agricultural Sciences in 2022. The preservation of the seeds was done at −18 C. Before the experiment, buckwheat seeds were disinfected in a 1% solution of sodium hypochlorite for 15 min, then cleaned with purified water until the pH became neutral. The seeds were then moistened in distilled water in darkness at 30 °C using a germination chamber. One hundred surface-sterilized seeds were placed in each germination tray (dimensions: 15 cm × 10 cm × 5 cm), with sterile filter paper (3 layers) used as the germination substrate. Distilled water and NaCl solution were applied via spray simultaneously every 12 h. The CK and NaCl seeds were simultaneously sprayed with 30 mL. Samples were randomly collected at 1, 3, and 5 days of germination and kept at −20 °C for future examination. Different seed treatments were applied: (1) CK, control, with distilled water spray; (2) NaCl stress treatment at 80 mM concentration. The NaCl concentration was selected based on preliminary screening (Supplementary Figure S1). (The NaCl reagent used in this experiment was purchased from Sinopharm Chemical Reagent Co., Ltd. (Shanghai, China) with a purity of ≥99.5%. The main impurities include trace sulfate (≤0.02%), chloride (≤0.005%), and heavy metals (≤0.001%). Detailed specifications are available in the product manual. A Millipore ultrapure water system was used for both the preparation of the NaCl solution and the experiments.)
A random sample of 30 buckwheat sprouts was selected, and the length of each sprout was measured using a vernier caliper with an accuracy of 0.01 cm, while the fresh weight was determined using an analytical balance with a precision of 0.001 g. Each treatment was conducted with three biological replicates, and 2 g of seedlings was randomly selected for each replicate. All measurements were performed with three technical replicates
4.2. Physiological and Biochemical Indicators
The malondialdehyde (MDA) determination was carried out based on the procedure reported by Zhuang [41]. A certain quantity of the sample was drawn and combined with trichloroacetic acid. The supernatant was removed and kept in the dark. A portion of the supernatant was, in turn, mixed with the thiobarbituric acid solution. The mixture was centrifuged once more after a quick cooldown and the reaction solution was harvested. OD_600 nm_, OD_532 nm_ and OD_450 nm_ were measured.
The determination of hydrogen peroxide (H_2_O_2_) was performed following the method described by Xue [42]. A measured amount of sample was homogenized with buffer solution. The mixture was centrifugated while the supernatant was collected. An aliquot of the supernatant was then added to the reaction solution, and absorbance was determined at OD_390nm_ and OD_593nm_.
4.3. Total Flavonoids and Total Phenolics
The total flavonoid content was estimated by using the procedure outlined by Yin et al. [43]. A given portion of the sample was mixed with ethanol, then the mixture was ultrasonicated and centrifuged. The supernatant was removed and diluted and OD_260nm_ absorbance was measured.
The phenolic content was enumerated using the process outlined by Mencin et al. [44]. One of the samples was centrifuged and a measured volume of the sample was mixed in methanol and subsequently centrifuged. A supernatant was prepared by taking an aliquot of the resultant and adding Folin–Ciocalteu reagent as well as Na_2_CO_3_ and letting the reaction proceed in the dark. OD_765nm_ absorbance was then measured.
4.4. Antioxidant Capacity
The determination of 1,1-Diphenyl-2trinitrophenylhydrazine (DPPH) and 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) radical scavenging activities was conducted following the methodology described by Zhang et al. [45].
4.5. Activity of Antioxidant Enzymes
Peroxidase (POD), catalase (CAT) and superoxide dismutase (SOD) activities were assessed following the method highlighted by Huang et al. [46]. A defined quantity of the sample was homogenized in sodium phosphate buffer, and the resulting supernatant was collected for subsequent enzymatic analysis. One unit of SOD and APX activity is defined as the amount of enzyme required to cause a change of 0.01 per minute in absorbance at OD_560nm_ and OD_290nm_, respectively. Similarly, one unit of CAT and POD activity is defined as the quantity of enzyme resulting in a change of 0.01 per minute in absorbance at OD_240nm_ and OD_470nm_, respectively.
4.6. Activity of Flavonoid Metabolism-Related Enzymes
The activity of CHI, 4CL, C4H and PAL was assessed as described by Yin et al. [24]. A certain portion of the sample was homogenized in Tris–HCl buffer and then centrifuged to separate the supernatant. A unit represented a 0.01 modification in absorbance per minute at OD_290nm_, OD_340nm_, OD_333nm_, or OD_381nm_.
4.7. Key Gene Expression Levels
4.7.1. RNA Extraction and Reverse Transcription
After grinding the buckwheat into powder using liquid nitrogen and transferring it to a centrifuge tube, add lysis buffer rapidly and mix thoroughly. Once lysis is complete, extract RNA using the RNA extraction kit (R6827, Omega, Norcross, GA, USA). Perform reverse transcription with the RNA reverse transcription kit (RR047A, TAKARA, Kyoto, Japan), then store the resulting cDNA at −80 °C for future use.
4.7.2. qPCR Primers
Table 1 presents the primer sequences for the target genes and the internal reference gene (Actin), based on the complete genome sequence of buckwheat published in the NCBI database and the gene primers reported in the relevant literature [21,47,48].
4.7.3. qPCR Reaction Conditions and Establishment of the Quantitative PCR System
Quantitative PCR was performed using TAKARA’s TB Green Premix Dimer Eraser (RR091A), TAKARA, Japan, Kyoto. The expression level of the reference gene (Actin) was normalized to 1. Genetic expression was measured using the 2^−ΔΔCt^ [43]. Based on the instructions for the TB Green Premix Dimer Eraser (RR091A, TAKARA, Japan), the specific parameters for the quantitative PCR reaction system and the PCR amplification program were established. The annealing temperature was 72 °C. Amplification efficiency ranged from 95% to 104%.
4.8. Processing of Data and Statistical Analysis
The results from the experiment are presented as mean ( ) ± standard deviation (SD), based on data from three autonomous tests. Statistical analysis was achieved using one-way analysis of variance (ANOVA), followed by Tukey’s multiple comparison test to assess differences between group means. Statistical relevance was assessed at the 0.05 level (p < 0.05). All data were scrutinized using DPS software (Version 9.0, China), and data visualization was conducted with Origin 2022.
5. Conclusions
This study found that both total flavonoid and phenolic contents in buckwheat buds reached their highest levels after 3 days of NaCl treatment. The NaCl-treated sprouts exhibited enhanced activities of important enzymes in the phenylpropanoid metabolic pathway and upregulated expression of related genes. Additionally, NaCl treatment increased antioxidant enzyme activity and the expression of their corresponding genes in buckwheat shoots. This study identified a germination treatment of 80 mM NaCl for 3 days, which enhanced the flavonoid content and antioxidant activity of buckwheat shoots, thereby offering a theoretical basis for the production of functional buckwheat sprouts. However, we observed that NaCl treatment inhibited the progress and development of the sprouts, an issue we aim to address in future research. As shown in Figure 7, The proposed hypothetical model is illustrated in the schematic diagram below.
A hypothetical model for the regulatory effects of NaCl treatment on the physiological metabolism and flavonoid biosynthesis in buckwheat sprouts.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mierziak J. Kostyn K. Kulma A. Flavonoids as Important Molecules of Plant Interactions with the Environment Molecules 201419162401626510.3390/molecules 19101624025310150 PMC 6270724 · doi ↗ · pubmed ↗
- 2Ma M. Wang P. Yang R. Gu Z. Effects of UV-B Radiation on the Isoflavone Accumulation and Physiological-Biochemical Changes of Soybean During Germination: Physiological-Biochemical Change of Germinated Soybean Induced by UV-B Food Chem.201825025926710.1016/j.foodchem.2018.01.05129412920 · doi ↗ · pubmed ↗
- 3Chavarín-Martínez C.D. Reyes-Moreno C. Milán-Carrillo J. Perales-Sánchez J.X.K. Canizalez-Román V.A. Cuevas-Rodríguez E.O. López-Valenzuela J.A. Gutiérrez-Dorado R. Effect of Germination and UV-B Elicitation on Chemical Compositions, Antioxidant Activities, and Phytochemical Contents of Underutilised Mexican Blue Maize Seeds Int. Food Res. J.20222930031010.47836/ifrj.29.2.08 · doi ↗
- 4Zhang J. Yang J. Yin Y. Germination Promotes Flavonoid Accumulation of Finger Millet (Eleusine coracana L.): Response Surface Optimization and Investigation of Accumulation Mechanism Plants 202413219110.3390/plants 1316219139204627 PMC 11360649 · doi ↗ · pubmed ↗
- 5Kim S.-H. Cui C.-B. Kang I.-J. Kim S.Y. Ham S.-S. Cytotoxic Effect of Buckwheat (Fagopyrum esculentum Moench) Hull Against Cancer Cells J. Med. Food 20071023223810.1089/jmf.2006.108917651057 · doi ↗ · pubmed ↗
- 6Zheng R.-L. Wang J. Liu S.-Y. Sun Z.-P. Zhao L.-Y. Chen G.-T. Screening and Extraction Process Optimization For Potential A-Glucosidase Inhibitors From Quinoa Seeds FMH 20241942000410.26599/FMH.2024.9420004 · doi ↗
- 7Xu L. Zhang L. Zhang S. Yang J. Zhu A. Sun J. Kalvakolanu D.V. Cong X. Zhang J. Tang J. Taxifolin Inhibits Melanoma Proliferation/Migration Impeding USP 18/Rac 1/JNK/β-catenin Oncogenic Signaling Phytomedicine 202412315519910.1016/j.phymed.2023.15519937995531 · doi ↗ · pubmed ↗
- 8Naponelli V. Piscazzi A. Mangieri D. Cellular and Molecular Mechanisms Modulated by Genistein in Cancer Int. J. Mol. Sci.202526111410.3390/ijms 2603111439940882 PMC 11818640 · doi ↗ · pubmed ↗