Animal Models for Swine Influenza Virus Research: Pathology, Viral Dynamics, and Immune Responses
Jingyu Zhang, Shuaiyu Jiang, Yupeng Fang, Jiahong Feng, Wenqing Zhang, Xiaoqing Zhang, Jie Zhang

TL;DR
This review discusses animal models used to study swine influenza virus, highlighting their roles in understanding disease mechanisms, transmission, and vaccine development.
Contribution
The paper introduces a tiered, multi-model strategy for SIV research, emphasizing the strengths and limitations of different animal models.
Findings
Animal models show significant differences in pathology, viral replication, and immune responses.
Pigs are essential for evaluating vaccine efficacy and safety due to their natural host status.
Ferrets and guinea pigs are useful for studying transmission dynamics.
Abstract
Swine influenza virus (SIV) continues to evolve and possesses notable zoonotic potential, making it an important respiratory pathogen of concern for both the global swine industry and public health. Owing to antigenic drift, genetic reassortment, and regional lineage diversity, vaccine efficacy against SIV shows marked variability across different epidemiological contexts. Therefore, establishing appropriate animal models to dissect its pathogenic mechanisms, transmission characteristics, and immune response patterns is of critical importance. This review systematically summarises the animal models commonly used in SIV research, including mice, ferrets, guinea pigs, pigs, and non-human primates, and provides an integrated analysis across three core dimensions: pathological manifestations, viral replication kinetics, and immune architecture. The evidence indicates that substantial…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
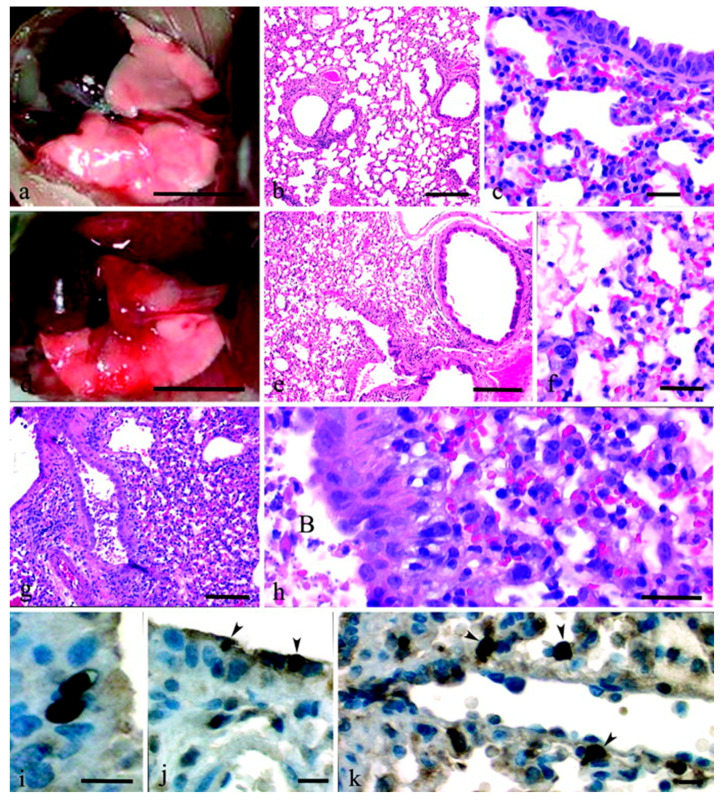 Figure 1
Figure 1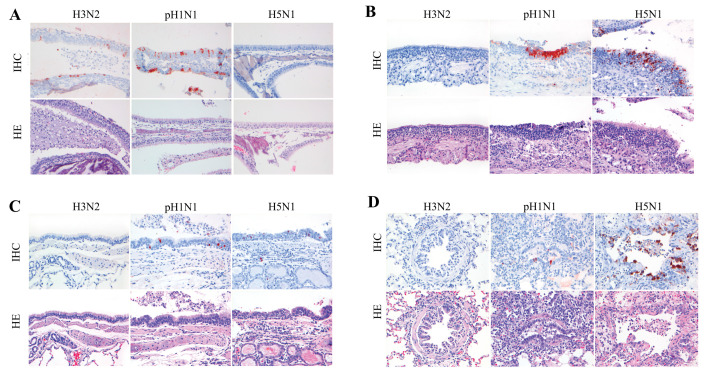 Figure 2
Figure 2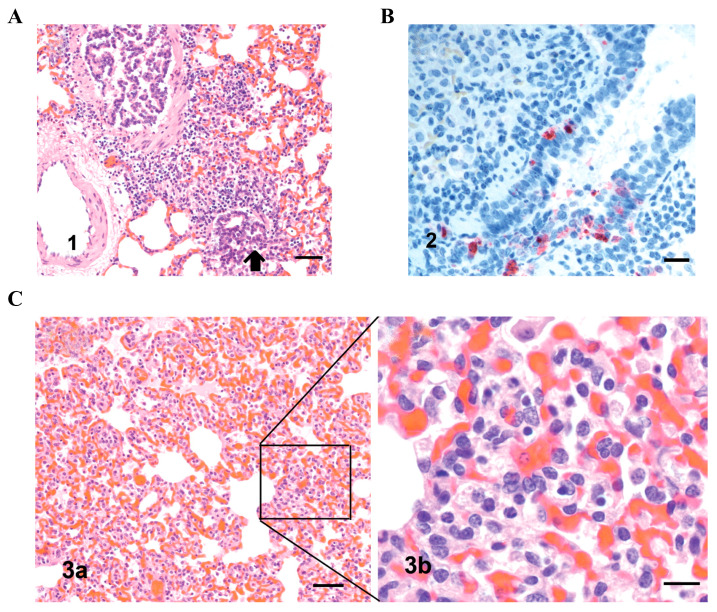 Figure 3
Figure 3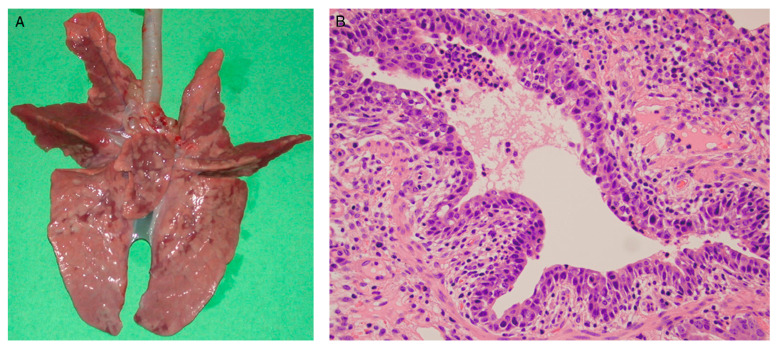 Figure 4
Figure 4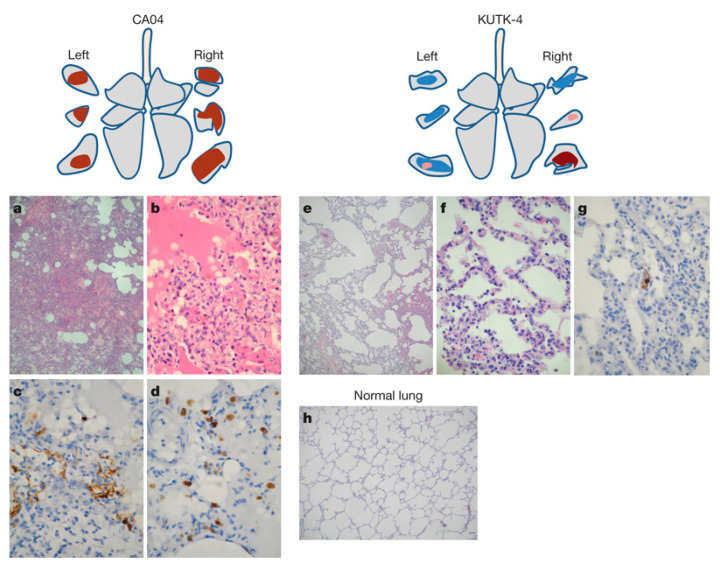 Figure 5
Figure 5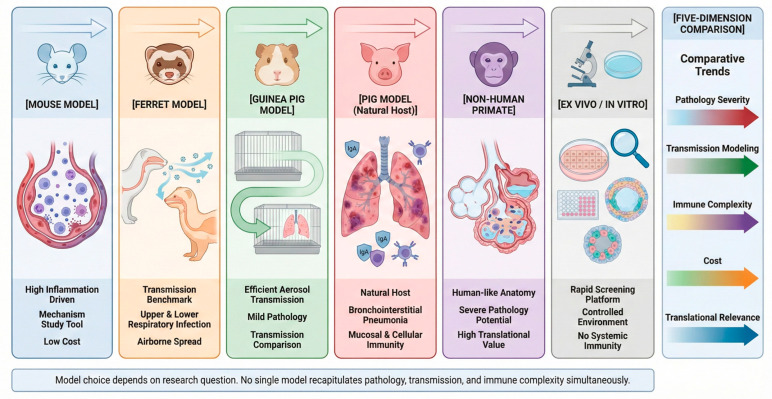 Figure 6
Figure 6- —National Natural Science Foundation of China
- —Special Project of Basic Scientific Research Funds for Colleges and Universities in Hebei Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInfluenza Virus Research Studies · Animal Disease Management and Epidemiology · Microbial infections and disease research
1. Introduction
Swine influenza virus (SIV) belongs to the family Orthomyxoviridae and is one of the major respiratory pathogens affecting the global swine industry. The viral genome consists of segmented negative-sense RNA, which readily undergoes genetic reassortment during co-infection of a host. Because porcine respiratory epithelial cells express both α-2,3 and α-2,6 sialic acid receptors, pigs are susceptible to both avian- and human-origin influenza viruses, making them an important “mixing vessel” for cross-species transmission and viral reassortment [1]. The 2009 pandemic H1N1 influenza virus originated from a triple-reassortant swine strain, highlighting the potential public health risk posed by SIV in emerging and re-emerging infectious diseases [2].
It is important to note that, although human influenza models have generated extensive mechanistic insights, their findings cannot be directly extrapolated to the swine influenza context. First, humans and pigs differ in respiratory tract architecture, pulmonary immune microenvironment, and the distribution of tissue-resident T cells [3]. Second, systematic interspecies differences exist in innate immune receptor expression profiles, inflammatory response intensity, and viral clearance kinetics. In addition, transmission dynamics within swine populations—such as herd density, housing conditions, and co-infection patterns—differ substantially from those in human populations [4]. Therefore, directly applying conclusions from human influenza studies to SIV research may lead to misinterpretation of immune protection mechanisms or vaccine efficacy.
At present, inactivated and live attenuated vaccines are widely used in swine populations. However, vaccine effectiveness varies markedly among regions and herds, largely due to antigenic drift, genetic reassortment, and region-specific lineage evolution of SIV [5]. Moreover, natural infection and vaccination differ significantly in their ability to induce mucosal immunity, local immune memory, and tissue-resident T cell responses [6], making accurate evaluation of cross-protective immunity an urgent challenge.
Accordingly, the establishment of appropriate animal models is essential for investigating SIV pathogenesis, host immune responses, transmission characteristics, and vaccine efficacy [7]. However, no single animal model fully recapitulates the pathological and immunological features of natural infection in pigs. Mouse models are genetically tractable and suitable for mechanistic studies but poorly reproduce porcine pulmonary mucosal immunity. Ferret and guinea pig models are valuable for transmission research but differ substantially from pigs in immune system architecture. Non-human primates share closer immune kinetics with humans but are limited by high cost and ethical constraints. In contrast, the pig model most faithfully reflects immune responses and disease course in the natural host, yet its use is restricted by experimental complexity and expense.
This leads to a central issue in current research: systematic differences in immune responses among animal models can directly influence the evaluation of vaccine efficacy and the interpretation of immune mechanisms.
Therefore, a comprehensive understanding of inter-model differences in innate immunity, humoral immunity, mucosal immunity, and tissue-resident T cell formation is critical for rational model selection and for improving the interpretability and translational relevance of vaccine studies. This review systematically compares commonly used SIV animal models from the perspective of immune response heterogeneity and discusses their implications for vaccine development and applied research.
2. Biological Characteristics and Epidemiological Shifts in SIV
2.1. Genome Architecture of SIV and Host Receptor Characteristics
Swine influenza virus (SIV) is an influenza A virus (IAV) with an eight-segmented, negative-sense RNA genome. Each segment encodes distinct structural or functional proteins, including haemagglutinin (HA), neuraminidase (NA), and nucleoprotein (NP) [3]. Porcine respiratory epithelial cells express both α-2,3- and α-2,6-linked sialic acid receptors, enabling pigs to be infected by influenza viruses of avian as well as human origin; consequently, pigs are often regarded as a “mixing vessel” for influenza virus reassortment. This receptor distribution provides a molecular basis for co-infection by viruses from different sources and subsequent genome segment reassortment, thereby creating key preconditions for the emergence and onward spread of novel influenza subtypes.
2.2. Epidemiological Dynamics of Swine Influenza Virus
SIV circulates predominantly as the H1N1, H1N2, and H3N2 subtypes, and the molecular evolutionary lineages and antigenic properties of circulating strains continue to change across regions worldwide. Following the 2009 influenza A(H1N1) pandemic, triple-reassortant viruses spread rapidly on a global scale and displaced multiple previously prevailing lineages [8]. In recent years, alongside increasing inter-regional movements of swine and the globalisation of pig production systems, the H1N1, H1N2, and H3N2 subtypes have remained in sustained circulation in China and globally. Ongoing emergence of novel reassortant genotypes and antigenic drift variants has further complicated vaccine strain selection, resulting in progressively greater challenges in matching vaccine strains to field viruses [9,10,11,12,13,14,15,16].
3. Establishment of SIV Animal Infection Models and Evaluation Parameters
The primary objective in establishing animal models of swine influenza virus (SIV) infection is to reproduce, to the greatest extent possible, the infection kinetics, histopathological progression, and immune-response trajectories observed in the natural host [17]. The quality of model construction directly determines the practical relevance of experimental outcomes. This section describes three key dimensions: the critical parameters for simulating natural infection, the evaluation endpoints, and the applicability of different animal models.
3.1. Key Parameters for Modelling Natural Infection
To generate experimental data with clinical relevance, the construction of animal models must carefully control infection routes and inoculation doses in order to minimise artefacts introduced by conventional experimental approaches [18].
(1) In swine populations, the natural transmission of SIV mainly occurs through aerosols, inhalation of respiratory droplets, and short-range contact. Although traditional high-dose intranasal inoculation or intratracheal administration can ensure high infection rates, these approaches often result in direct viral deposition in the deep lower respiratory tract and can induce non-physiological, acute severe pneumonia. Such artificial exposure may obscure the early steps of viral adhesion to and invasion of the respiratory mucosa. Therefore, to more accurately mimic natural infection, an ideal model should prioritise aerosol nebulization or environmentally simulated contact exposure [19]. These approaches better reflect low-dose, multi-site exposure and can more accurately capture the initial colonisation pattern along the respiratory epithelium.
(2) Concordance of viral replication and disease kinetics: A qualified model should reproduce, to a reasonable extent, the temporal dynamics observed during natural infection in pigs. Under natural conditions, infection typically follows a defined timeline: clinical signs emerge at approximately 2–3 days post infection (dpi), pulmonary lesions and viral shedding peak at 5–7 dpi, and viral clearance is largely completed by 10–14 dpi. If viral clearance occurs too rapidly (as seen in some non-adapted mouse strains) or if infection persists excessively, misinterpretation of vaccine durability or antiviral half-life may occur [20].
(3) Comprehensiveness of immune responses: Protective immunity against SIV depends on a coordinated network of systemic and mucosal immunity. An appropriate model must be capable of inducing and detecting key immunological readouts, including humoral immunity (serum HI and neutralising antibodies), mucosal immunity (nasal and pulmonary sIgA), and cellular immunity (Th1-associated cytokines and cytotoxic T lymphocyte responses) [21,22,23]. Reliance on a single serological parameter is often insufficient to accurately evaluate vaccines targeting this predominantly mucosal pathogen.
To avoid overgeneralization, it should be emphasised that peak viral load and replication kinetics are not fixed parameters but vary substantially depending on viral strain, inoculation dose, infection route, and host species. Highly pathogenic or host-adapted strains may replicate more rapidly and reach peak viral loads earlier, whereas low-dose aerosol exposure typically results in delayed replication and more spatially restricted infection. Likewise, compared with contact-based transmission models, intratracheal inoculation tends to artificially synchronise lower respiratory tract infection. Therefore, these variables should be carefully controlled and explicitly reported when interpreting cross-model comparisons.
3.2. Evaluation Parameters for Natural Infection-Mimicking Models
A standardised SIV animal model should demonstrate high stability and reproducibility across three key dimensions: pathological injury, viral replication kinetics, and immune phenotype [18].
(1) Pathological characteristics
Model animals should exhibit gross and microscopic lesions comparable to those observed in naturally infected pigs [24]. Gross pathology: Typical findings include well-demarcated purple-red consolidation of affected lung lobes, most commonly involving the cardiac and apical lobes, often accompanied by thickening of the alveolar septa. Histopathology: Characteristic necrotizing bronchitis should be evident, including necrosis and sloughing of tracheal/bronchial epithelial cells, neutrophilic exudation, and prominent peribronchiolar lymphocytic cuffing.
(2) Viral replication kinetics
The spatiotemporal distribution of viral load is a critical indicator of model sensitivity [25]. In a standard SIV model, viral titres typically peak at 1–2 days post infection, leading to viral pneumonia, and viral loads in lung tissue should be significantly higher than those in the upper respiratory tract. In addition, the model should exhibit differential replication efficiency among major subtypes (H1N1, H1N2, H3N2), thereby accurately reflecting differences in pathogenicity and immune evasion among circulating strains.
(3) Immune response profiling
Evaluation metrics should encompass local, systemic, and cellular immunity. For local immunity, virus-specific IgA can be detected in the nasal cavity (nasal wash/swab) and lower respiratory tract (bronchoalveolar lavage, BAL) following infection or immunisation in pigs, with IgA serving as a key isotype at mucosal surfaces [21]. For systemic immunity, serum hemagglutination inhibition (HI) antibody titres are commonly used in swine influenza studies and vaccine assessments as indicators of humoral responses [26]. For cellular immunity, CD4^+^ and CD8^+^ T cell responses can be evaluated in blood, BAL, and tracheobronchial lymph nodes, together with key cytokine expression profiles to characterise immune phenotypes and their correlation with protection [27].
3.3. Core Advantages and Limitations of Model Selection
Given the complexity of SIV infection and the fact that no single model can fully recapitulate the pathological features observed in pigs, the selection of an appropriate animal model requires consideration of multiple factors. A tiered selection strategy is therefore essential, and models should be chosen according to their alignment with the research objective, biological relevance to the natural host, and practical experimental constraints [28]. The major advantages and limitations of commonly used animal models in SIV research are summarized below, with detailed comparisons presented in Table 1.
(1)Mouse model: preferred platform for fundamental mechanistic studies
Mice possess a well-defined genetic background, abundant immunological reagents, and high experimental tractability [20]. Their primary advantage lies in the ability to dissect virus–host interactions through gene knockout and transgenic approaches, making them a core platform for early drug screening and mechanistic investigations [20]. However, it should be noted that not all swine-origin H1N1 strains replicate efficiently in mice without prior adaptation. Some strains exhibit cross-species replication barriers and often require serial passaging to acquire adaptive mutations for stable infection. This adaptation process may alter viral pathogenicity and thus deviate from the behaviour observed in the natural host. In addition, the distribution of respiratory epithelial receptors in mice differs from that in pigs, resulting in lower susceptibility to natural infection. Mouse lung pathology also poorly mimics the diffuse lesions seen in pigs, and aerosol transmission is relatively inefficient, limiting its utility for modelling natural transmission dynamics.
(2)Ferret model: gold standard for influenza transmission studies
Ferrets exhibit respiratory receptor distributions highly similar to those of humans and pigs and are highly susceptible to influenza virus replication and aerosol transmission. Therefore, they are widely regarded as an ideal model for studying transmission dynamics, viral adaptive evolution (e.g., antigenic drift and reassortment), and cross-species transmission potential [29]. Despite its unique value in evaluating viral spread and replication efficiency, the ferret model is constrained by high costs, limited availability of immunological reagents, and relatively small sample sizes. These limitations restrict its broad application in vaccine immunology and necessitate complementary use with other models [30].
(3)Guinea pig model: an intermediate option for drug screening and transmission studies
Guinea pigs display efficient aerosol transmission characteristics and are suitable for investigating viral spread and environmental stability [31]. However, guinea pigs generally lack typical influenza-like clinical signs, and their pulmonary pathological changes are relatively mild, preventing full recapitulation of the natural infection process in pigs. In addition, the limited availability of immunological reagents in this model constrains in-depth cellular immune profiling and mechanistic studies. Consequently, the guinea pig model is more appropriate as a complementary system for transmission dynamics alongside the ferret model rather than for comprehensive vaccine immunogenicity evaluation [32,33].
(4)Pig model: the gold standard closest to the natural host
As the natural host of SIV, pigs represent the gold standard model that most closely reflects natural infection. Pigs can faithfully reproduce clinical manifestations such as fever, lung injury, and respiratory distress, and their viral replication kinetics and host immune responses closely mirror those observed in field infections. Therefore, the pig model is the preferred system for evaluating vaccine protective efficacy, clinical outcomes, viral clearance, and potential immunopathological effects [18]. However, the pig model is associated with high experimental costs, strict biosecurity requirements, and longer study durations, which reduce its flexibility for large-scale screening and early mechanistic exploration compared with mice and ferrets.
(5)Non-human primate models: advanced systems for cross-species risk assessment
Non-human primates (e.g., rhesus macaques and cynomolgus macaques) possess immune systems highly similar to humans and are therefore valuable for assessing the cross-species transmission potential of SIV genotypes and their possible risks to human health [34,35]. Nevertheless, the high cost, stringent ethical oversight, and limited sample sizes restrict their use primarily to high-risk genotypes or advanced studies with direct public health relevance.
In addition to these in vivo systems, in vitro and ex vivo models provide important complementary tools. For example, primary porcine respiratory epithelial cells and precision-cut lung slices can simulate local host responses and tissue-level infection dynamics within a shorter experimental timeframe, serving as an intermediate bridge between in vitro mechanisms and in vivo models [36]; Cell lines such as MDCK and PK-15 are widely used for viral replication kinetics and antiviral drug screening, substantially reducing reliance on animal experiments [37]. Although these complementary systems cannot replace animal infection models, they provide efficient and controllable platforms for mechanistic exploration, virus–host interaction studies, and early therapeutic evaluation.
At present, no single animal model can fully recapitulate the complete biological characteristics of pigs as the natural host of swine influenza virus (SIV). The strengths and limitations of each animal model vary depending on the specific research objective. Therefore, reliance on a single model is insufficient to comprehensively cover key aspects of SIV research, including viral replication kinetics, histopathological changes, immune regulatory mechanisms, and vaccine immunogenicity.
To ensure both the comprehensiveness and accuracy of experimental findings, selecting the most appropriate animal model based on the study objective—or adopting a multi-model validation strategy—has become a critical approach in SIV pathogenesis research and vaccine evaluation. This section provides the methodological framework for the subsequent systematic comparisons of pathology, viral dynamics, and immune responses.
4. Animal Models of SIV Infection: Differences in Pathological Changes and Immune Responses
The strong interspecies transmission potential of SIV, together with the high heterogeneity among viral strains, results in pronounced differences across animal models in their ability to mirror the natural course of infection. These differences are particularly evident in patterns of tissue injury, anatomical sites of viral replication, magnitude of immune responses, and capacity for aerosol transmission, with each model presenting distinct strengths and limitations [38,39,40]. This section compares, in a model-by-model manner, the pathological manifestations and immunological features of the five most commonly used systems in current research—mice, ferrets, guinea pigs, pigs, and non-human primates—to inform subsequent vaccine evaluation and mechanistic studies of pathogenesis.
4.1. Mouse Models: High Genetic Manipulability but Substantial Immunological Limitations
As a classical small-animal experimental system, mice are widely used to investigate influenza virus pathogenesis and host immune responses [41,42,43]. Numerous studies have shown that swine-origin H1N1 viruses or recombinant influenza viruses from different sources can replicate efficiently in mice and reproducibly induce acute respiratory disease dominated by pulmonary injury, with relatively consistent pathological manifestations [44]. Following infection, gross lung lesions are commonly observed, characterised by varying degrees of pulmonary consolidation and focal or diffuse pneumonic foci; lesion severity correlates closely with viral replication levels in the lungs and with lethality in severe cases [45].
Histopathological examination indicates that murine influenza infection most frequently manifests as acute bronchitis or bronchiolitis accompanied by acute interstitial pneumonia (or bronchointerstitial pneumonia–like changes). Infected lungs show epithelial damage and sloughing within the airways, with cellular debris and exudates present in some lumina. Thickening of alveolar septa and prominent infiltration of inflammatory cells into alveolar spaces and the interstitium are also evident, typically dominated by neutrophils and macrophages, and may be accompanied by pulmonary oedema, haemorrhage, and atelectasis. Notably, infection with highly pathogenic or lethal strains or recombinant viruses often produces more pronounced necrotising bronchitis or bronchiolitis, together with severe alveolitis and pulmonary oedema as dominant pathological features. By contrast, infection with certain swine-derived 2009/H1N1 isolates tends to produce lesions characterised primarily by acute interstitial pneumonia, alveolar wall thickening, inflammatory infiltrates, and haemorrhage or atelectasis, which commonly become prominent several days post infection (e.g., 3–5 days post infection).
At the immunological level, influenza virus infection in mice rapidly induces acute inflammatory responses driven largely by innate immune activation. During the early stages of infection, type I interferon responses in the lung—particularly IFN-β—are markedly upregulated, accompanied by increased expression of pro-inflammatory cytokines such as TNF-α and multiple chemokines, including MIP-1α and MIP-2 [46,47,48,49]. Production of these mediators correlates closely with pulmonary viral loads and constitutes a major molecular basis for the development of acute inflammation and immunopathological lung injury. Concomitantly, large numbers of neutrophils are rapidly recruited to lung tissue and alveolar spaces and, together with activated alveolar macrophages, contribute to inflammatory amplification and tissue damage, forming the principal cellular components of early antiviral immunity and immunopathology in murine influenza models.
Multiple murine influenza studies further indicate that adaptive immunity progressively contributes to viral clearance as infection advances. During the mid to late stages of infection, virus-specific immune cell responses and cytokine changes can be detected in lung tissue and bronchoalveolar lavage fluid, while certain anti-inflammatory or immunoregulatory mediators, such as IL-10, are upregulated at specific time points, suggesting that negative-feedback regulatory mechanisms are gradually engaged following intense inflammatory responses [50].
Overall, murine influenza models are characterised by pronounced inflammatory reactions and innate immune cell infiltration accompanied by the participation of immunoregulatory pathways, providing an important experimental basis for dissecting influenza virus pathogenic mechanisms [51]. Representative gross and histopathological lung lesions observed in mice following influenza virus infection are shown in Figure 1 [52], which systematically illustrates typical patterns of pulmonary injury induced by different viral strains in this model.
4.2. Ferret Models: Human-like Transmission Patterns but Incomplete Pathology
Ferrets exhibit respiratory tract sialic acid receptor distributions highly similar to those of humans and are therefore among the most widely used small-animal models for studying the transmissibility and pathogenicity of human influenza viruses [53]. Following infection with swine-origin H1N1 or the 2009 pandemic H1N1 (pH1N1) virus, ferrets typically develop upper respiratory tract signs, including fever, sneezing, coughing, rhinorrhoea, and substantial nasal viral shedding [54]. Previous studies have further demonstrated that, in combined nasal and tracheal infection models, H5N1, pH1N1, and seasonal influenza viruses exhibit markedly different respiratory pathogenic dynamics, with characteristic variations in the timing of lesion development, the intensity of inflammation, and the anatomical sites involved [55].
Systematic comparisons of three representative influenza viruses—seasonal H3N2, 2009 pH1N1, and highly pathogenic H5N1—have shown distinct patterns of tissue tropism and lesion severity within the ferret respiratory tract. Replication of H3N2 is most restricted, occurring primarily in the nasal epithelium and causing only mild mucosal injury; pH1N1 replicates throughout the respiratory tract and induces moderate bronchopneumonia, whereas H5N1 infection is concentrated in alveolar regions, resulting in the most severe lower respiratory tract damage.
Histological sections obtained at 4 days post infection following inoculation with equivalent viral doses clearly illustrate these differences. In H3N2-infected ferrets, only a small number of antigen-positive cells are detected in the nasal cavity, with minimal bronchial and alveolar inflammation. In contrast, pH1N1 infection is associated with more prominent viral antigen expression and inflammatory infiltration in bronchial epithelium, bronchioles, and portions of the alveoli, whereas H5N1 viral antigen is widely distributed in the lower respiratory tract, accompanied by pronounced alveolitis, necrosis, and haemorrhage. These pathological changes correspond closely to the anatomical distribution of viral antigen and represent typical manifestations of virus-specific pathogenic features in the ferret model. As illustrated in Figure 2, this study provides a schematic summary of the spatiotemporal lesion patterns induced by the three representative strains in the ferret respiratory trac.
From an immunological perspective, ferrets typically develop detectable humoral immune responses, including haemagglutination inhibition and neutralising antibodies, approximately one week after natural influenza virus infection [56]. Concurrently, cellular immune activity characterised by IFN-γ-associated responses can be observed in infected respiratory tissues [57]. Overall, the kinetics of antibody production and patterns of local cellular immunity in ferrets closely resemble those observed in humans, underscoring their value for investigating influenza infection and immune responses [58].
Notably, repeated infection or prior immunisation in ferrets can result in immune imprinting effects similar to those described in humans, whereby exposure history substantially alters subsequent antibody specificity, cross-reactivity, and protective efficacy following reinfection or vaccination [59,60]. These findings indicate that primary exposure to a given viral strain can durably shape immune response patterns against heterologous viruses, making ferrets a valuable model for studying influenza immune memory and cross-protection.
4.3. Guinea Pig Models: Specialised Systems for Airborne Transmission Studies
Guinea pigs are highly susceptible to a wide range of human-, swine-, and avian-origin influenza viruses, yet typically develop only mild clinical signs, most often limited to transient weight loss and reduced activity [32,61,62,63]. In this context, guinea pig models show relatively limited sensitivity for assessing virulence, and pulmonary pathology is usually mild to moderate in severity. As illustrated in Figure 3, histological examination following infection commonly reveals bronchointerstitial pneumonia, mild degeneration of alveolar epithelial cells, interstitial oedema, and modest inflammatory cell infiltration dominated by lymphocytes and histiocytes, whereas extensive haemorrhagic pneumonia or classical diffuse alveolar damage is uncommon in this model.
From an immunological perspective, guinea pigs often mount relatively effective innate immune barriers following influenza virus infection. Previous studies have indicated that the complement system plays an important role in viral clearance and regulation of inflammatory responses, which may contribute to the relatively low mortality observed even after infection with highly pathogenic influenza viruses [64]. By contrast, pigs are more prone to developing severe pneumonia following high-dose SIV infection or secondary bacterial co-infection, underscoring substantial interspecies differences in immune response magnitude and immunopathological outcomes.
On the basis of these pathological and immunological features, guinea pig models are particularly well suited for comparing the pathogenicity of influenza viruses from different host origins within a single host species and for investigating aerosol transmission efficiency and early innate immune responses. However, because pulmonary lesions are comparatively mild and the availability of immunological reagents remains limited, guinea pigs retain notable constraints in faithfully modelling the immune responses associated with natural SIV infection in pigs.
4.4. Pig Models: The Natural Host, Capturing Complete Pathogenesis and Immunological Complexity
As the natural host of SIV, pig models most closely reflect field outbreaks with respect to both pathology and immune responses. Following SIV infection, pigs can reliably develop acute respiratory disease dominated by pulmonary injury, and the core pathological features are highly consistent across different viral strains and experimental conditions. Gross examination typically reveals characteristic dark red to purple areas of pulmonary consolidation, distributed in a multifocal or coalescing pattern, most commonly involving the cranial and middle lung lobes. The severity of these lesions correlates closely with pulmonary viral replication levels and the clinical severity of disease.
Histopathological evaluation indicates that lesions are characterised primarily by necrotising bronchiolitis and bronchointerstitial pneumonia. Typical changes include degeneration, necrosis, and sloughing of bronchiolar epithelial cells, with cellular debris and proteinaceous exudates within the lumen, accompanied by prominent peribronchiolar lymphocytic cuffing. In some regions, varying degrees of alveolar collapse and interstitial inflammatory cell infiltration can be observed. Acute lesions usually peak at approximately 3–5 days post infection and subsequently begin to resolve, with progressive reparative changes characterised by regenerative hyperplasia of the bronchial epithelium and relative attenuation of inflammation. Overall, although acute pulmonary injury can be marked, the pathological process is generally self-limiting and constitutes a typical pathological spectrum of SIV infection in the porcine lung. Representative pathological findings reported in prior studies are shown in Figure 4, which summarises gross lesions and histological injury patterns in pigs following SIV infection [65].
From an immunological perspective, SIV infection induces a highly coordinated set of immune responses in the porcine lung, characterised by concomitant innate immune activation, enhancement of adaptive immunity, and engagement of immunoregulatory mechanisms. During the early phase of infection, multiple innate and pro-inflammatory mediators are markedly upregulated in lung tissues, including IFN-α, IL-6, IFN-γ, and IL-12. The levels of these cytokines correlate positively with pulmonary viral loads and the extent of tissue injury and are important drivers of acute inflammation and clinical manifestations. Against this background, adaptive immunity is rapidly activated. Flow cytometric analyses of mononuclear cells isolated from lung tissues, bronchoalveolar lavage fluid, and tracheobronchial lymph nodes at day 3 and day 6 after SIV infection have shown marked enrichment and activation of multiple immune cell subsets at sites of infection, including CD4^+^ and CD8^+^ T lymphocytes, cytotoxic T cells, γδ T cells, natural killer cells, and cell populations enriched for dendritic cells, suggesting that SIV infection can rapidly elicit T cell-mediated adaptive immune responses during the acute phase [66].
In parallel, antigen-specific IgA responses can be detected in lung tissue and bronchoalveolar lavage fluid, indicating an important contribution of mucosal humoral immunity to viral clearance. Notably, alongside heightened inflammation, immunoregulatory components are also upregulated, including increased proportions of FoxP3^+^ regulatory T cells and enhanced expression of anti-inflammatory cytokines such as IL-10 and TGF-β. These findings suggest that negative-feedback mechanisms are simultaneously engaged during antiviral responses. Collectively, these immunological features indicate that SIV infection in pigs does not simply induce uncontrolled inflammation but rather involves a dynamic balance between inflammatory injury and immunoregulation [67,68].
Under conditions where the vaccine strain and the challenge strain are antigenically mismatched, non-neutralising antibodies induced against heterologous viruses may exacerbate pulmonary inflammation upon re-exposure and even aggravate lesion severity. This phenomenon is defined as vaccine-associated enhanced respiratory disease (VAERD) [69,70]. The existence of VAERD indicates that vaccine evaluation in the pig model should not focus solely on protection rates and viral clearance but should also systematically assess potential immunopathological risks. Because pigs are the natural host of SIV, their respiratory immune architecture can faithfully reflect immune dysregulation under antigen mismatch conditions, giving this model unique value for identifying such risks.
However, despite the high biological relevance of the pig model, its outbred nature may introduce substantial inter-individual variability. The high polymorphism of the swine major histocompatibility complex (SLA) can influence antigen presentation and T cell response magnitude, thereby contributing to variability in vaccine responsiveness among animals [71]. Therefore, experimental design should minimise genetic heterogeneity through source standardisation, age matching, and appropriate stratified statistical analysis.
4.5. Non-Human Primate Models: Immunological Relevance and Ethical and Economic Constraints
Commonly used non-human primate models for influenza include rhesus macaques and cynomolgus macaques. Studies have shown that infection of non-human primates with the 2009 swine-origin H1N1 virus generally results in pulmonary lesions dominated by peribronchiolar inflammation and mild-to-moderate interstitial pneumonia, rather than extensive diffuse alveolar damage, indicating relatively moderate pathogenicity consistent with the clinical features observed in human infections, as illustrated in Figure 5 [72]. By contrast, in non-human primate models, highly pathogenic influenza viruses such as the 1918 H1N1 strain can induce severe respiratory disease characterised by pronounced diffuse alveolar damage, pulmonary oedema, and more severe inflammatory pathology, closely reflecting viral virulence [73,74].
At the level of immune responses, non-human primates exhibit strong similarities to humans and pigs in cellular immune repertoires, Fc receptor composition, and cytokine networks, conferring clear advantages in immunological relevance. Analyses of immune gene expression have demonstrated that sustained and intense innate immune activation during early infection—particularly marked upregulation of type I interferons and pro-inflammatory mediators—is strongly associated with severe pulmonary injury and poor outcomes, whereas more moderate inflammatory responses coupled with timely viral clearance are typically linked to milder clinical and pathological manifestations [75]. These patterns have been repeatedly confirmed in comparative studies of highly pathogenic versus low-pathogenic influenza viruses and are consistent with the immunopathological features observed in severe human influenza cases and severe SIV infections in pigs.
Overall, the principal value of non-human primate models lies in the validation of the safety and key immunological endpoints of high-risk candidate vaccines, monoclonal antibodies, or antiviral interventions, rather than in routine screening of SIV pathogenicity or large-scale immunogenicity assessments. Their high costs, stringent ethical constraints, and limited sample sizes make it impractical for these models to replace pigs as the central experimental system for studying natural SIV infection and population-level immune protection.
4.6. Cross-Model Comparative Analysis: Differences in Pathology, Viral Kinetics, and Immune Architecture
Although each animal model provides unique value in SIV research, interpretation of their results must be grounded in systematic cross-model comparisons across key biological dimensions rather than isolated, model-specific descriptions, with detailed comparisons presented in Figure 6 and Table 2.
In terms of pathological severity, clear hierarchical differences are observed among species, and these patterns are influenced by viral subtype and virulence. The pig model most faithfully reproduces the characteristic necrotizing bronchiolitis and bronchointerstitial pneumonia seen in natural infection, with lesion distribution highly consistent with field cases in swine herds. In the ferret model, pathological changes show subtype-dependent variation: highly pathogenic strains can induce pronounced lower respiratory tract inflammation, whereas seasonal-like strains are more often confined to the upper respiratory tract. Mice commonly exhibit diffuse inflammatory infiltration; however, the lesion distribution does not fully match that of pigs, particularly with respect to bronchiolar necrosis and alveolar structural damage. Guinea pigs typically develop only mild bronchointerstitial changes with limited tissue destruction. The pathological severity observed in non-human primates generally falls between that of pigs and ferrets and is closely associated with viral virulence and inoculation dose.
Regarding viral replication kinetics, systematic differences also exist among models, and the timing of peak replication and viral load is influenced by viral strain, inoculation dose, and infection route. In pigs, the replication curve closely mirrors the timeline of field infection, showing a well-defined viral peak and a predictable clearance phase, and a stable shedding window can usually be detected in nasal swabs. Ferrets support highly efficient upper respiratory replication and exhibit stable aerosol transmission, making them a key model for studying transmission dynamics. Mice typically require host-adapted viral strains to achieve efficient replication, and the replication capacity of different swine-origin H1N1 isolates varies substantially in this model. Although guinea pigs support viral transmission, the correlation between clinical signs and viral load is relatively weak, making it difficult to infer true replication intensity from clinical presentation alone. Non-human primate models display replication efficiency and clearance kinetics that approximate human infection, but their experimental scalability is limited.
At the level of immune architecture, the differences among models are even more pronounced. The pig model can simultaneously reproduce mucosal IgA responses, CD4^+^/CD8^+^ T cell activation, γδ T cell participation, and immunoregulatory feedback mechanisms, thereby forming a relatively complete respiratory mucosal immune network. Ferrets exhibit human-like antibody kinetics and immune imprinting phenomena, but the depth of cellular immune characterisation remains limited. Mice are well suited for dissecting innate signalling pathways, inflammatory cascades, and cytokine networks; however, the complexity of their respiratory mucosal immune structure is lower than that of pigs. In guinea pigs, the relative scarcity of species-specific immunological reagents restricts fine-scale immune phenotyping. Non-human primates possess high translational relevance in immune architecture, allowing for observation of tissue-resident T cell formation and long-term immune memory; however, ethical and cost constraints limit their large-scale application.
Although a tiered, multi-model collaborative framework offers clear theoretical advantages, its practical implementation must be aligned with specific research objectives. Taking the evaluation of a hypothetical universal influenza vaccine candidate as an example, a stepwise experimental pathway can be constructed. First, at the in vitro stage, MDCK cells can be used for rapid screening of viral replication inhibition and preliminary antigen stability assessment, enabling early elimination of candidates with insufficient replication control or unstable antigen conformation [76]. Subsequently, the mouse model can be employed to dissect vaccine-induced innate immune activation, including interferon signalling, inflammatory cytokine profiles, and early CD8^+^ T cell expansion, thereby establishing a mechanistic immunogenicity foundation [77,78]. Building on these findings, the ferret model can be used to evaluate the vaccine’s ability to block aerosol transmission and shorten the viral shedding window, thereby assessing its potential population-level impact [79]. Finally, protective efficacy, safety, and durability of immunity should be validated in pigs to confirm real-world translational relevance in the natural host [79]. This tiered strategy progressively strengthens the evidence base while controlling cost and risk and reduces interpretive bias associated with reliance on a single model.
Taken together, these dimensions demonstrate that the various models exhibit complementary characteristics in pathological distribution, viral replication patterns, transmission efficiency, and immune complexity. No single model can fully capture the complete biological spectrum of natural host infection. Therefore, a multi-model integration strategy is not merely a methodological supplement but a necessary biological framework for reducing interpretive bias and improving the translational reliability of SIV vaccine evaluation.
5. Current Challenges and Future Perspectives
5.1. Major Challenges in Current Research
(1) Model-specific differences drive variability in validation outcomes
Existing animal models differ intrinsically from target hosts with respect to infection routes, receptor distribution, and immune composition. Immune response magnitudes vary not only among species but also among strains within the same model. Such interspecies and inter-individual variability complicates direct comparison of vaccine or intervention performance across models and limits extrapolation to target animals or field conditions.
(2) Biosafety constraints restrict critical validation studies
Research involving highly pathogenic or emerging agents typically requires biosafety level 3 (BSL-3) or higher containment. The availability, geographic distribution, and operational costs of high-containment facilities impose practical limitations, which can prolong study timelines and restrict systematic evaluation of certain models and strategies.
(3) Cost and ethical considerations limit large-animal studies
Large-animal models offer advantages in approximating natural infection and immune responses; however, high husbandry and experimental costs, stringent ethical review requirements, and limited sample sizes render them unsuitable for early-stage screening or large-scale validation. By contrast, murine models are economically efficient but possess simplified immune architectures that cannot fully substitute for target hosts.
(4) Natural infection processes remain incompletely reproduced
Marked differences among species in tissue tropism, immune regulation, and disease progression mean that most existing systems remain approximations and cannot comprehensively mimic natural infection or complex immunological outcomes. This limitation underscores the difficulty of relying on any single model as a sufficient predictor of vaccine performance.
5.2. Future Directions and Priorities
Future development of animal models is expected to shift from descriptive characterisation of infection outcomes toward finer-resolution analyses of immune processes and tissue microenvironments. Integration of multi-omics technologies, advanced in vitro platforms (such as organoids and organ-on-chip systems), and computational immunology approaches may partially mitigate the limitations of in vivo models. A more practical strategy is the establishment of tiered, multi-model evaluation frameworks: murine systems for early screening, intermediate-sized animals for calibration of immune response features, and eventual validation in target hosts. Coupled with mechanism-oriented integration of immunological endpoints, such approaches may reduce uncertainty arising from model differences and enhance the predictability and reproducibility of vaccine development pipelines.
6. Conclusions
Overall, the infection and immune processes of swine influenza virus (SIV) exhibit marked host dependence. Differences among animal models in patterns of pathological injury, immune response architecture, and transmission characteristics determine their functional positioning within the research framework. This host dependence is reflected not only in histopathological lesion distribution but also in key immunological dimensions, including thresholds of innate immune activation, the integrity of mucosal immune structures, and the capacity to generate tissue-resident T cells.
The mouse model offers clear advantages for mechanistic dissection and early-stage screening; however, its immune architecture deviates from that of the natural host. Ferret and guinea pig models are more suitable for studying transmission dynamics and early infection events, but they provide limited fidelity in recapitulating the natural disease course and immune complexity of swine influenza. In contrast, the pig model most comprehensively reproduces pulmonary pathology and the coordinated innate–adaptive immune responses observed in the natural host, making it an indispensable core system for vaccine efficacy evaluation and immunological mechanism studies. It also serves as a critical platform for identifying potential immunopathological risks under antigen mismatch conditions. Non-human primate models are primarily used for safety and immunological validation of high-risk strains or intervention strategies and provide complementary value in cross-species risk assessment and translational research.
Therefore, no single “universal model” represents an optimal solution for SIV research. The differences among models should not be viewed as limitations but rather as reflections of the biological diversity inherent in virus–host interactions across species. According to the research stage and specific scientific questions, a hierarchical, multi-model coordinated strategy can reduce bias arising from model-specific constraints and improve the interpretability and translational relevance of vaccine evaluation. This mechanism-oriented, target host-anchored model selection framework is likely to represent a more robust path for future SIV vaccine development and pathogenesis research, while also providing an experimental basis for risk identification and forward-looking assessment of emerging swine-origin reassortant strains.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zheng X. Cao F. Yu Y. Yu X. Zhou Y. Cheng S. Qiu X. Ao L. Yang X. Sun Z. The Ongoing Epidemics of Seasonal Influenza a(H 3N 2) in Hangzhou, China, and Its Viral Genetic Diversity Viruses 20251752610.3390/v 1704052640284969 PMC 12031544 · doi ↗ · pubmed ↗
- 2Sun H. Xiao Y. Liu J. Wang D. Li F. Wang C. Li C. Zhu J. Song J. Sun H. Prevalent Eurasian Avian-like H 1N 1 Swine Influenza Virus with 2009 Pandemic Viral Genes Facilitating Human Infection Proc. Natl. Acad. Sci. USA 2020117172041721010.1073/pnas.192118611732601207 PMC 7382246 · doi ↗ · pubmed ↗
- 3Ma W. Loving C.L. Driver J.P. From Snoot to Tail: A Brief Review of Influenza Virus Infection and Immunity in Pigs J. Immunol.20232111187119410.4049/jimmunol.230038537782856 PMC 10824604 · doi ↗ · pubmed ↗
- 4Pittman Ratterree D.C. Dass S.C. Ndeffo-Mbah M.L. Mechanistic Models of Influenza Transmission in Commercial Swine Populations: A Systematic Review Pathogens 20241374610.3390/pathogens 1309074639338936 PMC 11434764 · doi ↗ · pubmed ↗
- 5Ma W. Swine Influenza Virus: Current Status and Challenge Virus Res.202028819811810.1016/j.virusres.2020.19811832798539 PMC 7587018 · doi ↗ · pubmed ↗
- 6Holzer B. Martini V. Edmans M. Tchilian E. T and B Cell Immune Responses to Influenza Viruses in Pigs Front. Immunol.2019109810.3389/fimmu.2019.0009830804933 PMC 6371849 · doi ↗ · pubmed ↗
- 7Robinson N.B. Krieger K. Khan F.M. Huffman W. Chang M. Naik A. Yongle R. Hameed I. Krieger K. Girardi L.N. The Current State of Animal Models in Research: A Review Int. J. Surg.20197291310.1016/j.ijsu.2019.10.01531627013 · doi ↗ · pubmed ↗
- 8Salvesen H.A. Whitelaw C.B.A. Current and Prospective Control Strategies of Influenza a Virus in Swine Porc. Health Manag.202172310.1186/s 40813-021-00196-033648602 PMC 7917534 · doi ↗ · pubmed ↗