β-Xylosidase Overexpression Alters Pectin and Cellulose Distribution and Modulates Blast Disease Resistance in Rice
Takashi Ohara, Taichi Watanabe, Ryuya Bamba, Atsuko Nakamura, Hiroaki Iwai

TL;DR
Overexpressing a β-xylosidase gene in rice changes cell wall composition and affects resistance to blast disease in a tissue-specific way.
Contribution
This study reveals organ-specific effects of β-xylosidase overexpression on cell wall composition and blast resistance in rice.
Findings
Leaf pectin reduction correlates with increased blast susceptibility in rice leaves.
Leaf sheaths with increased pectin and cellulose show enhanced resistance to fungal penetration.
Hemicellulose changes have a secondary role in blast resistance compared to pectin and cellulose.
Abstract
Plant cell walls provide structural integrity and defense against biotic and abiotic stresses. In rice (Oryza sativa), xylan is the major hemicellulose, and β-xylosidase hydrolyzes xylan by removing xylose residues from non-reducing ends. We analyzed a transgenic rice line (OsXylGH3-1-FOX) that constitutively overexpresses a GH3-family β-xylosidase (Os03g0749100) under the maize ubiquitin promoter. Following inoculation with M. oryzae, OsXylGH3-1-FOX leaves exhibited increased lesion numbers and disease indices, indicating reduced resistance, whereas leaf sheaths showed fewer fungal penetrations, suggesting enhanced resistance. To investigate these organ-specific responses, we quantified cell wall components. In leaves, xylose and arabinose decreased by ~33%, and galacturonic acid (pectin) by ~50%. In leaf sheaths, xylose and arabinose were unchanged, while galacturonic acid and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
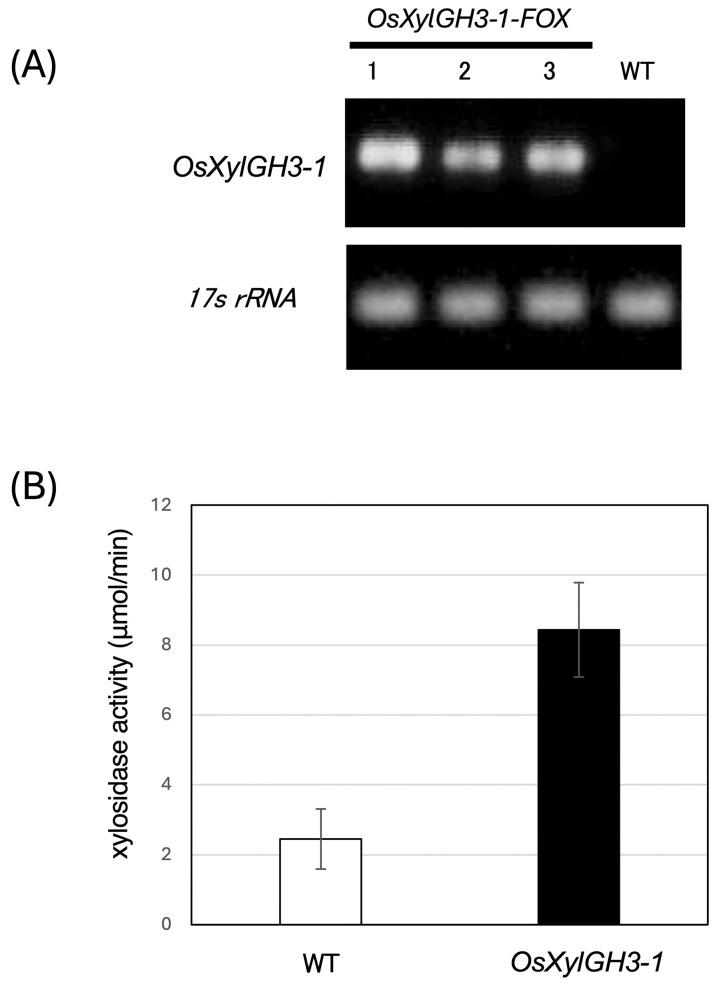 Figure 1
Figure 1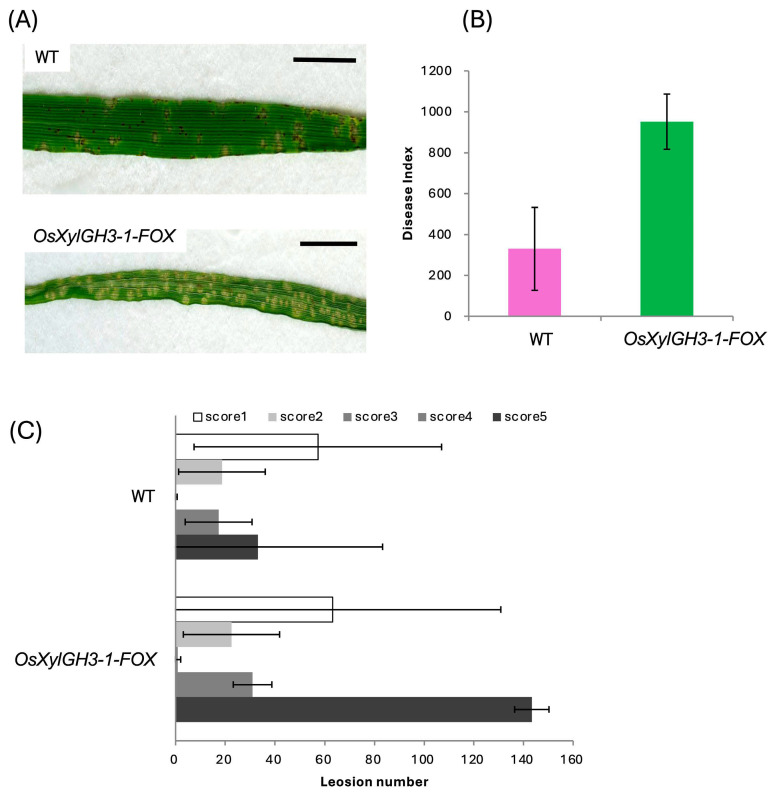 Figure 2
Figure 2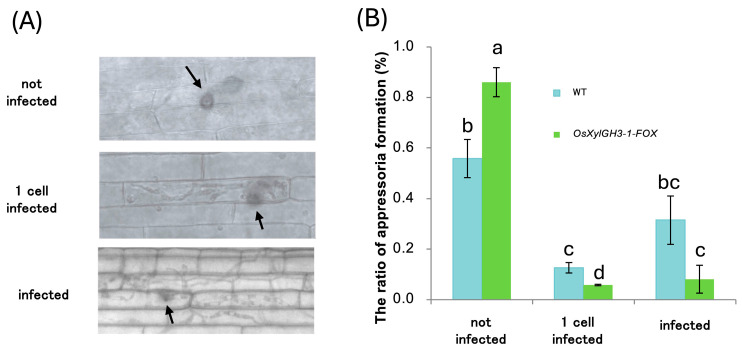 Figure 3
Figure 3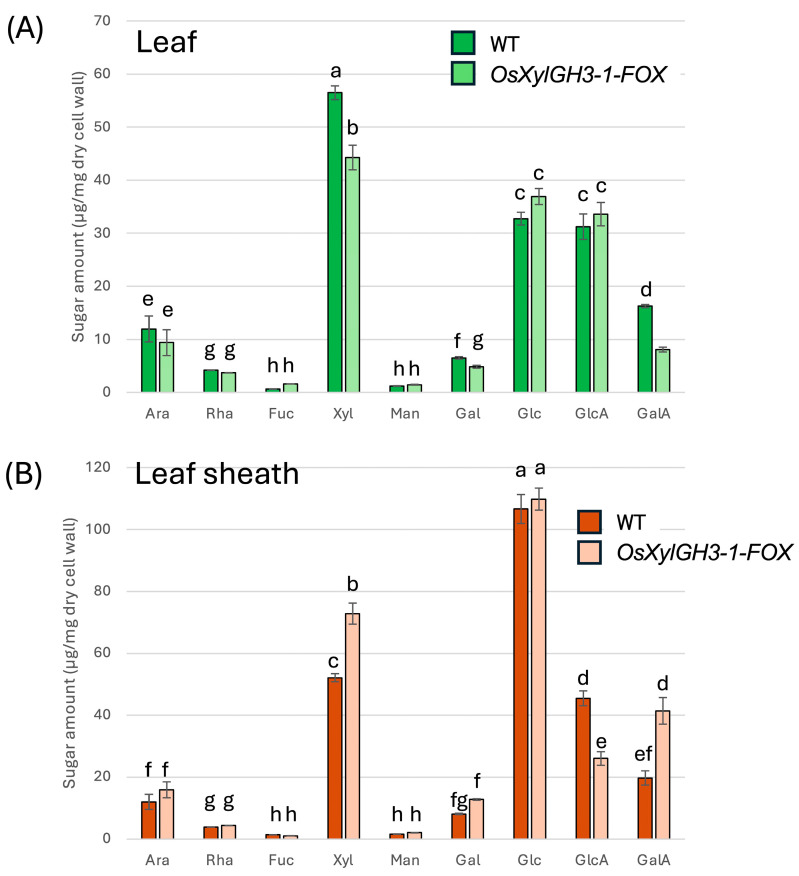 Figure 4
Figure 4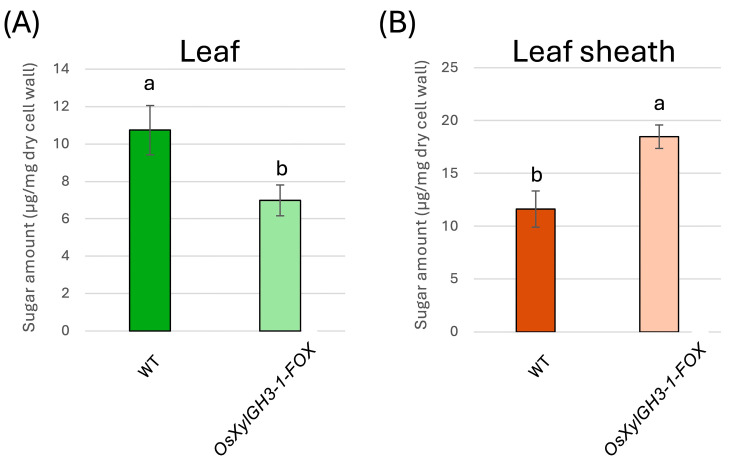 Figure 5
Figure 5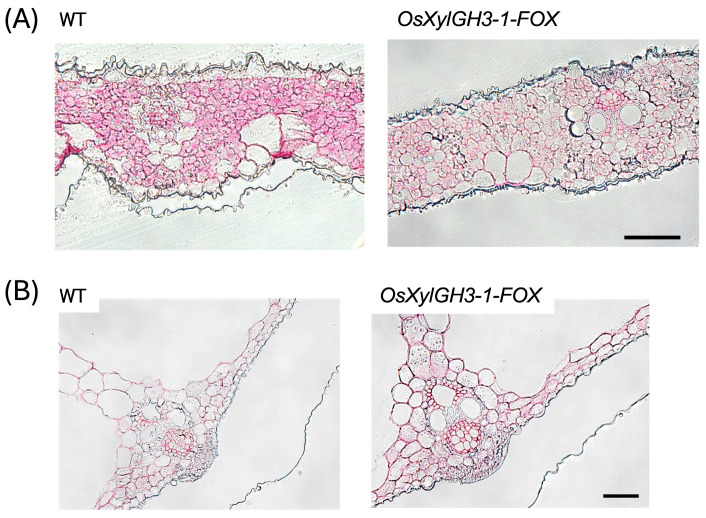 Figure 6
Figure 6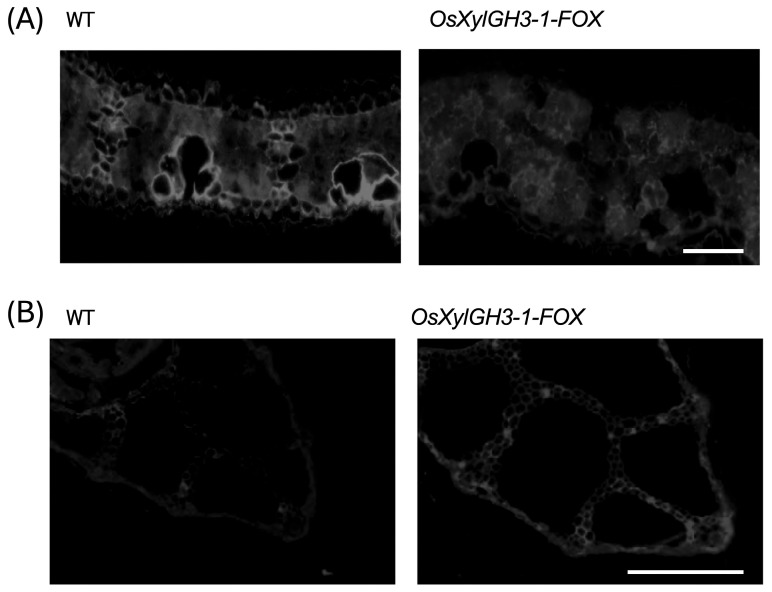 Figure 7
Figure 7- —ICHIMURA Foundation for New Technology
- —Tsukuba-Plant Innovation Research Center (T-PIRC), University of Tsukuba
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPolysaccharides and Plant Cell Walls · Plant-Microbe Interactions and Immunity · Biofuel production and bioconversion
1. Introduction
Plant cell walls are defining structures that maintain organ integrity, determine cell shape, guide differentiation and reproduction, and mediate a broad range of physiological functions [1]. As the outermost boundary of plant cells, the cell wall is the initial line of defense against biotic and abiotic stresses, such as herbivory, pathogen attack, oxidative stress, heat, intense light, salinity, and drought [2]. The primary cell wall is composed of three interwoven polysaccharide classes: cellulose, hemicelluloses, and pectins. These polysaccharides form a complex, cross-linked matrix that confers recalcitrance to saccharification and stress tolerance [3,4]. Cellulose, a β-1,4-glucan polymer, assembles into microfibrils through extensive hydrogen bonding, which imparts rigidity and load-bearing strength. Hemicelluloses (e.g., xylans, mixed-linkage glucans, and xyloglucans) associate with cellulose and lignin to stabilize the wall [5,6,7]. Pectins, which are rich in galacturonic acid (GalA), include homogalacturonan (HGA), rhamnogalacturonan I (RGI), rhamnogalacturonan II (RGII), and xylogalacturonan (XGA). The structures and substitutions of these pectins underlie wall plasticity and cell adhesion [1,8,9]. Pectin abundance varies among lineages. It is typically ~20–35% of the primary wall in dicots, but only ~5% in commelinid monocots, such as rice [10]. While cellulose composition is broadly conserved across land plants, hemicellulose composition varies by lineage: xyloglucan predominates in dicots, whereas xylan predominates in commelinid monocots (grasses) [11].
The major hemicelluloses in rice are arabinoxylans and mixed-linkage (1,3;1,4)-β-glucans. Arabinoxylans have a β-1,4-xylosyl backbone that is substituted with α-L-arabinofuranosyl side chains at O-2 and/or O-3 [12]. Xylan depolymerization occurs through the action of endo-β-1,4-xylanases (xylanases), which generate xylooligosaccharides (e.g., xylobiose), and exo-acting β-D-xylosidases (xylosidases), which remove successive xylosyl residues from the non-reducing ends of xylooligosaccharides [13,14]. Xylosidases are found in the glycoside hydrolase families GH3 and GH31; GH31 xylosidases are relatively well studied, while GH3 xylosidases are less characterized [15]. The rice gene Os03g0749100 encodes a GH3 family hydrolase with a predicted signal peptide and potential for secretion. Although GH3 enzymes are annotated with β-glucosidase activity (EC 3.2.1.21), they often exhibit broader substrate specificities, including β-xylosidase activity (EC 3.2.1.37). This implicates GH3 enzymes in xylan turnover and wall remodeling. Previous studies have shown that certain rice GH3 xylosidases, such as OsXyl1, can hydrolyze xylooligosaccharides (DP 2–6) and perform transglycosylation. This suggests that they play a role in cell wall recycling and have potential biocatalytic applications [15,16,17]. Elucidating the biochemical properties and physiological functions of Os03g0749100 could shed light on carbohydrate turnover and wall dynamics in rice.
Rice blast, caused by the fungus Magnaporthe oryzae, is the most destructive rice disease worldwide. After adhesion, M. oryzae forms an appressorium, accumulates glycerol, and generates extreme turgor pressure (up to ~8 MPa) to breach the host cell wall mechanically [18,19]. Leaf lesion severity is commonly assessed across standardized categories ranging from numerous brown specks (mild) to expanding whitish lesions (severe) [20]. To investigate how xylan catabolism influences wall architecture and disease resistance, we generated an Os03g0749100 FOX overexpression line under the maize ubiquitin-1 promoter [18,21]. Our results demonstrate that constitutive xylosidase overexpression produces organ-specific changes in wall composition and blast resistance, including decreased resistance in leaves and increased resistance in leaf sheaths. We also present biochemical and histochemical evidence that links pectin and cellulose content to these divergent outcomes.
2. Results
2.1. FOX Overexpression of a GH3 Xylosidase (Os03g0749100)
We screened ~14,500 rice FOX lines using the FOX hunting system by using expression libraries for full-length cDNAs (fl-cDNAs) from rice at a maximum of 28,000 fl-cDNA clones in total, individually overexpressed the fl-cDNAs in rice driven by the maize ubiquitin-1 gene promoter [22], and identified lines overexpressing the GH3-family xylosidase Os03g0749100. We refer to this line as OsXylGH3-1-FOX. Os03g0749100 transcript levels were elevated in OsXylGH3-1-FOX relative to wild type (WT) (Figure 1A). Consequently, xylosidase activity increased by approximately 3.4-fold in independent OsXylGH3-1-FOX line (line1, Figure 1A) compared to WT (Figure 1B). Under non-infected growth conditions, OsXylGH3-1-FOX plants did not show visible morphological or developmental abnormalities compared with WT Nipponbare (plant height, tiller number, leaf morphology, biomass).
2.2. Divergent Blast Resistance Responses in Leaves and Leaf Sheaths
Two weeks after sowing, the fourth leaves were spray-inoculated with M. oryzae. OsXylGH3-1-FOX plants exhibited significantly higher numbers of score-5 lesions (severe whitish lesions) at 3, 5, and 7 days post-inoculation (dpi) (Figure 2A). This resulted in a disease index that was >2-fold higher than that of WT (Figure 2B). Photographs illustrate the shift toward severe lesions in OsXylGH3-1-FOX (Figure 2C), as opposed to a uniform distribution of lesions ranging from mild brown to severe white in WT (Figure 2A,C). For leaf sheath assays, sheaths were excised at two weeks, injected with a 1.0 × 10^5^ spores/mL suspension, fixed, and stained after 48 h. Relative to WT, OsXylGH3-1-FOX leaf sheaths displayed more unpenetrated appressoria and fewer single-cell penetrations and multicell expansions (Figure 3A,B), indicating enhanced resistance at the entry stage. For each replicate, 100–150 appressoria were evaluated per sheath.
2.3. Monosaccharide Compositions of Hemicellulose/Pectin-Enriched Fractions
TFA-soluble fractions enriched in hemicellulose and pectin were analyzed by gas chromatography (GC). In leaves, OsXylGH3-1-FOX exhibited ~33% lower levels of xylose and arabinose (xylan signature sugars) and ~50% lower levels of galacturonic acid (GalA) (pectin signature) than WT (Figure 4A). In leaf sheaths, xylose and arabinose levels were similar to WT levels, while GalA levels increased by ~50% in OsXylGH3-1-FOX (Figure 4B). Statistical differences were assessed by one-way ANOVA followed by Tukey’s HSD; distinct letters indicate significant differences (p < 0.05).
2.4. Cellulose-Associated Sugar Quantification in TFA-Insoluble Fractions
The neutral sugars, primarily glucose derived from cellulose hydrolysates as determined by the anthrone assay, were quantified in the TFA-insoluble fraction. Cellulose content of the leaf blade was quantified from the TFA-insoluble fraction, as shown in Figure 5A. The fourth leaf of OsXylGH3-1-FOX contained approximately 35% less neutral sugar than the wild type (Figure 5A), suggesting reduced cellulose levels in OsXylGH3-1-FOX leaves. However, the leaf sheaths of OsXylGH3-1-FOX had about 70% more neutral sugars than the wild type (Figure 5B), suggesting increased cellulose abundance in the OsXylGH3-1-FOX sheaths. Statistical differences were assessed by one-way ANOVA followed by Tukey’s HSD; distinct letters indicate significant differences (p < 0.05).
2.5. Pectin and Cellulose Staining Patterns
Ruthenium red staining revealed lower pectin staining strength in OsXylGH3-1-FOX leaves compared to the WT (Figure 6A). Conversely, pectin staining strength was higher in OsXylGH3-1-FOX leaf sheaths than in the wild type (WT) (Figure 6B), which matches the biochemical increases in GalA. Leaf sections stained with toluidine blue revealed that cells in WT leaves were tightly aligned. In contrast, cells in OsXylGH3-1-FOX leaves were less aligned, and intercellular spaces were observed in some areas (Figure S1). Calcofluor white staining revealed localized regions of high cellulose in WT leaves (e.g., motor cells), while OsXylGH3-1-FOX leaves exhibited more uniform cellulose distribution (Figure 7A). In leaf sheaths, the staining strength of the cellulose staining was higher in the OsXylGH3-1-FOX leaves than in the WT leaves (Figure 7B). There were regularly localized regions of high cellulose in the OsXylGH3-1-FOX leaves.
To directly assess wall mechanics, we measured the mechanical strength of leaf blades. OsXylGH3-1-FOX leaves exhibited reduced breaking force compared with WT (Figure S2), consistent with compositional and staining data indicating compromised wall rigidity.
Calcofluor white staining revealed localized regions of high cellulose in WT leaves (e.g., motor cells), while OsXylGH3-1-FOX leaves exhibited more uniform cellulose distribution (Figure 7A). In leaf sheaths, the staining strength of the cellulose staining was higher in the OsXylGH3-1-FOX leaves than in the WT leaves (Figure 7B). There were regularly localized regions of high cellulose in the OsXylGH3-1-FOX leaves.
3. Discussion
Leaves of OsXylGH3-1-FOX exhibited significantly lower pectin content and weaker pectin staining than WT leaves (Figure 4A and Figure 6A). These leaves also showed formation of intercellular spaces (Figure S1). Both features closely resemble phenotypes reported for OsPG2-FOX rice lines, in which overexpression of a pectin degrading polygalacturonase reduces pectin, weakens cell adhesion, enlarges intercellular gaps, and facilitates M. oryzae ingress under conducive conditions [19]. Mechanistically, the pectin rich middle lamella is the principal mediator of plant cell–cell adhesion; perturbation of pectin abundance or cross linking compromises tissue integrity and increases wall porosity, thereby lowering the physical barrier to invasion [23,24]. These biophysical changes are significant because M. oryzae appressoria generate high turgor pressure (~8 MPa) and a rigid penetration peg, which depends on mechanical force to breach the cuticle and primary cell wall. Compared with the wild type, the leaves of OsXylGH3-1-FOX exhibited reduced cellulose content, which is a major determinant of cell wall strength, and diminished cellulose staining (Figure 5A and Figure 7A). Consequently, a decrease in wall strength or structural continuity can lower the threshold required for fungal [22,25]. Consistent with this interpretation, direct mechanical measurements showed reduced leaf mechanical strength in OsXylGH3-1-FOX relative to WT (Figure S2). We did not directly measure the mechanical properties of leaf sheaths, which is a limitation of the present work. A further limitation of this study is that the primary phenotypic analyses are based on a single FOX overexpression line. While FOX lines originate from independent full-length cDNA insertions, a more definitive understanding of the role of OsXylGH3-1 would require additional independent overexpression lines or loss-of-function mutants. Although we did not directly test Os03g0749100 activity on native rice xylan in vitro, several lines of evidence support its functional annotation as a GH3 β-xylosidase: (i) conserved GH3 catalytic domains and active-site features characteristic of β-xylosidases [11]; (ii) close homology to rice OsXyl1 with verified β-xylosidase activity on xylooligosaccharides (DP 2–6) [17]; and (iii) in vivo reductions in arabinoxylan signature sugars (xylose and arabinose) specifically in leaves (Figure 4A), consistent with xylan turnover in planta. In addition to differences in wall composition, leaf blades and leaf sheaths differ in development and anatomy. Leaf blades are photosynthetically active tissues that are highly expanded with thinner supporting structures and larger intercellular spaces. Leaf sheaths, on the other hand, consist of more compact, mechanically reinforced tissues that support the culm. These architectural and functional distinctions likely make blades more sensitive to reductions in pectin and cellulose while enabling sheaths to strengthen their wall matrices in response to elevated levels of these substances, thereby contributing to the divergent resistance phenotypes observed.
The observed decreases in xylose and arabinose in OsXylGH3-1-FOX leaves are consistent with β xylosidase overactivity on xylan/arabinoxylan side chains. A rice GH3 β-xylosidase (OsXyl1) efficiently hydrolyzes xylooligosaccharides, supporting a role in xylan turnover in planta [17]. Although a direct causal chain from xylan depletion to pectin loss is not yet defined in rice, covalent xylan–pectin–AGP linkages (the APAP1 proteoglycan) have been demonstrated in Arabidopsis, showing that hemicellulose and pectin domains can be structurally integrated; selective depletion of one domain can alter the extractability and organization of the other [21]. Together, these data support the inference that decreased pectin—and possibly secondary consequences of altered xylan–pectin connectivity—compromise adhesion and plausibly underlie reduced leaf resistance in OsXylGH3-1-FOX. In this study, β-xylosidase activity was quantified only in leaf extracts; we did not independently measure activity in leaf blades versus leaf sheaths, nor did we determine subcellular localization (apoplast vs. intracellular). Thus, tissue-specific differences in enzyme abundance, activation, or localization could contribute to the contrasting organ-level phenotypes. Future work employing tissue-resolved enzyme assays and localization analyses will help disentangle differential enzyme action from downstream remodeling.
In contrast, OsXylGH3-1-FOX leaf sheaths showed higher pectin content and staining (Figure 4B and Figure 6B), indicating improved adhesion. Moreover, cellulose content and staining strength were markedly increased (Figure 5B and Figure 7B). Because cellulose is the principal load-bearing component that confers mechanical rigidity, cellulose-rich walls strengthen the physical barrier at the penetration stage [26]. A stiffer, more continuous sheath wall should resist the penetration peg emergence that appressoria require to invade host tissue [25]. Consistent with this, several studies show that host cell wall strengthening—including lignification of epidermal mechanical tissues—can impede early penetration by blast fungus [27], and broader reviews of cell wall–associated immunity highlight coordinated remodeling of cellulose, pectin, and lignin as a front line defense [28,29].
Although levels of hemicellulose signature sugars decreased in OsXylGH3-1-FOX leaves, no significant change was detected in leaf sheaths (Figure 4). Relative to pectin (adhesion/plasticity) and cellulose (rigidity), hemicellulose appears secondary for determining the organ-specific resistance outcomes observed here. That said, recent biophysical work shows glucuronoarabinoxylan (GX) binds cellulose surfaces in a specific two-fold screw conformation and is critical for proper bundling and alignment of cellulose microfibrils in secondary walls; perturbing xylan can disrupt cellulose network architecture and mechanical performance [30]. Beyond structure, xylan-derived arabinoxylan oligosaccharides (AXOS) act as damage-associated molecular patterns (DAMPs) that trigger immune responses, and monocot xylanase inhibitors (TAXI/XIP/TLXI) contribute to wall-based immunity by inhibiting pathogen CWDEs [31,32]. Thus, hemicellulose can modulate resistance indirectly—via cellulose organization and immune signaling—even if its bulk content is not the primary determinant of the leaf vs. sheath phenotypes reported here.
A further limitation is that we did not assess immune signaling outputs (e.g., PR gene expression, ROS accumulation, or callose deposition) in leaves and sheaths before/after infection. Because cell wall remodeling interfaces with cell wall integrity sensing and downstream defense activation, future work should determine whether altered wall architecture in OsXylGH3-1-FOX modulates these pathways in addition to mechanical resistance.
Compared with OsPG2-FOX, which directly degrades homogalacturonan and recapitulates reduced adhesion and increased leaf susceptibility [19], OsXylGH3-1-FOX likely acts via xylan–pectin cross-domain interactions that secondarily impact pectin abundance/extractability. Notably, the organ-specific divergence we report here (leaf susceptibility vs. sheath resistance) has not, to our knowledge, been described for OsPG2-FOX or other wall-modifying lines, underscoring tissue-dependent consequences of β-xylosidase-mediated remodeling.
Constitutive overexpression of a GH3 β-xylosidase in rice produces organ-specific cell wall remodeling and divergent blast resistance phenotypes. In leaves, reduced pectin compromises adhesion and increases susceptibility. In leaf sheaths, increased pectin and cellulose strengthen wall integrity and improve resistance to penetration. These findings support a positive correlation between pectin/cellulose abundance and blast resistance, with hemicellulose contributing secondary effects. Future work should assess defense gene expression, lignification, and the biophysical properties of cell walls to further elucidate how xylosidase-driven remodeling shapes organ-specific immunity. Together with new mechanical data for leaves (Figure S2) and explicit limitations noted above, these results provide an integrated view of how xylosidase-driven remodeling shapes organ-specific immunity and mechanics.
4. Materials and Methods
4.1. Plant Material and Growth Conditions
The rice plants used were of the wild type (Oryza sativa cv. Nipponbare) and of the FOX line BP164, which carries an overexpression construct for OsXylGH3-1 (Os03g0749100). The plants were grown in a growth chamber at 28 °C with 115 µmol s^–1^ m^–2^ illumination. The presence of the constructs in the genomic DNA was confirmed by PCR using third-generation plants. Transgenic lines were selected on hygromycin-containing agar and tested for heritable expression patterns and altered sugar traits.
To analyze gain-of-function phenotypes, the FOX hunting system was developed using full-length cDNA (fl-cDNA) expression libraries from rice containing 28,000 fl-cDNA clones. They overexpressed the fl-cDNAs in rice under the control of the maize ubiquitin-1 promoter [18]. Among ~14,500 FOX rice lines, one overexpressed fl-cDNAs for xylosidase (accession no. AK102309; Rap-ID Os03g0749100) in GH family 3 and was named OsXylGH3-1-FOX. All FOX lines were compared to the WT Nipponbare background following standardized FOX-hunting procedures [18].
4.2. RNA Extraction and Gene Expression Analysis
The plant material was frozen in liquid nitrogen and ground using a TissueLyser II (Qiagen, Hilden, Germany) [19]. Total RNA was extracted using an RNeasy Plant Mini Kit (Qiagen, Hilden, Germany) and DNase I (Roche, Basel, Switzerland) following the manufacturer’s instructions.
cDNA was synthesized using ReverTra Ace^®^ (Toyobo, Tokyo, Japan) according to the manufacturer’s specifications. For the OsXylGH3-1-FOX line, transcripts were quantified using the primers OsXylGH3-1-forward (5′-TGCCACGTACTTGGTTCAA-3′) and OsXylGH3-1-reverse (5′-TAACCCCTAACTCCGGCTC-3′). The 17S rRNA transcript was used as an endogenous control because the small subunit rRNA in rice is a 17S molecule, as established by the complete sequencing of rice nuclear rDNA [33]. Primer specificity was confirmed by NCBI BLAST online tool against the Oryza sativa genome, and the amplicon size was verified by gel electrophoresis to match the expected product. We removed duplicated sentences and streamlined the paragraph for clarity.
4.3. β-Xylosidase Assay
The rice leaf blades were placed in a crushing tube, frozen in liquid nitrogen, and crushed using a TissueLyser (Qiagen). Then, one volume of extraction buffer (50 mM Tris-HCl, 3 mM EDTA, 1 M NaCl, and 10% glycerol [v/v]) was added. The sample was gently mixed and allowed to stand at 4 °C for two hours. Next, the sample was centrifuged at 15,000 rpm and 4 °C for 15 min. Finally, the supernatant was collected and used as the crude enzyme solution.
Enzymes were extracted in a buffer made with sodium acetate (20 mM) and NaCl (1 M). The pH of the buffer was 5.0. Each tissue was ground in liquid nitrogen, extracted in sodium acetate buffer, and then centrifuged at 15,000× g for 30 min at 4 °C. The hydrolytic activities of the cell wall proteins were determined. The cell wall proteins were extracted from each tissue. They were determined toward synthetic substrates. A reaction mixture was used. It consisted of the cell wall proteins, 25 mM acetate buffer (pH 5.0), and 1 mM PNP-glycoside substrate. The reaction was terminated by the addition of 200 mM sodium carbonate (800 µL) after being incubated at 37 °C for 2 h. The color change was measured at 420 nm. One unit of enzyme activity liberates 1 µmol of p-nitrophenol per minute.
4.4. Extraction and Analysis of Cell Wall Polysaccharides
Cell wall extraction and analysis were conducted according to Sumiyoshi et al., with slight modifications [9]. Full, mature leaves were frozen in liquid nitrogen and ground using a TissueLyser II (Qiagen) at 30 Hz for two minutes. The resulting powder was washed with 80% ethanol. After centrifugation for five minutes at 15,000 g, the supernatant was decanted and the pellet was treated with methanol and chloroform (MC; 1:1, v/v); acetone; and phenol, acetic acid, and water (PAW; 2:1:1, v/v). After amylase treatment to remove starch, the samples were air-dried, and the resulting alcohol-insoluble residue (AIR) was used as cell wall material. The AIR (2 mg) was hydrolyzed in 2 M trifluoroacetic acid (TFA), and the resulting pellets were hydrolyzed in 72% H_2_SO_4_. The resulting fractions were treated with methanol and hydrogen chloride. The methyl glycosides were then converted into trimethylsilyl derivatives and analyzed by gas–liquid chromatography (GC-2010; Shimadzu, Kyoto, Japan). The hexose and uronic acid contents were determined by the anthrone and meta-hydroxybiphenyl methods, respectively.
4.5. Microscopy
The following procedure is based on the method described by Hasegawa et al. [34]. A fully expanded leaf was fixed for 2 h in 4% paraformaldehyde, 0.25% glutaraldehyde, and 0.05 M phosphate buffer at room temperature. Transverse 3 µm sections were cut using a Leica RM2145 microtome (Leica Microsystems, Wetzlar, Germany). The sections were stained for 5 min with 0.02% (w/v) ruthenium red in DW and visualized by light microscopy. For cellulose staining, sections were treated with 0.01% (w/v) calcofluor white (Fluorescent Brightener 28, FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) for 1 min, washed three times with DW, and visualized by fluorescence microscopy (Leica, Wetzlar, Germany; DMRB).
4.6. Assay of Disease Resistance
For spray inoculation, intact rice seedlings (fourth leaf stage) were grown in a hydroponic culture for three weeks, as previously described [19]. The fourth leaves were excised and placed on moist filter paper in a clear plastic box. The leaves were spot-inoculated with a 10 µL suspension of M. oryzae (strain P91-15B; race 001; 86–137) conidia (1 × 10^5^ or 10^6^ mL^−1^) and incubated at 25 °C in the dark for two days. Subsequently, the leaves were incubated at 25 °C under continuous white light for five days. Disease symptoms were recorded at 3, 5, and 7 dpi. Lesion size was measured and the disease index was calculated according to Toyomasu et al. [35]. For the Blast Resistance Test by Leaf Sheath Inoculation, rice plants were grown in an incubator for two weeks, and cultured blast fungi were used as in the blast fungus spray experiment. The rice plants were cut approximately 1 cm from the root at the bottom and 1 cm from the leaf base at the top. The intermediate sections were used as leaf sheaths for the experiment. A spore suspension adjusted to a concentration of 1 × 10^5^ spores/mL was injected into the leaf sheaths using a syringe. Cardboard was placed in a square Petri dish and moistened with deionized water (DW) sufficient to maintain humidity for approximately one day. A toothpick was placed on top of the cardboard to keep the leaf sheaths from touching it directly. The injected leaf sheaths were then placed on top of the toothpick. The Petri dish was covered with a lid and sealed with mending tape. It was then left in the dark at 28 °C for 48 h. The mycelia that had penetrated the cells were stained with crystal violet and observed. For each replicate, 100–150 appressoria were evaluated. Magnification and field of view were standardized across samples. Penetration categories were scored as defined in Figure 3A.
4.7. Mechanical Strength Measurements
The mechanical properties of leaves were measured following the method described by Ohara et al. [19]. Mature leaves were cut approximately 12 cm from the tip, mounted with a 2 cm interval between both ends, and positioned so that the midpoint of the sample was placed over the sensor. Breaking force (N/mm) and strain (%) were measured using a force tester (MCT-1150, A&D COMPANY, Ltd., Tokyo, Japan).
4.8. Statistical Analysis
The data were expressed as the mean values ± standard deviations (SD) taken from 4–9 biological independent experiments. The experimental data of the samples were statistically analyzed through one-way analysis of variance (ANOVA) with Tukey’s post hoc test using Statistica 13.1 software (StatSoft, Inc., Tulsa, OK, USA). The results with p-value ≤ 0.05 and p-value ≤ 0.01 were considered statistically significant. Figure 3, Figure 4 and Figure 5 include statistical annotations; distinct letters denote significant differences (p < 0.05) based on ANOVA/Tukey’s HSD. For clarity, the number of biological replicates is summarized as follows: RT-PCR (n = 3), enzyme assays (n = 3), GC sugar analysis (TFA-soluble and TFA-insoluble; n = 4), leaf blade disease assays (n = 5–9 independent leaves), leaf sheath penetration assays (n = 4 sheaths; 100–150 appressoria/replicate), and histochemical staining (≥2 independent experiments with similar results). All comparisons are against WT Nipponbare. All statistical analyses were conducted within a hypothesis-driven framework in which predefined comparisons between the WT and the FOX line were evaluated using one-way ANOVA followed by Tukey’s HSD.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Iwai H. Virtual Issue: Cell Wall Functions in Plant Growth and Environmental Responses J. Plant Res.20211341155115810.1007/s 10265-021-01351-y 34613490 · doi ↗ · pubmed ↗
- 2Talbot N.J. On the Trail of a Cereal Killer: Exploring the Biology of Magnaporthe grisea Annu. Rev. Microbiol.20035717720210.1146/annurev.micro.57.030502.09095714527276 · doi ↗ · pubmed ↗
- 3Wyman C.E. Decker S.R. Himmel M.E. Brady J.W. Skopec C.E. Viikari L. Hydrolysis of Cellulose and Hemicellulose Polysaccharides Dumitriu S. CRC Press Boca Raton, FL, USA 20049951033
- 4Fangel J.U. Ulvskov P. Knox J.P. Mikkelsen M.D. Harholt J. Popper Z.A. Willats W.G.T. Cell Wall Evolution and Diversity Front. Plant Sci.201232596010.3389/fpls.2012.00152 PMC 339060322783271 · doi ↗ · pubmed ↗
- 5Kim Y.-J. Jeong H.Y. Kang S.-Y. Silva J. Kim E.-J. Park S.K. Jung K.-H. Lee C. Physiological Importance of Pectin Modifying Genes During Rice Pollen Development Int. J. Mol. Sci.202021484010.3390/ijms 2114484032650624 PMC 7402328 · doi ↗ · pubmed ↗
- 6Jeong H.Y. Nguyen H.P. Lee C. Genome-Wide Identification and Expression Analysis of Rice Pectin Methylesterases: Implication of Functional Roles of Pectin Modification in Rice Physiology J. Plant Physiol.2015183232910.1016/j.jplph.2015.05.00126072144 · doi ↗ · pubmed ↗
- 7Vogel J. Unique Aspects of the Grass Cell Wall Curr. Opin. Plant Biol.20081130130710.1016/j.pbi.2008.03.00218434239 · doi ↗ · pubmed ↗
- 8Carpita N.C. Gibeaut D.M. Structural Models of Primary Cell Walls in Flowering Plants: Consistency of Molecular Structure with the Physical Properties of the Walls during Growth Plant J.1993313010.1111/j.1365-313X.1993.tb 00007.x 8401598 · doi ↗ · pubmed ↗