An Efficient Micropropagation Protocol for Camellia chekiangoleosa ‘Ganhongyou 1’ via Stem Segment Culture
Anni Liu, Yixuan Peng, Xin Chen, Qiangqiang Cheng, Kang Zha, Qiang Wen

TL;DR
This paper develops an efficient method to propagate the Camellia chekiangoleosa ‘Ganhongyou 1’ plant using stem segment culture, achieving high survival and growth rates.
Contribution
A novel micropropagation protocol for Camellia chekiangoleosa ‘Ganhongyou 1’ with optimized growth regulator combinations and a phloem-rooting pattern confirmation.
Findings
Optimal bud induction achieved with 86.67% success using 6-BA and IAA on Hyponex medium.
A combined in vitro and ex vitro strategy resulted in an 88% survival rate after rooting and acclimatization.
Adventitious roots originated from phloem parenchyma cells, confirming a phloem-rooting pattern.
Abstract
The provincial-level registered superior cultivar Camellia chekiangoleosa ‘Ganhongyou 1’ boasts superior economic traits coupled with significant ornamental value, driving demand for an efficient propagation system. Consequently, this study aimed to develop a rapid micropropagation protocol by investigating culture conditions using semi-woody nodal segments with axillary buds as explants on Hyponex basal medium supplemented with varying combinations of plant growth regulators. Contamination was effectively minimized to 18% by a combined approach of surface sterilization (75% ethanol, 0.1% HgCl2, and 20% NaClO) and incorporating 1 mL/L bactericide into the induction medium. For bud induction, the optimal medium was 2 g/L Hyponex supplemented with 1.0 mg/L 6-benzylaminopurine (6-BA) and 0.2 mg/L indole-3-acetic acid (IAA), achieving an 86.67% induction rate. The best proliferation was…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2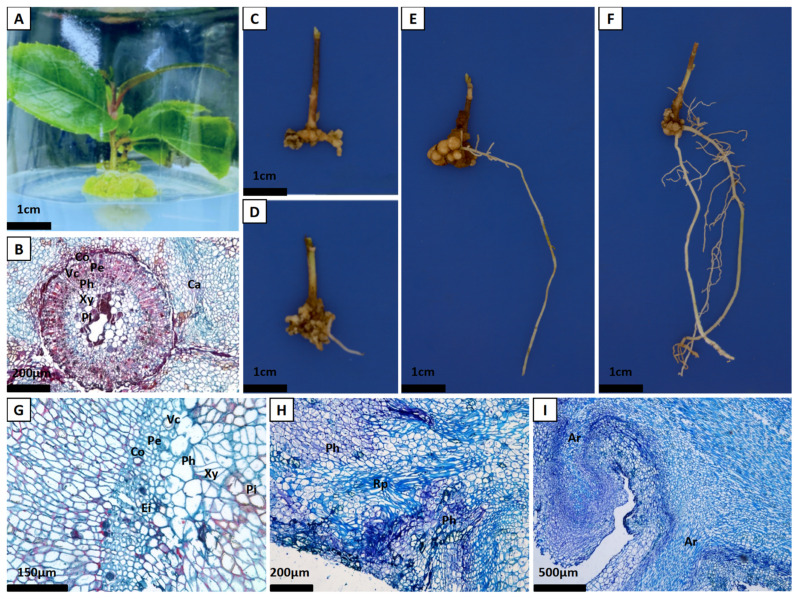 Figure 3
Figure 3 Figure 4
Figure 4- —Jiangxi Provincial Key Research and Development Program
- —Natural Science Foundation of Jiangxi Province, China
- —Jiangxi Provincial Early-Career Young Scientific and Technological Talent Training Program
- —Oil-Tea Special Research Project of Jiangxi Provincial Department of Forestry
- —Basic Research and Talent Research Special Projects of Jiangxi Academy of Forestry
- —Key Program of Jiangxi Provincial Natural Science Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant tissue culture and regeneration · Plant Molecular Biology Research · Flowering Plant Growth and Cultivation
1. Introduction
Oil-tea camellia, such as Camellia oleifera is one important woody oil crop, and its oil is rich in unsaturated fatty acids and various vitamins, which has been promoted by the Food and Agriculture Organization (FAO) as a healthy edible oil [1]. C. chekiangoleosa is one of the primary oil-tea camellia species in China. As an endemic species distributed in the mountainous regions of southern China, its seed oil surpasses the widely cultivated C. oleifera in several key aspects: extraction yield, content of unsaturated fatty acids (particularly oleic acid), and levels of natural antioxidants such as squalene and phytosterols [2,3,4]. Consequently, it possesses significant development potential for applications in the food, pharmaceutical, and cosmetic industries [5,6,7,8]. Despite these merits, conventional breeding of oil-tea camellia species, including C. chekiangoleosa, nonetheless faces significant constraints due to high heterozygosity and a long reproductive cycle, which lead to a shortage of superior germplasm resources [4,8]. Consequently, this bottleneck severely hampers varietal improvement and the sustainable development of the industry.
Plant tissue culture (PTC) serves as a pivotal biotechnology, playing an irreplaceable role in elucidating plant growth and development mechanisms, identifying gene functions, and creating transgenic plants [9,10,11,12]. It also provides a controlled experimental platform for germplasm conservation and efficient regeneration [13,14]. The successful establishment of this technical system highly depends on the appropriate selection of basal media and the precise regulation of plant growth regulators [14,15]. The type, concentration, and ratio of plant growth regulators directly determine the success or failure of key stages such as explant dedifferentiation, redifferentiation, and rooting [16]. The tissue culture of oil-tea camellia poses particular challenges. Its tissues are rich in polyphenols, making them prone to browning under in vitro conditions, and they exhibit high sensitivity to exogenous plant growth regulators, which therefore require careful screening and optimization [17]. Current studies mostly use buds or stem segments of oil-tea camellia as explants. These are cultured on basal media such as Murashige and Skoog medium (MS), woody plant medium (WPM), Hyponex, or 1/2 MS, supplemented with various combinations of cytokinins—for example, 6-benzylaminopurine (6-BA), benzyladenine (BA), zeatin (ZT), or thidiazuron (TDZ)—and auxins such as 1-naphthaleneacetic acid (NAA), indole-3-butyric acid (IBA), or indole-3-acetic acid (IAA) [18]. To improve culture outcomes, gibberellic acid (GA_3_) at concentrations ranging from 1.0 to 6.0 mg/L or casein hydrolysate at 500–600 mg/L is sometimes added as a supplement [19]. However, the rooting of shoots remains a critical bottleneck. Research indicates that IBA is widely used due to its high efficacy in inducing root primordia formation, and it can promote rooting either alone or in combination with NAA [20]. As conventional rooting media often yield limited results, a two-step method is frequently adopted: root primordia are first induced in a high-concentration auxin medium, followed by transfer to a hormone-free medium for further development [21], or alternatively, auxin soaking combined with ex vitro rooting is employed to improve efficiency [22]. Nonetheless, existing protocols still suffer from insufficient stability and reproducibility, and the responses to growth regulators vary significantly among different species and genotypes, which constrains the generalizability and broader application of the technique. Therefore, for C. chekiangoleosa, which has a relatively weak research foundation, systematically elucidating its response patterns to growth regulators and establishing an optimized, tailored system are of great significance for achieving efficient micropropagation and promoting its industrial application.
While micropropagation protocols have been developed for C. oleifera, no such system exists for C. chekiangoleosa. To address this gap, this study utilized nodal segments with buds of the superior cultivar ‘Ganhongyou 1’ to establish a complete micropropagation protocol. Key techniques including shoot induction, multiplication, and efficient rooting were systematically investigated. The aim was to develop a reliable in vitro propagation system, thereby providing a technical foundation for the large-scale production of high-quality planting material.
2. Results
2.1. Contamination Control with Different Disinfection Treatments
To establish an effective sterilization protocol, we evaluated seven different disinfection methods (i–vii). When explants were treated with 0.1% HgCl_2_ (method i) or 20% NaClO (method ii) for 7–10 min after 75% C_2_H_5_OH immersion, the contamination rate still exceeded 90% (Table 1). Although approximately 1% of the explants showed axillary bud swelling, their leaves failed to unfold; the remaining explants mostly remained unchanged or turned brown and died. When a pretreatment procedure was added (method iii)—repeatedly wiping stems with 75% ethanol and then rinsing under running water for 2–3 h—it not only lowered the contamination rate to 51% but also significantly increased the induction rate to 15.67%. Further reductions in contamination were achieved by additional measures: indoor pre-cultivation (method iv), an extra round of HgCl_2_ disinfection (method v), or an additional NaClO treatment (method vi). Among these, the supplementary NaClO treatment (method vii) was most effective, lowering the contamination rate to 24.67%. Building on method vii, a further and significant reduction in contamination—to 18.00%—was achieved by incorporating 1 mL/L bactericide into the induction medium. This combined approach was designated as method vii. Moreover, this combined approach not only achieved lower contamination but also markedly improved the bud induction rate and shoot vigor. The induced shoots exhibited robust growth, obvious elongation, long internodes, and numerous axillary buds—traits that are highly conducive to subsequent proliferation.
2.2. Axillary Bud Induction Using Different Media Formulations
To determine the optimal basal medium concentration, explants were cultured on Hyponex at varying levels (0.5, 1.0, 1.5, 2.0, 2.5, 4.0 g/L). At the lower end (0.5 g/L), nutrient insufficiency resulted in the lowest induction rate and poor bud development. Growth status improved with increasing concentration up to 2.0 g/L, which yielded the best outcomes. At the higher end (4.0 g/L), however, adverse effects including bud necrosis and leaf yellowing were observed (Table 2). Considering both the induction rate and growth performance, a Hyponex concentration of 2.0 g/L is determined to be optimal.
2.3. Axillary Bud Induction with Different PGR Combinations
Under a constant cytokinin (6-BA) concentration (0.5 or 1.0 mg/L), the addition of the same concentration (0.1 or 0.2 mg/L) of different auxins (IAA, NAA, or IBA) resulted in significant differences in both the induction rate and growth of axillary buds. As shown in Table 3, a higher concentration of 6-BA (1.0 mg/L) resulted in a greater bud induction rate compared to 0.5 mg/L. Specifically, based on this optimal concentration, the formulation with 1.0 mg/L 6-BA and 0.2 mg/L IAA was the most effective, achieving an induction rate of 86.67%. In terms of shoot morphology, buds from the 6-BA and IAA treatment were robust with dark green, smooth leaves, whereas those induced by 6-BA and IBA were relatively short despite moderate induction rates (Table 3; Figure 1A,B). The combination of 6-BA and NAA resulted in the lowest induction rate, accompanied by leaf browning and a failure of most axillary buds to sprout (Table 3; Figure 1C). Therefore, based on induction rate and shoot quality, the formulation of 2.0 g/L Hyponex supplemented with 1.0 mg/L 6-BA and 0.2 mg/L IAA was identified as optimal for axillary bud induction.
2.4. Shoot Proliferation with Different PGRs
To optimize the shoot proliferation medium, we examined the effects of different concentrations and combinations of 6-BA, IBA, and GA_3_. At conventional concentrations, the combination of 6-BA and IBA proved more effective than combinations with NAA or IAA, yielding a higher proliferation coefficient (Table 4 Group A; Figure 2A). A high IBA concentration (0.4 mg/L) resulted in good proliferation and longer internodes, but the leaves exhibited wilting and yellowing in the later stages of culture. Reducing the IBA concentration to 0.15 mg/L maintained a significant proliferation effect while promoting healthier shoot growth (Table 4 Group B; Figure 2B). The optimal 6-BA concentration was determined to be 1.0 mg/L. Higher concentrations (e.g., 4.0 mg/L) inhibited shoot growth—causing stunting, discoloration, and increased mortality—despite promoting proliferation (Table 4 Group C; Figure 2C). At 1.0 mg/L, proliferation remained effective while shoots grew vigorously. Furthermore, following screening across concentrations (0.2, 0.5, and 2.0 mg/L), the addition of 0.5 mg/L GA_3_ ultimately yielded the best proliferation outcome. This was characterized by an increased shoot number, smooth leaves, purplish-red new shoots, and a maximal proliferation coefficient of 6.53 (Table 4 Group D; Figure 2D). Based on the proliferation and growth performance, the optimal proliferation medium was identified as: 2.0 g/L Hyponex + 1.0 mg/L 6-BA + 0.15 mg/L IBA + 0.5 mg/L GA_3_.
2.5. A Two-Step Rooting and Acclimatization Strategy
Acknowledging the bottleneck of poor in vitro rooting in oil tea, we therefore developed and evaluated a novel two-step strategy combining in vitro pretreatment with ex vitro rooting. This protocol consisted of a 20-day in vitro pre-culture on an optimized medium, followed by ex vitro establishment. The pre-culture medium was formulated based on 1/2 strength Hyponex [23]. Through screening, the optimal pretreatment medium contained 1.0 mg/L NAA and 2.0 mg/L IBA. After a 20-day culture, explant bases remained flat with minimal callus (Table 5). It was noted that explants grown exclusively in vitro for 60 days showed no root emergence or primordium formation (Figure 3A,B), underscoring the limitation of purely in vitro conditions. Plantlets subjected to the two-step protocol were then dipped in 1.0 g/L ABT (a rooting powder complex) and transplanted. At 15 days post-transplantation, external base swelling and callus formation coincided with the internal dedifferentiation of bark parenchyma cells (Figure 3C,G). By 60 days, the emergence of adventitious roots corresponded to the development of distinct root primordia originating from phloem parenchyma, confirming a phloem-rooting pattern (Figure 3D,H). Subsequently, the roots underwent active elongation (75 days, Figure 3I) and matured into a fibrous system (90 days, Figure 3F). This integrated protocol achieved an 88% transplant survival rate, yielding vigorous seedlings with smooth, green leaves (Table 5).
3. Discussion
Effective control of microbial contamination is critical prerequisite for successfully establishing an aseptic tissue culture system in oil-tea camellia [13]. Excessive sterilization time or inappropriate disinfectant selection can easily lead to a high explant contamination rate, exacerbate browning, and even impede normal bud differentiation [24,25,26]. In plant tissue culture, the conventional combination of ethanol and mercuric chloride (HgCl_2_) is a commonly used surface sterilization method, but its efficacy is often unsatisfactory when applied to oil tea camellia. To address these limitations, improved strategies have been explored. Previous studies have shown that incorporating a preliminary running water rinse before chemical disinfection can significantly reduce the contamination burden on oil tea explants [14]. For oil-tea camellia explants, a 2–5 h tap water rinse markedly lowered contamination [15]. Furthermore, composite disinfection using both sodium hypochlorite and HgCl_2_, or the addition of novel fungicides (to control endogenous fungal contamination) and bacteriostatic agents (to control endogenous bacterium contamination) to the culture medium, has shown promise. In the cultivar ‘Haida Youcha 4’, a treatment involving 0.1% HgCl_2_ (12 min) and 2% NaClO (15 min), combined with 0.1 g/L of the bactericide in the induction medium, reduced the contamination rate to 16.7% [18]. Building upon these validated methods, the present study integrated strategies including preliminary running water rinsing, composite chemical disinfection, and the addition of a bactericide during the induction phase to systematically screen for the optimal sterilization protocol applicable to C. chekiangoleosa (Table 1). This integrated approach successfully minimized contamination and supported robust bud growth.
The selection of an appropriate basal medium is crucial for the efficient induction of axillary buds. Owing to its comprehensive composition, MS medium is the most widely used medium in the tissue culture of oil-tea camellia [2]. However, the present study found that for the ‘Ganhongyou 1’ cultivar of C. chekiangoleosa, the Hyponex medium demonstrated superior performance in bud induction, achieving an induction rate of 84.33% (Table 2). This discrepancy may be closely associated with the inorganic salt composition of the two media. MS medium is characterized by a high total concentration of inorganic salts and abundant nitrogen content [27]; In contrast, the Hyponex medium features a higher potassium content, with an N:P:K mass ratio of 7:6:19. In this study, it was observed that explants cultured on the Hyponex medium not only exhibited faster axillary bud sprouting but also developed into more robust seedlings. This may be attributed to its higher proportion of potassium (K), which plays a key role in maintaining cellular osmotic pressure, ion balance, and the buffering capacity of the medium, thereby contributing to the stability of seedling growth during the cultivation process [28]. Dai Xiaoying et al. observed a similar phenomenon in their study on axillary bud sprouting of C. chekiangoleosa [29]. Guo Chunxi et al. further noted that the content and ratio of N, P, and K significantly influence the growth and economic traits of oil-tea camellia, with a higher proportion of potassium being most beneficial for growth promotion [30]. In summary, the appropriate N:P:K ratio in the Hyponex medium better aligns with the nutritional requirements for bud induction and early-stage growth of ‘Ganhongyou 1’. This study also found that the concentration of Hyponex not only affects the number of axillary buds but also significantly influences their growth status, with 2.0 g/L Hyponex being most conducive to the induction and robust growth of axillary buds.
Plant growth regulators play a critical regulatory role in the induction and proliferation of axillary buds in C. chekiangoleosa. Existing studies have shown that in the tissue culture of oil-tea camellia, a combination of cytokinins (such as 6-BA, ZT, or TDZ) and auxins (such as NAA, IBA, or IAA) is typically employed to enhance culture efficiency, occasionally supplemented with an appropriate amount of GA_3_ or hydrolyzed casein [18,19]. This study systematically compared the effects of three auxins (IAA, IBA, and NAA) in combination with 6-BA. The results revealed that the combination of 6-BA and IAA yielded the optimal induction outcome, achieving an induction rate of 86.67% while producing buds of good quality (Table 3; Figure 1). This finding aligns with the conclusions from research on bud induction in the ‘Huashuo’ cultivar of C. oleifera [20]. However, responses to plant growth regulators vary among different cultivars. For instance, one study indicated that NAA plays a primary role in promoting axillary bud sprouting in the ‘Yangxin Miju’ cultivar of oil-tea camellia [31]. This highlights that genotype is a significant factor influencing hormonal effects and underscores the necessity of tailoring culture protocols according to specific varietal traits. Regarding concentration, the present study observed a positive correlation between 6-BA concentration and bud induction rate within the range of ≤1.0 mg/L (Table 3). Nevertheless, it is important to note that a higher cytokinin concentration is not always more effective. In their research on C. chekiangoleosa, Dai Xiaoying et al. reported that the number of induced axillary buds initially increased and subsequently decreased as the 6-BA concentration rose, suggesting that low concentrations are insufficient for effective induction, whereas high concentrations can be inhibitory. Consequently, future studies exploring higher 6-BA concentrations (>1.0 mg/L) should also focus on determining the concentration threshold of its inhibitory effects to more comprehensively optimize the induction system [29,32].
In the proliferation culture of oil-tea camellia, the cytokinin 6-BA is widely utilized, and a combination of a high concentration of 6-BA with a low concentration of auxin is generally considered an optimal formulation [18]. For example, in the ‘Xianglin 1’ cultivar, a formulation containing 5.0 mg/L 6-BA, 0.01 mg/L NAA, and 1.0 mg/L biotin achieved the highest proliferation coefficient [33]. However, excessively high concentrations of plant growth regulators can also inhibit the growth of tissue-cultured plantlets. The underlying mechanism may involve the activation of polyphenol oxidase, which in turn intensifies tissue browning and mortality [34]. The findings of this study are consistent with this observation: when the 6-BA concentration reached 4 mg/L, although the number of axillary buds was high, bud elongation was significantly inhibited, and the browning and mortality rates of the shoots increased (Figure 2; Table 4). To alleviate the dwarfing effect induced by cytokinin, GA_3_ was added to the proliferation medium in this study, leveraging its role in promoting cell elongation and division [35]. Ultimately, through the combined application of 6-BA, IBA, and GA_3_, vigorous plantlets with robust stems and dark green leaves were obtained while maintaining a high proliferation rate (Figure 2; Table 4). Similar results have also been reported in the ‘Huashuo’ and ‘Haida Youcha 4’ cultivars, where the addition of 6.0 mg/L GA_3_ significantly promoted bud proliferation and growth [33,35]. The subculture of red-flowered oil-tea camellia primarily relies on cluster shoot proliferation, which is attributed to the abundant meristems at the shoot base [29]. The continuous emergence of new shoots from the base results in uneven growth, with many exhibiting a purplish-red coloration. Independent plantlets differentiate only after the shoot height exceeds 2 cm. This phenomenon may be associated with insufficient light exposure at the base due to the relatively large leaves, consequently affecting chlorophyll synthesis and stem elongation. Therefore, it is recommended to use large-mouth vessels with ample volume and to divide the cluster shoots into smaller explants during subculture to improve light penetration, thereby enhancing shoot quality and morphology.
The in vitro rooting of oil-tea camellia plantlets presents considerable difficulty. When conventional rooting media containing NAA and IBA are used, the resulting rooted plantlets typically produce only a single main root with few fibrous roots [36]. Tang Guotao et al. conducted rooting experiments using basal media such as 1/4 MS, 1/2 MS, MS, and B5, supplemented with NAA, IBA, IAA, GA_3_, and 2,4-D. However, after numerous trials with multiple combinations, they failed to identify a suitable medium for oil-tea camellia rooting [18]. In response to the difficulty of in vitro plantlets rooting on conventional rooting media, scholars have investigated various methods to promote rooting. Among these, the “two-step method” is a common approach. It entails initially inducing the plantlets in a high-concentration auxin medium for a short period, then transferring them to a hormone-free medium to facilitate root system development. Alternatively, a bottle-free rooting method involves treating the base of the plantlets with an auxin solution and directly transplanting them for rooting [37]. Studies have shown that the conventional in-bottle rooting method often yields a low rooting rate, and the resulting root systems tend to be fragile, leading to poor transplant survival rates. In contrast, the ex vitro rooting method generally achieves a higher rooting rate, produces stronger and more robust roots, and demonstrates better transplant survival performance [38]. To address the difficulty of in vitro rooting also observed in C. chekiangoleosa, and considering its large, leathery leaves and sturdy stems which make the plantlets less prone to wilting during transplantation, this study adopted a “pre-treatment with auxin in vitro followed by ex vitro transplant rooting” approach, which effectively promoted rooting. This method effectively promoted rooting. It synchronizes rooting and acclimatization, is operationally simple, avoids root damage during transplantation, allows the new root system to integrate closely with the substrate, and results in a high survival rate. The concentration ratio of auxins during the pre-rooting stage is also crucial, as levels that are either too high or too low can adversely affect the rooting rate. This study found that the highest rooting rate was achieved when in vitro plantlets were cultured for 30 days in a medium supplemented with 1.0 mg/L NAA and 2.0 mg/L IBA, followed by a dip in a 1.0 g/L ABT solution (Figure 3; Table 5). This is consistent with the optimal rooting hormone ratio reported in studies on ‘Xiaoguo Youcha’ [39]. During the rooting process of white-flowered oil-tea camellia tissue culture seedlings, the addition of NAA promotes the formation and growth of primary roots, while supplementing with IBA or IAA further stimulates the development of lateral roots and enhances overall root density [40]. Furthermore, during the in-bottle pre-rooting culture stage, stems and petioles become pigmented, and the young stem tissues become more compact, achieving a higher degree of lignification within a short period. This enhances the stress tolerance of the seedlings and promotes rooting. Meanwhile, the acclimatized plantlets exhibit improved transpiration resistance, enhanced adaptability, significantly better photosynthetic performance, and stronger capacity to adapt to external environments [41], and dipping them in ABT rooting powder containing NAA and IAA as active ingredients can enhance the effectiveness of ex vitro rooting [42].
The stem serves as the most common cutting material in plant propagation, and its anatomical structure directly influences rooting capacity. Thus, the anatomical structure of the stem, along with root primordium types and rooting patterns, constitutes a key research focus in cutting anatomy [43]. The stem segment of C. chekiangoleosa primarily consists of the periderm, cortex, phloem, vascular cambium, xylem, and pith, with a ring of phloem fibers located between the cortex and phloem (Figure 3B) [44]. Some studies suggest that this fibrous layer physically obstructs the growth of root primordia; while others indicate that adventitious roots can still readily form if the tissue is non-continuous [45]. Therefore, this study aimed to elucidate its specific role. We found that cuttings lack preformed root primordia and possess a ring of sclerenchyma cells within the periderm. After formation, the root primordia must sequentially penetrate the phloem and breach this sclerenchymatous layer to emerge and form adventitious roots, which is likely a primary reason for their relatively long rooting cycle (approximately 60 days) (Figure 3B). Based on the timing of their formation, adventitious roots are classified as either latent or induced root primordia [46]. This study confirmed that C. chekiangoleosa cuttings possess no latent root primordia prior to insertion, and their adventitious roots originate entirely from primordia induced after cutting; therefore, they are classified as the induced-rooting type. These primordia originated primarily from the vascular cambium and phloem parenchyma, confirming a phloem-rooting pattern (Figure 3G–I). Furthermore, although callus formation is not directly linked to adventitious root production and occurs earlier than root primordium initiation, it plays a significant role in preventing pathogen invasion and reducing the loss of essential substances from the cuttings. Particularly during summer propagation, callus effectively inhibits cutting decay and serves as a temporary bridge for water and nutrient transport, thereby supporting the rooting process [47].
4. Materials and Methods
4.1. Plant Materials
The experimental material consisted of the provincial-level registered superior cultivar C. chekiangoleosa ‘Ganhongyou 1’ (Figure 4), selected by the Jiangxi Academy of Forestry. Explants were collected from healthy, semi-lignified branches with axillary buds taken from the base of superior trees located in Nanchang City, Jiangxi Province, China (28°41′ N, 115°51′ E). Tissue-cultured plantlets obtained after the initial induction of these explants were used as the study material.
4.2. Chemicals and Reagents
The chemicals and reagents used in this study are detailed in Table 6, which lists their full names, abbreviations, manufacturers, and specific compositions. All plant growth regulators, including 6-BA, IAA, NAA, IBA, and GA_3_, were obtained from Shanghai Zhihua Chemistry Technology Co., Ltd. (Shanghai, China). The rooting powder (ABT) was provided by Aibid Biotechnology Co., Ltd. (Beijing, China), with a total active ingredient content of 50% (20% NAA and 30% IAA). The bactericide, composed of 70% thiophanate-methyl and 30% thiram, was purchased from Zhongke Qiyuan Technology Co., Ltd. (Beijing, China), while the Hyponex fertilizer (N:P:K = 7:6:19) was sourced from Zhongda Yucheng Technology Co., Ltd. (Quanzhou, China).
4.3. Culture Conditions
All culture media contained 30 g/L sucrose and 7.0 g/L agar, with the pH adjusted to 5.8–6.0. Gibberellic acid (GA_3_) and the bactericide were filter-sterilized through a 0.22 μm membrane (Hangzhou Tangwei Filter Materials Co., Ltd., Hangzhou, China) and added aseptically to the medium after autoclaving, when it had cooled to 50–60 °C. The other plant growth regulators were added directly to the medium prior to autoclaving. The culture conditions were as follows: temperature, (25 ± 2) °C; light intensity, 2500 lux; and photoperiod, 14 h per day (14 h light/10 h dark cycle). For ex vitro rooting, the substrate was sprayed with a 500–600-fold dilution of 70% thiophanate-methyl wettable powder and left to stand for three days before use. Environmental conditions were controlled at 20–28 °C (day)/15–20 °C (night), 70–80% humidity, and 5000–10,000 Lux light intensity provided by 75% shading net. Irrigation was applied every 3–4 days.
4.4. Explant Sterilization
Newly sprouted semi-lignified shoots were collected on sunny days between June and July. After being transported to the laboratory wrapped in moist towels, the leaves were removed, and nodal segments containing axillary buds were excised. The segments were then scrubbed with a cleaning solution to remove surface contaminants, rinsed thoroughly with clean water, and subjected to seven different surface sterilization treatments. Following sterilization, damaged tissues were trimmed off, and the explants were inoculated onto the culture medium. Each treatment consisted of 30 bottles, replicated three times, with contamination and induction rates recorded ultimately. The seven disinfection methods applied were as follows: (i) Immersion in 75% ethanol for 30 s, followed by sterilization with 0.1% HgCl_2_ for 7–10 min; (ii) Immersion in 75% ethanol for 30 s, followed by sterilization with 20% sodium hypochlorite for 7–10 min; (iii) Wiping the stem segments repeatedly with 75% ethanol, rinsing under running water for 2–3 h, then immersing in 75% ethanol for 30 s, and finally sterilizing with 0.1% HgCl_2_ for 7–10 min; (iv) Hardwood branches were inserted into a cardboard box filled with moist river sand, and newly sprouted lateral buds were used as experimental materials, which were then treated according to Method iii; (v) Immersion in 75% ethanol for 30 s, rinsing with sterile water, repeating one additional ethanol immersion step, followed by sterilization with 0.1% HgCl_2_ for 7–10 min, and a final rinse with sterile water before repeating the disinfection cycle once; (vi) Immersion in 75% ethanol for 30 s, followed by sterilization with 0.1% HgCl_2_ for 7–10 min, supplemented with composite disinfection using 2.5% sodium hypochlorite for 7–10 min; and (vii) Following the disinfection procedure of Method vi, the bud induction medium was supplemented with 1 mL/L of bactericide.
4.5. Screening of Bud Induction Medium Concentration
The Hyponex basal medium powder, with an N-P-K ratio of 7:6:19, was used as the source of macroelements. Nodal segments with buds of ‘Ganhongyou 1’ were inoculated onto media containing Hyponex at concentrations of 0.5, 1.0, 1.5, 2.0, 2.5, and 4.0 g/L. All media were uniformly supplemented with the following components: 30 g/L sucrose as the carbon source, 7 g/L agar as the solidifying agent, and 1.0 mg/L 6-BA along with 0.01 mg/L NAA as plant growth regulators [18,19]. Each culture bottle was inoculated with one nodal segment. Each treatment included 30 bottles and was replicated three times. The axillary bud germination rate was assessed after 30 days of culture. The axillary bud induction rate was calculated as the percentage of segments with sprouted axillary buds relative to the total number of inoculated segments.
4.6. Optimization of PGR Combinations for Bud Induction
Using the modified Hyponex medium (2 g/L) as the basal medium, a total of twelve different plant growth regulator combinations were established, involving 6-BA (0.5, 1.0 mg/L), IAA (0, 0.1, 0.2 mg/L), IBA (0, 0.1, 0.2 mg/L), and NAA (0, 0.1, 0.2 mg/L). All media were supplemented with 30 g/L sucrose and 7 g/L agar, with the pH adjusted to 5.6. Following initial disinfection, the explants were inoculated, placing one nodal segment per culture bottle. Each treatment consisted of 30 bottles and was replicated three times. The axillary bud germination rate was recorded after 30 days of culture.
4.7. Shoot Proliferation Culture
Aseptic shoots from the initial culture were proliferated on the modified Hyponex medium. The experimental comprised four groups (A–D) designed to screen for optimal auxin type, auxin concentration, 6-BA concentration, and the best overall proliferation medium combination. Each treatment included five bottles. Cultures were initiated with two axillary buds per bottle and subcultured three times using four shoot clusters per bottle. The proliferation coefficient (final shoot number/initial shoot number) was recorded after 30 days of the third subculture cycle when growth had stabilized; shoot growth status was concurrently observed.
4.8. Rooting Pre-Treatment and Ex Vitro Rooting
Following proliferation, axillary shoots (3–4 cm tall) were selected, excised at the base, and transferred to a pre-rooting medium based on 1/2-strength Hyponex medium. The experiment comprised nine different plant growth regulator combinations, involving NAA (0.5, 1.0, 2.0 mg/L) and IBA (0.5, 1.0, 2.0 mg/L). For each treatment, 10 shoots were inoculated per bottle, with five bottles per replicate and three replicates. After 30 days, plantlets were removed, and the medium at the base was carefully washed off. The basal ends were then dipped in a 1.0 g/L ABT solution before being transplanted into seedling bags (6 cm × 8 cm) containing a mixture of 65% peat, 10% perlite, 10% coconut coir, 10% rice husks, and 5% plant ash. After transplanting, the substrate was thoroughly watered. A fungicide (Thiophanate-methyl, Hulian Bio-Pharmaceutical (Xiayi) Co., Ltd., Shanghai, China) was applied, and the plantlets were covered with a plastic film to maintain humidity. The film was periodically removed for ventilation. The transplant survival rate, defined as the percentage of plantlets that developed new roots relative to the total number transplanted, was assessed after two months.
4.9. Data Analysis
All data were subjected to one-way analysis of variance (ANOVA) using SPSS19.0. The assumption of homogeneity of variances was verified using Levene’s test. When a significant overall effect was detected (p < 0.05), Tukey’s honestly significant difference (HSD) post hoc test was applied for multiple comparisons among treatment means. Data in tables are presented as the mean ± standard deviation (SD). Different lowercase letters indicate statistically significant differences at p < 0.05 among treatments. The primary dataset from all screening experiments is provided in Table S1.
5. Conclusions
This study successfully established a complete and efficient micropropagation system for the provincial-level registered superior cultivar C. chekiangoleosa ‘Ganhongyou 1’. We systematically optimized the key stages: bud induction (86.67% rate on a medium of 2.0 g/L Hyponex, 1.0 mg/L 6-BA, and 0.2 mg/L IAA), shoot proliferation (6.53 proliferation coefficient with 2 g/L Hyponex, 1.0 mg/L 6-BA, 0.15 mg/L IBA, and 0.5 mg/L GA_3_), and rooting–acclimatization via an innovative two-step strategy (in vitro pre-culture + ex vitro treatment), which achieved an 88% transplant survival rate. Anatomical observations further elucidated the rooting mechanism: C. chekiangoleosa exhibits an induced-rooting type, with root primordia originating mainly from phloem parenchyma cells adjacent to the vascular cambium, representing a typical phloem-rooting pattern. This system effectively addresses common challenges in oil-tea camellia tissue culture, such as high contamination rates, severe browning, and rooting difficulties, thereby providing a reliable technical foundation for the large-scale production of this valuable species.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gao C. Yang R. Yuan D. Characteristics of Developmental Differences between Fertile and Aborted Ovules in Camellia oleifera J. Am. Soc. Hortic. Sci.201714233033610.21273/JASHS 04164-17 · doi ↗
- 2Zhang M. Wang A. Qin M. Qin X. Yang S. Su S. Sun Y. Zhang L. Direct and Indirect Somatic Embryogenesis Induction in Camellia oleifera Abel Front. Plant Sci.20211264438910.3389/fpls.2021.64438933841471 PMC 8034400 · doi ↗ · pubmed ↗
- 3Shen T. Huang B. Xu M. Zhou P. Ni Z. Gong C. Wen Q. Cao F. Xu L.-A. The Reference Genome of Camellia chekiangoleosa Provides Insights into Camellia Evolution and Tea Oil Biosynthesis Hortic. Res.20229 uhab 08310.1093/hr/uhab 08335039868 PMC 8789033 · doi ↗ · pubmed ↗
- 4Wei T. Dong L. Zhong S. Jing H. Deng Z. Wen Q. Li J. Chemical Composition of Camellia chekiangoleosa Hu. Seeds during Ripening and Evaluations of Seed Oils Quality Ind. Crops Prod.202217711449910.1016/j.indcrop.2021.114499 · doi ↗
- 5Cicero A.F.G. Derosa G. Pisciotta L. Barbagallo, C., on behalf of the SISA-PUFACOL Study Group Testing the Short-Term Efficacy of a Lipid-Lowering Nutraceutical in the Setting of Clinical Practice: A Multicenter Study J. Med. Food 2015181270127310.1089/jmf.2015.002426274827 PMC 4638196 · doi ↗ · pubmed ↗
- 6Sun Y. Gao L. Hou W. Wu J. β-Sitosterol Alleviates Inflammatory Response via Inhibiting the Activation of ERK/P 38 and NF-κ B Pathways in LPS-Exposed BV 2 Cells Bio Med Res. Int.20202020753230610.1155/2020/753230632596368 PMC 7273476 · doi ↗ · pubmed ↗
- 7Wang Z.-X. Wang Y.-Y. Evaluation of the Provincial Competitiveness of the Chinese High-Tech Industry Using an Improved TOPSIS Method Expert Syst. Appl.2014412824283110.1016/j.eswa.2013.10.015 · doi ↗
- 8Zhong S. Huang B. Wei T. Deng Z. Li J. Wen Q. Comprehensive Evaluation of Quality Characteristics of Four Oil-Tea Camellia Species with Red Flowers and Large Fruit Foods 20231237410.3390/foods 1202037436673466 PMC 9857641 · doi ↗ · pubmed ↗