Decoding the Liver–Blood Partitioning of Neonicotinoid Insecticides: Evidence from Paired Human Liver and Blood Biomonitoring
Jiaqi Shao, Tingna Chen, Yihan Li, Wenfei Yu, Hangbiao Jin, Qinghua Zhou, Yuanchen Chen

TL;DR
This study examines how neonicotinoid insecticides distribute between human blood and liver tissues, finding significant differences and associations with liver cancer.
Contribution
The study provides novel evidence on liver–blood partitioning of neonicotinoids in humans and links hepatic concentrations to liver cancer status.
Findings
Neonicotinoids were detected in 84.6% of blood and 87.2% of liver samples, with dinotefuran being the most abundant compound.
Blood-to-liver partition ratios varied with physicochemical properties and were lower in liver cancer patients.
Hepatic concentrations of specific neonicotinoids were statistically associated with liver cancer status after adjusting for covariates.
Abstract
Neonicotinoids (NEOs) are among the most widely used insecticides worldwide, and their increasing detection in environmental and human matrices has raised concerns about chronic exposure and potential health risks. However, human data on target-organ burdens and liver–blood partitioning of NEOs remain unclear. Here, we quantified nine NEOs in paired liver tissue and whole-blood samples from 234 individuals to characterize internal distribution patterns and liver–blood partitioning of NEOs in humans. Samples included both liver cancer patients and non-liver cancer individuals, enabling exploratory evaluation of disease-related differences. At least one NEO was detected in 84.6% of blood samples and 87.2% of liver samples, with median concentrations ranging from 0.15–3.52 ng/mL in blood and 0.39–10.99 ng/g in liver, respectively. Dinotefuran was the most abundant compound in both…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
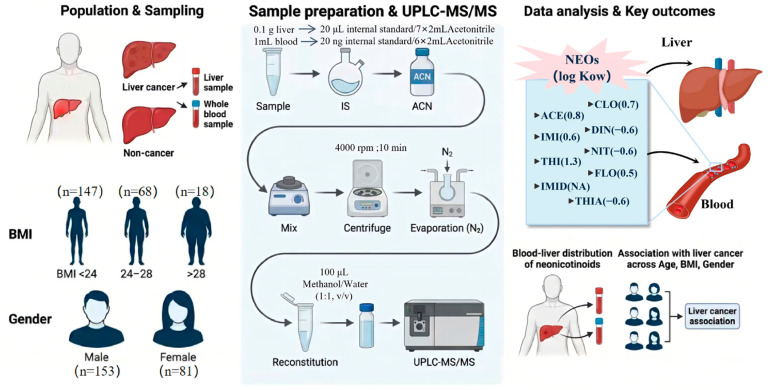 Figure 1
Figure 1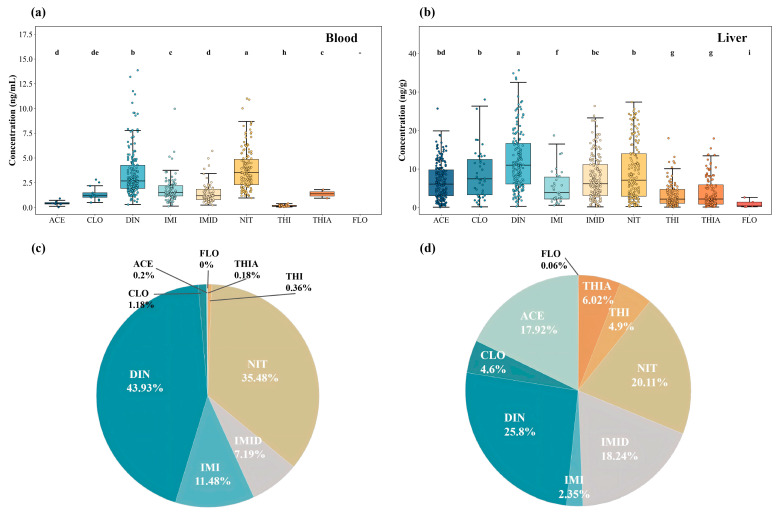 Figure 2
Figure 2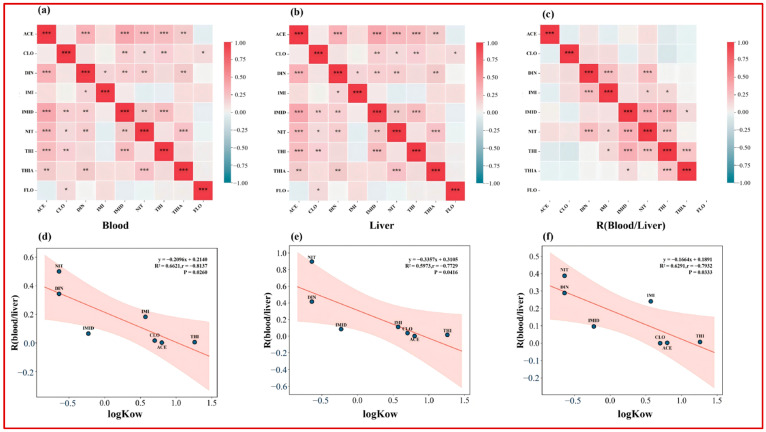 Figure 3
Figure 3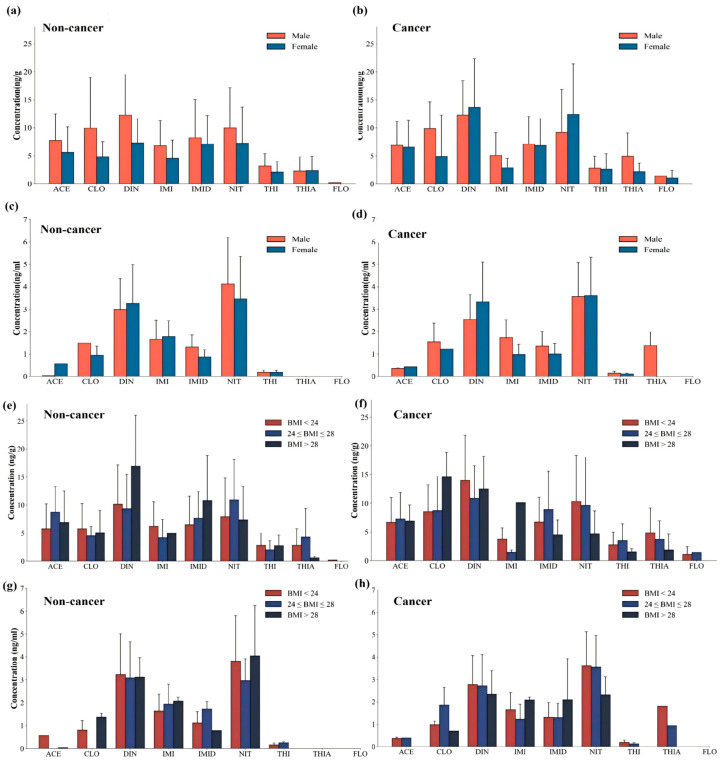 Figure 4
Figure 4- —National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect and Pesticide Research · Pesticide Residue Analysis and Safety · Insects and Parasite Interactions
1. Introduction
Neonicotinoid insecticides (NEOs) are systemic insecticides with nicotine-like structures and are widely used in agriculture and public health vector control due to their high efficacy and broad-spectrum activity [1,2,3,4]. With their extensive and long-term use, NEOs and their metabolites have been increasingly detected in environmental media and human biological samples, including surface waters, drinking water, urine, blood, and serum [5,6,7,8]. This widespread occurrence has raised ongoing concerns about chronic background exposure in the general population [3,9,10].
To date, most population-based biomonitoring studies have relied on easily accessible biological matrices—most commonly urine, serum, or whole blood—to estimate internal exposure to NEOs [5,6,11,12,13,14]. However, these matrices primarily reflect relatively recent intake and circulating levels and may not adequately capture target-organ burdens [15]. As a result, reliance on a single circulating matrix may lead to exposure misclassification when evaluating organ-specific health effects [16].
The liver plays a central role in xenobiotic uptake, biotransformation, and clearance and is a major target organ for the toxic effects of many environmental contaminants [17]. For several classes of pollutants, including per- and polyfluoroalkyl substances (PFAS), persistent organic pollutants (POPs), and metals, paired liver–blood measurements in humans have demonstrated pronounced compound-specific partitioning and substantial divergence between circulating levels and hepatic burdens [18,19,20]. These studies collectively indicate that liver–blood partitioning is governed by physicochemical properties and protein-binding characteristics, and that blood-based metrics alone may underestimate target-organ exposure [3,21].
In contrast, direct evidence on liver–blood partitioning of neonicotinoid insecticides in humans is still lacking. Although experimental studies have demonstrated hepatic uptake, distribution, and metabolism of NEOs [22,23,24,25,26,27], human data have been largely limited to single-matrix biomonitoring studies using urine or blood [11,12,13,14,28]. Whether blood-measured NEO concentrations reflect true hepatic burdens—and whether individual NEOs exhibit compound-specific partitioning patterns in humans—therefore remains unresolved.
In parallel, increasing attention has been directed toward potential liver-related health effects of NEO exposure. Toxicological studies suggest that several NEOs can induce oxidative stress, metabolic disruption, and inflammatory responses in hepatic tissue [22,23,24]. Population-level evidence has also linked NEO exposure to alterations in liver function indicators and metabolic outcomes [29]. However, without target-organ-specific exposure measurements, interpretation of such associations remains challenging, and the compounds most relevant to hepatic burden and liver-related outcomes may be misidentified [30].
Accordingly, the primary objective of this study was to establish a paired liver–blood biomonitoring framework for neonicotinoid insecticides in humans and to characterize their internal distribution and partitioning behavior. Using paired whole-blood and liver tissue samples from 234 individuals, we quantified nine individual NEOs (imidacloprid (IMI), acetamiprid (ACE), thiamethoxam (THIA), clothianidin (CLO), thiacloprid (THI), flonicamid (FLO), nitenpyram (NIT), imidaclothiz (IMID), and dinotefuran (DIN)), examined detection frequencies, concentration levels, and compositional differences across matrices, and calculated blood-to-liver partition ratios (R_B/L) to evaluate physicochemical determinants of internal distribution [30,31,32]. In addition, we explored whether liver–blood partitioning patterns and hepatic burdens differed between liver cancer patients and non-liver cancer individuals, providing organ-oriented context for interpreting epidemiological associations [33,34,35,36].
By explicitly integrating target-organ measurements with circulating biomarkers, this study aims to improve understanding of NEO internal disposition and to strengthen the interpretative framework for evaluating liver-related health outcomes associated with neonicotinoid exposure [16,30].
2. Methods and Materials
2.1. Study Population and Sample Collection
Between 2019 and 2021, we collected 234 paired liver tissue and whole-blood samples at the First Affiliated Hospital, Zhejiang University School of Medicine. The cohort included liver cancer patients (n = 125) and non-liver cancer participants (n = 109). To better represent contaminant occurrence in non-diseased hepatic tissue, normal liver tissue rather than tumor tissue was collected in this study. All participants provided written informed consent, and the study protocol was approved by the Clinical Research Ethics Committee of the First Affiliated Hospital, Zhejiang University School of Medicine. Demographic information was extracted from hospital medical records, including sex, age, body weight, height, and residential address (age groups: <40, 40–50, 51–60, and >60 years). The age of the participants ranged from 16 to 86 years; detailed sample size information is provided in Table S1. Disease status (liver cancer vs non-liver cancer) was considered in the subsequent analyses.
Liver tissue samples were collected immediately after hepatic resection and placed into polypropylene (PP) tubes, then temporarily stored on dry ice. At the same time, 10–15 mL of paired whole blood was collected into EDTA-containing BD Vacutainer^®^ tubes (Becton, Dickinson and Company, Franklin Lakes, NJ, USA). We selected whole blood as the circulating matrix because it includes both the plasma and cellular fractions (primarily red blood cells), and thus can better reflect total circulating burdens for compounds that partition into blood cells. For NEOs, compound-specific partitioning between plasma and red blood cells has been reported in humans, with THIA showing predominant presence in red blood cells while several other NEOs (e.g., IMI and DIN) are mainly detected in plasma. Accordingly, whole-blood measurements may reduce matrix-driven bias for compounds with non-negligible cellular partitioning, but the resulting concentrations may not be directly comparable to values reported in serum- or plasma-based biomonitoring studies. Field blanks were prepared during sampling using ultrapure water (5.0 mL, n = 3). All samples were subsequently stored at −80 °C until analysis. An overview of the methodological workflow is illustrated in Figure 1.
2.2. Sample Preparation
For liver tissue samples, 0.1 g of freeze-dried liver tissue was ground into a fine powder and spiked with 20 μL of an isotopically labeled internal standard mixture (10 ppm), including ACE-d_3_, IMI-d_4_, CLO-d_3_, DIN-d_3_ and THIA-d_3_. Subsequently, 7 mL of acetonitrile was added as the extraction solvent. The mixture was vortexed for 2 min, shaken for 30 min, and ultrasonicated for 30 min, followed by centrifugation at 4000 rpm for 10 min. The supernatant was collected into a new 15 mL centrifuge tube. The residue was re-extracted with an additional 7 mL of acetonitrile under the same conditions, and the two supernatants were combined and evaporated to dryness under a gentle nitrogen stream. The residue was reconstituted with 100 μL of methanol/water (1:1, v/v), centrifuged at 13,000 rpm for 8 min, and the supernatant was transferred to amber autosampler vials for UPLC–MS/MS analysis.
The extraction procedure for whole-blood samples was similar to that for liver tissue, with minor adjustments in sample amount, solvent volume, and internal standard addition. Briefly, after thawing, 1 mL of whole blood was transferred into a 15 mL centrifuge tube and spiked with 200 ng of the isotopically labeled internal standard mixture (ACE-d_3_, IMI-d_4_, CLO-d_3_, DIN-d_3_, and THIA-d_3_). Then, 6 mL of acetonitrile was added for extraction. The samples were processed using the same vortexing, shaking, and ultrasonication conditions, followed by centrifugation at 4000 rpm for 10 min. The supernatant was collected, and the residue was re-extracted with an additional 6 mL of acetonitrile. The combined supernatants were evaporated to dryness under nitrogen, reconstituted with 100 μL of methanol/water (1:1, v/v), centrifuged at 13,000 rpm for 8 min, and the final supernatant was transferred to amber autosampler vials for instrumental analysis.
2.3. Instrumental Analysis
Quantification of nine target compounds—ACE, IMI, CLO, THIA, FLO, THI, NIT, DIN, and IMID—was performed using ultra-performance liquid chromatography coupled with tandem triple quadrupole mass spectrometry (UPLC–MS/MS; Xevo TQ-S Waters Corporation, Milford, MA, USA). Detailed information on the instrumental parameters can be found in Supporting Information.
2.4. Quality Assurance and Quality Control
During sample analysis, one acetonitrile blank was inserted after every eight samples to minimize potential carryover and cross-contamination between consecutive injections. The limits of detection (LOD) and limits of quantification (LOQ) for the nine neonicotinoid insecticides were defined as concentrations corresponding to signal-to-noise ratios (SNRs) of 3 and 10, respectively. LODs and LOQs were determined based on three independent replicate measurements, and the mean values were used.
For the nine neonicotinoid insecticides, The LOD ranges in liver tissue and whole blood were 0.009–0.027 ng/g and 0.009–0.021 ng/mL, respectively, while the corresponding LOQ ranges were 0.027–0.083 ng/g and 0.026–0.064 ng/mL.
In addition, recovery experiments were conducted by spiking randomly selected liver and blood samples with known concentrations of mixed standard solutions and isotopically labeled internal standards, followed by analysis under the same sample preparation and instrumental conditions. The recoveries of neonicotinoid insecticides ranged from 74.1% (THIA) to 112% (DIN) in liver samples and from 85.6% (THI) to 105% (ACE) in blood samples. All analyte recoveries met analytical quality requirements, indicating that the sample preparation procedures and UPLC–MS/MS conditions used in this study were appropriate and reliable.
2.5. Calculation of Blood-to-Liver Partition Ratio
The blood-to-liver partition ratio (R_B/L) was calculated to characterize the relative distribution of individual NEOs between paired whole blood and liver tissue as follows:
where and represent the concentrations of individual NEOs in paired whole blood (ng/mL) and liver (ng/g), respectively. R_B/L values were calculated only when both blood and liver concentrations exceeded three times the LOD to reduce uncertainty near the detection limit.
2.6. Statistical Analysis
All statistical analyses were conducted using Python (v3.9). Data processing and management were performed using pandas (v1.3.5) and NumPy (1.21.6). Nonparametric tests and correlation analyses were implemented using SciPy (v1.7.3; scipy.stats). Logistic regression models were fitted using statsmodels (v0.13.2; Logit). Figures were generated using matplotlib (v3.4.3) and seaborn (v0.11.2). Continuous variables were assessed for distributional characteristics. Because NEO concentrations were right-skewed, nonparametric methods were applied. Differences between two independent groups were evaluated using the Mann–Whitney U test, and comparisons among more than two groups were assessed using the Kruskal–Wallis test. Correlations between continuous variables were assessed using Spearman’s rank correlation coefficient (ρ). Concentrations below the LOD were treated as non-detects and substituted with LOD/2 prior to subsequent analyses. ∑NEOs were calculated within each matrix as the sum of individual compounds after LOD substitution. For logistic regression analyses, NEO concentrations were entered as continuous predictors after natural log-transformation [ln(x)] to reduce right-skewness, limit the influence of extreme values, and improve model stability. Binary logistic regression was used to evaluate associations between NEO concentrations and liver cancer status, reporting odds ratios (ORs) with 95% confidence intervals (CIs). Both crude and multivariable-adjusted models were fitted; adjusted models included age, sex, and body mass index (BMI) as covariates. Values substituted as LOD/2 were included prior to ln-transformation. The R_B/L was calculated only when both liver and blood concentrations exceeded three times the LOD to reduce uncertainty near the detection limit; observations not meeting this criterion were treated as missing for partitioning analyses. All statistical tests were two-sided, and p < 0.05 was considered statistically significant. Where applicable, multiple-comparison adjustment was applied as specified in the corresponding analyses.
3. Result and Discussion
3.1. Levels and Composition of Neonicotinoids in Whole Blood and Liver
Paired whole-blood and liver tissue samples from 234 participants were analyzed. FLO was largely undetected (<LOD in most samples) and is therefore shown in figures and tables for completeness only; FLO was not included in inferential analyses and was not used to support interpretive conclusions throughout the Results and Discussion. In whole blood, at least one NEO was detected in 84.6% of samples. The mean and median ∑NEOs concentrations were 6.52 ± 4.44 ng/mL and 6.34 ng/mL, respectively, with concentrations ranging from non-detectable levels to 19.81 ng/mL (Table 1).
At the compound-specific level, the exposure composition in whole blood was highly uneven (Figure 2a,c). DIN and NIT dominated the mixture, accounting for 43.93% and 35.48% of ∑NEOs, respectively. DIN exhibited the highest detection frequency (84.6%) with a median concentration of 2.68 ng/mL. NIT was detected in 59.4% of samples and showed a median concentration of 3.52 ng/mL, contributing disproportionately among individuals with higher overall blood burdens. IMI and IMID contributed at intermediate levels (11.48% and 7.19%), with detection frequencies of 40.6% and 32.1% and median concentrations of 1.52 ng/mL and 1.19 ng/mL, respectively. The remaining compounds (ACE, CLO, THI, and THIA) were generally detected at lower frequencies or concentrations, and FLO was not detected in any whole-blood sample (Figure 2a).
Compared with previous biomonitoring studies, whole-blood concentrations of several high-frequency compounds were notably elevated. Earlier studies conducted in Guangdong Province reported mean DIN concentrations of 0.56 ng/mL and 0.95 ng/mL among students and individuals with obesity, respectively [12,37,38], whereas mean DIN concentrations in both liver cancer and non-liver cancer populations in the present study exceeded 3 ng/mL. Similarly, the mean IMI concentration in whole blood (1.8–2.0 ng/mL) was substantially higher than the previously reported level of approximately 0.21 ng/mL. These interstudy differences may reflect regional variation in crop structure and pesticide-use patterns, as well as differences in study populations, sampling periods, and analytical protocols [38,39,40].
In contrast to whole blood, neonicotinoid residues were substantially elevated in liver tissue. The mean hepatic concentration of ∑NEOs was 32.14 ± 19.61 ng/g, with a median of 32.69 ng/g, representing an approximately fivefold increase relative to whole-blood levels and a broader interindividual distribution. Hepatic ∑NEOs concentrations ranged from non-detectable levels (<LOD) to 88.45 ng/g (Table 1).
The compositional profile of neonicotinoids in liver tissue differed markedly from that in whole blood (Figure 2b,c). DIN, NIT, and IMID constituted the major contributors, accounting for 25.8%, 20.1%, and 18.2% of ∑NEOs, respectively. Notably, the contribution of ACE increased to approximately 18.0% in liver tissue, compared with only 3.0% in whole blood (Figure 2c). As illustrated in Figure 2d, the mean compositional structure of ∑NEOs in liver tissue was characterized by a more even distribution across multiple compounds than that observed in whole blood, with ACE, IMID, and THIA contributing substantially to the overall hepatic burden. DIN, NIT, and IMID also maintained high detection frequencies and concentration levels in liver tissue, with median concentrations of 10.99, 7.10, and 6.15 ng/g, respectively (Figure 2b).
Stratified analysis showed higher hepatic burdens among liver cancer patients than among non-liver cancer individuals. Notably, this comparison is cross-sectional and case–control in nature; therefore, the observed differences should be interpreted as associations rather than evidence of causation. The median hepatic concentration of ∑NEOs in the liver cancer group was 37.97 ng/g, exceeding that observed in the non-liver cancer group (32.53 ng/g). In liver cancer patients, detection frequencies of ACE, IMID, DIN, and NIT all exceeded 70%, with ACE detected in 89.6% of samples, substantially higher than the corresponding frequency in non-liver cancer individuals (77.1%) (Figure S1). As summarized in Figure S1, between-group contrasts were more evident in liver tissue than in whole blood, supporting the emphasis on hepatic burdens when interpreting liver cancer-related patterns. Because liver cancer may alter hepatic metabolism, transport processes, and blood flow, and may also involve treatment-related changes, disease-related shifts in liver–blood partitioning could contribute to the observed hepatic patterns; reverse causality cannot be excluded.
Previous studies on neonicotinoids in liver tissue have predominantly relied on animal exposure experiments, and population-level human evidence remains limited [16,25,26,27]. Here, we provide a systematic characterization of neonicotinoid residue levels and compositional profiles in human liver tissue based on paired biomonitoring. Given the liver’s central role in xenobiotic uptake, biotransformation, and clearance, and the fact that blood represents a dynamic circulating compartment, concurrent detection in both matrices is compatible with the presence of detectable parent compounds and/or metabolites in vivo and with measurable hepatic tissue burdens [16,41,42].
To further characterize the mixed-exposure structure, Spearman correlation analyses were conducted among individual neonicotinoids in whole blood and liver tissue (Figure 3a,b). In whole blood, significant positive correlations were observed for a limited number of compound pairs, mainly involving high-frequency analytes (e.g., DIN–IMI, DIN–THI, IMI–NIT, and IMI–THI). In liver tissue, correlations were more pervasive, with significant co-variation observed across multiple compound pairs involving ACE, DIN, IMID, NIT, THI, and THIA (p < 0.01). Overall, the broader correlation structure observed in liver tissue suggests that hepatic measurements integrate co-exposure signals while also reflecting compound-specific uptake, retention, and metabolism, consistent with previous human biomonitoring studies reporting correlated NEO profiles across biological matrices [12,28,41,43]. Taken together, the case–control differences and the subsequent hepatic association signals should be viewed as exploratory and hypothesis-generating. A primary implication is that reliance on blood-only biomarkers may lead to exposure misclassification when evaluating liver-related outcomes, underscoring the value of tissue-specific biomonitoring and internal partitioning information.
3.2. Liver–Blood Partitioning Characteristics Based on Population Biomonitoring
Previous studies, largely based on animal models or in vitro systems, have demonstrated that NEOs can enter the liver and undergo hepatic biotransformation [41]. Their internal distribution is therefore influenced not only by external exposure levels but also by molecular physicochemical properties and hepatic transport and metabolic processes [23,42,44,45]. Using paired whole-blood and liver tissue samples from the same individuals, we calculated blood-to-liver partition ratios (R_B/L) for individual NEOs to characterize their relative distribution between the circulating compartment and hepatic tissue.
Across the overall study population, R_B/L values varied substantially among compounds and were significantly negatively correlated with logKow (p = 0.026; Figure 2d). Compounds with lower hydrophobicity, such as DIN and NIT, exhibited higher median R_B/L values (approximately 0.5 and 1.5, respectively), indicating a relatively greater presence in whole blood [31]. In contrast, more hydrophobic compounds, including THI and ACE, showed markedly lower R_B/L values (approximately 0.09 and 0.07), reflecting a greater relative partitioning into liver tissue [32]. This compound-specific pattern is consistent with physicochemical property-dependent distribution, whereby increasing hydrophobicity favors association with lipid-rich phases and macromolecular binding within hepatic tissue, resulting in lower blood-to-liver ratios [23,45].
Liver–blood partitioning is unlikely to be determined by hydrophobicity alone, but rather reflects the combined influence of multiple processes, including membrane/lipid partitioning, binding to plasma and tissue macromolecules (i.e., the unbound fraction), ionization behavior, and transporter-mediated uptake and efflux [46,47]. Within certain chemical spaces, logKow may act as a proxy for factors that co-vary with lipophilicity [48], such as protein binding or hepatocellular binding capacity, and physicochemical properties including lipophilicity have been linked to interactions with hepatic uptake and efflux transporters, such as organic anion transporting polypeptides (OATPs) [49] and organic cation transporter 1 (OCT1) [50]. However, evidence for some neonicotinoids suggests that plasma protein binding does not systematically track lipophilicity, indicating that additional determinants beyond logKow contribute to tissue distribution. Because the present study did not directly measure plasma protein binding, tissue binding, transporter involvement, or intrahepatic metabolic profiles, the relative contributions of these mechanisms cannot be resolved. Therefore, the observed relationship between logKow and the liver-to-blood partition ratio should be interpreted as a descriptive physicochemical association rather than a causal mechanism, and as a hypothesis-generating basis for future mechanistic investigations.
Stratified analyses further showed that R_B/L values were generally lower in the liver cancer population than in non-liver cancer individuals (Figure S2). Consistently, the overall distribution of ΣNEOs exhibited a modest shift toward lower R_B/L in liver cancer patients (mean ± SD: 0.237 ± 0.234) compared with non-liver cancer individuals (0.287 ± 0.301), suggesting relatively greater hepatic enrichment. For DIN, the median R_B/L in liver cancer patients was 0.223 (IQR: 0.129–0.394), which was significantly lower than that observed in non-liver cancer individuals (median: 0.295; IQR: 0.188–0.714; Wilcoxon p = 0.009). NIT displayed a similar directional difference, with a lower median R_B/L in the liver cancer group (0.464) compared with the non-liver cancer group (0.646). Together, these results indicate an altered liver–blood partitioning pattern associated with liver cancer status, characterized by relatively lower blood-to-liver ratios and higher proportional hepatic burdens [28,36].
To explore whether shared determinants may underlie these partitioning patterns, we further examined the correlation structure among R_B/L values of individual NEOs using Spearman correlation analysis (Figure 3c). Several compounds exhibited significant positive correlations in their R_B/L values, suggesting partially shared factors influencing liver–blood distribution across NEOs. Overall, paired biomonitoring data support logKow-related liver–blood partitioning behavior of NEOs in humans and consistently indicate lower R_B/L values in liver cancer patients than in non-liver cancer individuals.
3.3. Sex- and BMI-Stratified Patterns of Neonicotinoid Burden
Using paired liver tissue and whole-blood samples from the same individuals, we examined sex- and BMI-stratified patterns of neonicotinoid burden. Sex differences were evaluated using the Mann–Whitney U test. BMI was analyzed both as a categorical variable (<24, 24–28, ≥28; Kruskal–Wallis test) and as a continuous variable using Spearman correlation analysis.
In liver tissue, sex differences were not observed for most analytes in the liver cancer population. However, among detected samples, THIA concentrations were significantly higher in males than in females (median: 3.08 vs. 1.85 ng/g, p = 0.028; Figure 4a,b). In the non-liver cancer population, males exhibited higher hepatic burdens for certain compounds: ACE concentrations were significantly higher in males than in females (median: 7.06 vs. 5.07 ng/g, p = 0.032; Figure 4a,b), and ∑NEOs were also elevated in males (38.10 vs. 30.66 ng/g, p = 0.027).
In whole blood, no significant sex differences were observed in the non-liver cancer population (p > 0.05). In contrast, sex-related differences were more evident in the liver cancer population. Among detected samples, males showed higher median IMI concentrations than females (1.66 vs. 0.91 ng/mL, p = 0.004; Figure 4c,d), and ∑NEOs were also higher in males (7.52 vs. 5.85 ng/mL, p = 0.045).
Sex-related differences in internal exposure have been reported inconsistently across previous neonicotinoid biomonitoring studies. Male-dominant patterns similar to those observed here have been reported in serum-based studies among elderly populations in southern China [51], whereas other studies have reported higher levels in females or no significant sex differences [12,52]. Such heterogeneity may reflect differences in exposure behaviors (e.g., occupational or agricultural contact, diet, and lifestyle) [53,54], physiological and toxicokinetic factors (e.g., body size, blood volume, hepatic blood flow, and sex hormone–regulated expression of CYP enzymes and transporters) [55,56], as well as methodological factors such as matrix selection and handling of non-detects. Notably, sex differences in whole blood were more pronounced in liver cancer patients than in non-liver cancer individuals, suggesting that disease status may modify systemic physiology and liver–blood exchange, thereby accentuating sex-specific contrasts for certain high-frequency compounds [57]. The determinants of these patterns remain uncertain and warrant confirmation in larger cohorts.
In liver tissue, BMI was associated with the burden of specific neonicotinoids. In the non-liver cancer population, ∑NEOs was positively correlated with BMI (p = 0.015), with median ∑NEOs increasing across BMI categories. Similarly, NIT concentrations were positively correlated with BMI in the non-liver cancer group (p = 0.040; Figure 4e). In contrast, THIA showed a negative correlation with BMI in both populations (non-liver cancer: p = 0.032; liver cancer: p = 0.028; Figure 4e,f), with lower post-detection median concentrations observed in individuals with obesity.
Group comparisons further indicated significant differences in hepatic ACE concentrations across BMI categories in the non-liver cancer population (Kruskal–Wallis p = 0.048; Figure 4g). The highest median ACE concentration was observed in the 24–28 BMI group (8.31 ng/g, IQR: 5.78–12.23), compared with the <24 group (4.51 ng/g, IQR: 1.82–8.62) and the ≥28 group (5.89 ng/g, IQR: 3.43–8.81).
By contrast, BMI was not consistently associated with ∑NEOs in whole blood (p > 0.05). Overall, these findings indicate that BMI-related variation in neonicotinoid burden was more evident in liver tissue than in whole blood. This pattern is consistent with the possibility that BMI may preferentially influence hepatic burden through alterations in the metabolic milieu of the liver—such as lipid accumulation, inflammatory status, intrahepatic blood flow, and transport processes—making such associations more detectable at the tissue level, while relationships in circulating blood appear less stable [58,59].
3.4. Liver Tissue-Based Associations Between Neonicotinoid Burden and Liver Cancer Status
To examine differences in NEO burden between liver cancer and non-liver cancer individuals, liver cancer status was specified as the outcome, and logistic regression models were constructed using quantitative measurements from whole blood (ng/mL) and liver tissue (ng/g), respectively. Crude odds ratios (ORs) and multivariable-adjusted ORs with 95% confidence intervals (CIs) were estimated (Table 2).
In whole blood, concentrations of individual NEOs were numerically higher in the liver cancer group; however, no compound showed a statistically significant association with liver cancer status in either crude or adjusted models (p > 0.05). Thus, within the present dataset, blood-based NEO concentrations did not provide statistically significant evidence for discrimination between liver cancer and non-liver cancer individuals.
In contrast, analyses based on liver tissue indicated more pronounced differences in NEO burden by disease status. Overall hepatic ∑NEOs concentrations were higher in liver cancer patients than in non-liver cancer individuals, with statistically significant differences observed for ACE, DIN, IMID, and THIA (p < 0.05; Table 2). Consistently, logistic regression analyses showed statistically significant associations between hepatic concentrations of ACE (OR = 0.75, 95% CI: 0.61–0.91, p = 0.004), DIN (OR = 0.83, 95% CI: 0.73–0.95, p = 0.007), IMID (OR = 0.84, 95% CI: 0.72–0.97, p = 0.019), and THIA (OR = 0.80, 95% CI: 0.67–0.95, p = 0.013) and liver cancer status. After adjustment for sex and BMI, these associations remained statistically significant: ACE (OR = 0.79, 95% CI: 0.64–0.99, p = 0.037), DIN (OR = 0.81, 95% CI: 0.70–0.93, p = 0.004), IMID (OR = 0.84, 95% CI: 0.71–0.98, p = 0.030), and THIA (OR = 0.79, 95% CI: 0.65–0.96, p = 0.017) (Table 2).
Overall, several NEOs exhibited statistically significant associations with liver cancer status when evaluated using hepatic tissue concentrations, whereas corresponding blood-based measurements did not. These findings highlight the importance of target-organ measurements for distinguishing internal exposure patterns by disease status [60,61,62]. From a biological perspective, the liver is the primary site of xenobiotic uptake, metabolism, and clearance, and toxicological studies have shown that multiple NEOs can induce oxidative stress, inflammatory responses, and metabolic perturbations in hepatic tissue, accompanied by altered biomarkers or histopathological changes [63]. This target-organ context may partly explain why disease-related differences were more evident in liver tissue than in whole blood [64].
Nevertheless, these associations should be interpreted cautiously. Liver cancer has multifactorial etiologies, and residual confounding by co-exposures, diet, pre-existing liver conditions, and treatment-related factors cannot be excluded. In addition, reverse causality and disease-related alterations in hepatic metabolism, transport, and chemical partitioning may influence tissue concentrations, complicating causal interpretation. Future studies incorporating mixture-based analytical approaches and sensitivity analyses, together with detailed clinical information on liver function, disease stage, and treatment history, will be essential for strengthening inference regarding neonicotinoid exposure and liver-related health outcomes [65,66].
4. Conclusions
This study reports population-based paired liver–blood biomonitoring data for NEOs from 234 individuals, providing direct evidence on internal distribution and liver–blood partitioning in humans. NEOs were frequently detected in both matrices, and hepatic ΣNEOs were markedly higher than whole-blood levels, indicating that blood measurements alone may underestimate target-organ burdens. Dinotefuran predominated in both matrices, yet mixture profiles differed between blood and liver, highlighting clear matrix-specific heterogeneity. R_B/L varied widely across compounds and showed a significant inverse relationship with logKow, consistent with physicochemical property-dependent partitioning in vivo. R_B/L values were generally lower in liver cancer patients, reflecting a shift toward hepatic enrichment under pathological conditions. In exploratory logistic regression models, several hepatic NEOs were associated with liver cancer status after adjustment for age, sex, and BMI, whereas corresponding blood-based metrics showed no comparable associations. Given the cross-sectional case–control design, these results are hypothesis-generating rather than causal and underscore the risk of exposure misclassification when liver-related outcomes are evaluated using blood-only biomarkers. Future work should prioritize longitudinal designs with richer clinical characterization and integrate mixture-aware analyses with toxicokinetic frameworks to clarify determinants of liver–blood partitioning and strengthen causal inference.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Jeschke P. Nauen R. Schindler M. Elbert A.J. Overview of the status and global strategy for neonicotinoids J. Agric. Food Chem.201159289729082056506510.1021/jf 101303 g · doi ↗ · pubmed ↗
- 2Chen Y. Yan C. Sun Z. Wang Y. Tao S. Shen G. Xu T. Zhou P. Cao X. Wang F.J.E.S. Organochlorine pesticide ban facilitated reproductive recovery of Chinese striped hamsters Environ. Sci. Technol.2021556140614910.1021/acs.est.1c 0016733797225 · doi ↗ · pubmed ↗
- 3Hladik M.L. Main A.R. Goulson D. Environmental risks and challenges associated with neonicotinoid insecticides Environ. Sci. Technol.2018523329333510.1021/acs.est.7b 0638829481746 · doi ↗ · pubmed ↗
- 4Thompson D.A. Lehmler H.-J. Kolpin D.W. Hladik M.L. Vargo J.D. Schilling K.E. Le Fevre G.H. Peeples T.L. Poch M.C. La Duca L.E. A critical review on the potential impacts of neonicotinoid insecticide use: Current knowledge of environmental fate, toxicity, and implications for human health Environ. Sci. Process. Impacts 2020221315134610.1039/c 9em 00586 b 32267911 PMC 11755762 · doi ↗ · pubmed ↗
- 5Zhang D. Lu S. Human exposure to neonicotinoids and the associated health risks: A review Environ. Int.202216310720110.1016/j.envint.2022.10720135344909 · doi ↗ · pubmed ↗
- 6Ospina M. Wong L.-Y. Baker S.E. Serafim A.B. Morales-Agudelo P. Calafat A.M. Exposure to neonicotinoid insecticides in the US general population: Data from the 2015–2016 national health and nutrition examination survey Environ. Res.201917610855510.1016/j.envres.2019.10855531288196 PMC 6710140 · doi ↗ · pubmed ↗
- 7Klarich K.L. Pflug N.C. De Wald E.M. Hladik M.L. Kolpin D.W. Cwiertny D.M. Le Fevre G.H. Letters T. Occurrence of neonicotinoid insecticides in finished drinking water and fate during drinking water treatment Environ. Sci. Technol. Lett.2017416817310.1021/acs.estlett.7b 00081 · doi ↗
- 8Berens M.J. Capel P.D. Arnold W.A. Neonicotinoid insecticides in surface water, groundwater, and wastewater across land-use gradients and potential effects Environ. Toxicol. Chem.202140101710333330118210.1002/etc.4959 PMC 8049005 · doi ↗ · pubmed ↗