Acomys cahirinus seurati as a Potential Reservoir Host of Leishmania major
Sergei Karlin, Barbora Bečvářová, Kamal Eddine Benallal, Tomáš Bečvář, Ghania Mezai, Mounir Zaabar, Mohamed Mefissel, Petr Volf, Jovana Sádlová

TL;DR
This study shows that Acomys cahirinus seurati can carry and spread Leishmania major, suggesting it may be a reservoir for cutaneous leishmaniasis in North Africa.
Contribution
The study experimentally confirms Acomys cahirinus seurati as a potential reservoir host for Leishmania major in North Africa.
Findings
Seven out of sixteen Acomys cahirinus seurati transmitted Leishmania major to sand flies between 10 and 25 weeks post-infection.
Parasites were mostly localized at the infection site with rare visceral spread.
Some animals transmitted parasites without visible skin lesions, indicating transmission is possible without ulceration.
Abstract
Cutaneous leishmaniasis (CL) caused by Leishmania major is a zoonotic disease transmitted by phlebotomine sand flies. Identification of reservoir hosts is critical for understanding transmission and guiding control. While Psammomys obesus and Meriones shawi are recognized reservoirs in North Africa, the role of other rodents remains unclear. We experimentally assessed the reservoir potential of Acomys cahirinus seurati from southeastern Algeria. Animals were intradermally infected, and clinical signs and infectiousness to sand flies were monitored. Parasite persistence in skin and visceral tissues was measured by quantitative PCR. All animals developed localized, self-limiting lesions. Seven of sixteen individuals transmitted parasites to Phlebotomus papatasi between 10 and 25 weeks post-infection. Parasites were mostly restricted to the inoculation site, with limited dissemination to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
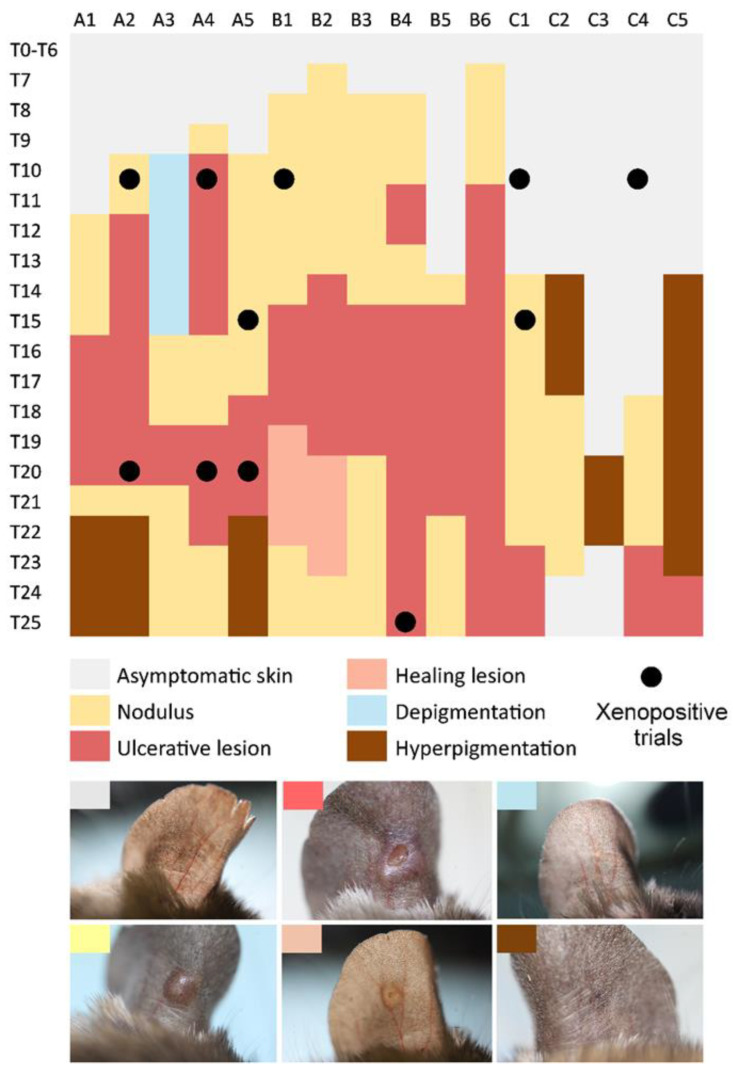 Figure 1
Figure 1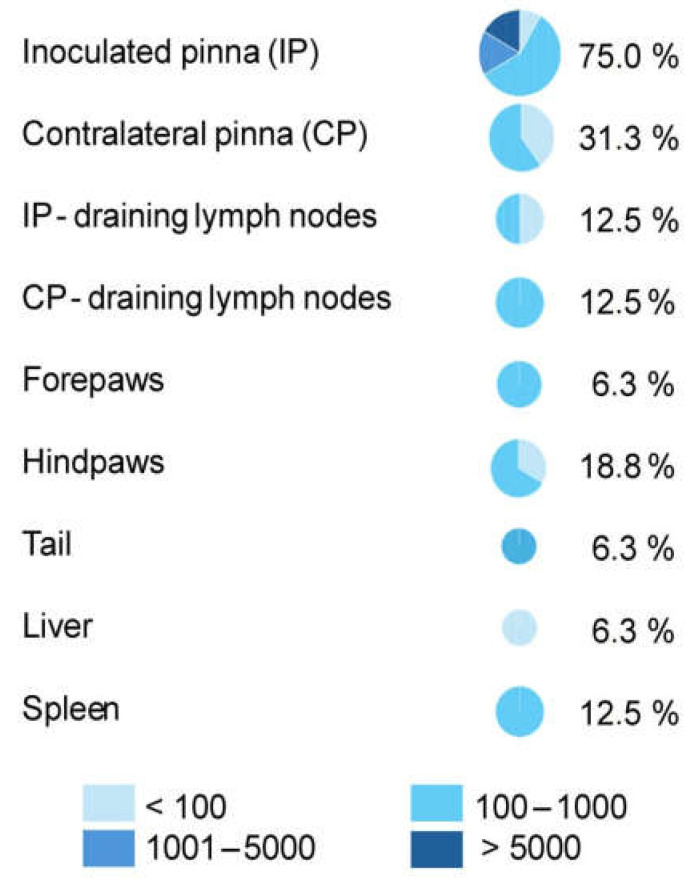 Figure 2
Figure 2- —Czech Science Foundation
- —European Union’s Horizon 2020
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsResearch on Leishmaniasis Studies · Parasites and Host Interactions · Trypanosoma species research and implications
1. Introduction
Cutaneous leishmaniasis (CL) is a zoonotic vector-borne disease caused by parasitic protozoans of the genus Leishmania Ross, 1903 (Kinetoplastida: Trypanosomatidae) and typically manifests as self-limiting cutaneous lesions. The parasites exhibit a digenetic life cycle, alternating between vertebrate hosts and blood-feeding female phlebotomine sand flies (Diptera: Psychodidae). CL is endemic in 89 countries worldwide, with the Eastern Mediterranean region and Algeria representing a major eco-epidemiological hotspot that accounts for approximately 79% of all newly reported cases [1]. Although CL has been present in Algeria for decades, its incidence has increased substantially in recent years, with recurrent local outbreaks and thousands of new human cases reported annually [2]. Leishmania major Yakimoff and Schokhor, 1914, is the dominant causative agent, whereas Leishmania tropica Wright, 1903, and Leishmania infantum Nicolle, 1908, occur less frequently and exhibit distinct eco-epidemiological patterns [3,4].
In the Maghreb region, L. major is primarily transmitted by Phlebotomus papatasi (Scopoli, 1786) and maintained in gerbil reservoir hosts, mainly Psammomys obesus Cretzschmar, 1828, and Meriones shawi (Duvernoy, 1842) [5]. Their reservoir competence is supported by both field and laboratory studies demonstrating high infection prevalence and prolonged infectiousness to sand fly vectors [6,7,8]. However, susceptibility to L. major is not restricted to gerbils, as shown by the involvement of Microtus guentheri (Danford & Alston, 1880) in Israel [9]. Notably, several endemic CL foci in Algeria lack any confirmed reservoir species, emphasizing the need to investigate alternative rodent hosts among the diverse rodent fauna of the country [10]. Recently, Gerbillus amoenus (de Winton, 1902) was proposed as a potential reservoir in Illizi, an area of sporadic CL outbreaks in the central Sahara [11]. The present study extends this work by evaluating the susceptibility of Acomys cahirinus seurati (Desmarest, 1819).
The genus Acomys I. Geoffroy, 1838, is widely distributed across Africa and the Middle East, comprising small, arid-adapted rodents commonly known as spiny mice. Taxonomic relationships within the genus remain unresolved [12]. Multilocus phylogenetic analyses identify the A. cahirinus group as one of five major lineages and the most genetically diverse, encompassing multiple clades awaiting formal taxonomic revision [13]. Although the Algerian form has been assigned species status by some authors (Acomys seurati Heim de Balsac, 1936) [10], its taxonomic status remains uncertain; therefore, the designation A. cahirinus seurati is adopted here, according to [12].
This study aimed to assess whether A. cahirinus seurati can sustain L. major infection and transmit the parasite to sand fly vectors, thereby fulfilling the criteria of a potential reservoir host in Algeria.
2. Materials and Methods
2.1. Rodents, Parasites and Sand Flies
A breeding colony of A. cahirinus seurati was established at the animal facility of the Pasteur Institute of Algiers using animals originating from Illizi, a province in the southeastern part of the central Sahara, Algeria (26°30′18″ N, 8°28′56″ E; 567 m Above See Level). A detailed description of the locality is provided elsewhere [11]. In 2022, experimental infections were performed on six animals in Algeria. Subsequently, four males and four females were transported to Czechia to establish a breeding colony at the animal facility of the Faculty of Science, Charles University, Prague.
Animals were housed in family groups in glass cages (40 × 60 × 55 cm) or T4 breeding containers (Velaz, Prague, Czech Republic) with bedding (Hapy Horse, SubliCZ.cz, Sojovice, Czech Republic), nesting material (Wood wool, Ratiboř, Czech Republic), and hay. Rodents were provided with a standard rodent diet (Krysík, SubliCZ.cz, Sojovice, Czech Republic), supplemented occasionally with fresh organic vegetables and Zophobas Lacordaire, 1859, larvae, and had access to water ad libitum. Environmental conditions were maintained at 22–25 °C, 40–60% relative humidity, and a 12 h light/12 h dark photoperiod.
A laboratory colony of P. papatasi originating from Turkey has been maintained at the Department of Parasitology, Charles University, Prague, under standard conditions (26 °C, 60–70% relative humidity, 14 h light/10 h dark photoperiod) with access to 50% sucrose, as previously described [14].
The human isolate L. major MHOM/DZ/09/LIPA100/MON-25 from the M’Sila region, Algeria, was used. Promastigotes were cultured in M199 medium (Sigma-Aldrich, Merck, Darmstadt, Germany) supplemented with 10% heat-inactivated fetal bovine serum (FBS, Sigma-Aldrich, Merck, Darmstadt, Germany), 1% BME vitamins, 2% sterile human urine, and 250 µg/mL amikacin (Medochemie Bohemia, Prague, Czech Republic).
2.2. Experimental Infections of Sand Flies
Phlebotomus papatasi females were experimentally infected with L. major as described previously [15]. Briefly, log-phase promastigotes were resuspended in heat-inactivated defibrinated ram blood at a concentration of 5 × 10^6^ parasites/mL. Sand flies were allowed to feed through a chick-skin membrane, and fully engorged females were maintained under standard insectary conditions. For rodent infections, sand fly females were dissected 8–10 days post-blood meal, when mature infections with accumulated metacyclic forms were present in the thoracic midguts.
2.3. Experimental Infections of Rodents
Experimental infections of rodents were first carried out in Algeria, as a preliminary experiment on a small number of individuals (6 females) infected with a higher dose (1 × 10^6^) of culture-derived promastigotes.
After the establishment of the colony in Prague, the offspring of the captured animals were tested with a natural infectious dose of promastigotes derived from the guts of experimentally infected sand flies in three independent repetitions: five males (A1–A5) in experiment A, three females and three males (B1–B6) in experiment B, and five males (C1–C5) in experiment C. Rodents aged 6–10 weeks were anesthetized using a ketamine (66 mg/kg) (Narkamon, Bioveta, Ivanovice na Hané, Czech Republic) and xylazine (26 mg/kg) (Xylapan, Vétoquinol, Nymburk, Czech Republic) mixture. Parasites isolated from thoracic midguts of infected sand flies were pooled in sterile saline and counted using a Bürker chamber. Salivary glands (SG) of P. papatasi females were pooled (10 glands per 10 µL saline), stored at −20 °C, and subjected to three freeze–thaw cycles in liquid nitrogen prior to use. The resulting SG lysate was mixed with the parasite suspension and intradermally injected into the left ear pinna in a total volume of 5.5 µL. Each animal received 2–5 × 10^4^ parasites and 0.5 SG.
The proportion of metacyclic promastigotes in the inoculum was determined from methanol-fixed, Giemsa-stained smears (Sigma-Aldrich, Merck, Darmstadt, Germany). Parasites were examined by light microscopy, photographed using an Olympus DP70 camera (Olympus Czech Group, Prague, Czech Republic), and measured with ImageJ software (https://imagej.net/software/imagej/) [15]. Promastigotes were classified as metacyclic when the flagellum length was at least twice the body length.
Animals were monitored weekly by visual inspection for clinical signs of infection and subjected to xenodiagnosis at 5-week intervals for up to 25 weeks post-infection (Prague experiments).
2.4. Xenodiagnosis
Rodents were anesthetized with ketamine/xylazine, and 30–40 female P. papatasi were allowed to feed on the inoculated pinnae using small plastic tubes covered with fine nylon mesh. On days 7–8 post-blood meal, sand flies were dissected, and their guts were examined by light microscopy. Infection intensity and localization were assessed as described previously [16].
2.5. Tissue Sampling and Parasite Detection
Rodents were euthanized by overdose with anesthetic. The following tissues were collected: whole inoculated and contralateral pinnae, ear-draining lymph nodes, spleen, liver (approximately one quarter), forepaws, hindpaws, and tail skin. Samples were stored at −20 °C until processing.
Total DNA was extracted using a tissue DNA isolation kit (Roche Diagnostics, Mannheim, Germany) according to the manufacturer’s instructions. Parasite loads were quantified by quantitative PCR (qPCR) using a LightCycler^®^ 480 system and SYBR Green chemistry (LightCycler^®^ 480 SYBR^®^, Green I Master, Mannheim, Germany). Primers targeting a 116 bp fragment of Leishmania kinetoplast minicircle DNA were used (forward primer (13A): 5′-GTGGGGGAGGGGCGTTCT-3′ and reverse primer (13B): 5′-ATTTTACACCAACCCCCAGTT-3′). PCR conditions consisted of an initial denaturation at 95 °C for 5 min, followed by 40 cycles of 95 °C for 10 s, 61 °C for 20 s, and 72 °C for 10 s. Standard curves were generated using 10-fold serial dilutions of L. major DNA corresponding to 5 × 10^5^ to 5 × 10^−1^ parasites per reaction. Quantification was performed by interpolation from the standard curve, and melting curve analysis (72–95 °C) was used to verify amplification specificity.
3. Results
3.1. Preliminary Experiment
Six animals were infected in Algeria with 1 × 10^6^ culture-derived L. major promastigotes and monitored for 15 weeks. Nodules, representing the first external signs of infection, appeared 6–7 weeks post-infection (p.i.) In two animals, nodules resolved within two weeks, and no further clinical signs developed. In contrast, four animals progressed to ulcerative lesions; lesion healing occurred in one individual by 14 weeks p.i., whereas lesions persisted in the remaining three animals until the end of the experiment. Post-mortem PCR analysis detected Leishmania DNA in all inoculated pinnae. Parasite DNA was additionally detected once in the contralateral pinna and in two cases each in the liver and spleen.
3.2. Course of Infection and Xenodiagnosis
After establishment of the colony in Prague, a total of 16 A. cahirinus seurati were infected in three independent repetitions (A, B, C) with (2–5) × 10^4^ sand fly–derived promastigotes plus 0.5 SG equivalents per animal. The proportion of metacyclic forms in the inocula was 33%, 18%, and 17% in experiments A, B, and C, respectively.
All animals developed cutaneous manifestations at the inoculation site during the 25-week observation period (Figure 1). In experiments A and B, the first visible signs appeared 7–10 weeks p.i., whereas in experiment C, the asymptomatic period lasted at least 14 weeks. The earliest clinical manifestation was typically a nodule, which progressed to an ulcerative lesion within 1–9 weeks. Lesions resolved within 2–9 weeks in most animals; however, in five individuals, lesions persisted until the end of the experiment (25 weeks p.i.). Given the overall self-limiting course of infection, lesion healing in these animals would likely have occurred with longer follow-up. Post-healing hyperpigmentation was observed in three animals, whereas in three others, hyperpigmentation preceded lesion development, and in one animal it represented the only skin manifestation. Transient depigmentation was recorded in a single individual for six weeks prior to nodule formation (Figure 1).
Xenodiagnosis was performed at five-week intervals, during which a total of 1598 female P. papatasi were allowed to feed on anesthetized rodents. Overall, 44% of animals (7/16) were infectious to sand flies, with four individuals transmitting parasites repeatedly. Infectiousness was most frequently detected at 10 weeks p.i., although positive xenodiagnostic results were also obtained at 15, 20, and 25 weeks p.i. (Figure 1; Table S1).
3.3. Post-Mortem Analysis
Post-mortem analyses revealed the presence of Leishmania DNA in all animals except individual A5, in which lesion healing was nearly complete by 21 weeks p.i., four weeks prior to the end of the experiment. Parasites were most frequently detected in the inoculated pinna (75% of animals; Figure 2). However, dissemination to other peripheral tissues was also observed, particularly to the contralateral pinna (31%) and hindpaws (19%). Visceral dissemination was rare, with parasite DNA detected in the spleen of 12% and in the liver of 6% of animals. Most tissues harbored low parasite loads (100–1000 parasites), whereas moderate (1000–5000) and high (>5000) parasite burdens were detected exclusively in the inoculated pinnae (Figure 2).
4. Discussion
This study provides novel experimental evidence supporting the inclusion of A. cahirinus among potential rodent reservoir hosts of L. major in North Africa. Identification of reservoir hosts is a critical prerequisite for the effective control of zoonotic diseases; however, it requires long-term, integrative field and laboratory investigations.
Recent parasitological surveys have revealed the presence of L. major DNA in a broad range of rodent species across North Africa. In addition to the well-established reservoir hosts P. obesus and M. shawi, natural infections have been documented in several other rodents, including Meriones libycus Lichtenstein, 1823, in Libya [17,18]; Ctenodactylus gundi (Rothman, 1776), Jaculus jaculus (Linnaeus, 1758) and Jaculus hirtipes (Lichtenstein, 1823) in Tunisia [19,20]; and Meriones crassus Sundevall, 1842, Meriones sacramenti Thomas, 1922, Rattus rattus (Linnaeus, 1758), Rattus norvegicus (Berkenhout, 1769), Gerbillus pyramidum I. Geoffroy, 1803, and Gerbillus andersoni de Winton, 1902, in Egypt [21,22,23]. Beyond rodents, L. major DNA has also been detected in insectivores and carnivores, including hedgehogs (Atelerix algirus Lereboullet, 1842, Paraechinus aethiopicus (Ehrenberg, 1832)) and the weasel Mustela nivalis Linnaeus, 1766 [24,25].
Several Leishmania species have previously been reported in rodents of the genus Acomys. Early evidence originates from Sudan, where inoculation of spleen material from Acomys albigena (Cretzschmar, 1828) resulted in infection of laboratory hamsters in a focus endemic for Leishmania donovani (Leishman & Donovan, 1903) [26]. More recently, L. tropica was detected molecularly in Acomys spp. in southern Ethiopia [27], and L. major DNA was identified in Acomys dimidiatus (Cretzschmar, 1826) in Iran [28]. However, detection of parasite DNA alone does not demonstrate reservoir competence.
To qualify as a reservoir host, a species must support long-term parasite persistence—ideally across at least one non-transmission season—and allow parasite localization compatible with sand fly infection [29]. Otherwise, infected animals may act only as parasite sinks, contributing little or nothing to onward transmission [30]. For this reason, laboratory-based experimental infections combined with xenodiagnosis are essential complements to field studies.
Our experimental data demonstrate that A. cahirinus seurati can remain infectious to P. papatasi for up to 25 weeks post-infection, with parasites predominantly persisting in the skin at the site of inoculation. Visceral dissemination to the spleen and liver was rare, which is consistent with observations in most rodent hosts of L. major [8,31,32]. Cutaneous lesions followed a self-limiting course and healed without extensive tissue destruction, an outcome consistent with the exceptional regenerative capacity of Acomys skin [33].
Importantly, the presence of ulcerative skin lesions is not a prerequisite for infectiousness to sand fly vectors. Previous studies have shown that asymptomatic M. shawi individuals were as infectious to P. papatasi as animals with active lesions [8]. Similarly, experimental infections of other African rodent species proposed as potential L. major reservoirs, such as Mastomys Thomas, 1915, and Arvicanthis Lesson, 1842, resulted only in mild skin alterations, including hyperpigmentation, without ulcer formation [32].
In the present study, Acomys individuals were infectious to sand flies from week 10 to week 25 post-infection; however, the overall proportion of infected P. papatasi females was low (0.8%). This finding aligns with previous observations indicating that even in hosts with substantial skin parasite loads, only a fraction of feeding sand flies acquire infection, likely due to the heterogeneous and patchy distribution of parasites in the skin [34,35]. In M. shawi, infection rates reached 11%, with a strong dependence on the precise feeding site: 37.9% of sand flies feeding at lesion margins became infected, compared with only 1.4% feeding on intact skin [35].
The present study was conducted under controlled experimental conditions using a single, well-characterized L. major strain and the primary vector species P. papatasi, which is considered the dominant and epidemiologically most relevant vector of L. major in North Africa. While this design ensured biological relevance and experimental reproducibility, future studies may examine whether reservoir competence of Acomys spp. varies among different parasite genotypes circulating across endemic regions. In addition, the study focused on a single population of A. cahirinus seurati originating from southeastern Algeria. Given the unresolved taxonomy and substantial genetic diversity within the genus Acomys, reservoir competence may differ among geographically or genetically distinct populations, which warrants further investigation.
5. Conclusions
In conclusion, this study provides the first experimental evidence that A. cahirinus seurati fulfills key criteria of a potential reservoir host for L. major, including sustained parasite persistence in the skin and the ability to infect P. papatasi over an extended period up to 6 months. Together with previous findings on G. amoenus, our results suggest that alternative rodent hosts may play an important role in maintaining L. major transmission in endemic foci where the classical reservoirs P. obesus and M. shawi are absent. This scenario is particularly relevant for sparsely populated Saharan regions such as Illizi in southeastern Algeria, where local ecological conditions favor distinct host assemblages. Recognition of such non-classical reservoir hosts is essential for improving risk assessment and designing targeted surveillance and control strategies for cutaneous leishmaniasis in North Africa.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ruiz-Postigo J.A. Jain S. Madjou S. Agua J.F.V. Maia-Elkhoury A.N. Valadas S. Warusavithana S. Osman M. Yajima A. Lin Z. Global leishmaniasis surveillance, 2022: Assessing trends over the past 10 years Wkly. Epidemiol. Rec.202398471488
- 2Relevé Épidémiologique Mensuel «R.E.M.» Algérie Available online: https://www.insp.dz/images/PDF/Epidemio/REM%20annuel%202024%20%20au%2003 AOUT.pdf(accessed on 3 February 2026)
- 3Aoun K. Bouratbine A. Cutaneous Leishmaniasis in North Africa: A review Parasite 2014211410.1051/parasite/201401424626301 PMC 3952656 · doi ↗ · pubmed ↗
- 4Benikhlef R. Aoun K. Boudrissa A. Ben Abid M. Cherif K. Aissi W. Benrekta S. Boubidi S.C. Späth G.F. Bouratbine A. Cutaneous Leishmaniasis in Algeria: Highlight on the Focus of M’Sila Microorganisms 2021996210.3390/microorganisms 905096233947003 PMC 8146893 · doi ↗ · pubmed ↗
- 5Ashford R.W. The leishmaniases as emerging and reemerging zoonoses Int. J. Parasitol.2000301269128110.1016/S 0020-7519(00)00136-311113254 · doi ↗ · pubmed ↗
- 6Ghawar W. Toumi A. Snoussi M.-A. Chlif S. Zâatour A. Boukthir A. Hamida N.B.H. Chemkhi J. Diouani M.F. Ben-Salah A. Leishmania Major Infection Among Psammomys Obesus and Meriones Shawi: Reservoirs of Zoonotic Cutaneous Leishmaniasis in Sidi Bouzid (Central Tunisia)Vector-Borne Zoonotic Dis.2011111561156810.1089/vbz.2011.071221919726 PMC 3263488 · doi ↗ · pubmed ↗
- 7Derbali M. Chelbi I. Ben Hadj Ahmed S. Zhioua E. Leishmania major Yakimoff et Schokhor, 1914 (Kinetoplastida: Trypanosomatidae) chez Meriones shawi Duvernoy, 1842 (Rodentia: Gerbillidae): Persistance de l’infection du mérion et de son infectivité pour le phlébotome vecteur Phlebotomus (Phlebotomus) papatasi Scopoli, 1786 (Diptera: Psychodidae)Bull. Soc. Pathol. Exot.201210539940210.1007/s 13149-012-0259-423055379 · doi ↗ · pubmed ↗
- 8Sadlova J. Vojtkova B. Lestinova T. Becvar T. Frynta D. Benallal K.E. Mekarnia N. Harrat Z. Volf P. Infectiousness of Asymptomatic Meriones shawi, Reservoir Host of Leishmania major Pathogens 20231261410.3390/pathogens 1204061437111500 PMC 10143307 · doi ↗ · pubmed ↗