First Detection of Human- and Dog-Associated Demodex Mites (Acari, Arachnida) in Southern European Wolves (Canis lupus)
Natalia Sastre, Manena Fayos, Luca Rossi, Olga Francino, Roser Velarde, Sebastian E. Ramos-Onsins, Lluís Ferrer

TL;DR
This study found human- and dog-associated Demodex mites in southern European wolves, suggesting recent cross-species transmission rather than ancient host specificity.
Contribution
The first detection of D. folliculorum and D. brevis in wild canids, challenging the assumption of strict host specificity in Demodex mites.
Findings
37.1% of wolves tested positive for Demodex DNA, with higher prevalence in Italian wolves.
Four Demodex species were identified, including those typically associated with humans and dogs.
Co-infestations with multiple Demodex species were observed for the first time in wild canids.
Abstract
Demodex mites are common commensals of mammalian skin, but under certain conditions, they can cause severe skin diseases. This study analyzed the presence, diversity, and phylogenetic relationships of Demodex species in two wolf subspecies from southern Europe to determine whether species-level differences exist between wild and domestic canids after thousands of years of divergence. A total of 1400 hair samples from 140 wolves were analyzed using a real-time PCR (qPCR) targeting mitochondrial 16S rRNA and nuclear 18S rRNA genes. Overall, 37.1% (52/140; 95% CI: 29.0–45.9%) of wolves were positive for Demodex DNA, with a higher prevalence in Italian (46%) than in Iberian (36%) wolves. The lip and chin areas were the most reliable sampling sites. Four Demodex species were identified in wolves: D. injai and D. canis (associated with dogs), and D. folliculorum and D. brevis (associated with…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
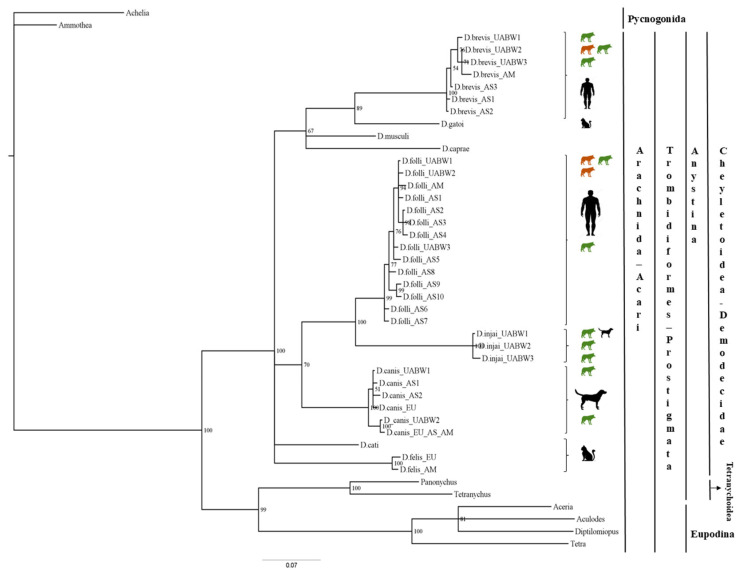 Figure 1
Figure 1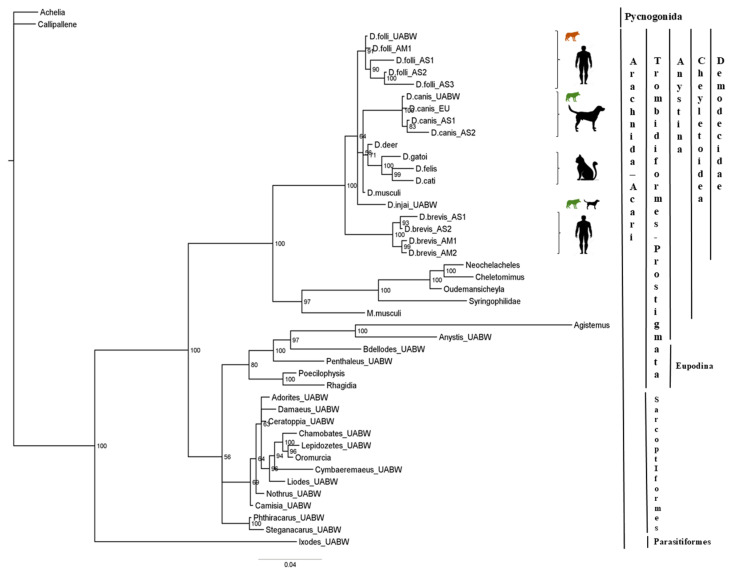 Figure 2
Figure 2- —European Interreg Program V-A Spain-France-Andorra (POCTEFA 2014-2020)
- —CERCA Programme, Generalitat de Catalunya
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDermatological diseases and infestations · Acne and Rosacea Treatments and Effects · Nail Diseases and Treatments
1. Introduction
Demodex mites are considered normal inhabitants in mammalian skin [1,2,3]. Currently, more than 80 species of Demodex spp. have been described, genetically and/or morphologically, in more than 80 wild and domestic mammals, including humans, dogs, cats, deer, ferrets, otters, and mice [2,4,5,6,7,8,9,10,11,12,13]. Most mammals harbor Demodex mites on the skin without developing lesions or any other clinical signs [1,13]. Multiple pieces of evidence and studies indicate that the immune system controls Demodex populations on the skin [1,3,14]. Although Demodex mites are considered commensals in small numbers, when they proliferate, they may cause severe dermatitis, for instance, rosacea in humans or demodicosis in dogs and cats [15,16,17]. In almost all cases, proliferation results from a compromised immune system, as occurs in transgenic mice [14,18] or in dogs or humans receiving immunosuppressive medical treatment [3,19,20].
Genetic sequencing has proven highly effective for detecting and identifying Demodex mites in both healthy and diseased mammals [4,7,9,13]. Because of the sensitivity and specificity of genetic techniques, mites can be detected where non-molecular methods, such as microscopy or trichoscopy, fail to yield results [21,22]. Demodex mites have been detected, identified, and classified in humans, dogs, cats, mice, and ferrets using different genetic approaches such as the mitochondrial 16S rRNA gene region [4,6,7,9], and the nuclear 18S rRNA gene region [2,9,23,24]. Ravera et al. (2013) [13], using a fragment of the chitin synthase gene, concluded that Demodex DNA can be detected in the skin of any dog if 20 or more skin sites are sampled. In wolves, Demodex mites have been detected in fecal samples [25,26,27]. Since Demodex spp. are hair follicle mites, their presence in feces could be explained by accidental ingestion during grooming or from the prey itself. Nevertheless, it remains unknown which Demodex species are present in wolves’ skin.
Research on Demodex mites is relevant not only to medicine but also to evolutionary biology. Their parasitic association with mammals may date back approximately 220 million years, coinciding with the emergence of hair follicles [8,28]. Roughly 100 million years ago, the clade Boreoeutheria diverged into two sister lineages: Euarchontoglires, which includes primates, and Laurasiatheria, which includes canids [29]. Demodex mites have been documented, both morphologically and genetically, in species from both groups, suggesting either a long-term stability of the hair follicle niche or a high capacity for mite transmission across and within placental mammals [4,5,7,9,24,30,31]. The genus Demodex therefore represents an exceptional case of parasitic parallelism [2,8,13,32].
The domestic dog originated from the gray wolf (Canis lupus) and is associated with Neolithic humans on most continents [33,34,35]. Axelsson et al. (2012) [36] suggest that dogs descended from scavenger wolves adapted to a starch-rich diet during the dawn of the agricultural revolution. However, it remains unclear where domestication occurred and whether it occurred once or multiple times worldwide. Archaeological records suggest different origins and domestication times: fossilized early dog remains have been found in Belgium (36,000 years ago; [37]), eastern Siberia (33,500 years ago; [38]), and northern Israel (12,000 years ago; [39]). Nevertheless, specimens older than 14,000 years do not appear related to modern dogs [38]. Genetic studies also yield different results. Autosomal single nucleotide polymorphism (SNP) data indicate contributions from Middle Eastern and European wolves [40], while shotgun sequencing data suggest an origin in Eurasia, possibly southern Asia [41]. Wang et al. (2015) [42] propose that domestication began around 33,000 years ago in East Asia, followed by migrations toward the Middle East and Europe. More recently, Bergström et al. (2022) [43] found that modern dogs are genetically closer to ancient wolves from eastern Eurasia than to those from western Eurasia. However, Freedman et al. (2014) [44] found that no extant wolf lineage is more closely related to dogs, implying that an extinct wolf lineage was their ancestor. In any case, dog domestication can be traced back at least 15,000 years.
The gray wolf (Canis lupus) is listed on the IUCN Red List (2018; https://www.iucnredlist.org/species/3746/247624660, accessed on 9 February 2026) as “Least Concern” and is included in Appendix II of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) (except for the populations of Bhutan, India, Nepal, and Pakistan, which are listed in Appendix I). Currently, five subspecies of Canis lupus are recognized in North America and seven in Eurasia. This classification is primarily based on genetic and morphological differences among populations, reflecting adaptations to distinct climates, latitudes, or habitats [45,46,47,48]. In Western Europe, the differentiation between the subspecies C. l. signatus of the Iberian Peninsula and C. l. italicus of the Italian Peninsula is the result of prolonged geographic isolation, which continues to this day [49,50,51,52].
Our study aims to determine whether there are species-level differences in Demodex mites between wild and domestic canids after this extensive period of divergence. For this purpose, we analyzed 1400 hair samples from 140 wolves belonging to the two southern European subspecies, using two gene fragments, 16S rDNA and 18S rDNA, widely used for the molecular identification of Demodex mites. We specifically aim to identify the Demodex species present in wolves, compare them with those found in domestic dogs, and assess whether host domestication and divergence may have influenced mite diversity. By addressing these questions, we test the hypothesis that long-term host divergence and domestication may have resulted in distinct Demodex species in wild and domestic canids, providing new insights into the evolutionary history of these mites.
2. Materials and Methods
2.1. Ethics Statement
In Cantabria, wolves were classified as a hunting species under Annex I of the Cantabrian Law, 12/2006. From 2017 to 2020, wolves were removed under a species-control program implemented by “Subdirección General del Medio Natural” of the Government of Cantabria, in accordance with Laws 2/2017, 5/2018, and MED/5/2019, under expedient numbers CVE-2017-1827, CVE-2018-17138, and CVE-2019-2950, to reduce wolf attacks on livestock in conflict areas. From 2021 to 2025, wolves in the Iberian Peninsula were protected, and hunting was banned. Individuals necropsied from 2021 onwards died accidentally. Natural Environment agents transported the carcasses to the Wildlife Rescue Center of Cantabria for necropsies. The three wolves from Aragón, Castilla y León and Catalunya died after being hit by a car. Hair samples were taken by agents of the Natural Environment for necropsies. Individuals from Italy (where wolf hunting is banned) died accidentally. Technicians from the European project Life12 NAT/IT/000807 WolfAlps moved carcasses to the Department of Veterinary Science (Università degli Studi di Torino) for necropsies. All procedures involving animal carcasses complied with national and regional regulations and did not involve the deliberate killing of animals for research purposes.
2.2. Sampling
Hair samples from Iberian and Italian wolves were obtained from northern Spain, including Cantabria (126 samples), Aragón (1), Castilla y León (1), and Catalunya (1), and from the Piemonte region of northern Italy (11) (Table 1). One hundred twenty-seven wolves (61 females and 66 males) from Cantabria and Castilla y León were morphologically and genetically identified as Canis lupus signatus, and 13 wolves (6 females and 7 males) from Piemonte, Catalunya, and Aragón were identified as Canis lupus italicus (Table 1). Between 2017 and 2025, we collected a total of 1400 hair samples from 140 wolf carcasses, with 10 samples per carcass obtained during necropsy examinations (Table 1 and Table S1). Five sites were from the face: periocular, lips, nose, chin, and entrance to the external ear canal, and five sites were from the rest of the body: dorsum, lumbar, abdomen, forelimb, and hindlimb. Using gloved hands and surgical mosquito forceps, hair was plucked in the direction of growth to include the hair bulb (root) for DNA extraction. Hair samples were stored at −20 °C until DNA extraction.
2.3. DNA Extraction and Real-Time PCR (qPCR) Amplification
Over the years, DNA from hair bulbs has been extracted using two different methods (Table S1): Tris-HCl approach, following Ravera et al. 2013 [13]; and PerkinElmer automated extraction, Chemagic 360 Instrument (Baesweiler, Germany), following the manufacturer’s protocol, including an incubation step at 56 °C overnight and a final elution volume of 50 µL DNA. An extraction blank was included in each 96-well plate. All extracted DNA was diluted 1:10 for qPCR amplification.
16S and 18S primers were used to amplify approximately 300-bp and 500-bp fragments of the mitochondrial 16S rRNA gene and the nuclear 18S rRNA gene, respectively [9]. All DNA samples were amplified by real-time qPCR and prepared under a laminar flow hood. Positive qPCR controls were obtained from known Demodex mite DNA [9]. Duplicates were amplified for each sample, and two extraction blanks and one negative qPCR control per gene were included in each 384-well plate to detect any qPCR exogenous DNA contamination. qPCR amplifications were performed in a QuantStudioTM 12K Flex Real-Time PCR System (Thermo Fisher Scientific, Waltham, MA, USA). Amplicons were sequenced and purified when melting curves showed Tm = 75 ± 1 °C for the 16S rRNA gene and Tm = 83 ± 2 °C for the 18S rRNA gene, and amplification cycles had Cp < 37. Sequences were separated on an ABI PRISM 3730 automated sequencer (Thermo Fisher Scientific) according to the manufacturer’s instructions.
2.4. Genetic Variability and Phylogenetic Analysis
Genetic diversity in Demodex mites was analyzed using DNASP 5.10 [53]. For comparisons and phylogenetic analysis, we used mite sequences from wolves (those ending in UAB) and those available in GenBank. Sequences were analyzed using SEQSCAPE 2.1.1 software (Thermo Fisher Scientific) and were compared with the GenBank database (www.ncbi.nlm.nih.gov/BLAST, accessed on 1 May 2025). Phylogenetic analysis for the mitochondrial 16S rRNA gene was carried out using 44 sequences and 302 bp (gaps included). Phylogenetic analysis for the 18S rRNA gene was carried out using 45 sequences and 491 bp (gaps included). The trees were rooted using outgroups from the class Pycnogonida, Achelia hispida (FJ862845) and Ammothea sp. (FJ862841) for the 16S rRNA gene, and Achelia echinata (AF005438) and Callipallene sp. (AF005439) for the 18S rRNA gene. MODELTEST 3.7 [54] was applied to select the best evolutionary model among 56 models of evolution by the Akaike information criterion. The Bayesian program MrBayes v3.2.7 [55] was implemented to generate the phylogenetic tree with 1,000,000 Markov Chain Monte Carlo iterations and a burn-in rate of 25%. Bootstrap values indicate the repeatability of the inferred clades and may provide a conservative estimate of their accuracy [56]. To display the phylogenies, we used the program FigTree (http://tree.bio.ed.ac.uk/software/figtree/, accessed on 1 September 2025).
3. Results
3.1. Mite Prevalence in Wolves
We screened 1400 hair samples from 140 wild wolves for mite DNA targeting the 16S rRNA and 18S rRNA genes. qPCR samples were considered positive when the melting curves (Tm) were close to Demodex control values (Tm = 75 °C ± 0.5 °C, 16S rDNA; Tm = 81.5 °C ± 0.5 °C, 18S rDNA), and at least one gene was successfully sequenced. The prevalence of Demodex mites in the hairs of 140 wolves was 37% (52/140; 95% CI: 29.0–45.9%). Of the Italian wolves, 46% (6/13; 95% CI: 19.2–73.9%) were positive for Demodex, while 36% were positive among Iberian wolves (46/127; 95% CI: 27.8–45.4%) (Table 1 and Table S1). Given the small sample size of Italian wolves, this prevalence should be interpreted with caution. No bias toward females and males (χ^2^ (1, N = 140) = 1.02, p > 0.05) was observed among Demodex-positive wolves.
Sampling 10 sites, 101 out of 1400 hair samples tested positive for Demodex by qPCR (Table S1). Twenty-three wolves (44%) were positive at one site, nineteen wolves (36%) at two sites, six wolves (12%) at three sites, two wolves (4%) at four sites, and two wolves (2%) at five and nine sites, respectively. Demodex positivity was significantly higher in face samples (71%) compared to body samples (χ^2^ (1, N = 1400) = 19.73, p < 0.001). The most likely site to detect Demodex mites was the lip area (20%), followed by the chin area (18%). The least common sites were the lumbar (4%) and the dorsal (3%) areas.
Regarding DNA extraction methods, the PerkinElmer automated extraction was significantly more efficient than Tris-HCl approach (χ^2^ (1, N = 1400) = 22.56, p < 0.0001), with 85/869 positives (10%) versus 16/531 (3%). Although the distribution of extraction methods differed between Italian (53% Perkin; N = 130) and Iberian (63% Perkin; N = 1270) wolves (χ^2^ = 4.87, p = 0.027), this did not affect the comparison of positive detection rates. A Cochran–Mantel–Haenszel test adjusting for both species confirmed a significant association between extraction method and positivity (common OR = 3.48, 95% CI: 2.02–6.00; χ^2^(1) = 22.7, p < 0.0001). However, the Breslow–Day test for homogeneity of odds ratios was not significant (χ^2^(1) = 0.94, p = 0.33), indicating that the Perkin method’s superior efficacy was consistent across wolf subspecies. A significant disadvantage of the Tris-HCL method is that DNA is not purified, reducing qPCR success due to higher inhibitor levels, which may lead to false-negative results. Its main advantages are its speed and low cost.
3.2. Identification of Demodex spp.
Using the 18S rRNA gene, 83 samples were positive for Demodex spp., 21 samples could not be identified due to different mite types in the same sample (double sequence), and 88 samples were positive for other acarid mites (Table S1). Among the positive Demodex samples, we identified Demodex injai (52), Demodex canis (26), and Demodex folliculorum (5), but not Demodex brevis (Table 2 and Table S1). Using the 16S rRNA gene, we identified 101 positive samples, including D. injai (54), D. canis (30), D. folliculorum (12), and D. brevis (5) (Table 2 and Table S1).
Interestingly, the Demodex species commonly reported in dogs, such as D. canis and D. injai, were not detected in any of the Italian wolves examined. Instead, two species commonly associated with humans, D. folliculorum and D. brevis, were identified. Differential sampling effort (0 positives in 13 Italian wolves versus 45 positives in 127 Iberian wolves) could potentially bias detection probabilities, even though it was statistically significant (Fisher’s exact test, p = 0.015). A larger sample size would be necessary to determine whether D. canis and D. injai are absent from this wolf subspecies, especially considering that a recent study found evidence of extensive wolf–dog hybridization in peninsular Italy [57]. In contrast, all four Demodex species, those usually reported in both dogs and humans, were identified in Iberian wolves. D. injai showed the highest prevalence, present in more than half of the individuals (64%), followed by D. canis (24%), while D. brevis and D. folliculorum had lower rates (8% and 4%, respectively). Additionally, co-infestation with D. canis and D. injai and D. canis and D. brevis were observed in three and one Iberian wolves, respectively, and D. brevis and D. folliculorum in one Italian wolf (Table 2 and Table S1).
3.3. 16S rDNA Sequence Variability and Phylogenetic Relationships
We identified 11 Demodex sequences (haplotypes) among 101 sequences of the mitochondrial 16S rRNA gene from 52 wolves (Table 2 and Table S1). The 11 Demodex haplotypes were submitted to GenBank because they had not been previously described in wolves and, in most cases, represented new records (Table 2).
They were identified as follows: (1) Demodex injai (three haplotypes): D.injai_UABW1 (N = 19 sequences, identical to the sequence KT259449 from GenBank), D.injai_UABW2 (N = 29 sequences), and D.injai_UABW3 (N = 6 sequences); (2) Demodex canis (two haplotypes): D.canis_UABW1 (variant cornei) (N = 27 sequences, identical to sequences JX390979 and MN161404 from GenBank), and D.canis_UABW2 (N = 3 sequences, identical to several sequences from GenBank such as JF784000); (3) Demodex folliculorum (three haplotypes): D.folliculorum_UABW1 (N = 7 sequences, identical to sequences HQ844221, FN424245, and FN424246 from GenBank), D.folliculorum_UABW2 (N = 3 sequences), and D.folliculorum_UABW3 (N = 2 sequences); and (4) Demodex brevis (three haplotypes): D.brevis_UABW1 (N = 2 sequences), D.brevis_UABW2 (N = 2 sequences), and D.brevis_UABW3 (N = 1 sequence). Haplotypes from D. injai and D. canis were exclusive to the Iberian wolves, as well as haplotypes D.folliculorum_UABW3, D.brevis_UABW1, and D.brevis_UABW3 (Table 2). The exclusive haplotype found in Italian wolves was D.folliculorum_UABW2. Finally, haplotypes D.folliculorum_UABW1 and D.brevis_UABW2 were shared between wolf populations.
We conducted a phylogenetic analysis that included the 11 Demodex sequences, 31 sequences from the order Trombidiformes, and two outgroups obtained from GenBank (Figure 1). In total, we aligned 44 fragments of 302 bp each (gaps included). The Tamura-Nei substitution model, which assumes equal base frequencies and accounts for among-site rate heterogeneity using a gamma distribution (TrNef+G) was identified as the best-fit model for the 16S rRNA gene.
The 16S-Bayesian tree (Figure 1) showed two major splits among the Trombidiformes order (bootstrap value 100%). All our samples belong to the Anystina cohort, which includes the family Demodecidae. However, the Demodecidae sequences did not cluster by host species. For instance, D. brevis (host: human) was sister taxa with D. gatoi (host: cat), and D. folliculorum clade (host: human) was sister to the D. injai clade (host: dog) with strong bootstrap support (100%). Despite the short length of the 16S fragment, which limits deeper evolutionary inference, these results suggest that the evolutionary relationships among Demodex mites may not strictly follow host phylogeny, indicating possible host-switching events over evolutionary time.
3.4. 18S rDNA Sequence Variability and Phylogenetic Relationship
We identified three Demodex sequences (haplotypes) among 83 sequences of the nuclear 18S rRNA gene from 40 wolves (Table 2 and Table S1). The three Demodex haplotypes were submitted to GenBank because they had not been previously described in wolves (Table 2). They were identified as Demodex injai, Demodex canis and Demodex folliculorum, and they have been described previously in dogs and humans (Table S1).
In addition to Demodex-positive samples, we sequenced 49 samples that amplified at Cp < 37 and melting curves Tm = 82 °C + 3 °C (Table S1). We amplified DNA from 11 mites belonging to the order Oribatida, 3 to the order Prostigmata, and one to the order Ixodida, the only hematophagous mite detected in an Iberian wolf (Table S1). The remaining mites were most likely soil-dwelling and not host-associated. Table S1 shows the sequences of the 15 mites, along with their percentage identity relative to the reference sequence in GenBank. The genus names used in the phylogenetic tree should be considered tentative, as the order-level classification is reliable, but species-level identification is not possible with the available data. These findings indicate that the 18S rDNA fragment is not an optimal marker for detecting Demodex in wolves.
The 18S-Bayesian tree (Figure 2) showed a clear split (bootstrap value 100%) between the Parasitiformes (Ixodida) and the Sarcoptiformes (Oribatida) and the Trombidiformes (Prostigmata) Orders. However, Oribatida did not form a distinct cluster, whereas Prostigmata split into several clusters, including the Eupodina and Anystina cohorts. The Demodecidae family constitutes a monophyletic group of the Anystina cohort that includes four large clusters: (1) the D. folliculorum cluster (host: human), (2) the D. canis cluster (host: dog), (3) the D. gatoi, D. cati, and an unnamed Demodex species in a cat (here referred to as D. felis) cluster (host: cat), and (4) the D. brevis cluster (host: human). Again, D. injai appears to be more closely related to D. folliculorum than to D. canis; however, this result should be interpreted with caution due to the low bootstrap support (<70%).
Phylogenetic analyses of Demodex spp. The tree was estimated using MrBayes based on aligned fragments of the 16S rRNA gene. Branch support is based on 10,000 bootstrap replications. Bootstrap values are shown as percentages. The scale at the bottom represents genetic distances, expressed as the number of nucleotide substitutions per site. Green: Canis l. signatus; Brown: Canis l. italicus.
Phylogenetic analyses of Demodex spp. The tree was estimated using MrBayes based on aligned fragments of the 18S rRNA gene. Branch support is based on 10,000 bootstrap replications. Bootstrap values are shown as percentages. The scale at the bottom represents genetic distances, expressed as the number of nucleotide substitutions per site. Green: Canis l. signatus; Brown: Canis l. italicus.
4. Discussion
4.1. Detection, Prevalence, and Identification of Demodex spp.
Demodex mites were detected in over one-third of the wolves, with a slightly higher prevalence in Italian than in Iberian wolves and no sex bias, suggesting widespread infestation in both populations. Detection of Demodex DNA, however, does not necessarily reflect an active infestation or viable mites. Furthermore, it is important to emphasize that the wolves in this study were wild. In a previous study, in which we examined five body regions from 14 Mexican (Canis lupus baileyi) and 8 Alaskan (Canis lupus occidentalis) captive wolves, we were unable to detect any positive cases with confidence [9]. The fact that these captive animals were vaccinated annually, fed regularly, and kept in sheltered conditions possibly reduced mite proliferation and, consequently, the likelihood of detection, despite our assumption that all wolves harbor Demodex mites in their pilosebaceous complexes. Notably, to detect 37% of wolves positive for Demodex, each wolf had to be sampled at 10 different skin sites, yielding a total of 1400 samples, of which only 7% tested positive. Therefore, it is more effective to examine fewer wolves across multiple sites than to explore a larger number of wolves at a single site [1,9,13]. Based on our results, the number of sampling sites for detecting Demodex mites can be limited to the face, with the most suitable sites being the lips and chin. The dorsal and lumbar regions can be excluded. Reducing the number of sites decreases laboratory workload and overall costs. The resulting time and resources are recommended for use in a DNA extraction method incorporating a purification step to minimize the risk of false negatives, despite its higher cost relative to non-purified DNA extraction.
Regarding the genetic markers, our results suggest that the 16S rRNA gene is more suitable for Demodex detection and phylogenetic inference than the 18S rRNA gene. Amplification of the 18S marker may lead to the unintended detection of non-target mites, such as soil mites, or, in some cases, to the failure to detect Demodex mites. The mitochondrial 16S marker, instead, showed greater sequence variability and, consequently, higher resolution at short evolutionary distances than the nuclear 18S marker. However, despite this increased variability, bootstrap support for the nodes separating the major Demodex lineages remained low, generally failing to reach the 0.9–0.95 threshold, except for the folliculorum–injai clade. A similar lack of strong support was observed in the 18S-based phylogeny. These results indicate that some uncertainty persists in the inferred relationships among Demodex species, highlighting the need for additional loci and increased sampling to robustly resolve species boundaries and evolutionary relationships within the genus.
4.2. Co-Infestations and Host Specificity Concerns
To date, no peer-reviewed studies have explicitly documented co-infestations of D. canis and D. injai, and D. canis and D. brevis, within canids. Most reports describe these species in isolation, as dogs exhibit clinical signs of demodicosis [1,17,58]. Similarly, studies conducted in healthy dogs have not reported co-infestations [13,21,22]. In our study, no known diseases were present in either population at the time, which supports previous findings that Demodex mites can be carried asymptomatically across various mammalian hosts [9,13,59]. The absence of confirmed co-occurrence in dogs may indicate underdiagnosis due to limitations in sampling strategies, as co-infestations of D. canis with D. folliculorum and D. brevis have been documented in two healthy cats [7]. Co-infestation of D. brevis and D. folliculorum has also been documented in humans, particularly in relation to dermatological and ophthalmological conditions, such as chronic blepharitis and rosacea [60,61,62]. These two species usually inhabit different ecological niches: D. folliculorum in hair follicles and D. brevis in sebaceous or meibomian glands, but they can coexist in the same individual [62], and their combined presence has been suggested as a potential factor in the development of these diseases [63,64].
The observation of interspecific co-infestations in some wolves, along with findings that the Iberian wolf harbors Demodex species found in dogs and humans, suggests substantial ecological plasticity and raises important questions about the strict host specificity paradigm. Previous studies have raised similar concerns by reporting the presence of D. canis not only in dogs and wolves, but also in a variety of hosts, including bats, ferrets, cats, and mice, and, exceptionally, in a single bear and a single human [7,9,11,31,65]. These findings support a revision of the host–parasite model for Demodex, proposing that at least some species, especially those infecting carnivores, may function as opportunistic generalists rather than obligate specialists. Molecular studies across a broader range of wild and domestic hosts are needed to further assess the extent of host specificity versus host plasticity in this group.
4.3. Inference on the Process of Dog Domestication
Contrary to our initial hypothesis, our results indicate that wolves and domestic dogs share closely related Demodex species, suggesting that long-term host divergence and domestication may not have led to the evolution of distinct mite species in these canids. The identification of D. canis, D. injai, D. folliculorum, and D. brevis in wild wolves is a remarkable finding that challenges the traditional view of strict host specificity in Demodex mites. While D. canis and D. injai are typically associated with domestic dogs, and D. folliculorum and D. brevis with humans, their concurrent presence in wolves raises several questions regarding their evolutionary history and host associations. The detection of D. canis and D. injai in Iberian wolves is not entirely surprising, whereas their absence from the skin of Italian wolves is unexpected. Further analysis of additional individuals would be necessary to confirm the presence of these mites in the Italian population. And even more surprising is the detection of human-associated Demodex in wolves. Two possible explanations can be proposed for this unexpected detection: 1, wolves served as a reservoir and potential source of transmission to humans, occurring through early interactions between canids and hominids during the process of dog domestication [2], and 2, a more plausible scenario involving ecological contact and host-switching events, given the recent isolation history of these two wolf populations. The Iberian and Italian wolf populations have been geographically isolated since the early 20th century, when wolves became extinct in France [52]. Since then, the population sizes of Spanish and Italian wolves have also declined dramatically, with minima reached in both countries during the 70s of the 20th century [51,52,66,67]. The presence of human-associated Demodex species in wolves may result from cross-species transmission, potentially facilitated by overlapping habitats, direct contact (e.g., in captivity or rehabilitation centers), or indirect contact through domestic dogs. However, to date, no reports have been published of dogs harboring human-associated Demodex on their skin. Future studies employing longer or more variable genetic markers alongside broader host and geographic sampling will be crucial to disentangle the evolutionary relationships of Demodex mites, clarify potential host-switching events, and enhance our understanding of their ecology and evolution in both wild and domestic hosts.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ferrer L. Ravera I. Silbermayr K. Immunology and Pathogenesis of Canine Demodicosis Vet. Dermatol.201425427-e 6510.1111/vde.1213624910252 · doi ↗ · pubmed ↗
- 2Thoemmes M.S. Fergus D.J. Urban J. Trautwein M. Dunn R.R. Ubiquity and Diversity of Human-Associated Demodex Mites P Lo S ONE 20149 e 10626510.1371/journal.pone.010626525162399 PMC 4146604 · doi ↗ · pubmed ↗
- 3El-Moamly A. El-Swify O. Raising Awareness of Demodex Mites: A Neglected Cause of Skin Disease Infection 2025531273129810.1007/s 15010-025-02521-z 40319165 PMC 12316822 · doi ↗ · pubmed ↗
- 4Zhao Y.-E. Wu L.-P. Phylogenetic Relationships in Demodex Mites (Acari: Demodicidae) Based on Mitochondrial 16S r DNA Partial Sequences Parasitol. Res.20121111113112110.1007/s 00436-012-2941-722581346 · doi ↗ · pubmed ↗
- 5Izdebska J.N. Rolbiecki L. Demodex lutrae n. sp. (Acari) in European Otter Lutra lutra (Carnivora: Mustelidae) with Data from Other Demodecid Mites in Carnivores J. Parasitol.201410078478910.1645/14-532.124945076 · doi ↗ · pubmed ↗
- 6Silbermayr K. Horvath-Ungerboeck C. Eigner B. Joachim A. Ferrer L. Phylogenetic Relationships and New Genetic Tools for the Detection and Discrimination of the Three Feline Demodex Mites Parasitol. Res.201411474775210.1007/s 00436-014-4243-825468382 · doi ↗ · pubmed ↗
- 7Ferreira D. Sastre N. Ravera I. Altet L. Francino O. BardagíM. Ferrer L. Identification of a Third Feline Demodex Species through Partial Sequencing of the 16S r DNA and Frequency of Demodex Species in 74 Cats Using a PCR Assay Vet. Dermatol.201526239-e 5310.1111/vde.1220626178604 · doi ↗ · pubmed ↗
- 8Palopoli M.F. Fergus D.J. Minot S. Pei D.T. Simison W.B. Fernandez-Silva I. Thoemmes M.S. Dunn R.R. Trautwein M. Global Divergence of the Human Follicle Mite Demodex folliculorum: Persistent Associations between Host Ancestry and Mite Lineages Proc. Natl. Acad. Sci. USA 2015112159581596310.1073/pnas.151260911226668374 PMC 4703014 · doi ↗ · pubmed ↗