Exploring the Antibacterial Properties of a Newly Isolated Microviridae Phage Against Multidrug-Resistant Escherichia coli
Yoana Kizheva, Maria Pandova, Zoltan Urshev, Yoana Gladicheva, Tsvetelina Paunova-Krasteva, Sergei Ivanov, Petya Hristova

TL;DR
This paper explores a newly discovered phage that can kill drug-resistant E. coli, offering a potential alternative to antibiotics.
Contribution
The study characterizes a novel Microviridae phage (vB_SEC_3) with antibacterial potential against multidrug-resistant E. coli.
Findings
The phage vB_SEC_3 effectively lysed two MDR E. coli strains with high efficiency.
The phage is a putative new species within the genus Alphatrevirus of the Microviridae family.
The phage tolerated a wide pH and temperature range and showed minimal titer loss during storage.
Abstract
In response to the alarming rise in antimicrobial resistance, bacteriophages have re-emerged as a promising alternative to conventional antibiotic therapy. The main objective of this paper was to characterize a newly isolated phage (vB_SEC_3) in the context of its suitability for phage therapy against MDR E. coli, which is considered a priority pathogen. The phage was characterized at the morphological, genomic, and biological levels relevant to phage therapy. TEM analyses revealed a non-enveloped icosahedral capsid lacking tail structure. Phylogenetic and tANI analyses placed the phage within the α3 phages (genus Alphatrevirus) of the less-studied family Microviridae and revealed <95% similarity to its closest relatives, suggesting vB_SEC_3 is a putative novel species within this genus. The genome (6085 bp, GC content 45.3%) displayed the conserved organization typical for these…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
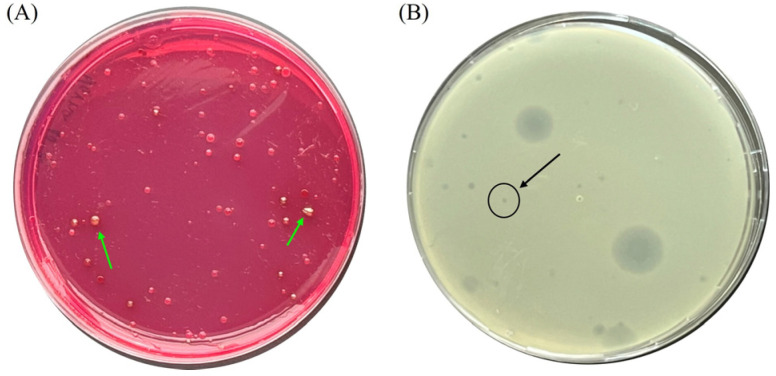 Figure 1
Figure 1 Figure 2
Figure 2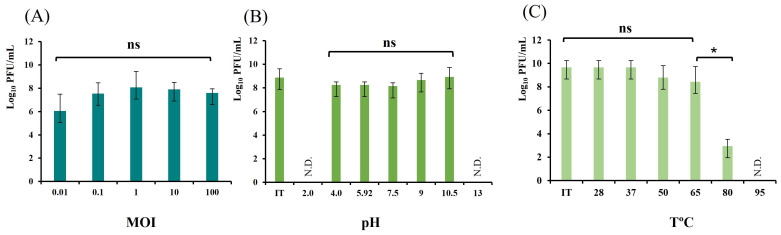 Figure 3
Figure 3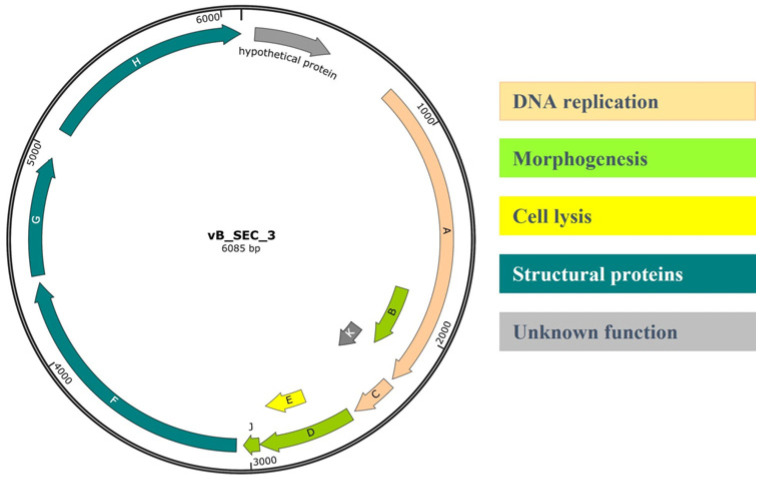 Figure 4
Figure 4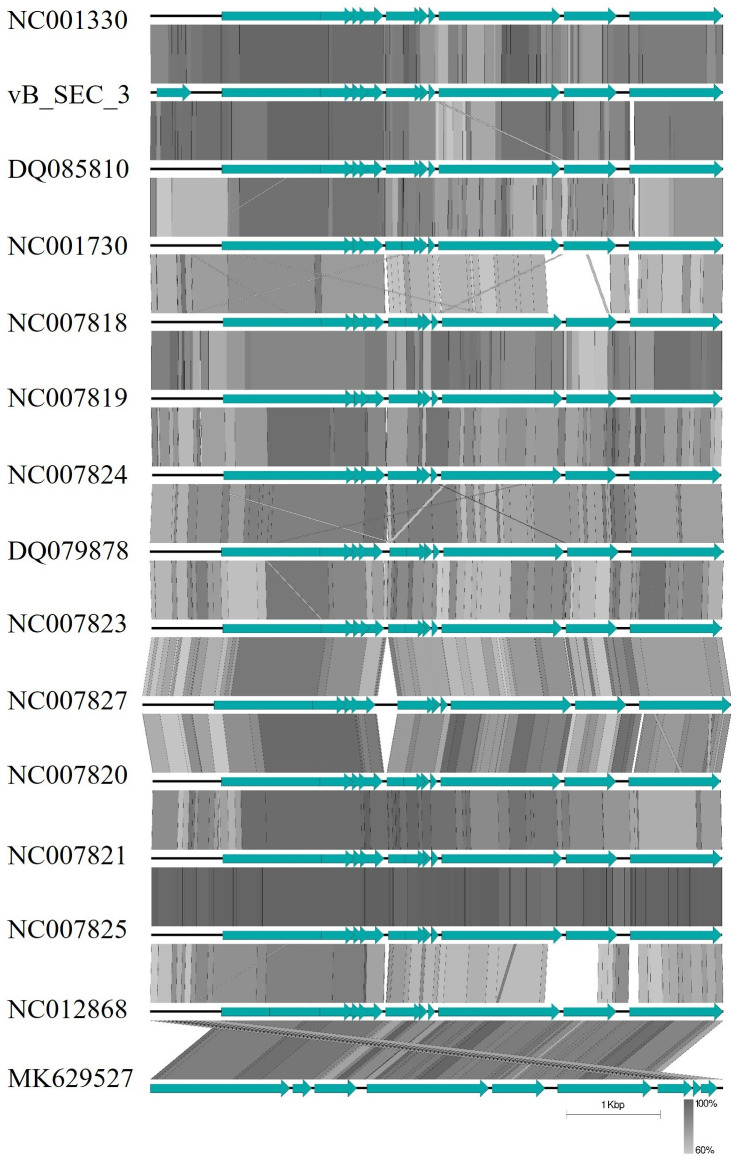 Figure 5
Figure 5 Figure 6
Figure 6- —National Recovery and Resilience Plan of the Republic of Bulgaria
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Monoclonal and Polyclonal Antibodies Research · Fecal contamination and water quality
1. Introduction
Historical records of antibiotic discovery and their use date back to the 19th century [1]. Since then, up to the present day, the development of various antibiotic formulations and their application in the treatment of severe bacterial infections have undergone dramatic changes, ultimately resulting in the widespread emergence and development of antibiotic-resistant bacteria (ARB), primarily due to their uncontrolled and inappropriate overuse. Nowadays, antimicrobial resistance (AMR) is considered a global concern, having an impact not only on human and animal health but also on economic, social, and environmental systems [2]. Overall, water is recognized as a major pathway for the spread of AMR across humans, animals, soil, and food systems [3]. In particular, wastewater serves as a key reservoir of AMR due to the presence of antibiotic residues, resistant bacteria, and resistance genes, promoting selective pressure and horizontal gene transfer (HGT), which accelerate its dissemination [4,5].
The bacterial pathogens most frequently identified as actively involved in the transfer of AMR in wastewater include Enterococcus faecalis, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, Enterobacter spp., and Escherichia coli [6,7]. The latter species is a normal inhabitant of the gastrointestinal tracts (GITs) of humans and animals and is consequently present in fecal excreta. For this reason, E. coli has been widely used as an indicator microorganism for fecal contamination in water [8]. The species is among the leading causative agents of severe diseases and was listed by the World Health Organization (WHO) as a priority bacterial pathogen in 2024 [9]. Moreover, according to the Global Burden of Disease study, nearly 50% of AMR-attributable deaths in high-income countries are associated with E. coli and S. aureus [10]. Additionally, E. coli possesses multiple mechanisms that confer resistance to a wide range of antibiotics (particularly to β-lactams), resulting in multidrug resistance, and its ability to readily transfer this resistance further increases the concern [11]. Thus, in light of pessimistic projections indicating that the global mortality associated with ARB could reach ten million deaths per year by 2050 if urgent action is not taken to address the problem, new and effective treatment strategies should be developed and implemented to manage E. coli-associated diseases, particularly those caused by MDR strains [12]. Thus, following a decline in interest after the discovery of antibiotics, bacteriophages (phages for short) and phage therapy have regained scientific attention in recent years as promising alternatives or adjuncts to conventional antibiotic treatments.
As a target bacterial pathogen, E. coli is extensively investigated for the development of phage-based formulations. In brief, the majority of published studies have primarily focused on the therapeutic potential of double-stranded DNA (dsDNA) tailed bacteriophages formerly classified within the families Siphoviridae, Myoviridae, and Podoviridae (order Caudovirales), now included in the class Caudoviricetes, as well as on the recently established taxon within this class, the subfamily Stephanstirmvirinae [13,14,15,16,17,18,19]. However, E. coli serves as a host for a far greater diversity of phages, which, for various reasons, have remained relatively poorly studied. Such is the case, for example, with virulent single-stranded (ss) DNA (+) phages of the family Microviridae. Representatives of this viral family, along with crAss-like phages, are considered among the most stable colonizers of the healthy human gut phagome, as shown by longitudinal metagenomic analyses and genome-based taxonomic clustering [20]. On the other hand, summarized data indicate that in non-GIT habitats, such as wastewater, these phages occur at lower abundances than tailed dsDNA phages, typically comprising only 2% to 37% of all somatic coliphages (SC), which could explain this gap [21,22,23,24]. Members of the family Microviridae are classified into two subfamilies, Gokushovirinae and Bullavirinae. The latter comprises three genera that include phages infecting members of the family Enterobacteriaceae and E. coli, in particular Alphatrevirus (α3 phages), Gequatrovirus (G4 phages), and Sinsheimervirus (phiX174 phages) [25]. Despite their lytic nature, these phages are normal members of the human gut phagome, which likely renders them tolerated by the immune system, potentially making them less susceptible to recognition by the human immune system and subsequent excessive inflammatory responses following potential administration [26]. However, in light of the development of phage therapy against pathogenic and MDR E. coli, the potential of Microviridae phages remains relatively poorly explored, as only a limited number of studies highlighting their therapeutic potential have been reported [26,27,28].
Based on the limited data on the less-explored use of Microviridae phages in phage therapy against E. coli, the present study aimed to contribute to the existing knowledge regarding the suitability of these phages for phage therapy. The object of the study was a newly isolated phage (vB_SEC_3), found to be a member of the genus Alphatrevirus, family Microviridae. The obtained data showed that the phage possesses several desirable characteristics—lytic life cycle, clear genome (no known lysogeny, virulence, and antibiotic resistance genes were found), good tolerance to pH and temperature, good survival rate at common laboratory storage (4 °C) and, most importantly, capable of lysing MDR E. coli strains—which highlight its potential for application in phage therapy. Based on this, we believe that our study contributes to the global understanding of the biology and diversity of Microviridae phages and further advances research highlighting their potential in combating MDR bacteria.
2. Materials and Methods
2.1. Collection and Processing of the Wastewater Samples
Wastewater treatment plants (WWTPs) of three of the largest cities in Bulgaria—Sofia, Burgas, and Varna—were selected for sampling. The sampling period covered two months from February to March 2024. Influent water samples (n = 6, representing one monthly sample collected from each of the three sampling sites) were collected in sterile 1 L plastic containers and transported under refrigerated temperature (4 °C) to the laboratory, where they were processed immediately. The initial processing of the raw water samples was as described elsewhere [29]. After removing the large debris from the samples, they were used for the isolation of both bacteria and phages.
2.2. Isolation and Identification of E. coli from Wastewater
For the purposes of this study, a total of fifteen bacterial isolates, presumably belonging to the E. coli species, were obtained from wastewater samples (Table 1). Endo agar (EA) was used as a selective medium for the targeted isolation of E. coli (Merck KGaA, Darmstadt, Germany). One milliliter from each processed wastewater sample was serially 10-fold diluted in sterile saline (pH 7.0), and 100 µL aliquots from dilutions up to 10^−3^ were surface-inoculated onto EA and incubated for 24 h at 37 °C. Only single colonies exhibiting a characteristic morphology (red colonies with a permanent metallic sheen), typical of E. coli growth on this medium, were isolated. Pure bacterial cultures were obtained after three consecutive cultivations on non-selective medium nutrient agar (NA) (Merck KGaA, Darmstadt, Germany) and stored at −20 °C for further analyses. Subsequently, all isolates were Gram-stained, and the DNA from all isolates was extracted from overnight broth cultures in nutrient broth (NB) using a commercial Genomic DNA Purification Kit, following the manufacturer’s instructions (Thermo Fisher Scientific Inc., Waltham, MA, USA). Species identification was performed via species-specific PCR with primer pair UAL-754 (5′-AAAACGGCAAGAAAAAGCAG-3′)/UAR-900 (5′-ACGCGTGGTTACAGTCTTGCG-3′) [30]. The reaction mixture (25 µL) contained 16.5 µL sterile ultra-pure H_2_O, 6.5 µL Red Taq DNA Polymerase 2X Master Mix 2.0 mM MgCl_2_ (VWR, Darmstadt, Germany), 0.5 µL of each primer, and 1 µL DNA template. The reaction conditions were as described by Bej and colleagues [30]. The resulting amplicons were separated in 1.5% agarose gel and observed under UV light. The detection of a 147 bp product was considered reliable evidence for E. coli identification.
2.3. Phenotypic Antibiotic Susceptibility Testing of the Wastewater-Derived E. coli Isolates
The phenotypic antibiotic susceptibility (PAS) of the wastewater-derived E. coli isolates to seventeen antibiotics, representatives of eight antimicrobial categories (Ampicillin (AMP), 10 µg/disk; Ampicillin/Sulbactam (A/S), 10–10 µg/disk; Amoxicillin/Clavulanic acid (AMC), 20–10 µg/disk; Cefuroxime (CXM), 30 µg/disk; Imipenem (IPM), 10 µg/disk; Meropenem (MRP), 10 µg/disk; Aztreonam (AT), 30 µg/disk; Levofloxacin (LE), 5 µg/disk; Norfloxacin (NX), 10 µg/disk; Pefloxacin (PEF), 5 µg/disk; Amikacin (Am), 30 µg/disk; Gentamicin (GEN), 10 µg/disk; Eravacycline (ERV), 20 µg/disk; Tigecycline (TGC), 15 µg/disk; Fosfomycin (FO), 200 µg/disk; Nitrofurantoin (NIT), 100 µg/disk; Trimethoprim (TR), 5 µg/disk) was screened according to EUCAST recommendations (break point table version 15.0, valid from 1 January 2025) [31]. Log-phase bacterial cultures were obtained after overnight cultivation on NA at 37 °C. Bacterial suspensions were prepared in sterile saline (10^8^ CFU/mL, McFarland standard 0.5) and surface inoculated on Mueller–Hinton agar plates (MHA, Merck KGaA, Darmstadt, Germany). After placing the antibiotic-impregnated paper disks (HiMedia, Mumbai, India), the Petri dishes were cultivated at 37 °C for 16–24 h. The interpretations of the results were carried out according to EUCAST recommendations. Isolates exhibiting resistance to at least three classes of antimicrobials were considered MDR [32].
2.4. Bacteriophage Isolation, Purification, and Propagation
Prior to phage isolation, one wastewater sample (collected from the WWTP near Sofia in March) was passed through a 0.22 µm pore-size membrane filter (Corning Incorporated, Corning, NY 14831, USA) to obtain a bacteria-free suspension. A non-pathogenic laboratory strain of E. coli, NBIMCC 8432, was used as the initial host strain for bacteriophage recovery from wastewater. The bacterial strain was initially cultivated on NA for 24 h at 37 °C and subsequently used for the preparation of a bacterial suspension in sterile saline (10^8^ CFU/mL). Double agar overlay plaque assay (DAOPA) was applied for phage isolation [33]. A mixture containing equal volumes (100 µL) of bacterial suspension and filtered wastewater sample, 30 µL 1 M CaCl_2_, and 3 mL semisolid NA (0.45% agar content) was poured onto solid NA agar plates supplemented with CaCl_2_ to a final concentration of 10 mM (NA-Ca). Double agar petri dishes were cultivated for 24 h at 37 °C. A selected single plaque resulting from bacterial lysis was picked with a sterile plastic needle and transferred into 10 mL of sterile NB-Ca (10 mM CaCl_2_ content), followed by cultivation (37 °C, 24 h) with the host strain. Pure phage lysate was obtained after membrane filtration of the obtained culture through a membrane filter (pore size 0.22 µm). Phage purification was accomplished following three successive cultivations in NB-Ca with the corresponding host strain. The newly isolated phage was stored as pure lysate at 4 °C and used for further analyses.
2.5. Study of the Phage Host Range and Efficiency of Plating Against MDR E. coli Strains
Spot testing assay (STA) was applied to determine the host range of the newly isolated phage [34]. In total, twenty-seven bacterial strains, with representatives of eleven bacterial species (E. coli, E. cloacae, E. asburiae, E. kobei, K. oxytoca, K. pneumoniae, A. baumannii, A. guillouiae, P. mirabilis, S. aureus, and P. aeruginosa) were used. Fifteen of the E. coli strains used were isolated from wastewater in this study, and one was an ATCC strain isolated from feces (Table 1). Non-Escherichia species that were not obtained from publicly available repositories (ATCC, NBIMCC) used in this study were part of a laboratory collection of the Department of General and Industrial Microbiology, Sofia University “St. Kliment Ohridski”, and were previously isolated and identified (data not published). Log-phase bacterial cultures obtained after overnight cultivation on NA were used to prepare bacterial suspensions (10^8^ CFU/mL) in sterile saline. Solid NA-Ca plates were prepared, and mixtures containing 100 µL of the respective bacterial suspension, 10 mM CaCl_2_, and 3 mL semisolid NA-Ca were poured onto them. Ten µL of pure phage lysate (10^9^ PFU/mL), obtained after cultivation of the newly isolated phage with its initial host (E. coli 8432), was spotted onto a semisolid agar surface, and the petri dishes were cultivated at 37 °C for 24 h. The observation of the bacterial lysis area after cultivation was considered evidence for bacterial susceptibility to phage infection. The relative efficiency of plating (EOP) of the phage on susceptible E. coli strains was further evaluated. Briefly, double-layer agar plates were prepared as described above. The phage lysate was serially diluted tenfold in phage buffer up to 10^−7^, and 10 µL aliquots of each dilution were spotted onto the agar surface [35]. The relative EOP was calculated as the ratio of average PFU/mL obtained for the tested strain to the average PFU/mL obtained for the original host strain E. coli 8432 [36]. The EOP of the phage on E. coli 8432 was considered 1. The experiment was conducted in triplicate, and the results were expressed as mean ± SD.
2.6. Determination of Plaques and Virion Morphology of the Phage
Plaque morphology was examined on single plaques formed after 24 h of cultivation on E. coli 8432 lawn using DAOPA. The general plaque appearance (clear or turbid; presence or absence of a halo) and plaque dimensions (measured with an electronic caliper) were recorded. Measurements were conducted on at least ten individual plaques, and the final dimensions were expressed as the mean ± SD. The virion morphology was studied via transmission electron microscopy (TEM). Fifty μL of pure phage lysate (10^10^ PFU/mL) was spotted onto pre-coated Formvar grids for TEM analyses. The grids were negatively stained with 1% uranyl acetate in 70% methanol for 30 min in the dark. The observations were carried out on Hitachi HT7800, operating at 100 kV (Hitachi High-Tech Corporation, Tokyo, Japan). Phage dimensions were determined from at least ten individual measurements of single virions and were reported as mean ± SD.
2.7. Determination of Optimal Phage-to-Bacteria Ratio for Yielding High-Titer Lysates
The optimal phage-to-bacteria ratio, resulting in the highest phage titers (i.e., multiplicity of infection, MOI), was determined for the newly isolated phage. The phage isolate and its respective bacterial host (E. coli 8432) were cultivated in 10 mL NB-Ca at different ratios corresponding to MOI values of 0.01, 0.1, 1, 10, and 100. After incubation at 37 °C for 4 h, phage titers were measured, and the MOI yielding the highest titers was considered optimal for phage propagation. The experiments were carried out in triplicate, and the final titers were expressed as mean ± SD.
2.8. Influence of Temperature and pH on Phage Viability
Pure phage lysates with known titers were stored at 4 °C for 6 months. Phage stability under these non-specific laboratory conditions was evaluated by comparing titers at the start of the experiment with those measured after the storage period. Phage stability under various temperatures and pH was studied according to the procedures described by Park and colleagues, with slight modifications [37]. Pure phage lysates were prepared in NB-Ca, and the initial titer was counted via STA with E. coli 8432 as the host. Phage viability at different temperatures was evaluated by measuring titer reductions following 2 h incubation of 1 mL of pure phage lysate at 28 °C, 37 °C, 50 °C, 65 °C, 80 °C, and 95 °C, respectively. The analyses were carried out in a heating dry block under stable thermal conditions (BioSan, Riga, Latvia). The effect of pH on phage viability was evaluated by mixing 100 µL of pure phage lysate with known titer with 900 µL of phage buffer (10 mM Tris–HCl, 10 mM MgSO_4_, 68 mM NaCl, and 1 mM CaCl_2_) adjusted to pH 2.0, 4.0, 5.92, 7.5, 9.0, 10.5, and 13.0. The mixtures were incubated for 3 h, after which the number of viable phage particles was determined via STA. Reductions in phage titers were interpreted as evidence of phage susceptibility to the respective pH. The experiments were carried out in triplicate, and the final results were expressed as mean ± SD.
2.9. Phage Genome Sequencing and Feature Analysis
Crude phage lysate was prepared in 30 mL NB-Ca according to optimal MOI, and the titer was assessed via STA (10^9^ PFU/mL). Phage genetic material was extracted following the previously described method [29]. Sequencing was performed at Microbes NG (Birmingham, UK) using a hybrid approach that combined Oxford Nanopore Technologies (R10.4.1 flow cells) with 2 × 250 bp Illumina short reads. Raw reads were analyzed in Phage Galaxy by Kraken (Galaxy Version 1.3.1), metaviralSPAdes (Galaxy Version 4.2.0+galaxy0), and Megahit (Galaxy Version 1.2.9+galaxy2) [38,39,40,41]. The assembled genome was analyzed in CLC sequence viewer (Version 7.6) and annotated both manually and using two web-based services, Pharokka software (Galaxy Version 1.3.2) and the RAST Server: Rapid Annotations using Subsystems Technology [42,43]. The presence/absence of tRNA, virulence-associated, and antibiotic resistance genes in the phage genome was screened via tRNA prediction (tRNAscan) (Galaxy version 0.4) and ABRicate (Galaxy Version 1.0.1.), respectively [44,45].
2.10. Phylogenetic Analyses Based on Whole Genome Comparison
The phylogenetic relationship between the newly isolated phage and related bacteriophages was analyzed using the web-based platform VICTOR, a widely used tool for genome-based phylogeny and classification of prokaryotic viruses [46]. FASTA sequences of the complete genomes of 40 phages, members of the family Microviridae, were obtained from GenBank and used in the analyses. The phylogenetic tree was constructed based on pairwise comparisons of whole-genome nucleotide sequences using the Genome-BLAST Distance Phylogeny (GBDP) method with parameters recommended for prokaryotic viruses (including 100 pseudo-bootstrap replicates) [46,47]. Comparative genome alignment and the corresponding graphical representation were performed using EasyFig v.2.2.5. [48]. Sequence similarity between the genome of the newly isolated phage and its relatives was calculated using VClust v.1.2.8., a web-based tool that performs average nucleotide identity (ANI)-based genome clustering in accordance with the standards of the International Committee on Taxonomy of Viruses (ICTV) [49].
2.11. Statistical Analysis
The data were analyzed by Microsoft Excel v. 2508 using XLMiner Analysis ToolPak with ANOVA single factor. The number of antibiotic resistance profiles among isolates from different cities and sampling times was analyzed by Welch’s t-test. A p-value < 0.05 was considered statistically significant.
3. Results
3.1. Assessment of the Phenotypic Antibiotic Resistance Profiles of the Newly Isolated E. coli Strains
For the purposes of this study, fifteen bacterial isolates (WWTP Sofia (n = 6), WWTP Burgas (n = 6), and WWTP Varna (n = 3)) were obtained following cultivation of the wastewater samples on the selective EA (Table 1). Presumptive E. coli strains were selected based on colony morphology on EA, characterized by a red color with a permanent metallic (fuchsin) sheen, indicative of lactose fermentation (Figure 1A). All isolates were found to be Gram-negative short rods and were subsequently identified as E. coli based on species-specific PCR analysis, with all isolates yielding the expected 147 bp PCR product [30]. PAS testing showed that all newly isolated E. coli strains were susceptible to eight out of the seventeen antibiotics tested, representing five antimicrobial categories (aminoglycosides, tetracyclines, carbapenems, miscellaneous agents, and penicillin + β-lactamase inhibitors). The same strains were 100% resistant to five other substances, representing four antimicrobial categories—phosphonic acid derivative antibiotics, a unique class of antimicrobials (FO), cephalosporin antibiotics (CXM), monobactams (AT), penicillin/penicillin + β-lactamase inhibitors (AMP and AMC, respectively)—in accordance with EUCAST guidelines (Supplementary Table S1). Additionally, resistance to antibiotics in two more antimicrobial categories—quinolone/fluoroquinolone antibiotics (NX and PEF) and dihydrofolate reductase inhibitors (antifolates) (TR)—was also detected in individual isolates (B2, B6, and V7). The most antibiotic-resistant isolate was found to be V7. Based on the obtained results and in accordance with the guidelines listed in EUCAST, it can be concluded that all E. coli strains isolated in this study could be classified as MDR [32]. However, no statistically significant differences were observed when comparing the number of antibiotic-resistant profiles among isolates obtained from different WWTPs (cities of Sofia/Varna, Varna/Burgas, and Sofia/Burgas; p = 0.423, p = 0.788, and p = 0.185, respectively), nor between the two sampling time points (February and March; p = 0.926).
3.2. Bacteriophage Isolation, Host Range Determination, and EOP Evaluation
A non-pathogenic E. coli 8432 strain was used as the initial host for phage isolation from wastewater (Table 1). Following cultivation of the wastewater sample with the host bacterium, several plaques with distinct morphologies were observed on the surface of the double-agar plates. One small, clear plaque was chosen for phage isolation (Figure 1B). After purification via three consecutive cultivations with the host bacterium, a pure phage culture was obtained. The newly isolated phage was designated as vB_SEC_3. The host range analysis revealed that the phage can lyse only representatives of the species E. coli. Beyond the phage activity against its initial host (strain 8432), vB_SEC_3 was found to lyse another three strains (S1, B5, and 8739) out of the 16 tested E. coli strains (18.6%). No lytic activity was observed against the other tested species (Table 1). On this basis, we can conclude that vB_SEC_3 has a narrow host range. Moreover, two of the susceptible E. coli strains were defined as MDR (S1 and B5) based on their antibiotic resistance profiles (Supplementary Table S1). Therefore, we further evaluated the phage EOP against these two strains. The results revealed that vB_SEC_3 was highly effective against MDR strains S1 and B5, with EOP of 0.735 and 0.961, respectively. Although these differences in the original host were found to be statistically significant (p < 0.05), it can be concluded that the phage demonstrates high efficacy against both isolates, effectively lysing them.
3.3. Investigation of the Morphology of vB_SEC_3
Phage’s basic characterization includes the determination of both plaque and virion morphology. The newly isolated phage formed plaques with a small clear center with a diameter (d, mm) of 1.473 ± 0.197 surrounded by a turbid zone (d = 3.215 ± 0.35 mm) on an E. coli 8432 lawn (Figure 2A). The virion morphology, examined by TEM, was found to be similar to that of Microviridae phages, displaying a small icosahedral capsid (45.16 ± 2.01 nm in diameter) with no tail structures observed (Figure 2B).
3.4. Determination of the Optimal MOI of the Phage
Determination of the optimal MOI is an essential step in characterizing a newly isolated phage, as it identifies the best phage-to-bacteria ratio in the cultivation medium to achieve high-titer lysates. The comparison of the obtained average final titers with ANOVA revealed no statistical differences (p = 0.206). However, out of five tested ratios, we established that the highest average titer was obtained in MOI 1 (8.068 PFU/mL ± 1.36) (the ratio between phage particles and bacterial cells in the cultivation medium is equal), after 4 h cultivation with the initial host in NB-Ca (Figure 3A). Thus, this phage-to-bacterium ratio was selected as optimal and applied in subsequent phage propagation experiments where appropriate.
3.5. Tolerance of vB_SEC_3 to Various pH Values and Temperatures
In this study, we investigated the effect of seven pH values ranging from 2.0 to 13.0 on phage viability. Temperature tolerance was evaluated in two aspects: the effect of storage temperature (4 °C) on phage viability, and the thermal stability of virion particles across a temperature range of 28 °C to 95 °C. Generally, no viable phage particles were detected after incubation of vB_SEC_3 at extremely acidic and extremely basic conditions (pH 2.0 and 13.0). At the same time, the phage exhibited strong tolerance across a wide pH range (4.0–10.5) (Figure 3B). ANOVA analysis revealed no statistically significant differences in phage titers (p = 0.311) following incubation under these pH conditions, indicating that phage activity was not adversely affected. The effect of standard refrigeration conditions (4 °C) on phage viability and ability to lyse its host bacterium was studied over a six-month period by comparing phage titers measured before and after storage. The obtained results showed a small decrease in phage titer across the tested period (up to 0.16 log_10_ PFU/mL). Briefly, no statistically significant differences in phage titers compared to the initial values (9.67 ± 0.58 log_10_ PFU/mL) were observed after incubation at temperatures up to 65 °C (p = 0.302), indicating that this temperature range had no adverse effect on phage activity (Figure 3C). Incubation at 80 °C resulted in a significant reduction in phage titer (a decrease by 6.72 log_10_ PFU/mL) compared to the initial value. This finding was supported by ANOVA, which revealed highly significant differences in phage titers between samples incubated at 65 °C and 80 °C (p < 0.05). No viable phage particles were detected after incubation at 95 °C.
3.6. Genome Organization of vB_SEC_3
The genetic material of the newly isolated phage was extracted, and the complete genome was sequenced, after which the obtained raw reads were processed. The total number of reads mapped with the Kraken software (Galaxy Version 1.3.1) to the Microviridae family was 33,742, which, after assembling, yielded a single genome with completeness of 100% with a coverage of 1663×. The annotated genome was deposited at GenBank under the accession number PX907427. The phage was found to possess one circular fragment of single-stranded DNA (+) (ssDNA) with 6085 bp length and GC content of 45.3%. The genome contained nine open reading frames (ORFs) and 11 coding sequences (CDS) and was found to possess a typical organization of known microvirus phages, with the most distinctive feature being the presence of overlapping reading frames (Figure 4). In brief, the genome of vB_SEC_3 contains three genes (B, K, and E) embedded within other CDS. These three genes were annotated as encoding protein K with unknown function, protein E, involved in cell lysis, and the highly flexible internal scaffolding protein B. The remaining seven genes were annotated as coding for products involved in virus replication (A and C), virion morphogenesis (D and J), or structural (capsid) proteins (F, G, and H). Functional annotations were derived from Kodaira and colleagues [50]. No genes associated with lysogeny, antibiotic resistance, virulence, or tRNA-related regions were identified in the genome of vB_SEC_3. Preliminary whole-genome BLASTN analysis revealed high similarity to two Enterobacteria (Escherichia) phages alpha3 (NC001303 and DQ085810), with nucleotide identities of 94.99% and 95.06%, respectively, and 100% query coverage. Subsequently, comparative genome alignment was performed to further analyze similarities and differences in genome organization and ORF positioning between vB_SEC_3 and other known α3 phages, revealing a high degree of genetic similarity (Figure 5).
3.7. Comparative Genome Analyses and Phylogenetic Relationships
To more precisely determine the taxonomic position of the newly isolated phage, a phylogenetic tree was constructed based on pairwise comparisons of whole-genome nucleotide sequences of vB_SEC_3 and other members of the family Microviridae (Figure 6). Complete genome sequences of 37 bacteriophages belonging to the established groups α3, phiX174, and G4 within subfamily Bullavirinae were retrieved from GenBank and used for comparative analysis. The results showed that the newly isolated phage clustered together with the two phages mentioned above (NC001303 and DQ085810), revealing the phylogenetic relationship between them, thus confirming that these phages are the closest relatives of vB_SEC_3. Both phages are also known as Alphatrevirus alpha3 and are classified within the genus Alphatrevirus. Based on these findings, we could assume that vB_SEC_3 is a member of the group α3 phages within the family Microviridae, genus Alphatrevirus. To further validate these findings, intergenomic pairwise similarity analyses were conducted among the α3 group members (n = 14) included in the phylogenetic analysis. The calculated total ANI (tANI) confirmed our previous assumption, as high nucleotide identity was observed between vB_SEC_3 and the two closest relatives mentioned above (94.8% and 94.9%, respectively) (Table 2, Supplementary Figure S1). Nevertheless, despite the observed similarity percentages, which remained below 95%, vB_SEC_3 meets the ICTV criteria for genus and species demarcation (70% and 95%, respectively), suggesting it is a putative novel species within the genus Alphatrevirus.
4. Discussion
The rapid global increase in AMR, alongside the declining pipeline of effective antibiotics, has prompted renewed interest in alternative treatments, notably phage therapy [51]. Supporting global efforts to combat AMR and advancing phage therapy—both worldwide and locally—requires comprehensive, coordinated research, beginning with the exploration of diverse bacteriophages and the evaluation of their therapeutic potential against targeted bacterial pathogens. While this rediscovered approach has gained increased visibility and experienced rapid growth in Western regions, phage research in countries such as Bulgaria has accelerated over the past several years, building on earlier, largely isolated investigations [52]. In this regard, the present study describes the characterization of a newly isolated bacteriophage (vB_SEC_3) and presents data on its capacity to target and lyse E. coli, with a particular focus on MDR strains.
Influent wastewater (WWTP, Sofia, Bulgaria) was used as the source for phage isolation, consistent with other studies reporting the isolation and characterization of phages targeting enterobacteria, particularly E. coli [19,28,51,53,54]. A non-pathogenic E. coli strain (8432) was used as the initial host for phage isolation to maximize the chances of phage discovery, avoiding the naturally occurring resistance often observed in wild E. coli strains from the same habitat, as previously suggested [53]. The host range of the phage was examined across a wide range of test bacteria, most of which were representatives of the family Enterobacteriaceae. For the purposes of this study, alongside phage isolation, a total of fifteen E. coli isolates were obtained from influent wastewater and included in the host range analyses. The tested PAS of all newly isolated E. coli strains revealed that all of them exhibit MDR, primarily showing resistance to β-lactam antibiotics (CXM, AT, AMP, and AMC) along with FO. According to analyses from the Antimicrobial Testing Leadership and Surveillance (ATLAS) project, resistance to cephalosporins is central to classifying these strains as critical pathogens [55]. Additionally, the observed resistance to FO—an antibiotic previously recommended for treating urinary tract infections caused by extended-spectrum β-lactamase (ESBL- producing bacteria, and generally exhibiting low resistance rates—raises concerns about the efficacy of currently available effective antibiotics against MDR E. coli strains [56,57]. In this regard, we established that although vB_SEC_3 exhibited a narrow host range (targeting only E. coli strains), it was able to target two MDR E. coli isolates (S1 and B5), showing EOP > 0.5 (0.735 and 0.961, respectively) relative to the initial non-pathogenic strain (8432). According to the widely accepted EOP scale, in which phage infectivity is classified as high (EOP ≥ 0.5), moderate (0.2–0.5), low (0.001–0.2), and inefficient (≤0.001), the newly isolated phage exhibited a high EOP, highlighting its potential against MDR E. coli strains [58].
The genotypic and morphological characterization of vB_SEC_3 revealed that it possessed features typical of the known α3 phages, members of the genus Alphatrevirus, subfamily Bullavirinae, family Microviridae. These are small, non-enveloped, ssDNA (+) phages that exhibit a highly conserved genome organization and exclusively target members of the Enterobacteriaceae family [59,60]. Our observations established that vB_SEC_3 possessed a small icosahedral capsid with no visible tail structures, typical of Microviridae phages, although its dimensions (d = 45.16 ± 2.01 nm) were larger than previously reported [24,61]. Indeed, our morphological analyses were performed using TEM, following the standard pipeline for phage characterization, whereas most studies examining capsid structure, assembly, and mature virions of Microviridae phages employ primarily cryo-electron microscopy (cryo-EM). Notably, virion diameters measured via cryo-EM are typically reported in angstroms, which could partly explain this discrepancy [27,28,62]. Sequencing analyses revealed that the genome organization of vB_SEC_3 was highly similar to that previously reported for the Microviridae phages [50]. The taxonomic position of the newly isolated phage was confirmed through phylogenetic and comparative genome alignment analyses, which identified its two closest relatives—NC001303 and DQ085810—both members of the genus Alphatrevirus, family Microviridae. Moreover, based on the calculated tANI and the criteria established by the ICTV, vB_SEC_3 has the potential to be considered a putative novel species within this genus. Interestingly, the phylogenetic analysis based on the whole genome sequence similarity positioned the new vB_SEC_3 phage in one of two distinctive subclusters of α3 phages (one represented by the reference genome, acc. No. NC001303, the other by coliphage NC35, acc. No. NC007820) (Figure 6). Similarly, two distinct groups of α3 phage genome sequences, one of which included vB_SEC_3, were observed when comparing them by tANI calculation (Table 2, Supplementary Figure S1). These two analyses suggest possible intrageneric diversification within α3-like phages, an observation that merits further attention.
Additionally, the genome features of vB_SEC_3 were studied in the context of its suitability for application in phage therapy; no known genes for lysogeny, tRNA, virulence, or antibiotic resistance were established. The gene encoding phage lysis protein (gene E) was also established in the genome of vB_SEC_3. Interestingly, ssDNA phages mediate cell lysis in a manner fundamentally different from the endolysin–holin system used by dsDNA phages, although this process remains poorly explored [63]. Gene E encodes a membrane protein that inhibits MraY, an enzyme essential for murein synthesis, functioning similarly to penicillin antibiotics, which are known to inhibit cell wall synthesis [64]. Given its universal presence and essential role in bacterial survival, MraY is a promising target for antibacterial drug development [65]. With the rise of resistance to β-lactam antibiotics (also observed in our analyses) through the development of ESBL-producing bacteria, it is intriguing to consider whether MraY inhibitors, such as those encoded by the Microviridae phages, could offer therapeutic benefits against these resistant strains. Further research is warranted, as these phages show promising potential against ESBL-producing bacteria.
Nevertheless, despite their potential against MDR bacteria, these phages have been studied mostly in terms of their distribution, role in host-phage interactions in the human and animal gut, and genetic diversity, rather than their therapeutic applications, with few limited exceptions [26,27,28,66,67,68,69]. This discrepancy may arise because standard DNA isolation and sequencing protocols often fail to detect small ssDNA genomes, as successful isolation using conventional methods requires the phage DNA to be in its double-stranded form, which represents a transient stage in the replication strategy of ssDNA phages (known as rolling circle) [61]. In contrast, our investigation focused primarily on general phage characteristics most frequently studied for phages intended for phage therapy. The newly isolated phage was assessed for long-term viability under non-specific laboratory conditions (4 °C), as well as for thermal inactivation and pH stability. Despite its distinct biology, the newly isolated phage exhibited propagation patterns similar to those of the more extensively studied dsDNA-tailed phages, including its stability across a wide pH range of 4.0–10.5 and thermal inactivation following incubation at temperatures of 65–80 °C [29,58]. We established that only a slight decrease in phage titer occurred after storage of the crude phage lysate for 6 months at 4 °C, and that the phage produced a high number of progeny within 4 h at an MOI of 1. Such rapid production of progeny, facilitated by the small genome size and the phage’s employed replication strategy, can be considered desirable for phages intended for phage therapy. On the other hand, such phages are more prone to mutations, and although they are considered obligately lytic, in the context of phage therapy, genetic variability and potential adaptation dynamics should be monitored [61].
5. Conclusions
In the rapidly evolving field of strategies aimed at combating antibiotic resistance among pathogenic bacteria, particularly through bacteriophage-based approaches, this study provides new data highlighting the potential of the newly isolated phage vB_SEC_3 against MDR E. coli strains. As a representative of the relatively understudied family Microviridae in the context of phage therapy, the obtained results support further investigation of Microviridae phages as potential antibacterial agents. Moreover, the bacterial pathogen addressed in this study is among those of greatest interest for the development of phage-based therapies, as highlighted in the WHO report on antibacterial agents in clinical and preclinical development. A reference to this document shows that Annex 13, focusing on phage-based products and phage-derived enzymes, indicates that three out of ten reviewed products are designed to target E. coli [70]. In Bulgaria, previous investigations into the lytic potential of E. coli phages have been limited and sporadic, conducted decades ago, and the present study provides new data to advance this field [71]. However, further in-depth research is essential, particularly with regard to the phage’s efficacy against clinical MDR E. coli strains, the potential for resistance development, its pharmacodynamic and pharmacokinetic properties, the evaluation of endotoxins and enterotoxins in phage lysates, and cytotoxicity, before it can be considered for clinical application.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Durand G.A. Raoult D. Dubourg G. Antibiotic discovery: History, methods and perspectives Int. J. Antimicrob.20195337138210.1016/j.ijantimicag.2018.11.01030472287 · doi ↗ · pubmed ↗
- 2Sambaza S.S. Naicker N. Contribution of wastewater to antimicrobial resistance: A review article J. Glob. Antimicrob. Resist.202334232910.1016/j.jgar.2023.05.01037285914 · doi ↗ · pubmed ↗
- 3Prestinaci F. Pezzotti P. Pantosti A. Antimicrobial resistance: A global multifaceted phenomenon Pathog. Glob. Health 201510930931810.1179/2047773215 Y.000000003026343252 PMC 4768623 · doi ↗ · pubmed ↗
- 4Barancheshme F. Munir M. Strategies to combat antibiotic resistance in the wastewater treatment plants Front. Microbiol.20188260310.3389/fmicb.2017.0260329387043 PMC 5776126 · doi ↗ · pubmed ↗
- 5Amato M. DasíD. González A. Ferrús M.A. Castillo M.Á. Occurrence of antibiotic-resistant bacteria and resistance genes in agricultural irrigation waters from Valencia city (Spain)Agric. Water Manag.202125610709710.1016/j.agwat.2021.107097 · doi ↗
- 6Petrovich M.L. Zilberman A. Kaplan A. Eliraz G.R. Wang Y. Langenfeld K. Microbial and viral communities and their antibiotic resistance genes throughout a hospital wastewater treatment system Front. Microbiol.20201115310.3389/fmicb.2020.0015332140141 PMC 7042388 · doi ↗ · pubmed ↗
- 7Marano R.B. Gupta C.L. Cozer T. Jurkevitch E. Cytryn E. Hidden resistome: Enrichment reveals the presence of clinically relevant antibiotic resistance determinants in treated wastewater-irrigated soils Environ. Sci. Technol.2021556814682710.1021/acs.est.1c 0061233904706 · doi ↗ · pubmed ↗
- 8Abdelgalel R.R. Ibrahem R.A. Mohamed D.S. Ahmed A.B.F. Multidrug-resistant Escherichia coli in wastewater sources: A comparative study and identification of resistance hotspots BMC Microbiol.20252549810.1186/s 12866-025-04244-540790549 PMC 12341253 · doi ↗ · pubmed ↗