Effect of Epichloë Endophytes on Growth of Grass Pathogenic Fungi
Cuiling Wan, Xiuzhang Li, Qian Shi

TL;DR
This study explores how Epichloë fungi can inhibit the growth of plant pathogenic fungi, suggesting their potential as biocontrol agents.
Contribution
The study identifies specific Epichloë strains with strong antifungal activity and links alkaloid concentrations to inhibition rates.
Findings
Twelve Epichloë strains showed varying inhibition rates against five pathogenic fungi.
Strain F2 inhibited spore germination of B. sorokiniana by 87.73%, while strain H3 had minimal inhibition on F. avenaceum.
Ergonovine and ergine concentrations correlated with antifungal activity, though the strength varied by pathogen.
Abstract
Endophytic fungi widely colonize plant tissues without causing severe disease, protect hosts from pathogenic microorganisms, and represent a key potential resource for novel biocontrol agents. To explore the biocontrol potential of endophytic Epichloë fungi and their correlation with alkaloids, 12 Epichloë strains were isolated from six different geographic sites of Festuca sinensis, Achnatherum inebrians, and Hordeum brevisubulatum. The antifungal activity of these strains was evaluated against five phytopathogenic fungi (Alternaria alternata, Bipolaris sorokiniana, Curvularia lunata, Fusarium avenaceum, and Drechslera erythrospila) using dual-culture assays, which measured the inhibition of both colony growth and spore germination. Concurrently, the concentrations of ergonovine and ergine were quantified in the liquid cultures of each Epichloë strain. The results showed that 12…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
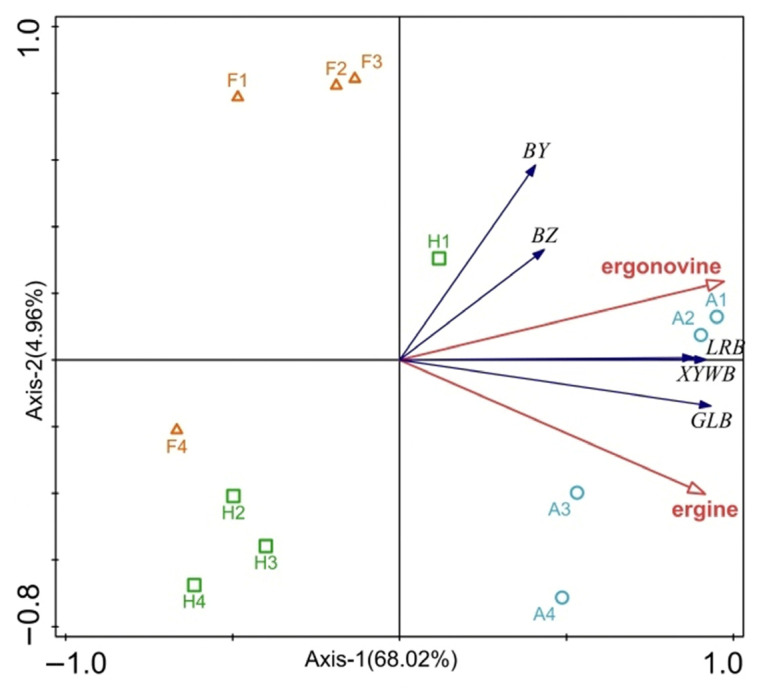 Figure 1
Figure 1- —The National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and fungal interactions · Turfgrass Adaptation and Management · Plant Pathogens and Fungal Diseases
1. Introduction
Grasses are threatened by numerous phytopathogenic microorganisms, which cause substantial damage to host plants [1]. A wide variety of pathogens can infect grasses, including fungi, bacteria, viruses, phytoplasmas, viroids, and nematodes. Among these, fungal pathogens account for more than 80% of total plant diseases [2]. Although considerable progress has been made in understanding fungal pathogenicity and developing control strategies over the past century, plant fungal diseases still represent a major constraint in agricultural production [3]. Plant diseases are the main factors affecting the health of natural grasslands. They not only reduce the yield and quality of forage grasses, but also restrict the sustainable utilization of grasslands and the development of animal husbandry [4]. By applying chemical pesticides, the losses caused by diseases can be mitigated to a certain extent. However, chemical pesticides not only cause environmental problems but also select for strains with resistance [5,6]. Therefore, applying beneficial endophytic microorganisms as biocontrol agents for disease management is an economical and effective strategy [7]. In pastoral, turfgrass, and natural grassland ecosystems, many cool-season grasses form symbiotic associations with endophytic fungi of the genus Epichloë. These endophytes are well known for their ability to produce antimicrobial metabolites, which effectively inhibit the growth of pathogens both in vitro and in planta [1,8,9,10].
Grass endophytes, comprising both sexual (Epichloë spp.) and asexual (Neotyphodium spp.) clavicipitaceous fungi (Clavicipitaceae, Hypocreales, Ascomycota), form symbiotic associations with some cool-season grasses (family Poaceae subfamily Pooideae) [11]. Nowadays, the Neotyphodium and Epichloë genera are commonly referred to collectively as Epichloë endophytes [12]. Endophytic fungi often exhibit host specificity, with each species typically colonizing only one or a few grass hosts [13]. These endophytes grow systemically in the aboveground plant parts, obtaining nutrients from the apoplast of developing leaves and stems near the shoot meristems [14]. In addition, they can be vertically transmitted via host seeds [15]. There is growing research interest in these fungal endophytes due to their ability to confer multiple benefits on host plants. These benefits include enhanced growth and increased tolerance to both abiotic stresses (e.g., drought, salt, temperature, and heavy metals) [16,17,18,19,20] and biotic stresses (e.g., herbivores and nematodes) [21]. The long-term co-evolution and stable symbiotic relationship between grasses and Epichloë fungi imply that these endophytes play an important role in plant–pathogen interactions [22]. Studies have shown that Epichloë endophytes can inhibit the growth and pathogenicity of various pathogenic fungi through multiple in vitro and in planta mechanisms [23]. However, this effect varies depending on the specific pathogenic fungus and endophyte strain. Although alkaloids have been hypothesized to play a role in plant–pathogen interactions [24], direct evidence for an association between alkaloid levels and pathogen suppression remains insufficient, and the underlying mechanisms warrant further investigation.
Achnatherum inebrians is one of the most toxic wild grasses in the northern natural grasslands of China, yet as a plant adapted to saline habitats in arid and semi-arid regions, it holds potential for ecological restoration and soil conservation [25]. Festuca sinensis and Hordeum brevisubulatum are important high-quality forage grasses in China. F. sinensis contributes to the restoration of degraded alpine meadows through soil conservation, while H. brevisubulatum shows promise in the improvement of saline–alkaline land; both grasses possess high forage value and play vital roles in maintaining regional ecological stability [26,27]. All three have a relatively high rate of Epichloë endophytic fungal infection in the natural grasslands [28,29,30,31]. The A. inebrians endophyte symbiotic system has been extensively studied in China’s grass endophyte research. Researchers have conducted comprehensive investigations into its physiological, biological, and chemical aspects [24,29,32,33]. Studies have shown that Epichloë can significantly enhance drought tolerance and powdery mildew resistance in A. inebrians [34,35]. The biological and ecological characteristics of the F. sinensis-Epichloë endophyte symbiont have also been characterized [28]. Similarly, the Epichloë symbiont improves seed germination and cold resistance in F. sinensis [17,19,36,37]. For H. brevisubulatum, endophyte infection significantly enhances host salt tolerance and waterlogging tolerance [38,39], while also improving nutrient uptake under salt stress [40]. Furthermore, studies have shown that the presence of Epichloë endophytes in H. brevisubulatum increases acetone content. This increase in acetone may significantly enhance antifungal activity against Alternaria alternata, Bipolaris sorokiniana, Fusarium avenaceum, and Trichoderma viride [41].
Evidence indicates that Epichloë endophytes can directly inhibit the growth of plant-pathogenic fungi [42,43,44], thereby enhancing host resistance [45,46,47,48]. To extend these findings and assess the potential role of specific alkaloids, it was hypothesized that geographically diverse Epichloë strains exhibit variable but direct antifungal activity in vitro, and further, that this activity is positively correlated with their concentration of the ergot alkaloids ergonovine and ergine. To test this hypothesis, in vitro dual-culture assays were conducted using 12 Epichloë strains isolated from three grass species (F. sinensis, A. inebrians, and H. brevisubulatum) across six sites, against five grass pathogenic fungi (Alternaria alternata, Bipolaris sorokiniana, Curvularia lunata, Fusarium avenaceum, and Drechslera erythrospila). The inhibitory effects on both colony growth and spore germination were measured, and the concentrations of ergonovine and ergine in liquid cultures were measured to analyze their correlation with antifungal activity. This work adds to the growing evidence of Epichloë-mediated pathogen inhibition and lays the groundwork for evaluating their potential as biocontrol agents in agricultural development.
2. Materials and Methods
2.1. Biological Materials
Endophytic fungi were isolated from F. sinensis, A. inebrians, and H. brevisubulatum with high infection rates. Plant samples were collected from six different sites, and two strains were selected from each site as experimental backups. The specific sampling locations, together with their latitude, longitude, and altitude, are listed in Table 1. The collected single-plant samples were examined under a microscope using the aniline blue staining method of stem pith [49] to determine whether they carried endophytic fungi. The stems of the plants detected with endophytic fungi were cut into small segments over 1 cm in length and then surface sterilized (75% absolute ethanol for 2 min, 1% sodium hypochlorite for 3 min, and rinsed with sterile water three times). After removing surface moisture with sterile filter paper, the stem segments were inoculated onto antibiotic potato dextrose agar (ABPDA) medium and incubated in the dark at 25 °C. Five fungal pathogens were used in this study. B. sorokiniana, F. avenaceum, and A. alternata were isolated from A. inebrians, while D. erythrospila and C. lunata were isolated from Lolium perenne.
The F. sinensis samples were collected from Xiahe County, Gansu Province, and Guide County, Qinghai Province, China; the isolated Epichloë strains were designated F1, F2, F3, and F4. The H. brevisubulatum samples were collected from Linze County and Ganzhou District, Gansu Province, China; the isolated Epichloë strains were designated H1, H2, H3, and H4. The A. inebrians samples were collected from Sunan Yugur Autonomous County and Hezuo City, Gansu Province, China; the isolated Epichloë strains were designated A1, A2, A3, and A4 (Table 1).
2.2. The Inhibition Rate of Grass Pathogenic Fungi by Epichloë Endophytes In Vitro
A mycelial plug (4 mm diameter) of each endophytic fungus was taken from the margins of actively growing colonies and transferred face down in the centre of a 90 mm diameter Petri dish of ABPDA. After the cultures of Epichloë endophytes were grown on ABPDA at 22 ± 1 °C in the dark for four weeks, they were inoculated with grass pathogenic fungi. For each treatment, three biological replicates (endophyte-inoculated dishes) were prepared. For each biological replicate, three mycelial plugs (4 mm in diameter) from a one-week-old culture of the fungal pathogen were evenly spaced and inoculated on the dish, serving as technical replicates. Meanwhile, five different pathogenic fungal colonies (4 mm in diameter) were evenly spaced and inoculated on the edge of a blank ABPDA medium as a blank control. All dishes were sealed with laboratory film (Parafilm, Pechiney Plastic Packaging, Menasha, WI, USA), then put the dish at 22 ± 1 °C in the dark for one week. After one week of observing the growth condition of pathogenic fungi, we measured the pathogenic fungal colony diameter and the distance between the endophytic fungi and pathogenic fungi and calculated the inhibition rate using the formula:
Ir is inhibition rate and Dck is the mean colony diameter of the pathogenic fungus in the control group. Dpa is the mean colony diameter of the pathogenic fungus in the endophyte-treated group.
2.3. Effect of the Culture Supernatant of Endophytic Fungi on Pathogenic Fungi Colony Growth
ABPDA medium (150 mL) was dispensed into a 250 mL Erlenmeyer flask and sterilized. Following sterilization and cooling, two 4 mm mycelial plugs of the endophytic fungus were added aseptically. After sealing with Parafilm, the flasks were incubated in the dark at 22 ± 1 °C with agitation at 140 rpm on an orbital shaker. After four weeks, we transferred the endophytic fungi liquid culture to a 50 mL centrifuge tube and centrifuged it at 3500 rpm. After centrifugation, we took the supernatant and stored it in a 4 °C refrigerator for later use.
Sterilized 90 mm Petri dishes were individually supplemented with 1 mL of the endophytic fungal culture and 20 mL of melted, sterile ABPDA medium. After the medium solidified, a pathogenic fungal mycelial plug (4 mm in diameter) was inoculated in the center of each prepared plate. For each treatment, three biological replicates were prepared, each consisting of three technical replicates (plates). As a blank control, another set of sterilized 90 mm Petri dishes was prepared by adding 1 mL of sterile liquid PDA medium and 20 mL of melted, sterile ABPDA medium. Five different pathogenic fungi were then individually inoculated onto these control plates. All plates were sealed with laboratory film (Parafilm, Bemis Flexible Packaging, Neenah, WI, USA) and incubated in the dark at 22 ± 1 °C for one week. After a week, we measured the colony diameter and calculated the inhibition rate. The formula for calculating the inhibition rate is the same as Formula (1).
2.4. Effects of Epichloë Endophytes on Spore Germination of Grass Pathogenic Fungi
Under aseptic conditions, 5 mL of sterile water was added to each plate colonized by the pathogenic fungus. The mycelial mat was then scraped off using a sterile spreader and filtered through sterile gauze into a pre-prepared Erlenmeyer flask. The resulting suspension was homogenized thoroughly using sterile inoculation loops to ensure complete spore dispersion. A small aliquot was taken using a rubber-tipped dropper to prepare a wet mount for microscopic observation. The spore concentration was adjusted with sterile water until the spore density reached no less than 100 spores per field of view under 400× magnification.
For the experimental treatment, 1 mL of endophytic fungal culture supernatant and 1 mL of pathogenic fungal spore suspension were mixed on a sterile double-concave slide and covered with a coverslip to prepare a wet mount. For the blank control (CK), 1 mL of sterile liquid ABPDA medium was mixed with 1 mL of the same pathogenic fungal spore suspension on a separate slide. Each mixture was gently stirred with a dissecting needle to ensure uniformity. The spore concentration was examined under a microscope at 400× magnification and adjusted to 80–100 spores per field of view. The prepared slides were placed in 11 cm diameter Petri dishes under humid conditions and incubated at 22 ± 1 °C. Spore germination of the grass pathogens was examined every 2 h until the germination rate in the blank control group exceeded 65%. For each treatment, three biological replicates were examined, with three fields of view counted per replicate as technical replicates, and germinated spores were counted using a hemocytometer. The inhibition rate was calculated using the following formula:
Ir’ is the inhibition rate, Nck is the mean number of germinated spores in the control group, and Npa is the mean number of germinated spores in the treatment group.
2.5. Determination of Ergot Alkaloids
The concentrations of ergonovine and ergine were determined by HPLC analysis following a modified method adapted from Zhang et al. [50] and Liu [51]. Briefly, 500 μL of endophytic fungal culture supernatant was mixed with 1 mL of 20% acetic acid in a 2 mL centrifuge tube. The mixture was sonicated for 5 min, vortexed for 2 min, and then centrifuged at 1000× g for 5 min. A 0.5 mL aliquot of the resulting supernatant was loaded onto a PCX column pre-activated with 2 mL of methanol. The column was sequentially rinsed with 2 mL of purified water and eluted with 1 mL of a 95% methanol solution containing 5% ammonia. The eluate was collected, and a 0.25 mL portion was passed through a 0.22 μm organic phase filter into a 1.5 mL amber vial. All prepared samples were stored in the dark at 4 °C until analysis. HPLC analysis was performed on an Agilent 1100 system equipped with an Agilent C18 column (250 mm × 4.6 mm, 5 μm particle size). The mobile phase consisted of (A) 0.1 mol·L^−1^ ammonium acetate and (B) acetonitrile:0.1 mol·L^−1^ ammonium acetate (3:1, v/v), delivered at a flow rate of 1.0 mL·min^−1^. The elution gradient was programmed as follows: 95% A (0–10 min), linearly decreased to 80% A (10–20 min), further decreased to 50% A (20–30 min), and finally returned to 95% A (30–35 min). Detection was carried out at 312 nm. Alkaloid quantification was achieved by comparing peak areas of 20 μL injections with external standard calibration curves.
2.6. Statistical Analysis
Data are presented as the mean ± standard error of three replications. The normality of the data was tested using the Shapiro–Wilk test, and the data conformed to a normal distribution. Then, the experimental data were statistically analyzed using one-way analysis of variance (ANOVA), followed by Duncan’s post hoc test at the 0.05 level, with SPSS statistical software (Version 27.0, Chicago, IL, USA). An RDA (redundancy analysis) was conducted using the Canoco software program (Version 5.02 trial, Windows release) to analyze the relationship between the concentration of ergonovine and ergine alkaloids in liquid culture medium by Epichloë endophytes and the inhibition rate of the fungal pathogen. All data were centered and standardized prior to analysis. The significance of the RDA axes was tested using Monte Carlo permutation tests as implemented in the software, and the overall model was significant.
3. Results
3.1. Effect of the Colony Growth of Grass Pathogenic Fungi in the Culture Medium Inoculated by Epichloë Endophytes
Compared with the controls, all strains of Epichloë endophytes significantly (p < 0.05) inhibited the colony growth of D. erythrospila, B. sorokiniana, C. lunata, F. avenaceum, and A. alternata (Table 2). Strain A1, isolated from A. inebrians, exhibited the highest inhibition rate against B. sorokiniana (73.24%), while strain H2 from H. brevisubulatum showed the lowest inhibition rate against F. avenaceum (8.41%).
All five fungal pathogens were inhibited to varying degrees by the Epichloë endophytes, with significant differences in inhibitory effects among endophytic strains against the same pathogen (p < 0.05). Notably, strain A1 exhibited significantly stronger inhibition than the other tested strains against most pathogens, particularly D. erythrospila, B. sorokiniana, C. lunata and F. avenaceum. In contrast, strain H3 showed the highest inhibitory activity specifically against A. alternata.
The inhibitory effect of a given Epichloë strain also varied significantly across different pathogens (p < 0.05). Overall, B. sorokiniana was the most sensitive pathogen, with the majority of endophytic strains exerting strong inhibitory effects against it. In contrast, the other four pathogens displayed strain-specific susceptibility: F. avenaceum, C. lunata, A. alternata and D. erythrospila were most strongly inhibited by strains A1, H1, H3 and H4, respectively.
3.2. Effect of the Growth of Grass Pathogenic Fungi Under Liquid Culture Medium by Epichloë Endophytes
3.2.1. Effect of the Colony Growth of Grass Pathogenic Fungi Under Liquid Culture Medium by Epichloë Endophytes
Compared with the controls, all liquid culture media of Epichloë endophytes significantly (p < 0.05) inhibited the colony growth of D. erythrospila, B. sorokiniana, C. lunata, F. avenaceum, and A. alternata (Table 3). Strain A1, isolated from A. inebrians, showed the most significant inhibitory effect on the colony growth of B. sorokiniana, with an inhibition rate of 74.68%. In contrast, strain H2 from H. brevisubulatum exhibited the weakest inhibition against F. avenaceum (9.83%).
All five fungal pathogens were inhibited to varying degrees by the liquid culture medium of Epichloë endophytes, and significant differences in antifungal activity were detected among endophytic strains (p < 0.05). Overall, strain A1 displayed significantly stronger inhibitory effects than most other strains against D. erythrospila, B. sorokiniana, and C. lunata. Strain A2 also showed high inhibitory activity against F. avenaceum, while strain F2 exhibited strong inhibition against D. erythrospila and A. alternata. In addition, strain F4 was among the most effective isolates against A. alternata.
The inhibitory effects of the same Epichloë endophytic strain varied significantly across different grass pathogenic fungi. Strains F1, F2, F3, F4, H1, H2, H3, and H4 demonstrated significantly higher inhibition rates against D. erythrospila than against the other four pathogenic fungi (p < 0.05). Meanwhile, strains A1, A2, A3, and A4 showed the most pronounced inhibition against B. sorokiniana.
3.2.2. Effect of the Fungal Pathogen Spore Germination Under Liquid Culture Medium by Epichloë Endophytes
Compared with the controls, all liquid culture media of Epichloë endophytes significantly (p < 0.05) inhibited spore germination of D. erythrospila, B. sorokiniana, C. lunata, F. avenaceum, and A. alternata (Table 4). Strain F2, isolated from F. sinensis, exhibited the highest inhibition rate against B. sorokiniana spore germination (87.73%), whereas strain H3 from H. brevisubulatum showed the weakest inhibition against F. avenaceum (7.89%).
All five fungal pathogens showed varying degrees of spore germination inhibition by Epichloë endophytes. Significant differences in inhibitory effects were observed among different endophytic strains against the same pathogen (p < 0.05). Overall, strain F3 exhibited the strongest inhibitory effect on D. erythrospila spore germination, outperforming other tested strains. For B. sorokiniana and C. lunata, strain F2 was the most effective inhibitor. Meanwhile, strain A3 showed the highest inhibitory activity against F. avenaceum. In addition, strains F2 and F3 displayed significantly stronger inhibition of A. alternata spore germination. Their inhibitory effects were notably higher than those of other strains.
The inhibitory effects of the same Epichloë endophytic strain varied significantly across different grass pathogenic fungi. All 12 strains exhibited the strongest inhibitory effect on the spore germination of B. sorokiniana, while showing the weakest inhibition against F. avenaceum.
3.2.3. Relationship Between the Concentration of Ergonovine and Ergine Alkaloids in Liquid Culture Medium by Epichloë Endophytes and the Inhibition Rate of the Fungal Pathogen
The first two RDA axes explained 68.02% and 4.96% of the total variance, respectively, cumulatively accounting for 72.98% of the total variation (Figure 1). The concentrations of two ergot alkaloids (ergonovine and ergine) in the liquid culture medium of Epichloë endophytes showed a positive correlation with the inhibition rates against five pathogenic fungi (D. erythrospila, A. alternata, C. lunata, B. sorokiniana, and F. avenaceum), but their inhibitory effects varied among different pathogens. Specifically, compared to ergine, ergonovine exhibited a stronger correlation with the inhibition rates of D. erythrospila and A. alternata, and its correlation with the inhibition rates of B. sorokiniana and C. lunata was also significantly stronger than that of ergine. In contrast, ergine showed a stronger correlation with the inhibition rate of F. avenaceum. Among the endophytic fungal strains, strains A1 and A2 had the highest ergonovine concentration and simultaneously displayed the strongest inhibitory effects on B. sorokiniana and C. lunata. In contrast, strains F4, H2, H3, and H4 had relatively low concentrations of both alkaloids and generally weak inhibitory activity against all five pathogenic fungi.
The result of RDA analysis among the concentration of alkaloids in liquid culture medium by Epichloë endophytes and the inhibition rate of the fungal pathogen. Note: BY indicated Drechslera erythrospila; BZ indicated Alternaria alternata; XYWB indicated Curvularia lunata; LRB indicated Bipolaris sorokiniana; GLB indicated Fusarium avenaceum.
4. Discussion
The mechanisms by which endophytic fungi enhance disease resistance in host grasses may involve the direct inhibition of spore germination [43,52], the enhancement of host plant resistance, and the production of antifungal compounds by the Epichloë endophytic symbiont [53,54]. Alternatively, endophytic infection leads to the formation of fungal mycelial networks or hyphal sheaths within host plant tissues. These structures occupy specific ecological niches in grass tissues and may trigger rejection responses from pathogenic fungi [55]. The antifungal activities of 12 Epichloë strains isolated from three grass species were evaluated against five pathogenic fungi using dual culture and liquid culture assays, with impacts on colony growth and spore germination assessed. Consistent with the hypothesis proposed in Section 1, all 12 Epichloë endophytes exhibited varying degrees of inhibition on both colony growth and spore germination of the pathogenic fungi. A significant positive correlation was observed between the inhibition rates against the five pathogenic fungi and the alkaloid concentrations in the liquid culture media of the different Epichloë endophyte strains. Different Epichloë strains exhibit varying alkaloid concentrations in the liquid culture medium, with corresponding differences in their inhibitory effects on pathogenic fungi.
This study indicates that strains A1 and A2 produced the highest ergonovine concentrations and simultaneously showed the strongest inhibitory effects against B. sorokiniana and C. lunata. This is consistent with the previous research by Sun et al. [24], who reported the presence of ergonovine and ergine in endophyte-infected A. inebrians, while these alkaloids were absent in endophyte-free plants. Their study also demonstrated that extracts from endophyte-infected plants exhibited higher inhibition of pathogenic fungal growth, and that alkaloid concentration was positively correlated with the inhibition of spore germination. Therefore, ergonovine and ergine alkaloids produced by Epichloë endophytes may contribute to the enhancement of disease resistance observed in their host grasses. It is important to note, however, that the inhibitory effect of endophytic fungi on plant pathogenic fungi is not solely dependent on alkaloids. Various non-alkaloid secondary metabolites have also been implicated, including sesquiterpenes [54], phenolic glycerides [56], hydroxyl unsaturated fatty acids [57], aromatic sterols [58], indole derivatives (indole-3-acetic acid (IAA) and indole-3-ethanol), diacetamides [54], as well as volatile compounds such as Chokol K and methyl esters [59]. Different Epichloë species associated with different grass hosts produce distinct metabolite profiles, which in turn lead to varying inhibitory effects on pathogenic fungal growth [60].
The present study revealed that Epichloë strains derived from different host grass species differed in their inhibitory effects on pathogen colony growth and spore germination. In terms of spore germination inhibition, strains isolated from F. sinensis generally exhibited stronger antifungal activity than those from A. inebrians and H. brevisubulatum. This host-dependent variation in inhibitory activity is consistent with the findings of Xie et al. [61], who also reported that the antifungal potential of Epichloë endophytes differed significantly according to their original host species. Furthermore, Yang et al. [41] compared the antifungal activity of acetone extracts from endophyte-infected and endophyte-free host grasses against four fungi: A. alternata, B. sorokiniana, F. avenaceum and Trichoderma viride. They found that extracts from infected plants showed significantly stronger inhibition at all tested concentrations (p < 0.05), and markedly suppressed mycelial growth, conidial germination, and germ tube elongation. These results clearly demonstrated that endophytic infection significantly enhances the antifungal activity of host grass acetone extracts. Previous studies have indicated that the inhibitory effects of endophytic fungi on pathogens are influenced by endophyte genotype [60], which is consistent with the results of the present study.
Different Epichloë strains isolated from different geographic sites of the same species showed significant differences in their inhibitory rates of pathogenic fungi. The inhibition rates of colony growth of D. erythrospila and C. lunata by the liquid culture media of strains A1 and A2, isolated from A. inebrians in Sunan, were significantly different (p < 0.05) compared with those of strains A3 and A4, isolated from A. inebrians in Hezuo. These two sampling sites are approximately 510 km apart. Furthermore, such inhibitory variation also exists among different Epichloë strains. Similarly, Liu (2019) observed that strain XH from Xiahe and strain PA from Ping’an, both isolated from F. sinensis, displayed significantly distinct inhibitory activity against A. alternata [51]. This phenomenon may be attributed to the diverse environmental pressures that shape the host–endophyte symbiosis. Distinct habitats can modulate the physiological and metabolic profiles of Epichloë endophytes, thereby altering the production, composition, and yield of antifungal metabolites synthesized by the symbiotic system. These metabolic differences ultimately lead to the observed variation in inhibitory activity against pathogenic fungi [62]. However, further in-depth studies are needed to understand the specific inhibiting molecules.
There are some differences in the inhibition activity of pathogenic fungi between different strains of Epichloë endophytes isolated from the same geographic sites of the same species of plant. For example, strain H1 inhibited the colony growth of the five pathogenic fungi in the following order of decreasing inhibition rates: C. lunata, B. sorokiniana, A. alternata, D. erythrospila, and F. avenaceum, but the order for strain H2 was B. sorokiniana, C. lunata, D. erythrospila, A. alternata, and F. avenaceum. This result is similar to Li’s findings on the interaction between E. gansuensis and pathogens [46]. In their study, when tested against four pathogenic fungi, strain L112 inhibited colony growth in the following order of decreasing sensitivity: B. sorokiniana, C. lunata, F. avenaceum and A. alternata, but the order for L113 was B. sorokiniana, F. avenaceum, A. alternata and C. lunata. L112 and L113 are E. gansuensis, which were isolated from the same geographic populations of A. inebrians, but their inhibitory effects on pathogenic fungi are significantly different. Niones and Takemoto also reached similar results. Although both E364 and E365 were isolated from Festuca longifolia in Switzerland, only E364 exhibited inhibitory effects against C. graminicola [63]. This differential activity is likely attributable to underlying genetic heterogeneity between the isolates.
The results of this study also showed that endophytic fungi exhibit varying inhibitory effects on pathogenic fungi under different culture conditions. Taking strain F4 as an example, its inhibitory activity against the five tested pathogenic fungi differed between solid and liquid culture media. Such differences in inhibitory activity under different culture conditions were also observed in other endophyte strains. For instance, Ma and Nan [2] reported that strain N-A1 isolated from L. perenne also showed distinct inhibitory effects on pathogen colony growth between solid and liquid culture assays. This condition-dependent variation in inhibitory activity is likely due to different inhibitory mechanisms under distinct culture conditions [42]. Previous studies have found that the main mechanism is hyperparasite or niche competitive when the colony of Epichloë endophytes and pathogenic fungi has a direct action on the culture medium. The Epichloë endophytes invade pathogen hyphae and enzymatically hydrolyze the pathogenic fungus cells [61], or compete against pathogenic fungus nutrients and niche space, and achieve antagonism between pathogenic fungi [30]. In liquid culture, however, Epichloë endophytes mainly inhibit pathogenic fungi by secreting active compounds such as indole derivatives, sesquiterpenes, and alkaloids [54,64,65]. Furthermore, whether different culture media and conditions affect the types and quantities of antifungal metabolites produced by Epichloë endophytes requires further investigation.
The results indicate that all 12 strains exhibited significant inhibition of pathogenic fungal growth. This finding, however, contrasts with that of Li et al., who reported that both N. lolii and N. coenophialum promoted the growth of C. lunata and A. alternata, and that strains NgL121, NgL141, NgL232, and NgS1a1 enhanced the sporulation of Fusarium acuminatum [46]. Generally, the variations in the inhibitory effects of endophytic fungi result from the combined action of multiple factors, including pathogen strains, grass genotypes or varieties, growth conditions, endophyte status, and so on [42,44,66]. Although some endophytes produced antifungal substances in vitro [8], they may not be able to produce these compounds in sufficient quantities in the grass to protect it from fungal diseases [52,56]. Alternatively, the inhibitory substances produced in vitro may differ from those synthesized in planta [42]. After all, external culture conditions in vitro are quite different from the native growth conditions of endophyte fungi. Furthermore, regarding secondary metabolites, which are crucial for symbiotic protection, the concentrations produced by the endophyte alone are far lower than those of compounds such as alkaloids generated by endophyte–grass symbiosis [60]. Therefore, future research should focus on identifying the specific antagonistic compounds produced by these Epichloë endophytes and elucidating their mechanisms of action. Such efforts will support the potential of Epichloë endophytes as promising biocontrol agents for the sustainable management of grass fungal diseases.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Fernando K. Reddy P. Hettiarachchige I.K. Spangenberg G.C. Rochfort S.J. Guthridge K.M. Novel antifungal activity of Lolium-associated Epichloë endophytes Microorganisms 2020895510.3390/microorganisms 806095532599897 PMC 7355949 · doi ↗ · pubmed ↗
- 2Ma M.Z. Nan Z.B. Effects of fungal endophytes from perennial ryegrass on the growth of plant pathogens Pratacult. Sci.201128962968
- 3Wang X.Y. Zhou Y. Ren A.Z. Gao Y.B. Effect of endophyte infection on fungal disease resistance of Leymus chinensis Acta Ecol. Sin.2014346789679610.5846/stxb 201303080377 · doi ↗
- 4Zhang Y.W. Nan Z.B. Xin X.P. Response of plant fungal diseases to beef cattle grazing intensity in Hulunber grassland Plant Dis.20201042905291310.1094/PDIS-03-20-0683-RE 32915707 · doi ↗ · pubmed ↗
- 5Li F. Deng J. Guo Y.E. Gao P. Li Y.Z. Duan T.Y. Influence of interactions between grass endophyte and arbuscular mycorrhizal fungi on growth and leaf spot disease of perennial ryegrass J. Plant Prot.20194635236110.13802/j.cnki.zwbhxb.2019.2017180 · doi ↗
- 6Yakti W. Kovács G.M. Franken P. Differential interaction of the dark septate endophyte Cadophora sp. and fungal pathogens in vitro and in planta FEMS Microbiol. Ecol.201995 fiz 16410.1093/femsec/fiz 16431609451 PMC 6864363 · doi ↗ · pubmed ↗
- 7Deng J. Li F. Duan T.Y. Effects of AM fungus and grass endophyte on the infection of Lolium perenne by the pathogen Bipolaris sorokinianum in a greenhouse Acta Pratacult. Sin.20192810311310.11686/cyxb 2019104 · doi ↗
- 8Fernando K. Reddy P. Spangenberg G.C. Rochfort S.J. Guthridge K.M. Metabolic potential of Epichloë endophytes for host grass fungal disease resistance Microorganisms 2022106410.3390/microorganisms 10010064 PMC 878156835056512 · doi ↗ · pubmed ↗