Phenotypic Antimicrobial Resistance Profiles and Provisional Epidemiological Cut-Off Values of Edwardsiella anguillarum Isolated from Farmed Nile Tilapia (Oreochromis niloticus) in Brazil, with Exploratory Data on Edwardsiella tarda
Natália Amoroso Ferrari, Vittória Cueva Segura da Silva, Pamela Giovana Turini, Julia Faria de Souza, Raffaella Menegueti Mainardi, Mayza Brandão da Silva, Alene Santos Souza, Gabriel Diogo Guimarães, Maisa Fabiana Menck-Costa, Marco Rozas-Serri, Mariene Miyoko Natori

TL;DR
This study examines antibiotic resistance in Edwardsiella bacteria from farmed Nile tilapia in Brazil and proposes new thresholds to monitor resistance in aquaculture.
Contribution
The paper introduces provisional epidemiological cut-off values for Edwardsiella anguillarum and reports resistance patterns in aquaculture settings.
Findings
Most Edwardsiella anguillarum isolates remained susceptible to several antibiotic classes.
Non-wild type and multidrug-resistant profiles were detected with a MAR index of 0.68.
Local, species-specific pECVs are recommended for resistance surveillance in aquaculture.
Abstract
Antimicrobial resistance in bacteria associated with aquaculture, such as Edwardsiella spp., represents an emerging challenge because of their relevance to fish health and their potential impact on animal, environmental, and human health. In this study, we primarily investigated the antimicrobial susceptibility profiles of Edwardsiella anguillarum isolated from farmed Nile tilapia (Oreochromis niloticus) in Brazil. Based on our findings, herein, we propose provisional local epidemiological cut-off values (pECVs) using the normalized resistance interpretation method, with data for Edwardsiella tarda included as an exploratory context. Fifty isolates (31 E. anguillarum and 19 E. tarda) collected between 2017 and 2025 were tested against 28 antibacterial agents using the disk diffusion method. Based on the pECVs, isolates were classified as wild type (WT) or non-WT (NWT), and the multiple…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
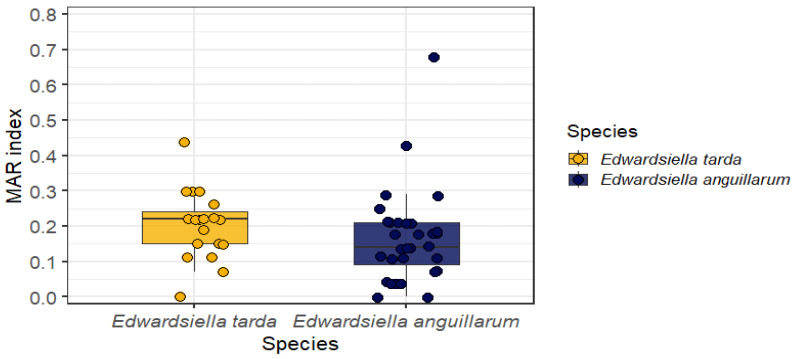 Figure 1
Figure 1- —National Council for Scientific and Technological Development
- —Coordination of Superior Level Staff Improvement (CAPES), Brazil
- —CNPq/MCTI/CT-Saúde
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAquaculture disease management and microbiota · Pharmaceutical and Antibiotic Environmental Impacts · Aquaculture Nutrition and Growth
1. Introduction
Edwardsiellosis is a septicemic disease that affects a wide range of aquatic hosts, resulting in substantial economic losses and posing a threat to aquaculture systems worldwide [1]. The disease is caused by species of the genus Edwardsiella, four of which affect fish: E. tarda, E. anguillarum, E. ictaluri, and E. piscicida [2]. Edwardsiella tarda is most frequently associated with clinical conditions in fish, whereas E. anguillarum has been described as an emerging and highly virulent pathogen, particularly affecting tilapia from tropical regions [2,3,4]. Outbreaks have been associated with high fish mortality rates, reduced growth performance, increased treatment costs, and trade losses, collectively resulting in substantial financial impacts in regions with intensive production. In an outbreak in oscar fish (Astronotus ocellatus), an ornamental species, E. tarda has been reported to cause 100% mortality within 240 h [5]. In another outbreak affecting tilapia in Korea, a strain of E. anguillarum has been reported to cause significant mortality and economic loss [6]. Recent studies have indicated that infections caused by E. anguillarum can lead to acute septicemia and rapid mortality, reinforcing its increasing relevance in warm-water aquaculture [7,8]. Taxonomic revisions have direct implications for antimicrobial susceptibility studies because historical data may reflect heterogeneous species assignments.
These bacteria are gram-negative bacilli that, in addition to compromising animal health and productivity, hold potential public health relevance. Edwardsiella tarda has been linked to human infections, including bacteremia and septicemia, which are often associated with the consumption of raw fish or environmental exposure to infections [9]. The zoonotic potential of E. tarda, combined with its widespread distribution in the intestinal microbiota of fish and other aquatic organisms [1], highlights the need for integrated surveillance strategies from a One Health perspective [5], particularly in regions where raw or minimally processed fish are commonly consumed [6,10]. In addition, there is limited knowledge regarding the zoonotic potential of E. anguillarum and the molecules used for its treatment.
In this context, the use of antimicrobials remains a main measure for controlling bacterial infections in fish, including those caused by Edwardsiella spp. [1,10,11]. However, the pressure exerted by these drugs favors the selection of drug- or multidrug-resistant (MDR) strains [1,12]. Furthermore, the indiscriminate or prolonged use of these agents accelerates and intensifies this phenomenon, transcending aquaculture and posing a global public health threat under the One Health Framework [13]. Moreover, Edwardsiella spp. play an important role in the dissemination of resistance genes, often carried by mobile genetic elements, such as plasmids [1]. Classical genes, such as blaTEM (β-lactamase gene) and tetA (tetracycline resistance gene), have been reported in E. tarda, reinforcing its function as a reservoir and vector of resistance to other microorganisms [3,10]. In several countries, only a few molecules have been validated and authorized for use in fish, such as in Brazil, where only oxytetracycline and florfenicol have been officially approved [14]. Based on routine diagnostic cases in Brazil (unpublished data), reduced therapeutic responses to these compounds have been occasionally observed in infections caused by Edwardsiella spp., a scenario also suggested by in vitro findings reported in the literature [10].
Additionally, species within the genus Edwardsiella exhibit differences in ecological distribution and host predilection [1]. For example, E. anguillarum has been isolated from Nile tilapia and other species, such as seabream, eels, and milkfish [4,7], whereas E. tarda has an even broader host range, including freshwater and marine fish [10,15,16]. These species-specific associations suggest that distinct lineages circulate in different ecological niches, potentially impacting their antimicrobial resistance profiles.
Despite the growing number of reports on edwardsiellosis, significant methodological limitations remain. There are no official interpretive criteria, recommended by the CLSI (Clinical and Laboratory Standards Institute)/ EUCAST (European Committee on Antimicrobial Susceptibility Testing), specific to Edwardsiella spp., and the absence of standardized epidemiological cut-off values (ECVs) complicates the detection of early resistance and comparisons across studies [10]. Furthermore, although some studies have generated ECVs or susceptibility data for specific Edwardsiella spp., direct comparative evaluations of E. tarda and E. anguillarum, two species with increasing relevance in tropical aquaculture, are scarce [10]. This gap is particularly relevant because these species exhibit differences in their host preferences and pathogenicity profiles in cultured fish [7] and in the One Health context.
Considering the limited availability of isolates from each species in most routine aquaculture diagnostics, including in the present study, generating large datasets suitable for formal ECV determination is often not feasible. Thus, preliminary datasets are valuable as they provide the first comparative signals needed to guide future surveillance, refine hypotheses, and support the gradual construction of robust reference values.
Therefore, continuous monitoring of antimicrobial susceptibility in fish-associated bacteria is essential for elucidating the emergence and spread of resistance. In this scenario, even preliminary comparative data may make meaningful contributions by revealing the early differences between bacterial pathogen species.
In this study, we aimed to characterize the antimicrobial susceptibility profiles of E. anguillarum isolated from Nile tilapia and to propose provisional local ECVs (pECVs) using phenotypic data. The results obtained for E. tarda are presented as exploratory and contextual information rather than for a formal interspecies comparison.
2. Materials and Methods
2.1. Origin and Selection of Isolates
The Edwardsiella spp. isolates analyzed in this study were obtained from farmed Nile tilapia and submitted to the Fish Bacteriology Laboratory of the State University of Londrina (LABBEP–UEL) for microbiological diagnosis between 2017 and 2025.
After necropsy, the eye, brain, kidney, liver, and spleen fragments were aseptically collected and inoculated onto Mueller–Hinton blood agar (MHBA), which contains Mueller–Hinton agar (MHA; Himedia, Mumbai, India) supplemented with 5% defibrinated sheep blood. The plates were incubated at 29 ± 1 °C for 24–48 h for bacterial isolation. Initially, the colonies were evaluated for morphology and Gram staining, in addition to biochemical tests for genus-level identification.
As it is not possible to differentiate Edwardsiella species using only phenotypic tests, DNA was extracted using the DNeasy Blood and Tissue Kit (QIAGEN, Hilden, Germany), following the manufacturer’s instructions, for the molecular identification of species using multiplex polymerase chain reaction, as proposed by Da Costa et al. [17]. After identification, the isolates were preserved at −80 °C in a solution containing brain–heart infusion broth (Himedia, Mumbai, India) and 20% glycerol.
In this study, 19 isolates of E. tarda and 31 of E. anguillarum were collected from Nile tilapia (Oreochromis niloticus) originating from six Brazilian states, as shown in Table 1.
2.2. Antimicrobial Susceptibility Tests
The selected isolates were subcultured from frozen stocks onto MHBA and subjected to the disk diffusion method on MHA following the recommendations of the CLSI guideline VET03 for bacteria isolated from aquatic animals [18]. After a 24 h incubation at 28 °C, a bacterial inoculum was prepared and standardized to 0.5 McFarland (approximately 1.5 × 10^8^ CFU [colony-forming units]/mL) and subsequently seeded onto MHA plates. A total of 28 antibacterial agents were evaluated (piperacillin + tazobactam, aztreonam, meropenem, ceftazidime, amoxicillin, sulfazotrim, norfloxacin, tobramycin, ampicillin, cefazolin, imipenem, levofloxacin, cefoxitin, amoxicillin + clavulanic acid, ceftiofur, cefepime, ceftriaxone, cefotaxime, cefuroxime, florfenicol, amikacin, gentamicin, cephalexin, ciprofloxacin, tetracycline, enrofloxacin, streptomycin, and marbofloxacin). The antimicrobial panel was primarily composed of agents recommended for the susceptibility testing of Enterobacterales, ensuring coverage of the main antimicrobial classes relevant to this group. In addition, antimicrobial agents routinely used in the diagnostic workflow for fish bacterial isolates at the LABBEP–UEL were included because of their relevance in treating bacterial diseases in fish.
The inhibition zone diameters were measured in millimeters. The phenotypic production of extended-spectrum β-lactamases (ESBLs) and ampicillin cephalosporinase β-lactamases (ACBLs) was also assessed. For this purpose, antibiotic discs were strategically arranged on MHA plates to facilitate the detection of the characteristic inhibition patterns associated with these resistance mechanisms. Escherichia coli ATCC 25922 was included as the reference strain to monitor assay performance and verify disk viability. Moreover, internal laboratory controls previously characterized as positive were used to validate the phenotypic identification of ESBL and ACBL production.
2.3. Determination of ECVs and Multiple Antibiotic Resistance (MAR) Index
Considering that the CLSI and BrCAST (Brazilian Committee on Antimicrobial Susceptibility Testing)/ EUCAST interpretive criteria extrapolate values that have not been validated for fish pathogens, the isolates were not classified using these systems.
Instead, pECVs were determined using the normalized resistance interpretation (NRI) method available at https://www.bioscand.se/nri/ (accessed on 13 October 2025). Based on the pECVs, isolates were classified as wild type (WT) or non-WT (NWT) [19]. Considering the sample size, the pECVs calculated for E. anguillarum (n = 31) are presented as local and provisional reference values, whereas those estimated for E. tarda (n = 19) are presented as preliminary and exploratory. According to the authors’ recommendations, when disc diffusion datasets include fewer than 50 observations obtained at 28 °C, the standard deviation (SD) upper limit recommended for assays performed at 22 °C should be used for the analysis. As our dataset met these criteria (<50 observations at 28 °C), we adopted the corresponding threshold and considered an upper SD limit of <6.49 mm. NWT isolates were interpreted as resistant.
For each isolate, the MAR index was calculated as the ratio of the number of antibacterial agents to which the strain was resistant (NWT) to the total number of antibacterial agents used. The MAR index is a tool used to indicate the level of antibiotic pressure in a given source, with a threshold of 0.2 commonly used to identify environments at a high risk of antibiotic exposure [20].
For E. tarda, no pECV was established for tetracycline, owing to numerous small inhibition zones that prevented analysis; thus, 27 antibacterial agents were considered for the MAR index calculation. Isolates exhibiting resistance to three or more classes of antibacterial agents were classified as MDR [21].
2.4. Data Analyses
The results were compiled in Microsoft Excel and analyzed using RStudio v.2025.05.1 + 513. The English language of the manuscript was reviewed and polished using the AI language model ChatGPT (version GPT-5.2, OpenAI) to enhance clarity, grammar, and overall readability.
3. Results
The results are presented based on antimicrobial class to facilitate the interpretation of phenotypic susceptibility patterns between species. Phenotypic antimicrobial susceptibility testing revealed heterogeneous inhibition zone diameters among E. tarda (n = 19) and E. anguillarum (n = 31) isolates obtained from farmed Nile tilapia (O. niloticus) (Table 2 and Table 3; Supplementary Figures S1–S28). The quality control strain Escherichia coli ATCC 25922 yielded inhibition zone diameters within the expected reference ranges for all antimicrobial agents tested, validating the disk diffusion assays (Supplementary Table S1). The internal quality controls used for the phenotypic screening of ESBL and ACBL production also performed as expected, confirming the technical reliability of these procedures (Supplementary Figure S29).
Based on the pECV classification, both species showed high proportions of WT isolates for several antimicrobial classes. In the piperacillin + tazobactam treatment, 95% of E. tarda and 81% of E. anguillarum isolates were classified as WT. Third- and fourth-generation cephalosporins also exhibited high WT proportions, including ceftazidime (95% WT in E. tarda and 94% WT in E. anguillarum), cefepime (100% and 94% WT, respectively), and cefotaxime (89% and 81% WT) (Table 2).
Carbapenems showed high WT proportions in both species. In the imipenem treatment, 89 and 100% of E. tarda and E. anguillarum isolates were classified as WT, respectively, whereas in the meropenem treatment, 95 and 90% of the corresponding isolates were classified as WT, respectively. Aminoglycosides also showed predominantly WT profiles, particularly tobramycin (100% WT in E. tarda and 97% WT in E. anguillarum), amikacin (95% and 100% WT, respectively), and gentamicin (84% and 100% WT, respectively).
In contrast, an increased proportion of NWT isolates was observed for several antimicrobial agents. Among penicillins, in the amoxicillin and ampicillin treatments, 42% of E. tarda isolates were classified as NWT for each compound, whereas 3% of E. anguillarum isolates were classified as NWT. Amoxicillin and clavulanic acid elevated NWT proportions in both species, reaching 68% in E. tarda and 58% in E. anguillarum. When grouped by antimicrobial class, penicillins and fluoroquinolones presented higher NWT proportions than those of carbapenems, aminoglycosides, and advanced-generation cephalosporins.
For cephalosporins, E. anguillarum exhibited higher pECVs for ceftazidime, cefotaxime, cefuroxime, and cefazolin than did E. tarda. Ceftriaxone and cephalexin exhibited high WT proportions in E. tarda (89 and 95%, respectively), whereas in the cephalexin treatment, 48% of E. anguillarum isolates were classified as NWT.
Fluoroquinolone susceptibility profiles differed between the species. For E. tarda, norfloxacin and levofloxacin classified 89% and 58% of the isolates as NWT, respectively, whereas ciprofloxacin and marbofloxacin classified 95% and 100% of the isolates as WT, respectively (Table 3). In E. anguillarum, NWT proportions were observed for ciprofloxacin (42%), enrofloxacin (29%), norfloxacin (23%), and marbofloxacin (16%), whereas 90% of the isolates were classified as WT for levofloxacin.
Florfenicol classified 53% of E. tarda and 52% of E. anguillarum isolates as WT. Provisional pECVs for tetracycline could not be established for E. tarda, whereas E. anguillarum showed a distribution that allowed the classification of 52% of the isolates as WT and 48% as NWT.
Multidrug resistance, defined as resistance to three or more antimicrobial classes based on the pECV classification, was detected in 15 of the 19 E. tarda isolates (78.95%) and 22 of the 31 E. anguillarum isolates (70.97%). Non-MDR profiles were observed in 4 E. tarda isolates (21.05%) and 9 E. anguillarum isolates (29.03%).
The MAR index ranged from 0.00 to 0.44 in E. tarda (mean = 0.20) and from 0.00 to 0.68 in E. anguillarum (mean = 0.19). MAR index values ≥ 0.20 were observed in 11 of 19 E. tarda isolates (57.89%) and in 12 of 31 E. anguillarum isolates (38.71%). The highest MAR index value (0.68) was observed for E. anguillarum isolate BEP228. The distribution and individual variability of the MAR indices are shown in Figure 1.
Phenotypic screening for β-lactamase activity did not detect ESBL phenotypes in either species. One E. anguillarum isolate (BEP228) showed a phenotypic profile suggestive of ACBL production, which was reproducible in independent assays (Supplementary Figure S30).
4. Discussion
The present study was designed to establish pECVs for E. anguillarum isolated from Nile tilapia (O. niloticus) farmed in Brazil using the NRI approach. Considering the sample size obtained for E. anguillarum, the proposed pECVs provide a consistent framework for local epidemiological surveillance, allowing for discrimination between WT and NWT subpopulations. In contrast, the data generated for E. tarda should be interpreted as preliminary and exploratory, serving mainly to contextualize antibiotic resistance patterns within the genus rather than to support direct interspecies comparisons.
The importance of defining local pECVs is evident when considering the substantial heterogeneity in antimicrobial susceptibility profiles reported for Edwardsiella spp. across different geographic regions, host species, and production systems. Rahmawaty [4] demonstrated wide phenotypic and genotypic variability among Edwardsiella isolates from Taiwan, including both E. tarda and E. anguillarum, highlighting that resistance patterns are strongly impacted by local antimicrobial usage and ecological conditions. Similarly, Rocha [10] showed that pECVs derived from E. tarda varied according to fish species and farming systems in Brazil, reinforcing the need for species-specific epidemiological benchmarks rather than generalized cut-off values.
All isolates analyzed in the present study originated from Nile tilapia on commercial farms, a species typically produced under intensive farming conditions characterized by high stocking densities and frequent therapeutic interventions [1,7,10]. Such farming conditions favor antimicrobial exposure and sustained selective pressure, contributing to the emergence and maintenance of resistant phenotypes [7,10,13]. Thus, the pECVs proposed for E. anguillarum likely reflect the resistance distributions associated with intensive tilapia farming systems and should not be directly extrapolated to native fish species or low-input fish production systems [10,15,22]. This interpretation is supported by Reis et al. [15], who reported lower MAR indices and distinct resistance profiles in E. tarda isolated from tambaqui (Colossoma macropomum), a species generally reared under relatively less intensive conditions.
Although no unique intrinsic resistance patterns have been formally described for Edwardsiella spp., their profile is consistent with that of other Enterobacterales, characterized by natural resistance to narrow-spectrum penicillins and several non-β-lactam classes, while maintaining baseline susceptibility to most β-lactams [1]. In the members of this family, this basal resistance is primarily attributed to the structural features of β-lactamase genes, such as reduced outer membrane permeability, and their presence, with no basal chromosomal activity [9]. According to Nantongo [23], resistance to drugs such as oxacillin and penicillin in E. tarda is a typical family trait, often resulting from porin channels that restrict the entry of hydrophobic or large-molecular-weight molecules. Therefore, clinically significant β-lactam resistance observed in this genus likely reflects acquired mechanisms, such as the production of plasmid-mediated β-lactamases, e.g., blaTEM and blaCTX-M enzymes, coupled with alterations in membrane permeability rather than a uniquely elevated intrinsic resistance [9].
From a mechanistic perspective, the conserved susceptibility of the isolates to carbapenems, advanced-generation cephalosporins, aminoglycosides, and sulfamethoxazole/ trimethoprim is consistent with the modes of action of these antimicrobials [24]. Carbapenems and advanced cephalosporins inhibit bacterial cell-wall synthesis through high-affinity binding to penicillin-binding proteins and exhibit enhanced stability against most β-lactamases, whereas aminoglycosides target the 30S ribosomal subunit to inhibit protein synthesis [24]. Furthermore, the sulfamethoxazole/ trimethoprim combination acts as a folate pathway inhibitor, interfering with the synthesis of essential precursors for bacterial nucleic acid production [13,21]. Moreover, the observation that these isolates remained susceptible suggests that in this Brazilian aquaculture setting, broad-spectrum resistance mechanisms remain uncommon among Edwardsiella populations [10]. This interpretation is supported by recent studies in Brazil that reported a low frequency of -lactamase-associated resistance genes in Edwardsiella spp., in contrast to the high prevalence of ESBL-producing strains reported in other regions, such as Egypt and Asia [10].
In contrast, higher proportions of NWT isolates than those of WT isolates were observed for penicillins, fluoroquinolones, florfenicol, and tetracycline. Resistance to penicillins may be attributed to the basal chromosomal β-lactamase activity, reduced outer membrane permeability, or the presence of inhibitor-susceptible enzymes, as suggested by their variable responses to amoxicillin/ clavulanic acid [25]. The absence of ESBL phenotypes among all isolates, confirmed by phenotypic detection, is epidemiologically relevant and contrasts with reports from Egypt and parts of Asia, where the genotypic detection of bla_CTX-M and bla_TEM in E. tarda exceeded 80% of the isolates [1]. These findings suggest that, although resistance to penicillins and some cephalosporins is present, ESBLs are not yet widespread among Brazilian Edwardsiella isolates [10].
Therefore, resistance to cephalosporins such as ceftriaxone and aztreonam, even in the absence of ESBL production, may be associated with alternative mechanisms. Similar patterns have been described in Edwardsiella spp., including E. tarda, isolated from ornamental fish and humans. In such species, mutations affecting porin proteins (OmpF/OmpC) and overexpression of efflux pumps (AcrAB–TolC) contributed to intermediate resistance levels without detectable ESBL genes [11,25,26]. These resistance mechanisms are compatible with the heterogeneous inhibition zone distributions observed in the present study.
Only one E. anguillarum isolate exhibited a phenotypic profile suggestive of ACBL production. Although this result was reproducible across independent assays, its exclusive phenotypic nature requires cautious interpretation and molecular confirmation to determine the genetic basis in this case. Rocha et al. [10] reported a low frequency of β-lactamase-associated resistance genes in Edwardsiella spp., supporting the view that enzymatic β-lactam resistance remains limited in Brazilian aquaculture; however, its emergence cannot be ruled out under sustained selective pressure.
Fluoroquinolone resistance displayed marked heterogeneity among E. anguillarum isolates, reflecting the diversity of the mechanisms involved in resistance to this antimicrobial class. Fluoroquinolones inhibit DNA gyrase and topoisomerase IV, and resistance may arise through point mutations in quinolone resistance-determining regions, plasmid-mediated quinolone resistance genes (qnrA and qnrS), efflux pump overexpression, or reduced outer-membrane permeability [1,13]. The coexistence of WT and NWT subpopulations observed in the present study suggests continuous selection and dissemination of resistance determinants within tilapia farming systems, as indicated by other studies [3,10].
Resistance to florfenicol and tetracycline deserves particular attention as these are the only antimicrobials officially approved for therapeutic use in fish in Brazil [27]. Both compounds inhibited bacterial protein synthesis by targeting the 50S and 30S ribosomal subunits. Resistance to these classes is commonly mediated by transferable genes such as floR, fexA, and tet variants, which are frequently located on plasmids, transposons, and other mobile genetic elements [1,10]. Therefore, the intermediate WT/NWT distributions observed for E. anguillarum are consistent with the active role of horizontal gene transfer in shaping resistance profiles in aquaculture environments [3].
The MAR index and high frequency of MDR isolates observed in E. anguillarum further indicate sustained selective pressures in intensive tilapia farming systems. Similar MAR thresholds associated with high-risk environments have been reported in other studies involving Edwardsiella spp. [5,10]. Notably, the presence of MDR isolates with relatively low MAR values highlights heterogeneous resistance profiles arising from different combinations of resistant antimicrobial classes rather than from the uniform accumulation of resistance mechanisms.
From a One Health perspective, the implications of these findings extend beyond fish health. Aquaculture environments provide favorable conditions, including high bacterial densities, biofilm formation, and exposure to antimicrobial residues at sub-inhibitory concentrations, for horizontal gene transfer [3]. Edwardsiella is recognized as a core member of the aquatic resistome owing to its abundance and ability to harbor diverse antimicrobial resistance genes (ARGs) on chromosomes and plasmids [3]. Furthermore, it serves as a crucial indicator organism for monitoring the accumulation of ARGs in aquatic habitats, acting as a reservoir for detecting resistance determinants in the environment and transmitting them to the relatively broader microbiome [3]. These genes may be exchanged for opportunistic human pathogens, contributing to the expansion of environmental resistomes and posing substantial risks to animal, environmental, and human health.
These results highlight the dynamic interplay between antimicrobial usage, resistance mechanisms, and environmental factors in aquaculture while emphasizing the importance of integrated phenotypic and molecular surveillance strategies aligned with the One Health principle.
5. Conclusions
In this study, pECVs for E. anguillarum isolated from Nile tilapia (O. niloticus) in Brazil were determined, providing a practical phenotypic framework for distinguishing between WT and NWT populations using NRI. In addition, the data presented herein provide preliminary information on the antibacterial resistance of E. tarda strains and other relevant fish pathogens. Brazilian Edwardsiella lineages appear to be at an intermediate stage of antimicrobial resistance dissemination, with their heterogeneous susceptibility profiles shaped by selective pressure in intensive tilapia farming. Although most isolates remain susceptible to several antimicrobial classes, the presence of NWT and MDR isolates, with MAR indices ≥0.2, observed across regions and years, indicates ongoing selection driven by antimicrobial use. Finally, this study makes available, and subject to comparison, relevant data on antibacterial resistance and local pECVs, and provides preliminary insights into the two major fish pathogens of the genus Edwardsiella.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Algammal A.M. Mabrok M. Ezzat M. Alfifi K.J. Esawy A.M. Elmasry N. El-Tarabili R.M. Prevalence, Antimicrobial Resistance (AMR) Pattern, Virulence Determinant and AMR Genes of Emerging Multi-Drug Resistant Edwardsiella tarda in Nile Tilapia and African Catfish Aquaculture 202254873764310.1016/j.aquaculture.2021.737643 · doi ↗
- 2Katharios P. Kalatzis P.G. Kokkari C. Pavlidis M. Wang Q. Characterization of a Highly Virulent Edwardsiella anguillarum Strain Isolated from Greek Aquaculture, and a Spontaneously Induced Prophage Therein Front. Microbiol.20191014110.3389/fmicb.2019.0014130787917 PMC 6372524 · doi ↗ · pubmed ↗
- 3Leung K.Y. Wang Q. Zheng X. Zhuang M. Yang Z. Shao S. Achmon Y. Siame B.A. Versatile Lifestyles of Edwardsiella: Free-Living, Pathogen, and Core Bacterium of the Aquatic Resistome Virulence 20221351810.1080/21505594.2021.200689034969351 PMC 9794015 · doi ↗ · pubmed ↗
- 4Rahmawaty A. Chen M.Y. Byadgi O.V. Wang P.C. Chen S.C. Phenotypic and Genotypic Analysis of Edwardsiella Isolates from Taiwan Indicates Wide Variation with a Particular Reference to Edwardsiella tarda and Edwardsiella anguillarum J. Fish Dis.2022451659167210.1111/jfd.1368835916068 · doi ↗ · pubmed ↗
- 5Vishnupriya V. Swaminathan T.R. Dharmarathnam A. Sharma S.R.K. Preena P.G. Virulent and Multi-Drug-Resistant Edwardsiella tarda Infection in Oscar Fish: Unveiling the Threat of Mass Mortality and AMR Dissemination Curr. Microbiol.20248117410.1007/s 00284-024-03698-638753164 · doi ↗ · pubmed ↗
- 6Oh W.T. Jun J.W. Kim H.J. Giri S.S. Yun S. Kim S.G. Kim S.W. Kang J.W. Han S.J. Kwon J. Characterization and Pathological Analysis of a Virulent Edwardsiella anguillarum Strain Isolated From Nile Tilapia (Oreochromis niloticus) in Korea Front. Vet. Sci.202071410.3389/fvets.2020.0001432047760 PMC 6997428 · doi ↗ · pubmed ↗
- 7Elgendy M.Y. Sherif A.H. Kenawy A.M. Abdelsalam M. Phenotypic and Molecular Characterization of the Causative Agents of Edwardsiellosis Causing Nile Tilapia (Oreochromis niloticus) Summer Mortalities Microb. Pathog.202216910562010.1016/j.micpath.2022.10562035690232 · doi ↗ · pubmed ↗
- 8Martínez-Lara P. Hernández-López J. Vargas-Albores F. Yepiz-Plascencia G. Gollas-Galván T. Martínez-Porchas M. Montes-Vara R. Coronado-Molina D. First Identification of Pathogenic Edwardsiella anguillarum, Nocardia Asteroides, and Streptococcus agalactiae in Tilapia (Oreochromis niloticus) Farmed in Mexico Microb. Pathog.202520710793410.1016/j.micpath.2025.10793440716469 · doi ↗ · pubmed ↗