First Report of Desmodium styracifolium as a Novel Host for ‘Candidatus Phytoplasma australasiaticum’—Related Strains in China
Yafei Tang, Zhenggang Li, Mengdan Du, Guobing Lan, Lin Yu, Shanwen Ding, Zifu He, Xiaoman She

TL;DR
This paper reports the first case of a phytoplasma infection in the medicinal plant Desmodium styracifolium in China, identifying it as a new host for a specific phytoplasma strain.
Contribution
The study establishes D. styracifolium as a novel host for ‘Candidatus Phytoplasma australasiaticum’ in China.
Findings
Phytoplasma infection was confirmed in D. styracifolium using molecular techniques.
The phytoplasma strain was identified as belonging to the 16SrII-A subgroup of ‘Ca. Phytoplasma australasiaticum’.
This is the first report of this phytoplasma in D. styracifolium in China.
Abstract
Desmodium styracifolium (Osb.) Merr., a member of the Leguminosae family, is an important medicinal plant widely used in traditional Chinese medicine. In September 2024, D. styracifolium plants exhibiting symptoms of little leaf and stunted growth were observed in a field of Zhanjiang, Guangdong province, China. Since the symptoms resembled those associated with phytoplasma infections, total DNA was extracted from the leaves of four symptomatic plants and one healthy plant for molecular identification. Universal primer pairs (P1/P7, R16mF2/mR1) for phytoplasma detection were used to amplify the 16S rDNA fragments (~1.8 kb and ~1.4 kb), while a specific primer pair secY-F/secY-R was employed to amplify a ~1.4 kb segment of the secY gene. Target fragments were successfully amplified from all symptomatic samples but not from the healthy control. These amplicons were cloned and sequenced.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1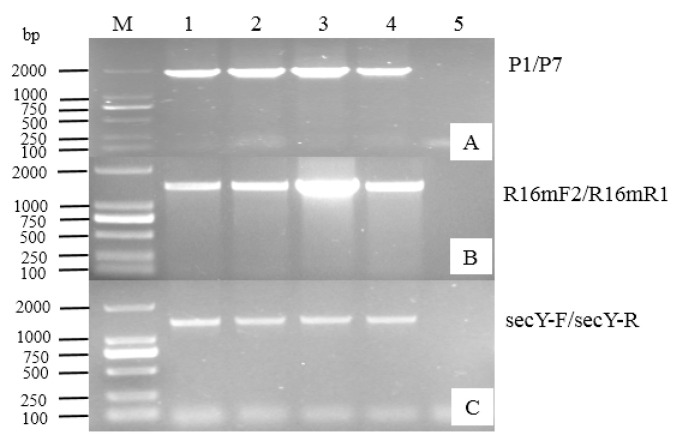 Figure 2
Figure 2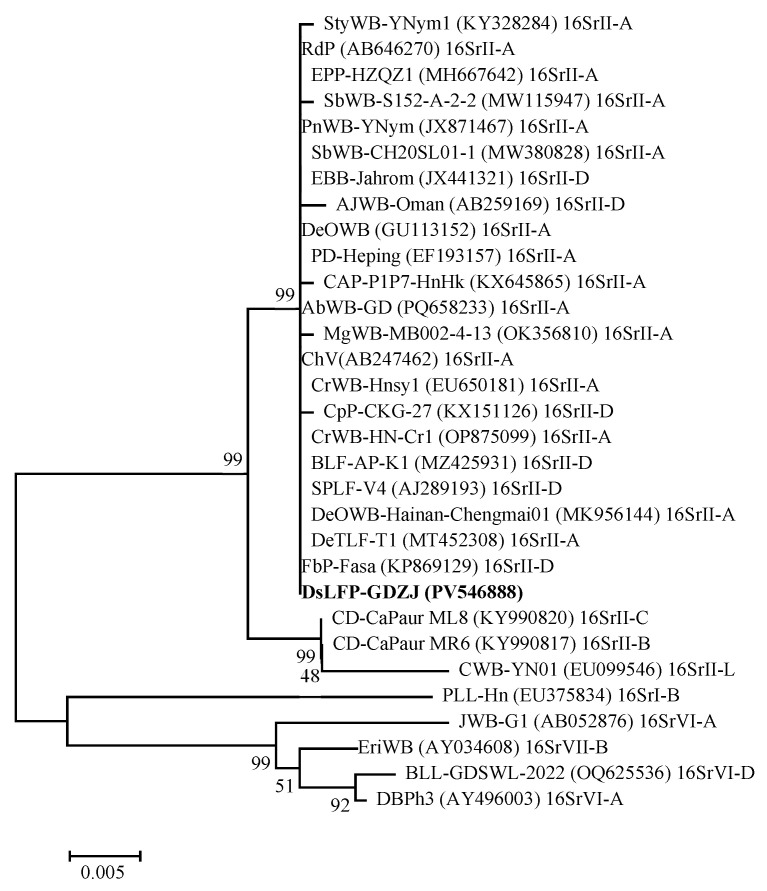 Figure 3
Figure 3 Figure 4
Figure 4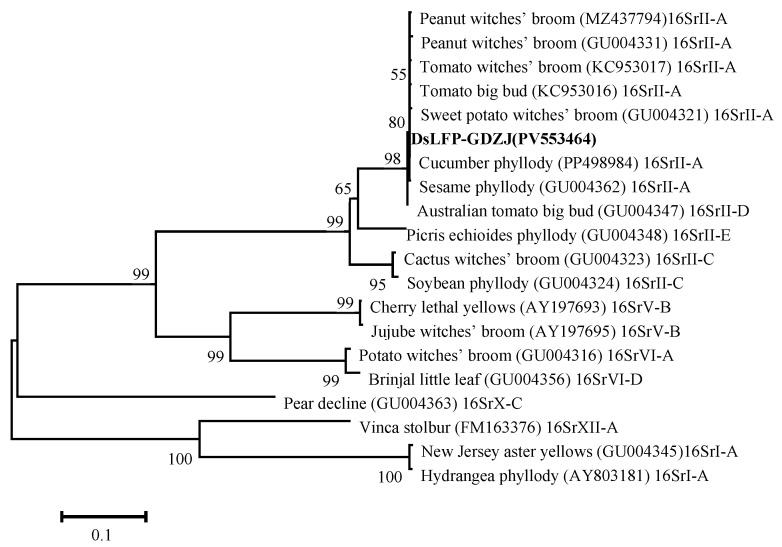 Figure 5
Figure 5- —Modern Agricultural Industrial Technology System of Guangdong Province
- —Project of Collaborative Innovation Center
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhytoplasmas and Hemiptera pathogens · Research on scale insects · Cocoa and Sweet Potato Agronomy
1. Introduction
Phytoplasmas, which belong to the genus Candidatus Phytoplasma within the class Mollicutes, are significant obligate prokaryotic bacteria that lack a cell wall and cannot be cultured outside of their hosts. These organisms exhibit pleomorphic forms, with sizes ranging from 200 to 800 nm, and possess relatively small genomes varying from 680 to 1600 kb [1,2]. They primarily proliferate within plant phloem tissues and insect haemolymph [3,4]. Phytoplasmas are transmitted by insect vectors, mainly leafhoppers, planthoppers, and psyllids, as well as through dodder and grafting. Notably, recent studies have also identified that phytoplasma can be transmitted by seed [5,6,7,8,9,10]. Phytoplasmas have a wide host range and can infect thousands of plant species, including fruits, vegetables, ornamental plants, and shade and timber trees [11]. Infected plants exhibit a series of typical symptoms such as phyllody, virescence, witches’ broom, yellowing, little leaf, proliferation, necrosis, dieback, stunting, and bunchy top [8,9,12,13,14]. Phytoplasmas have constituted a class of highly damaging plant pathogens, posing a significant constraint on productivity and quality in global agriculture and forestry [1,15].
Symptom observation and molecular detection are widely utilized approaches for phytoplasma disease diagnosis. Recently, 16S rRNA gene-based molecular analysis has been used to identify and classify phytoplasmas. If the full-length or nearly full-length 16S rDNA sequence (>1500 bp) of a strain sharing greater than 98.65% nucleotide identity with the reference strain is considered a member of the respective ‘Ca. Phytoplasma’ species. When the 16S rDNA sequence alone is insufficient to differentiate ‘Ca. Phytoplasma’ species, additional conserved or housekeeping genes, such as rp, tuf, secY, and secA, are suggested to confirm or support the ‘Ca. Phytoplasma’ species identification. The threshold for these genes is set at 97.5% for rp and tuf, 95.7% for secY, and 95.0% for secA, enabling clear differentiation among them [2,16]. The classification into groups and subgroups is based on restriction fragment length polymorphism (RFLP) patterns of the F2nR2 fragment (about 1250 bp) of the 16S rDNA. To date, 49 named ‘Ca. Phytoplasma’ species have been identified [2,16,17].
Desmodium styracifolium (Osb.) Merr. is a member of the Desmodium genus within the Leguminosae family. In China, it is known as “Guang Jin qian cao” and is an important medicinal plant widely used in traditional Chinese medicine for treating many different human diseases [18,19]. To date, only two Desmodium plants have been reported to be infected by phytoplasma, D. ovalifolium and D. triflorum. D. ovalifolium witches’ broom phytoplasma was found in Hainan Province, and D. triflorum little leaf phytoplasma was found in Taiwan [20].
In this study, D. styracifolium plants showing typical little leaf symptoms were found in Zhanjiang city of Guangdong province, China. To identify the causative phytoplasma, molecular detection, phylogenetic, and virtual RFLP (restriction fragment length polymorphism) analyses were employed to classify phytoplasmas infecting these symptomatic plants. Our findings represent the first report of a phytoplasma associated with D. styracifolium little leaf disease in China. This study will contribute to a better understanding of the distribution and molecular diversity of phytoplasmas in China.
2. Materials and Methods
2.1. Field Investigation and Sample Collection
In September 2024, D. styracifolium plants exhibiting little leaf symptoms were found in Zhanjiang city of Guangdong province, China (21°15′8″ N, 110°6′11″ E). Disease incidence was investigated according to symptoms observed in the field. Leaf samples were randomly collected from four diseased plants and one healthy plant.
2.2. DNA Extraction and Molecular Identification
Total DNA was extracted from the collected five leaf samples using the Easypure Plant Genomic DNA Kit (Beijing TransGen Biotech Co., Beijing, China). PCR detection was performed with universal ribosomal primers P1/P7 [21,22] and R16mF2/R16mR1 [23] targeting the 16S-23S ribosomal gene, as well as the specific primers secY-F/secY-R (5′-GCGGAAGAAGCTATTAT-3′/5′-CGAATAACATAATATAATTGATTCC-3′) for the secY gene [24]. The amplification protocols are 94 °C for 10 min, 35 cycles each of 94 °C for 30 s, 50 °C for 30 s, and 72 °C for 2 min, and a final extension at 72 °C for 10 min. PCR products were analyzed by electrophoresis on 1% agarose gel, visualized under a UV transilluminator (Bio-Rad, Hercules, CA, USA). The amplified fragments were purified and ligated into the pMD-19T vector (Takara Bio Inc., Kusatsu, Japan). The ligation mixtures were transformed into Escherichia coli DH5α competent cells using the heat shock method. Positive clones were confirmed by bidirectional DNA sequencing by the Sanger method at Sangon Biotech Co. (Shanghai, China).
2.3. Sequence Analysis
Initial identification of ‘Ca. Phytoplasma’ species was performed by sequence comparison using the BLASTn algorithm in the GenBank database (www.ncbi.nlm.nih.gov, accessed on 20 May 2025). The homology analysis of 16S rRNA and secY gene sequences was performed using the online MUSCLE tool (https://www.ebi.ac.uk/Tools/msa/muscle/, accessed on 27 May 2025). Phylogenetic trees were constructed using the neighbor-joining method implemented in MEGA 6.0 with 1000 bootstrap replicates. Two separate datasets retrieved from the GenBank database were analyzed: one comprising the 16S rRNA gene sequences of DsLFP-GDZJ and 30 other phytoplasma strains (Table S1), and the other comprising the secY gene sequences of DsLFP-GDZJ and 19 other phytoplasma strains (Table S2). Virtual RFLP analysis of the F2nR2 fragment of the 16S rRNA gene was conducted to define the 16Sr group and subgroup using the online interactive tool iPhyClassifier (https://plantpathology.ba.ars.usda.gov/cgi-bin/resource/iphyclassifier.cgi, accessed on 27 May 2025) [25].
3. Results
3.1. Field Surveys
In September 2024, D. styracifolium plants in the Mazhang district of Zhanjiang, Guangdong province, China, displayed typical little leaf symptoms, characterized by the emergence of abnormally small leaves (Figure 1). These symptoms strongly indicated a potential phytoplasma infection. About 300 plants were surveyed in the field, among which 10 exhibited symptoms, resulting in a disease incidence of 3.33%.
3.2. Molecular Detection
Four symptomatic and one healthy sample were detected by PCR using phytoplasma 16S rDNA universal primer pairs, P1/P7 and R16mF2/mR1. The expected fragments of approximately 1.8 or 1.4 kb were obtained from four symptomatic samples, whereas no fragment was obtained from the healthy sample (Figure 2). The PCR result indicated that these D. styracifolium plants with little leaf were infected by phytoplasma. To further verify the presence of phytoplasma, we employed a specific primer pair, secY-F/secY-R, to amplify the secY gene. The PCR results also showed that the expected fragment of approximately 1.4 kb was obtained from all diseased samples, with no fragment in the healthy sample (Figure 2). These results conclusively confirmed the presence of phytoplasma in these diseased D. styracifolium plants. The identified phytoplasma strain was tentatively named DsLFP-GDZJ.
3.3. Sequence Analysis of 16S rDNA
The target fragment amplified using primer pair P1/P7 was gel-purified, cloned, and sequenced. The fragment amplified from the total DNA of D. styracifolium symptomatic samples in Guangdong was 1806 bp in length. The nucleotide sequence was submitted to the National Center for Biotechnology Information (NCBI) to get the GenBank accession number PV546888. Sequence analysis revealed that the fragment contains the 16S rDNA (1–1525 nt), the 16S-23S rDNA intergenic spacer region (1526–1751 nt), which includes the rDNA-Ile gene (1604–1680 nt), and a partial segment of the 23S rDNA (1752–1806 nt).
BLAST analysis revealed that the 16S rDNA sequence of DsLFP-GDZJ shared the highest identity with those of phytoplasma strains belonging to the peanut witches’ broom group (16SrII group). Further, MUSCLE analysis results (Table 1) showed that the 16S rDNA sequence of DsLFP-GDZJ shared the highest identity (99.67–100%) with those of ‘Ca. Phytoplasma australasiaticum’ (subgroup 16SrII-A and 16SrII-D) strains. Among them, DsLFP-GDZJ shared 100% identity with 11 phytoplasma strains associated with adzuki bean witches’ broom (PQ658233), eggplant phyllody (MH667642), Cleome rutidosperma witches’ broom (OP875099), soybean witches’ broom (MW680828), Crotalaria witches’ broom (EU650181), Chrysanthemum virescence (AB247462), sweet potato little leaf (AJ289193), Desmodium ovalifolium witches’ broom (GU113152 and MK956144), Desmodium triflorum little leaf (MT452308), and pear decline (EF193157). The identity with other strains (subgroup 16SrII-B, 16SrII-C, and 16SrII-L) ranged from 98.00% to 98.56%. In contrast, the similarity with strains from the ash yellows group (16SrVII group), elm yellows group (16SrV group), and aster yellows phytoplasma group (16SrI group) was much lower (89.25–91.12%). These results indicate that DsLFP-GDZJ is a strain related to ‘Ca. Phytoplasma australasiaticum’.
The phylogenetic analysis (Figure 3) of 16S rDNA revealed that DsLFP-GDZJ was clustered within a major branch alongside 25 related phytoplasma strains belonging to the 16SrII group, indicating a close phylogenetic relationship. In contrast, it showed a distant relationship with strains from groups 16SrI, 16SrVI, and 16SrVII. Further analysis revealed that DsLFP-GDZJ formed a tight subclade with strains from 16SrII-A and 16SrII-D subgroups, demonstrating the closest genetic affinity.
The online phytoplasma classification tool, iPhyClassifier, was used to conduct the virtual RFLP analysis of the F2nR2 fragment of the DsLFP-GDZJ 16S rRNA gene. The results (Figure 4) showed that DsLFP-GDZJ exhibited an identical restriction pattern to the reference strain of subgroup 16SrII-A (L33765), with a similarity coefficient of 1.00, which further demonstrates that DsLFP-GDZJ belongs to the 16SrII-A subgroup.
3.4. Sequence Analysis of the secY Gene
The target fragment amplified using primer pair secY-F/secY-R was gel-purified, cloned, and sequenced. The amplified fragment from diseased D. styracifolium samples in Guangdong was 1425 bp in length, and contained the full length of the secY gene (74–1336 nt). The nucleotide sequence was submitted to NCBI to get the GenBank accession number PV553464.
The BLAST analysis revealed that the secY sequence of DsLFP-GDZJ shared 100% identity with several phytoplasma strains belonging to the 16SrII group, including four strains: NCHU2014, NCHU2022, WF_GM2021, and SPWB of Ca. Phytoplasma australasiaticum (CP040925, CP097312, CP133702, CP171825), and other strains associated with Vigna angularis witches’ broom (PQ619118), Perilla frutescens witches’ broom (MW310228), cowpea virescence (KC953013), Gendarussa vulgaris witches’ broom (MN543069), cauliflower phyllody (KC953012), long bean phyllody (AB703251), cucumber phyllody (PP498984), black gram witches’ broom (AB703249), Crotalaria witches’ broom (JF834194), and shaggy button witches’ broom (AB703252). Furthermore, the phylogenetic tree constructed based on secY gene sequences of DsLFP-GDZJ and 19 other phytoplasmas showed that the DsLFP-GDZJ strain was clustered with seven strains belonging to subgroup 16SrII-A and one strain belonging to subgroup 16SrII-D (Figure 5). These findings collectively suggest that the DsLFP-GDZJ strain is a member of the 16SrII-A subgroup of phytoplasmas.
4. Discussion
Phytoplasma diseases are one of the most devastating plant diseases worldwide. Phytoplasmas can infect over 1000 plant species, including critical food crops, vegetables, and fruit trees, often causing severe yield losses and substantial economic damage [11]. In China, over 100 different plant species have been reported to be infected by diverse phytoplasmas, resulting in significant economic losses to agricultural and forestry production [26].
In 2024, D. styracifolium plants exhibiting little leaf symptoms were first observed at a field in China. PCR testing confirmed that the samples were infected with a phytoplasma, and the strain was tentatively designated as D. styracifolium little leaf phytoplasma (DsLFP-GDZJ). The 16S rDNA sequence of DsLFP-GDZJ showed 99.67% to 100% similarity with strains of ‘Ca. Phytoplasma australasiaticum’ from 16SrII-A and 16SrII-D subgroups. The phylogenetic analysis of 16S rDNA also revealed that DsLFP-GDZJ is clustered into 16SrII-A and 16SrII-D subgroups. According to the iPhyClassifier analysis results, DsLFP-GDZJ belonged to the 16SrII-A subgroup. According to current international phytoplasma classification standards [16], DsLFP-GDZJ is a new strain of ‘Ca. Phytoplasma australasiaticum’ (belonged to the 16SrII-A group). The secY gene sequence analysis also revealed that DsLFP-GDZJ shared the highest similarity and closest phylogenetic relationship with strains of the 16SrII-A subgroup. This study represents the first report of a phytoplasma disease on D. styracifolium, specifically caused by a strain of ‘Ca. Phytoplasma australasiaticum’.
In China, phytoplasmas display high genetic diversity and a wide geographical distribution, with 12 distinct groups identified, namely 16SrI, 16SrII, 16SrV, 16SrVII, the X-disease group (16SrIII), clover proliferation group (16SrVI), loofah witches’ broom group (16SrVIII), apple proliferation group (16SrX), rice yellow dwarf group (16SrXI), stolbur group (16SrXII), bermudagrass white leaf group (16SrXIV), and Malaysian periwinkle virescence group (16SrXXXII) [20,26]. Guangdong province is located in the tropical and subtropical regions of southern China and is renowned for its abundant biological resources and genetic diversity. However, Guangdong also suffers significantly from phytoplasma-associated diseases. To date, at least four groups (16SrI, 16SrII, 16SrV, and 16SrVI) of phytoplasmas have been reported to be distributed in Guangdong. Specifically, 16SrI phytoplasma was associated with rice orange leaf, mulberry yellow dwarf, gum tree yellowing, and witches’ broom [27,28,29]. 16SrV phytoplasma was associated with jujube witches’ broom [30]; 16SrVI phytoplasma with Breynianivosa little leaf [31]; and 16SrII phytoplasmas with eggplant phyllody, peanut witches’ broom, cowpea phyllody, tomato big bud, and cucumber phyllody [31,32,33,34,35,36]. Thus, the 16SrII phytoplasmas may represent the dominant group affecting Solanaceae, Leguminosae, and Cucurbitaceae in Guangdong, China. This study reveals that D. styracifolium (Osb.) Merr. serves as a new host plant for phytoplasmas of the 16SrII-A subgroup.
This study contributes to understanding the distribution, host range, and molecular diversity of phytoplasmas present in China. Phytoplasmas are mainly transmitted to plants by insects such as leafhoppers, planthoppers, psyllids, etc. To prevent serious crop losses caused by phytoplasmas, disrupting the transmission pathways is the most effective strategy. Therefore, further studies should be performed to investigate various transmission routes of phytoplasmas and plan, develop, and implement effective vector management and disease control strategies.
5. Conclusions
In 2024, a new disease was discovered in D. styracifolium plants in the fields of Zhanjiang city, Guangdong province, characterized by typical phytoplasma-induced symptoms such as little leaf. The causative phytoplasma, designated as DsLFP-GDZJ, was detected and identified using molecular biology methods. According to the results of sequence similarity analysis and phylogenetic analysis of 16S rRNA and secY genes, and virtual RFLP analysis of the 16S rRNA gene, DsLFP-GDZJ is a strain of ‘Ca. Phytoplasma australasiaticum’, and belongs to the 16SrII-A subgroup. To our knowledge, this is the first report of ‘Ca. Phytoplasma australasiaticum’ (16SrII-A group) associated with D. styracifolium little leaf disease in China, thereby establishing D. styracifolium (Osb.) Merr. as a new host plant of phytoplasma.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bertaccini A. Duduk B. Paltrinieri S. Contaldo N. Phytoplasmas and phytoplasma diseases: A severe threat to agriculture Am. J. Plant Sci.201451763178810.4236/ajps.2014.512191 · doi ↗
- 2Wei W. Zhao Y. Phytoplasma taxonomy: Nomenclature, classification, and identification Biology 202211111910.3390/biology 1108111935892975 PMC 9394401 · doi ↗ · pubmed ↗
- 3Hogenhout S.A. Oshima K. Ammar E.-D. Kakizawa S. Kingdom H.N. Namba S. Phytoplasmas: Bacteria that manipulate plants and insects Mol. Plant Pathol.2008940342310.1111/j.1364-3703.2008.00472.x 18705857 PMC 6640453 · doi ↗ · pubmed ↗
- 4Koinuma H. Maejima K. Tokuda R. Kitazawa Y. Nijo T. Wei W. Kumita K. Miyazaki A. Namba S. Yamaji Y. Spatiotemporal dynamics and quantitative analysis of phytoplasmas in insect vectors Sci. Rep.202010429110.1038/s 41598-020-61042-x 32152370 PMC 7062745 · doi ↗ · pubmed ↗
- 5Sugio A. Mac Lean A.M. Kingdom H.N. Grieve V.M. Manimekalai R. Hogenhout S.A. Diverse targets of phytoplasma effectors: From plant development to defense against insects Annu. Rev. Phytopathol.201149759510.1146/annurev-phyto-072910-09532321838574 · doi ↗ · pubmed ↗
- 6Salehia M. Rasoulpour R. Izadpanah K. Molecular characterization, vector identification and partial host range determination of phytoplasmas associated with faba bean phyllody in Iran Crop Prot.201689122010.1016/j.cropro.2016.06.016 · doi ↗
- 7Akhtar K.P. Sarwar G. Dickinson M. Ahmad M. Haq M.A. Hameed S. Iqbal M.J. Sesame phyllody disease: Its symptomatology, etiology, and transmission in Pakistan Turk. J. Agric. For.20093347748610.3906/tar-0901-23 · doi ↗
- 8Bertaccini A. Phytoplasmas: Diversity, taxonomy, and epidemiology Front. Biosci 20071267368910.2741/209217127328 · doi ↗ · pubmed ↗