Rainfall-Driven Mobilisation of Clinically Relevant Burkholderia pseudomallei in a Groundwater-Connected Urban Creek, Northern Australia
Kaitlin Janssen-Groesbeek, Jennifer Elliman, Catherine Rush, Jeffrey Warner

TL;DR
This study shows that rainfall can stir up dangerous Burkholderia pseudomallei bacteria in urban creeks connected to groundwater, increasing the risk of melioidosis in northern Australia.
Contribution
The study identifies groundwater-connected urban creeks as a mobile reservoir for clinically relevant B. pseudomallei under rainfall-driven conditions.
Findings
B. pseudomallei was detected in turbid, flowing water after ≥30 mm of rainfall in both wet and dry seasons.
MLST identified 18 sequence types, including 12 novel types, with six matching clinical isolates from Townsville.
The findings suggest that rainfall-driven processes in groundwater-connected creeks increase environmental exposure risk.
Abstract
Burkholderia pseudomallei is a saprophytic environmental bacterium and the causative agent of melioidosis, a serious opportunistic infection in tropical regions, including northern Australia. Infection occurs following environmental exposure via percutaneous inoculation, ingestion, or inhalation; however, the environmental reservoirs and transmission pathways responsible for human disease remain poorly defined. Groundwater has been implicated as a potential source of infection, but the factors influencing the persistence and mobility of B. pseudomallei in surface waters in North Queensland are not well understood. Water samples were collected from a groundwater-connected seasonal creek in Townsville, North Queensland, over a 12-month period encompassing wet and dry seasons. Samples were cultured on Ashdown agar and confirmed as B. pseudomallei by qPCR. Multi-locus sequence typing (MLST)…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5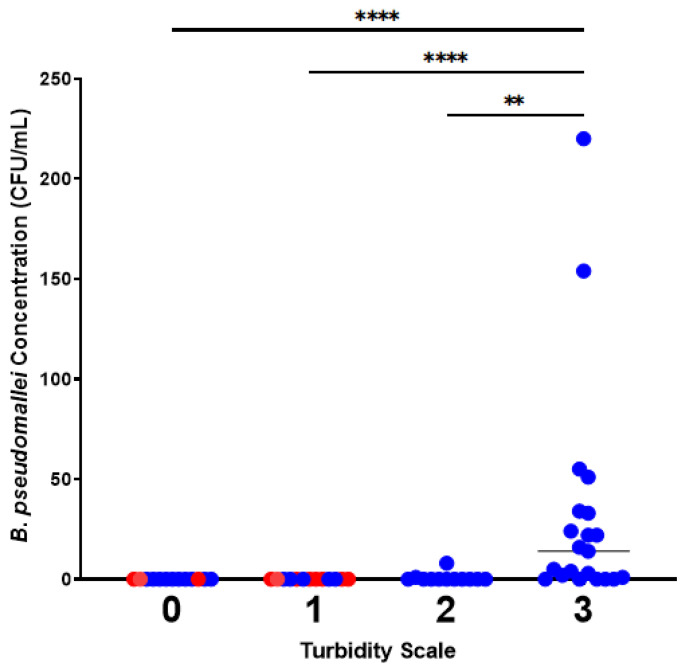 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9- —James Cook University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBurkholderia infections and melioidosis · Research on Leishmaniasis Studies · Plant Pathogenic Bacteria Studies
1. Introduction
Burkholderia pseudomallei is a Gram-negative environmental bacterium and the causative agent of melioidosis, a potentially severe opportunistic infection of humans and animals in tropical regions. Infection follows exposure to environmental B. pseudomallei via inoculation, ingestion, or inhalation, yet the specific reservoirs and transmission pathways responsible for human disease are often difficult to identify [1,2,3]. In northern Australia, melioidosis remains a major public health concern, with incidence increasing in recent years [4], underscoring the need to better define environmentally driven exposure pathways.
Groundwater is now recognised as a key environmental reservoir and exposure pathway for B. pseudomallei in northern Australia [5,6,7,8,9]. Repeated detection of the organism in groundwater discharging to surface watercourses implicates groundwater–surface water interfaces as important points of human and animal contact [5,8,10]. Hydrologic settings that promote discharge, including springs, seepages, and bank outflows, provide plausible exposure routes consistent with observed epidemiological patterns [7]. In North Queensland, the spatial clustering of melioidosis has been linked to geology, soil characteristics, and surface and subsurface water movement, emphasising the role of catchment hydrology in governing mobilisation and availability of B. pseudomallei [11,12,13].
International studies reinforce the importance of hydrologically mediated exposure. In Laos, rainfall-driven runoff has been used to characterise B. pseudomallei distribution and genetic diversity, demonstrating mobilisation into surface waters during wet conditions [14]. Melioidosis incidence in Northeast Thailand shows strong seasonal variation correlating with monsoon rainfall, where rice paddy water and runoff fields are primary reservoirs for human exposure [15,16]. In Papua New Guinea, lagoons and near-shore aquatic environments have been implicated as exposure settings, highlighting the role of aquatic habitats alongside soils as pathogen reservoirs [17,18]. Across Australia, landscape modification and altered recharge–runoff regimes further influence B. pseudomallei persistence and transport, particularly in groundwater-connected systems [9,19,20,21].
Climatic variability strongly modulates these hydrologic connections and melioidosis risk. Heavy rainfall is associated with increased incidence in Darwin, Northern Territory [22], while in Townsville, North Queensland, humidity and rainfall, particularly prolonged rainfall, are important predictors of case occurrence [23]. Queensland surveillance data demonstrate marked interannual variability in notifications, with sharp increases during unusually wet years [24,25]. Climate projections indicate increasing rainfall intensity and flash-flood risk across northern Australia [26], conditions likely to enhance aquifer–stream exchange and expand environmental exposure to B. pseudomallei.
Despite growing recognition of groundwater as a reservoir, the temporal dynamics of B. pseudomallei mobilisation and persistence within seasonal surface-groundwater systems remain poorly characterised, particularly in urban and peri-urban North Queensland. Seasonal creeks represent dynamic interfaces where groundwater discharge, runoff, and sediment transport converge, yet their role as transient exposure pathways has not been systematically examined.
This study characterises the temporal dynamics of B. pseudomallei in a seasonal creek in northern Queensland, focusing on rainfall, water flow, and turbidity, and assesses the clinical relevance of environmental isolates using genomic epidemiology. By integrating longitudinal environmental sampling with sequence-based analysis, this work clarifies how hydrologically driven processes generate short-term exposure risk in groundwater-connected urban waterways.
2. Materials and Methods
2.1. Longitudinal Study at Selected Location
A total of four sample sites were initially screened for suitability for use in this study. The selection criteria and results of selection are included in Supplementary Materials S1–S8. The inclusion criteria comprised detection of high concentrations of B. pseudomallei, proximity of the sampling site to James Cook University, and representation of a broad hydrological catchment, defined as multiple upstream watercourses converging at a single downstream sampling location (Figure 1). From the four sample sites, Goondaloo Creek (latitude 19.323687° S, longitude 146.762842° E) was selected for longitudinal investigation after it was determined to fulfil all the inclusion criteria (Supplementary Materials S1–S8). Water samples were collected weekly for over a year (February 2022–February 2023) whenever the creek contained water. In addition, the sampling frequency was increased opportunistically during periods of sustained rainfall. Daily rainfall data were collected from the Bureau of Meteorology, Climate Data Online [25]. A visual turbidity scale, with four qualitative categories, was used to assess collected water samples (Figure 2). The presence or absence of flowing water was recorded.
2.2. Sample Processing and Isolation of B. pseudomallei
One litre of groundwater from the study site was collected into sterile bottles. Ten replicate 100 µL volumes of sample water were inoculated onto Ashdown agar (ASH-A), incubated for two days at 37 °C in ambient air, and analysed for typical colonial morphology of B. pseudomallei to determine CFU/mL [28,29]. The sum of the characteristic B. pseudomallei colonies from all ten replicate plates was used to determine CFU/mL. The remaining water was filtered through 0.22 µm filters, and the paper was incubated in Ashdown broth (ASH-B) using similar incubation conditions. Approximately 10 µL of ASH-B was dilution-streaked onto ASH-A from each sample after one, two and five days of incubation. The remaining water samples were filtered and processed to enhance the sensitivity of B. pseudomallei detection. ASH-A and ASH-B media were selected in accordance with published consensus guidelines [1]. Comparative evaluations of selective media for B. pseudomallei are limited; consequently, Ashdown agar has remained the preferred selective medium in endemic regions. Although Ashdown agar is considered the standard and demonstrates specificity approaching 100%, it may inhibit the growth of mucoid and smooth colony variants of B. pseudomallei, as well as gentamicin-susceptible strains [30,31,32,33].
Quantification of B. pseudomallei colonies was determined by recognition of characteristic colony morphology and standard plate count methods. A subset of 10% or the square root of total colonies, if 10% was below 10 colonies, was collected to represent diversity [34,35,36]. A subset of suspect colonies was confirmed by Type III Secretion System (TTSS) qPCR modified from [5] (Supplementary Materials, Table S9). Briefly, the PCR reactions consisted of 1× GoTaq Colourless Master Mix (Promega, Madison, WI, USA), 0.4 µM of forward and reverse primers (Sigma-Aldrich, St. Louis, MO, USA), 0.256 µM of probe (Sigma-Aldrich), 50–100 ng of DNA, and molecular-grade H_2_O to 20 µL. Thermocycling conditions consisted of a 5 min initial denaturation at 95 °C, followed by 45 cycles of 15 s at 95 °C and 15 s at 59 °C. A single colony of bacteria was directly added to each PCR reaction (Colony PCR). A Mic qPCR Cycler (Bio Molecular Systems, Upper Coomera, QLD, Australia) (micPCR version 2.10.1) was used for all PCR reactions. Confirmed B. pseudomallei colonies were stored in 20% Glycerol Tryptic Soy Broth and frozen at −80 °C.
2.3. MLST PCR and Sequencing of B. pseudomallei Isolates
In the James Cook University Townsville PC3 facility, isolates were cultured on ASH-A, and DNA was extracted (Roche High Pure PCR Template kit (Roche Diagnostics, Mannheim, Germany)) from a single colony. Internal fragments of the seven MLST loci (ace, gltB, gmhD, lepA, lipA, narK, and ndh) were amplified using PubMLST primers and conditions (Supplementary Materials, Table S10) [37]. The reactions consisted of 1x GoTaq Colourless Master Mix (Promega), 0.4 µM of forward and reverse primers (Sigma-Aldrich), 50–100 ng of DNA and molecular-grade H_2_O to 30 µL. The thermocycling conditions for ace, gltB, lipA, narK, and ndh consisted of a 3 min initial denaturation at 95 °C, followed by 40 cycles of 30 s at 95 °C, 30 s at 62 °C, and 30 s at 72 °C, ending with a 10 min final elongation step at 72 °C. For gmhD and lepA, the annealing step was conducted at 59 °C after optimisation using a PCR temperature gradient. Amplification was performed on a CFX96 Real-Time System C1000 Touch Thermal Cycler (Bio-Rad Laboratories, Hercules, CA, USA), and products were verified by 1.5% agarose gel electrophoresis with a 100 bp HyperLadder (Meridian Bioscience, Eveleigh, NSW, Australia).
The seven MLST amplicons from each isolate were pooled, cleaned (Wizard SV Gel and PCR Clean-up System kit (Promega, Madison, WI, USA)), quantified (Qubit 3.0 Fluorometer (Thermo Fisher Scientific, Scoresby, VIC, Australia) and Qubit dsDNA HS Assay kit (Thermo Fisher Scientific, Scoresby, VIC, Australia)), and normalised (62 ng or approximately 200 fmol of DNA). Libraries were prepared with the ONT Native Barcoding kit (SQK-NBD114.24) (Oxford Nanopore Technologies, Littlemore, OX4 4DQ, United Kingdom), with up to 24 barcoded DNA samples pooled, and then run on a R10.4.1 FLO-MIN114 MinION flow cell. High-accuracy basecalling with barcode splitting was used; FASTQ files were exported per the Mk1C manual. Read quality and length were assessed with NanoPlot (version 1.42.0) [38,39]. For the ST assignment, approximately 8000 reads per isolate (more than 1000 reads per locus) were analysed via the Center for Genomic Epidemiology (CGE) webservice “https://cge.food.dtu.dk/services/MLST/ (accessed on 15 May 2024)” (v2.0.9 (11 May 2022)) using k-mer alignment (KMA; minimum depth ≥ 80×) against the B. pseudomallei pubMLST database (19 June 2023) [40,41]. KMA consensus sequences were also queried on pubMLST to confirm allele calls “https://pubmlst.org/organisms/burkholderia-pseudomallei (accessed 15 May 2024)” [42]. Twelve isolates underwent Illumina sequencing to validate the ONT sequencing results. Novel alleles and STs were submitted to PubMLST (Supplementary Materials, Tables S21 and S22).
2.4. Data Analysis
All descriptive statistics, measures of normality, and graphs were generated in GraphPad Prism (v10.0.1); all tests were nonparametric. The proportion of culture-positive samples was calculated with a Fraction of Total test. B. pseudomallei concentrations were compared between flowing versus non-flowing water by a Mann–Whitney test, and across turbidity categories by a Kruskal–Wallis test with Dunn’s Multiple Comparisons. Associations with climatic variables were assessed using Spearman’s Rank Correlation Coefficient and simple linear regression. Observed versus expected distributions of Goondaloo Creek STs were tested with a Chi-Square Test of Good Fitness test.
For phylogenetic analysis, concatenated MLST sequences from Townsville B. pseudomallei isolates, together with associated metadata spanning 1996–2023, were exported from pubMLST [42], merged with Goondaloo Creek STs, and rooted with Thailand ST10 [14]. Alignments were built with MAFFT (v7.505), and trees were inferred in IQ-TREE (v2.2.2.2) using ModelFinder with 1000 nonparametric bootstraps (nodes ≥ 80 shown) [14,43,44,45,46,47,48,49]. Trees were annotated and edited in iTOL (v6.9.1) [14,50].
3. Results
3.1. Annual Prevalence of B. pseudomallei in Creek Water
The prevalence of B. pseudomallei in creek water collected over a year from Goondaloo Creek is presented in Figure 3. Water samples collected from both weekly and opportunistic sampling revealed 18 out of 59 samples (30.5% [95% confidence interval (CI): 20.25 to 43.15]) were culture-positive for B. pseudomallei (Supplementary Materials, Table S11). TTSS qPCR was used for confirmation of selected colonies (Supplementary Materials, Table S12). Of the 669 presumptive B. pseudomallei colonies observed on ASH-A, 66 (approximately 10%) were selected and subsequently confirmed as B. pseudomallei by TTSS-targeted qPCR.
3.2. Prevalence and Concentration of B. pseudomallei Correlates with Extent of Rainfall
Burkholderia pseudomallei was isolated from creek water only during the two days following heavy rainfall (Figure 3). The concentration of B. pseudomallei was strongly correlated with rainfall (p ≤ 0.0001; r = 0.8141; 95% CI: 0.7010–0.8872) (Figure 4). However, the total extent of rainfall was only a weak predictor of B. pseudomallei concentration (R^2^ = 0.4932; F(1, 57) = 55.47; p ≤ 0.0001), with a fitted regression model of y = 0.5810x − 3.631. Notably, B. pseudomallei was not detected until cumulative rainfall reached approximately 30 mm, suggesting the presence of a rainfall threshold associated with mobilisation or persistence (Supplementary Materials, Table S13).
3.3. Detection of B. pseudomallei Correlates with Water Flow and Water Turbidity
Burkholderia pseudomallei was exclusively isolated from flowing and turbid creek water compared with water that was not flowing and clear (Figure 5 and Figure 6) (Supplementary Materials, Tables S11 and S14). B. pseudomallei was only detected in turbidity-level-two or -three water (Figure 6 and Supplementary Materials, Table S15). Turbidity-level-two water yielded concentrations below 10 CFU/mL, while level-three water yielded concentrations up to 220 CFU/mL (level zero vs. three (p ≤ 0.0001), level one vs. three (p ≤ 0.0001), and level two vs. three (p = 0.0011)). These findings suggest that turbidity is a strong indicator of B. pseudomallei presence in water and may reflect the contribution of rainfall-driven soil erosion and surface runoff processes to the mobilisation of B. pseudomallei into aquatic environments.
3.4. Genomic Epidemiology of Goondaloo Creek Isolates
A total of 50 B. pseudomallei isolates were sequenced using ONT sequencing, two B. pseudomallei controls (ST276 and ST814) and 48 Goondaloo Creek B. pseudomallei isolates. Creek isolates chosen for sequencing were representative of STs present in water samples and across all sampling timepoints (Supplementary Materials S16–S18 and S20).
Targeted ONT sequencing of 48 isolates resolved 18 STs, comprising 12 novel STs and six previously reported in pubMLST. Two novel alleles were detected: lipA (single-nucleotide variant; accepted by pubMLST) and lepA (3 bp insertion; not accepted). Consequently, 45/48 isolates received complete ST assignments, while three clonally related isolates remained incomplete due to the novel lepA allele. Both alleles were confirmed by Illumina WGS (Supplementary Materials, Tables S19 and S22). The most frequent STs were ST283 (15/48, 31.3%), ST1966 (5/48, 10.4%), and ST2072 (5/48, 10.4%) (Figure 7 and Supplementary Materials, Table S23). The ST frequencies deviated from a uniform distribution (expected 5.556% per ST; p ≤ 0.0001).
Nine STs (ST276, ST283, ST624, ST1664, ST1966, ST1969, ST2070, ST2074, and ST2075) matched STs previously recorded from human and/or animal cases in pubMLST (Figure 8 and Supplementary Materials, Table S21). ST283 had the most clinical links (five human and five animal submissions).
Concordance with Townsville clinical epidemiology was evident: Goondaloo Creek ST1966, ST276, ST283, ST2070, ST2074, and ST2075 matched prior Townsville human clinical STs (Figure 9), and ST276 also matched previously detected environmental ST276 in Townsville. Additional creek STs (ST624, ST1969, ST2071–ST2080, and Unknown 1) were grouped within clades containing Townsville clinical STs, although these placements lacked bootstrap support.
4. Discussion
Groundwater and groundwater–surface water interfaces are increasingly recognised as important reservoirs and exposure pathways for B. pseudomallei in northern Australia [5,6,7,8,9]. This study provides longitudinal evidence that a seasonal creek functions as a hydrologically triggered conduit, rather than a static reservoir, for the mobilisation of B. pseudomallei into surface water following rainfall.
Isolation of B. pseudomallei was consistently preceded by rainfall events of approximately ≥30 mm and largely confined to periods of active flow and elevated turbidity. Concentrations increased with rainfall intensity and persisted for up to two days following heavy rainfall, extending during sustained precipitation. These patterns are consistent with sediment-linked mobilisation via lateral subsurface flow, erosion, and suspended sediment transport observed in other hydrologic systems [10,51,52]. In contrast, stagnant and typically clear creek conditions were associated with little or no detection, indicating that short-term mobilisation, rather than in situ persistence, governs presence in this system. The strong association between turbidity and B. pseudomallei concentration supports a sediment-mediated transport mechanism. Similar relationships have been reported in Laos, where rivers act as carriers for B. pseudomallei following rainfall [10,14]. Collectively, these findings suggest that seasonal creeks generate brief but potentially high-risk exposure windows immediately following rainfall events. Exposure appears to be associated with eroded soils containing B. pseudomallei, which contaminate groundwater and seasonal creeks following periods of high rainfall.
Genomic analysis identified substantial strain diversity, including multiple sequence types previously associated with human and animal infection. Several creek isolates matched Townsville clinical sequence types, including ST283 and ST276, supporting the clinical relevance of organisms mobilised through this system [44,53]. Clustering within established Townsville clades indicates longstanding local establishment. While MLST resolution was sufficient to demonstrate lineage overlap, whole-genome sequencing will be required to more precisely resolve transmission pathways and dispersal dynamics.
The short detection window contrasts with studies reporting broader wet-season persistence in drains and depositional environments [8,14], likely reflecting hydrologic context. Flowing creek systems may rapidly mobilise and export organisms downstream, whereas slower-moving or depositional systems permit longer persistence. Washed-out organisms may subsequently accumulate in downstream habitats, extending exposure risk beyond the immediate site.
These findings have important public health implications. In Darwin, increased hydrologic connectivity during La Niña conditions was hypothesised to facilitate expansion of specific B. pseudomallei lineages [9]. In Townsville, shifts in sequence type prevalence over time have been documented without a defined mechanism [44,53]. Groundwater-mediated transport through connected surface-water networks represents a plausible contributor to the redistribution of established local lineages across catchments, particularly during extreme rainfall events.
The sampling year was characterised by frequent environmental detection of B. pseudomallei (this study), alongside elevated regional melioidosis notifications during the same period [24]. Although direct attribution cannot be made, the concordance between rainfall intensity, environmental detection, and case incidence mirrors established associations observed in Darwin and Townsville [22,23] and aligns with outbreak reports following extreme rainfall elsewhere in Australia [2,54,55].
From a surveillance perspective, groundwater sampling from urban creeks provides an efficient approach for detecting B. pseudomallei across broad catchments, complementing soil-based investigations that are labour-intensive and spatially heterogeneous [19,56]. Incorporating rainfall thresholds, turbidity metrics, and hydrologic connectivity into monitoring frameworks may improve identification of high-risk exposure windows, particularly under projected climate scenarios characterised by increasing rainfall intensity and flooding [26].
Several limitations warrant consideration. This study focused on a single seasonal creek, and applicability to other catchments with differing geomorphology and aquifer connectivity remains uncertain. Weekly sampling may not have captured very short-lived mobilisation events, and rainfall data were derived from a station approximately nine kilometres from the site. Genomic analyses were limited to MLST. Future studies should integrate higher-frequency sampling, on-site hydrometeorological monitoring, sediment characterisation, and whole-genome sequencing across multiple catchments.
In North Queensland, where groundwater discharge, climatic variability, and urban expansion intersect, seasonal creeks represent dynamic interfaces capable of rapidly mobilising clinically relevant B. pseudomallei. As climate projections indicate increasing rainfall intensity and flash flooding [26], rainfall-triggered mobilisation events are likely to become more frequent. Integrating hydrologic monitoring, genomic epidemiology, and environmental surveillance will be critical for defining spatiotemporal risk and informing targeted public health interventions in a changing climate.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Limmathurotsakul D. Dance D.A. Wuthiekanun V. Kaestli M. Mayo M. Warner J. Wagner D.M. Tuanyok A. Wertheim H. Yoke Cheng T. Systematic review and consensus guidelines for environmental sampling of Burkholderia pseudomallei P Lo S Negl. Trop. Dis.20137 e 210510.1371/journal.pntd.000210523556010 PMC 3605150 · doi ↗ · pubmed ↗
- 2Yip T.W. Hewagama S. Mayo M. Price E.P. Sarovich D.S. Bastian I. Baird R.W. Spratt B.G. Currie B.J. Endemic melioidosis in residents of desert region after atypically intense rainfall in Central Australia, 2011 Emerg. Infect. Dis.2015211038104010.3201/eid 2106.14190825988301 PMC 4451904 · doi ↗ · pubmed ↗
- 3Chen P.S. Chen Y.S. Lin H.H. Liu P.J. Ni W.F. Hsueh P.T. Liang S.H. Chen C.L. Chen Y.L. Airborne transmission of melioidosis to humans from environmental aerosols contaminated with B. pseudomallei P Lo S Negl. Trop. Dis.20159 e 000383410.1371/journal.pntd.000383426061639 PMC 4462588 · doi ↗ · pubmed ↗
- 4Smith S. Horne P. Rubenach S. Gair R. Stewart J. Fairhead L. Hanson J. Increased incidence of melioidosis in Far North Queensland, Queensland, Australia, 1998–2019 Emerg. Infect. Dis.2021273119312310.3201/eid 2712.21130234808088 PMC 8632158 · doi ↗ · pubmed ↗
- 5Baker A. Tahani D. Gardiner C. Bristow K.L. Greenhill A.R. Warner J. Groundwater seeps facilitate exposure to Burkholderia pseudomallei Appl. Environ. Microbiol.2011777243724610.1128/AEM.05048-1121873480 PMC 3194885 · doi ↗ · pubmed ↗
- 6Mayo M. Kaesti M. Harrington G. Cheng A. Ward L. Karp D. Jolly P. Godoy D. Spratt B. Currie B. Burkholderia pseudomallei in unchlorinated domestic bore water, tropical Northern Australia Emerg. Infect. Dis.2011171283128510.3201/eid 1707.10061421762588 PMC 3381386 · doi ↗ · pubmed ↗
- 7Baker A.L. Warner J.M. Burkholderia pseudomallei is frequently detected in groundwater that discharges to major watercourses in northern Australia Folia Microbiol.20166130130510.1007/s 12223-015-0438-326620184 · doi ↗ · pubmed ↗
- 8Rachlin A. Mayo M. Webb J.R. Kleinecke M. Rigas V. Harrington G. Currie B.J. Kaestli M. Whole-genome sequencing of Burkholderia pseudomallei from an urban melioidosis hot spot reveals a fine-scale population structure and localised spatial clustering in the environment Sci. Rep.202010544310.1038/s 41598-020-62300-832214186 PMC 7096523 · doi ↗ · pubmed ↗