Taxonomy and Phylogeny of Three New Euplotid Ciliates (Ciliophora, Euplotidae) Isolated from High Latitudes
Huan Li, Mikhail Tribun, Yuxuan Wang, Yunhan Wang, Sitong Li, Chunyu Lian, Xuming Pan

TL;DR
This paper describes three new species of euplotid ciliates found in high-latitude regions and uses molecular data to confirm their classification.
Contribution
The study introduces and validates three new euplotid ciliate species from high latitudes, expanding known biodiversity in these regions.
Findings
Three new euplotid ciliate species (Euplotes aspergilliformis, E. borealis, and E. verebkovi) were identified and described.
Molecular phylogenetic analysis confirmed the distinctiveness of the three new species.
Detailed morphological characteristics were provided for each new species.
Abstract
The biodiversity of euplotid ciliates in high-latitude regions has long been underestimated due to a historical research focus on mid- to low-latitude areas. This study describes three new species collected from high latitudes, Euplotes aspergilliformis sp. nov., E. borealis sp. nov., and E. verebkovi sp. nov. Euplotes aspergilliformis sp. nov. is characterized by medium body size (60–85 × 40–60 μm). It possesses nine frontoventral cirri, two marginal cirri and two caudal cirri with forked distal ends. It also has eight dorsal kinetids (with 13–17 dikinetids in the mid-dorsal kinety) and a double-eurystomus type silverline system. Euplotes borealis sp. nov. is a medium-sized (45–55 × 25–35 μm) freshwater ciliate. It can be recognized by nine frontoventral cirri, one marginal and two caudal cirri. It has nine dorsal kineties with about nine basal bodies in the mid-dorsal kinety, and a…
Click any figure to enlarge with its caption.
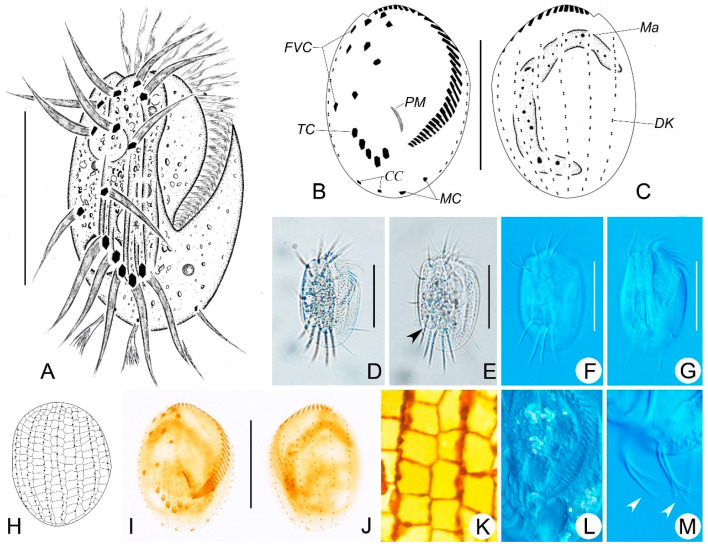 Figure 2
Figure 2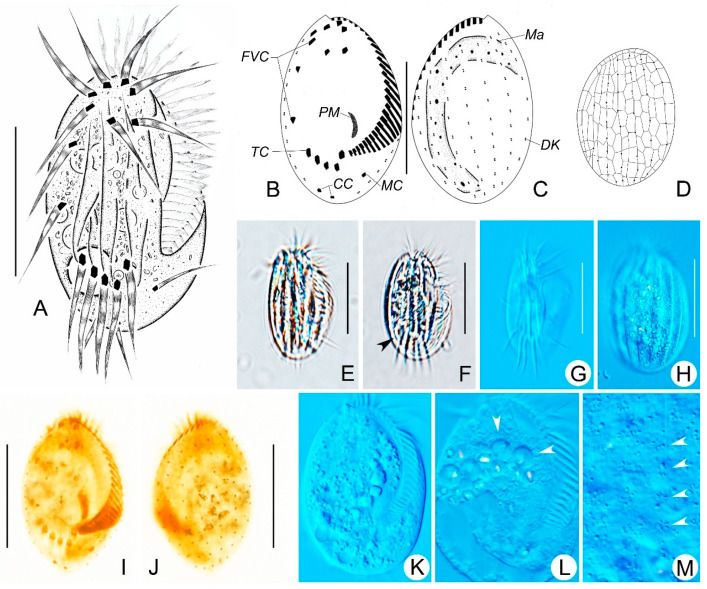 Figure 3
Figure 3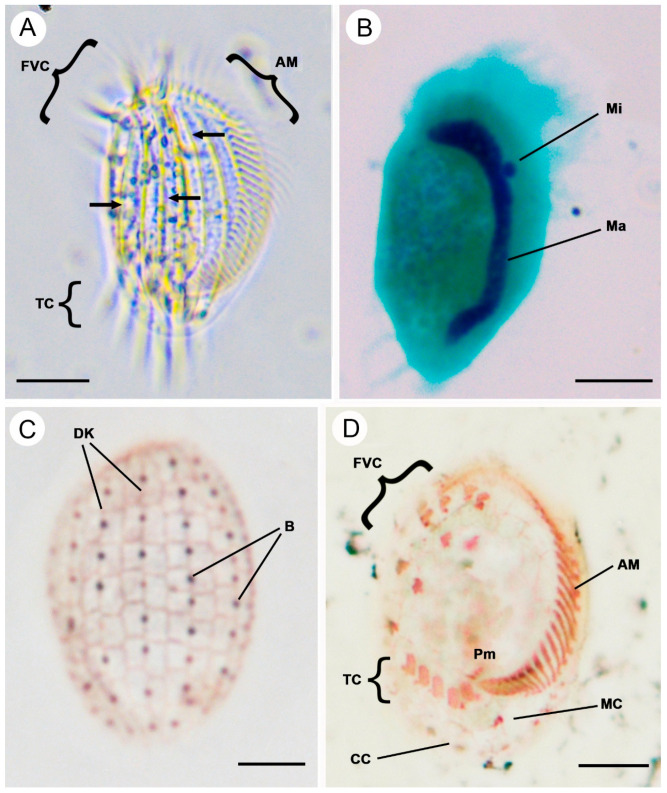 Figure 4
Figure 4| Character | Species | Min | Max | Mean | Med | SD | CV | n |
|---|---|---|---|---|---|---|---|---|
| Body length in μm |
| 63 | 81 | 69.7 | 68 | 5.9 | 8.5 | 19 |
|
| 39 | 55 | 46.0 | 45 | 4.6 | 10.0 | 25 | |
|
| 40 | 54 | 47.6 | 47.1 | 3.7 | 7.9 | 20 | |
| Body width in μm |
| 42 | 59 | 50.0 | 50 | 5.1 | 10.2 | 19 |
|
| 24 | 37 | 29.8 | 29 | 3.6 | 12.0 | 25 | |
|
| 22 | 31 | 27.2 | 26.8 | 2.8 | 10.4 | 20 | |
| Length of AZM |
| 44 | 52 | 47.2 | 47 | 2.0 | 4.2 | 19 |
|
| 26 | 40 | 31.2 | 31 | 2.9 | 9.3 | 25 | |
|
| 25 | 36 | 31.2 | 31.6 | 3.1 | 9.9 | 20 | |
| No. of AZM |
| 32 | 37 | 33.8 | 34 | 1.2 | 3.6 | 19 |
|
| 25 | 33 | 28.8 | 29 | 2.0 | 6.9 | 25 | |
|
| 27 | 34 | 29.8 | 29 | 1.9 | 6.6 | 13 | |
| No. of FVC |
| 9 | 9 | 9.0 | 9 | 0 | 0 | 19 |
|
| 9 | 9 | 9.0 | 9 | 0 | 0 | 25 | |
|
| 10 | 10 | 10.0 | 10.0 | 0 | 0 | 14 | |
| No. of TC |
| 5 | 5 | 5.0 | 5 | 0 | 0 | 19 |
|
| 5 | 5 | 5.0 | 5 | 0 | 0 | 25 | |
|
| 5 | 5 | 5.0 | 5 | 0 | 0 | 15 | |
| No. of MC |
| 2 | 2 | 2.0 | 2 | 0 | 0 | 19 |
|
| 1 | 1 | 1.0 | 1 | 0 | 0 | 25 | |
|
| 1 | 1 | 1.0 | 1.0 | 0 | 0 | 14 | |
| No. of CC |
| 2 | 2 | 2.0 | 2 | 0 | 0 | 19 |
|
| 2 | 2 | 2.0 | 2 | 0 | 0 | 25 | |
|
| 2 | 2 | 2.0 | 2.0 | 0 | 0 | 14 | |
| No. of DK |
| 8 | 8 | 8.0 | 8 | 0 | 0 | 19 |
|
| 9 | 9 | 9.0 | 9 | 0 | 0 | 25 | |
|
| 7 | 7 | 7.0 | 7.0 | 0 | 0 | 14 | |
| No. of dikinetids in mid-DK |
| 13 | 17 | 14.9 | 15 | 1.1 | 7.7 | 19 |
|
| 8 | 10 | 8.7 | 9 | 0 | 7.0 | 25 | |
|
| 8 | 9 | 8.3 | 8 | 0.4 | 5.8 | 15 | |
| No. of dikinetids in left-DK |
| 10 | 15 | 12.0 | 12 | 1.3 | 10.8 | 19 |
|
| 4 | 6 | 4.8 | 5 | 0 | 12.5 | 25 | |
|
| 6 | 10 | 8.2 | 8 | 0.9 | 11.6 | 15 | |
| No. of dikinetids in right-DK |
| 10 | 14 | 12.3 | 12 | 1.1 | 9.0 | 19 |
|
| 7 | 14 | 10.3 | 10 | 1.9 | 18.1 | 25 | |
|
| 5 | 8 | 6.4 | 6 | 1.1 | 17.4 | 15 |
| Body Size | AZM, n | DK, n | Mid-DK, n | Ma | Isolation Location | Original Description | |
|---|---|---|---|---|---|---|---|
|
| 60–85 × 40–60 | 32–37 | 8 | 13–17 | C | FW, China | Present |
|
| 140–240 × 80–160 | 52–62 | 14 | 28 * | 3 | Cameroon | [ |
| ca. 180 × ? | 13 | Chad | [ | ||||
| 140–240 × ? | 52–62 | 14 | 26–31 | 3 | Cameroon | [ | |
| ca. 169 × ? | 53–67 | 12–15 | 21–28 | 3 | Rwanda | [ | |
| 130–200 × 70–100 | 60–65 | 12–14 | 18–26 | 3 | FW, China | [ | |
|
| 180–220 × 110–155 | 63–93 | 12–13 | 24–37 | 3 | FW, China | [ |
|
| 105–160 × ? | 56 * | 8 | 32 * | 3 | FW | [ |
| 121.5–135 × 73.5–85.5 | 48–52 | 8–9 | 28–31 | C | FW, USA | [ | |
| 100–160 × 40–90 | 50–65 | 8–12 | 17–25 | 3 | FW | [ | |
| 140–180 × 95–135 | 45–55 | 9–11 | 27 * | 3 | FW, USA | [ | |
| 88–125 × 55–78 | 44–53 | 8–9 | 15–23 | C or 3 | FW, China | [ | |
| 98–133 × 67.2–105 | 35–40 | 3 | Soil, Paithan | [ | |||
|
| 60 × 40 | 19–22 | 7 | 10 | C or J | Soil, Austria | [ |
|
| 120–165 × 90 | T | [ | ||||
| 128–150 × 75–95 | 51–58 | 9 | 24–30 | T | FW, USA | [ | |
| 83–140 × 58–90 | 53–99 | 8–9 | 30 * | T | BW, Africa | [ | |
| 130–160 × 90–110 | 57–64 | 10 | 23–28 | T | FW, China | [ | |
| 110–200 × 70–130 | 76–85 | 9 | 28–35 | S | BW, USA | [ | |
| 130–153 × 81–95 | ca. 60 | 10–11 | T | FW, China | [ | ||
| 90–140 × 60–90 | 56–67 | 9 | 17–24 | T * | Marine, China | [ | |
| 110–160 × 60–80 | 59–70 | 9–10 | 19–24 | T or Y | FW, China | [ | |
| 100–1132 × 68–82 | 40–56 | 9–10 | 20–21 | T | FW, India | [ | |
|
| 130.7–133.3 × 82.2–85.2 | C | BW or FW, USA | [ | |||
| 40 | 8 | C | BW or FW | [ | |||
| 105–122 × 65–77 | 43–51 | 8–9 | 18–26 | C | FW, USA | [ | |
| 105–165 × 60–110 | 40–60 | 8–9 | 24 * | 3 | [ | ||
| 120–130 × 80–85 | 40–50 | 8 | C | FW, China | [ | ||
| 150–170 × 100–120 | 47–55 | 8 | 21–26 | C | FW, China | [ | |
| 105–165 × 60–110 | 48–50 | 8 | 24–30 | FW, Slovakia | [ | ||
| 107–119 × 72–82 | 42–46 | 8 | 18–24 | 3 | FW, India | [ | |
|
| 49–52 × 40–46 | 20–25 | 7 | 11–16 | C | FW, India | [ |
| Body Size | AZM, n | DK, n | Mid-DK, n | Ma | Isolation Location | Original | |
|---|---|---|---|---|---|---|---|
|
| 35–55 × 20–40 | 25–33 | 9 | 8–10 | C | FW, China | Present |
|
| 40–55 × 25–35 | 18–26 | 7 | 6–9 | C | FW, China | [ |
|
| 65 ** × ? | 35–38 | 7 | 13 * | C * | Caspian Sea | [ |
|
| 80–90 * × ? | S * | Germany | [ | |||
| 34–45 × 27–31 | 17 * | 8 | 5–7 | C * | Tidal marsh | [ | |
| 45 × ? | FW | [ | |||||
|
| 38 × 26 | 18–20 | 7 | Max 9 | 3 | FW, UK | [ |
|
| 46–58 × 32–40 | 28–32 | 8 | 12–16 | hook- to a horseshoe-shaped or 3 | BW in Chile | [ |
|
| 34–45 × 25–30 | 23–25 | 6 | 9–11 | hook-shaped or C | Marine, Chile | [ |
|
| 25–30 × ? | 25 * | 8 * | 5 * | C | Mobile Bay, | [ |
- —The National Natural Science Foundation of China
- —Startup Foundation for Doctors of Harbin Normal University
- —Excellent Youth Fund of Heilongjiang Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProtist diversity and phylogeny · Marine and coastal ecosystems · Genetic and Kidney Cyst Diseases
1. Background
Euplotid ciliates represent one of the most species-rich groups within the phylum Ciliophora, having garnered considerable attention from protozoologists over the past few decades [1,2,3,4,5,6]. The most speciose genus within this group, Euplotes Ehrenberg, 1830, represents one of the most complex and diverse taxa, with a cosmopolitan distribution across freshwater, marine, and terrestrial biotopes [7,8,9,10].
To date, over 160 nominal species have been recognized in Euplotes [11,12,13,14,15,16]. Traditionally, species identification in this genus relied primarily on morphological characters of living specimens. With the adoption of integrative taxonomic approaches, additional species-specific characteristics (e.g., ciliary pattern, silverline system) have been incorporated into the identification process [17,18,19]. In recent years, molecular data have also become widely utilized to resolve taxonomic ambiguities and delimit new species [20,21,22,23,24,25,26,27].
Despite this long history of morphological and taxonomic research on euplotids, investigations have predominantly concentrated on mid- to low-latitude regions. In contrast, high-latitude areas characterized by extreme environmental conditions, including prolonged ice cover, low temperatures, and pronounced seasonal fluctuations in light availability, have remained largely unexplored. The extreme selective pressures of high-latitude regions shape unique microbial communities in which Euplotes plays a crucial ecological role, and the recent discovery of considerable species diversity there underscores the critical need for more comprehensive surveys [28,29,30,31]. Currently, high-latitude regions remain critically undersampled for ciliate biodiversity, leaving a substantial gap in our understanding of the biogeography and evolutionary adaptations of the genus Euplotes. In the present study, we take an initial step toward addressing this gap by describing three new Euplotes species isolated from high-latitude areas and elucidating their phylogenetic positions.
2. Materials and Methods
2.1. Sampling and Morphological Methods
Euplotes aspergilliformis sp. nov. was collected on 22 November 2024 from the Hulan River (45°57′2″ N, 126°34′46″ E), Harbin, Heilongjiang Province, China, where the water temperature was 3 °C (Figure 1A–C). Euplotes borealis sp. nov. was found on 7 April 2025 from a small pond near Anda People’s Court (45°24′43″ N, 125°18′5″ E), Suihua, Heilongjiang Province, China, where the water temperature was 11.8 °C (Figure 1A,B,D). Euplotes verebkovi sp. nov. was isolated from the private farm “Verebkovo” (57°44′40.8″ N, 27°34′08.1″ E) in the Pskov region of Russia, which specializes in rainbow trout breeding.
Although attempts to establish clonal cultures were unsuccessful, no co-occurring morphologically similar Euplotes species were detected in the cultures. Moreover, each protargol-stained preparation, comprising approximately 30–50 individuals, displayed consistent morphological features, providing strong evidence that each isolate represents a single species. Initial cultures were established and maintained at room temperature (about 25 °C) in Petri dishes containing sterile distilled water with rice grains added to enrich the growth of bacteria as a food source for the ciliates. Living cells were observed in vivo using bright field and Nomarski differential interference contrast microscopy at magnifications between 100× and 1000× (Zeiss Axio Imager A2, Gottingen, Germany). The infraciliature and nuclear apparatus were revealed using Feulgen staining and protargol staining [32,33], while dry silver nitrate staining [34] was used to reveal the silverline systems. Drawings of stained specimens were made with the help of photomicrographs. Terminology is mainly according to Curds (1975) [11].
2.2. DNA Extraction, PCR Amplification, and Gene Sequencing
Living cells were isolated from the raw culture and repeatedly washed with sterile distilled water to remove potential contamination. The cells were then transferred to a 1.5 mL microfuge tube with a minimum volume of water. Genomic DNA was extracted from one to five cells using the DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions. The SSU rRNA gene was amplified with the primers Euk A: 5′-AAC CTG GTT GAT CCT GCC AGT-3′, Euk B: 5′-TGA TCC TTC TGC AGG TTC ACC TAC-3′, F9: 5′-CTGGTTGATCCTGCCAG-3′, and R1513 Hypo: 5′-TGATCCTTCYGCAGGTTC-3′ [35,36]. The PCR conditions for Euplotes aspergilliformis and E. borealis were as follows: denaturation at 98 °C for 30 s, followed by 18 cycles of amplification (98 °C, 10 s, 69–52 °C touch down, 30 s; 72 °C, 1 min); another 18 cycles amplification (98 °C, 10 s, 51 °C, 30 s; 72 °C, 1 min); and a final extension step at 72 °C for 5 min. For E. verebkovi, DNA amplification was performed in a T100 Thermal Cycler (BioRad, Hercules, CA, USA) with the following program: 3 min at 95 °C; 39 cycles of 30 s at 95 °C, 1 min at 55 °C, and 2 min at 72 °C; and a final extension at 72 °C for 6 min. To improve sequence quality, subsequent processing differed between the studied species. For E. aspergilliformis and E. borealis, the PCR products were either directly sequenced or purified by TIANgel Midi Purification Kit (TIANGEN BIOTECH, Beijing, China), cloned using PMD 18-T vector cloning kit (Takara Biomedicals, Shiga, Japan), and a randomly selected clone was sequenced bidirectionally in Shanghai Sangon Biotechnology Company (Shanghai, China). For E. verebkovi, the amplicons were cloned using the InsTAclone PCR Cloning Kit (Fermentas, Carlsbad, CA, USA) according to the manufacturer’s protocol. Colonies were tested by PCR amplification using vector-specific M13 primers. PCR products were sequenced using Sanger sequencing by ABI310 (ABI Prism, Foster City, CA, USA). All sequences were generated using bidirectional sequencing. The quality of the obtained sequences can be observed through the corresponding diagram. Individual sequencing reactions were manually assembled into contigs, with careful reference to the diagram to ensure accuracy.
2.3. Phylogenetic Analyses
The newly obtained SSU rDNA gene sequence of three Euplotes species and those of 83 other euplotids downloaded from GenBank database were used for phylogeny analyses. Six discocephalids were used as outgroup taxa. Sequences were aligned using MAFFT version 7 server (https://mafft.cbrc.jp/alignment/server/ (accessed on 26 January 2026)). After alignment, the primer binding sites were trimmed manually by Bioedit v.7.2.5 [37], resulting in a final alignment including 2172 sites.
Maximum likelihood (ML) analysis was performed using RAxML-HPC2 on ACCESS v8.2.12 on the online server CIPRES Science Gateway (http://www.phylo.org/portal2/login!input.action (accessed on 26 January 2026)) with the GTRGAMMA model (Miller et al., 2012; Stamatakis, 2014) [38,39]. The reliability of internal branches was assessed using a nonparametric bootstrap method with 1000 replicates. Bayesian inference (BI) analyses were carried out using MrBayes on Access v3.2.x [40] on the CIPRES Science Gateway with GTR + I + G model selected by Akaike Information Criterion (AIC) in MrModeltest v2.2 [41]. Markov chain Monte Carlo (MCMC) simulations were run for 4,000,000 generations, with sampling every 100 generations; the first 10,000 trees were discarded as a burn-in. MEGA 11 v11.0.13 was used to visualize the tree topologies [42].
For interpretation of bootstrap values, we followed the protocol described by Vd’ačný and Rajter [43], that is, values ≥ 95 are considered to be high, 71 to 94 are moderate, 50 to 70 are low, and <50 have no support [44]. Bayesian posterior probabilities < 0.95 are considered as low and values ≥ 0.95 as high [45].
ZooBank Registration of present work: urn:lsid:zoobank.org:pub:C7711067-3EE8-40FF-B748-715C1AF1A025.
3. Results
3.1. Euplotes aspergilliformis sp. nov. (Figure 2A–M, Table 1)
Diagnosis: Freshwater species with a size of 60–85 × 40–60 μm in vivo; outline generally oval-shaped, buccal field about 70% of body length with approximately 34 adoral membranelles; nine frontoventral cirri; two marginal and two caudal cirri; five transverse cirri; consistently eight dorsal kineties with about 15 dikinetids in mid-dorsal kinety; macronucleus C-shaped; double-eurystomus type silverline system.
Locality: Water sample from the Hulan River (45°57′2″ N, 126°34′46″ E), Harbin, China, where the temperature was about 3 °C.
Materials: The holotype slide (registration number: LH2024112203-1) with protargol-stained holotype specimens (circled with black ink on the back of slide; Figure 2B,C,I,J), three paratype slides with protargol-stained specimens (Nos. LH2024112203-2, LH2024112203-5, LH2024112203-6), and one paratype slide with dry silver-nitrate-stained specimens (No. LH2024112203-11) are deposited in the Laboratory of Protozoology, Harbin Normal University.
Morphology and infraciliature of Euplotes aspergilliformis sp. nov. in vivo (A,D–G,L,M), after protargol staining (B,C,I,J) and dry silver nitrate (H,K) staining. (A,D) Ventral views of the representative individual. (B,C,I,J) Ventral (B,I) and dorsal (C,J) views of the holotype specimen, showing the infraciliature and nuclear apparatus. (E,F) Ventral views of different individuals; arrowheads indicate contractile vacuole. (G) Showing the ridges on ventral sides. (H,K) Silverline system on dorsal side. (L) Showing the endoplasm of a cell. (M) Posterior part of an individual; arrowheads show the caudal cirri. Scale bars: 50 μm.
Etymology: The species-group name aspergilliformis (meaning brush-shaped) refers to the brush-like caudal cirri of the living cells.
ZooBank registration of E. aspergilliformis: urn: lsid: zoobank. org: act: urn:lsid:zoobank.org:act:DC2FC703-8892-40F9-9BF8-5DC6761607A7.
Morphological Description:
Cells measuring 60–85 × 40–60 μm in vivo. Both left and right margins are convex, anterior and posterior ends are smoothly rounded (Figure 2A,D–G). Three conspicuous ventral ridges extending posteriorly to transverse cirri with some shorter ridges between them (Figure 2A,G). Cytoplasm is colorless, with numerous food vacuoles of varying sizes in the center, making the cell somewhat opaque in contrast to the highly transparent margin (Figure 2D,E,L). The contractile vacuole is about 11 μm in diameter, located posterior to the rightmost transverse cirrus (Figure 2A,E).
The buccal field is approximately 70% of the body length, composed of 32–37 membranelles, with bases up to 19 μm long (Figure 2A,B,I). Invariably, there are nine frontoventral cirri (cilia about 28 μm long); five transverse cirri, cilia all about 33 μm long; two marginal cirri (cilia about 20 μm long); and two caudal cirri (cilia about 16 μm long) with forked distal ends (Figure 2A,B,I,M). There are eight dorsal kineties, with 13–17 dikinetids in the mid-dorsal kinety and about 12 basal bodies in the leftmost dorsal kinety (Figure 2C,J). It has a dorsal silverline system of double-eurystomus type (Figure 2H,K).
3.2. Euplotes borealis sp. nov. (Figure 3A–M, Table 1)
Diagnosis: Small freshwater Euplotes, in vivo about 45–55 × 25–35 μm; generally oval-shaped; buccal field about 75% of body length with approximately 29 adoral membranelles; nine frontoventral cirri; one marginal and two caudal cirri; five transverse cirri; consistently nine dorsolateral kineties with about nine dikinetids in mid-dorsal kinety; macronucleus C-shaped; dorsal silverline system double-eurystomus type.
Locality: Freshwater from a small pond near Anda People’s Court (45°24′43″ N, 125°15′5″ E), Heilongjiang, China, where the temperature was about 11.8 °C.
Materials: One slide (No. LH2025040714-1) with protargol-stained holotype specimens (circled with black ink in the back of slide; Figure 3B,C,I,J), four paratype slides (Nos. LH2025040714-2, LH2025040714-3, LH2025040714-4, LH2025040714-5) with protargol-stained holotype specimens, and one paratype slide (No. LH2025040714-17) with the dry silver-nitrate-stained specimens are deposited in the Laboratory of Protozoology, Harbin Normal University.
Morphology and infraciliature of Euplotes borealis sp. nov. in vivo (A,E–H,K–M), after protargol staining (B,C,I,J) and dry silver nitrate (D) staining. (A,E) Ventral views of the representative individual. (B,C,I,J) Ventral (B,I) and dorsal (C,J) views of the holotype specimen, showing the infraciliature and unclear apparatus. (D) Silverline system on dorsal side. (F) Ventral view of a different individual; arrowheads indicate contractile vacuole. (G,H) Showing the ridges on ventral (G) and dorsal (H) sides. (K,L) Showing the endoplasm; arrowheads indicate the food vacuoles. (M) Sub-pellicular rosette-like structures around dorsal cilia; arrowheads indicate the basal positions of dorsal cilia. Scale bars: 30 μm.
Etymology: The species-group name borealis (meaning north) refers to the fact that this species was first described from a northern freshwater habitat (Heilongjiang Province, China).
ZooBank registration of Euplotes borealis: urn: lsid: zoobank. org: act: CD9D8E56-EA42-41AF-BD7F-00A6A4159CE1.
Morphological Description:
Cells in vivo are about 45–55 × 25–35 μm. Both the left and right margins are convex; the anterior end is narrowly rounded (Figure 3A,E–G). There are six or seven longitudinal ridges on the dorsal side (Figure 3H). About six ellipsoid granules surround each dorsal cilium, forming rosette rows along the dorsal kinety (Figure 3M). The cytoplasm is colorless, with opaque endoplasmic particles in the mid-body region (Figure 3E,F,K,L). The contractile vacuole is 10 μm in diameter, adjacent to the rightmost transverse cirrus, pulsating at intervals of about 30 s (Figure 3A,F).
The adoral zone is approximately 75% of the body length, composed of 25–33 membranelles, with bases up to 12 μm long (Figure 3A,B,I). Invariably, there are nine frontoventral cirri (cilia about 18 μm long); five transverse cirri, cilia all about 19 μm long; one marginal cirrus (cilia about 14 μm long); and two caudal cirri with cilia about 12 μm long (Figure 3A,B,I). There are nine dorsal kineties, with 8–10 dikinetids in the mid-dorsal kinety and about five basal bodies in the leftmost dorsal kinety (Figure 3C,J). It has a double-eurystomus type of dorsal silverline system (Figure 3D).
3.3. Euplotes verebkovi sp. nov. (Figure 4A–D, Table 1)
Diagnosis: Medium-sized freshwater Euplotes ciliates, in vivo about 40–54 × 22–31 µm. The cell shape is an elongated oval with rounded ends. The buccal field is about 2/3 of the cell length, with about 30 adoral membranelles. The macronucleus is C-shaped, and the micronucleus is spherical. There are seven conspicuous dorsal ridges and seven dorsal kineties, with eight or nine bristles in the central row. It has a dorsal silverline system of double-eurystomus type. There are ten frontoventral cirri, five transverse cirri, two caudal cirri and one marginal cirrus.
Locality: Private trout farm “Verebkovo” (57°44′40.8″ N; 27°34′08.1″ E) in the Pskov region, Russia.
Materials: Slides with silver nitrate-impregnated cells and Feulgen-stained cells of the E. verebkovi sp. nov. are available from the Zoological Institute RAS (Laboratory of Cellular and Molecular Protistology).
Morphology and infraciliature of Euplotes verebkovi sp. nov. in vivo (A), after Feulgen (B), and silver nitrate impregnation (C,D) staining. (A) Ventral view of the representative individual; arrows show the dorsal ridges. (B) Nuclear apparatus stained after Feulgen. (C,D) Dorsal (C) and ventral (D) views of representative specimens after silver nitrate impregnation. AM, adoral zone of membranelles; B, bristles; CC, caudal cirri; DK, dorsal kineties; FVC, frontoventral cirri; Ma, macronucleus; MC, marginal cirri; Mi, micronucleus; Pm, paroral membrane; TC, transverse cirri. Scale bars: 10 μm.
Etymology: The species name refers to the area where the sample was collected (trout farm “Verebkovo”).
ZooBank registration of Euplotes verebkovi: urn:lsid:zoobank.org:act:7944DF1C-DA95-489A-B0B1-078FC668E277.
Morphological Description:
Cells in vivo are about 40.3–54.0 µm in length and about 22.3–31.4 µm in width (average 47.6 ± 3.7 × 27.2 ± 2.8 µm), with a body length/width ratio of about 1.7–1.8. It generally has an elongated oval shape with rounded ends. The dorsal side is decorated with seven ridges (Figure 4A). The cytoplasm is colorless and highly transparent at the marginal area (Figure 4A). The contractile vacuole is not visible. The macronucleus is C-shaped with rounded ends (Figure 4B). The micronucleus is single, spherical (compact type), and usually localized in the cavity of the macronucleus (Figure 4B).
The adoral zone comprises 27–34 membranelles (Figure 4A,D). The ventral cirral pattern is stable, composed of invariably ten frontoventral cirri, five transverse cirri, one marginal cirrus, and two caudal cirri (Figure 4D). The dorsal surface bears seven kineties, with eight or nine bristles in the mid-dorsal row, the leftmost row consists of six to ten bristles and the rightmost row consists of five to eight bristles (Figure 4C). The dorsal argyrome is of the double-eurystomus type (Figure 4C).
3.4. SSU rDNA Sequences and Phylogenetic Analyses
The small subunit ribosomal RNA (SSU rRNA) gene sequences of Euplotes verebkovi, E. aspergilliformis, and E. borealis have been deposited in GenBank under the accession numbers PQ656668, PZ068345, and PZ068344, respectively. The length and GC contents are as follows: E. verebkovi (1996 bp, 43.24%), E. aspergilliformis (1790 bp, 42.79%), and E. borealis (1887 bp, 43.19%).
Phylogenetic trees were constructed based on the SSU rRNA gene sequences using both maximum likelihood (ML) and Bayesian inference (BI) methods, with a broad selection of taxa in the order Euplotida. The topologies of ML and BI trees were almost identical; thus, only the ML tree with supported values from both methods is presented here (Figure 5).
Consistent with previous studies, the four families of Euplotida are all monophyletic. It should be noted that the monophyly of Certesiidae could not be assessed, as only one species was included. The monophyly of the family Euplotidae received full support (100 ML/1.00 BI). With full support (100 ML/1.00 BI), the newly obtained E. aspergilliformis groups with E. patella (EF094964) and clusters tightly with E. octocarinatus (EF094963, 99 ML/1.00 BI). The newly sequenced E. verebkovi grouped with E. daqingensis with full support (100 ML/1.00 BI) and then formed a no-support clade (49 ML/0.91 BI) with E. elegans (DQ309868). Euplotes borealis (PZ068344) and Euplotes sp. (LN870038) are positioned outside this clade with moderate support (74 ML/0.99 BI).
4. Discussion
4.1. Euplotes aspergilliformis sp. nov.
Comparison with related congeners. Considering the double-eurystomus type silverline system, nine frontoventral and two marginal cirri, six Euplotes species should be compared with E. aspergilliformis sp. nov., namely, E. amieti Dragesco, 1970, E. paramieti Han et al., 2024, E. woodruffi Gaw, 1939, E. eurystomus (Wrzesniowski, 1870) Kahl, 1932, E. aediculatus Pierson, 1943, and E. finki Foissner, 1982 (Table 2).
Euplotes amieti can be clearly separated from E. aspergilliformis sp. nov. by its (1) triangular-shaped adoral zone (vs. nearly C-shaped), (2) 3-shaped macronucleus (vs. inverted C-shaped), (3) larger cell size (130–240 × 70–160 vs. 60–85 × 40–60), (4) higher number of adoral membranelles (52–70 vs. 32–37), (5) more dorsal kineties (12–15 vs. eight), and (6) more basal bodies in the middle kinety (18–32 vs. 13–17) [46,47,48,49,50].
Euplotes paramieti differs from E. aspergilliformis sp. nov. in (1) the appearance of the adoral zone (proximal ventral membranelles arranged in sigmoidal shape, vs. evenly bending, nearly C-shaped), (2) the shape of the macronucleus (3-shaped vs. inverted C-shaped), (3) cell size (180–220 × 110–155 vs. 60–85 × 40–60), (4) the number of adoral membranelles (63–93 vs. 32–37), (5) the number of dorsal kineties (12–13 vs. eight), and (6) the number of basal bodies in the middle kinety (24–37 vs. 13–17) [15].
Euplotes eurystomus can be clearly distinguished from E. aspergilliformis sp. nov. by (1) the shape of the adoral zone (conspicuous collar positioned at anterior end vs. evenly bending, nearly C-shaped), (2) the shape of the macronucleus (3-shaped vs. C-shaped), (3) longer cell (88–180 vs. 60–85), (4) higher number of adoral membranelles (44–65 vs. 32–37), and (5) basal bodies in the middle kinety (15–31 vs. 13–17) [11,22,51,52,53,54,55,56,57,58,59].
Euplotes finki differs from E. aspergilliformis sp. nov. in (1) the biotope (soil vs. freshwater), (2) the arrangement of transverse cirri (a gap between the left two and the remaining three divided them into two groups vs. a continuous group), (3) the shape of caudal cirri (absence vs. presence of a forked distal), (4) the number of adoral membranelles (19–22 vs. 32–37), (5) the number of dorsal kineties (seven vs. eight), and (6) the number of basal bodies in the middle kinety (about 10 vs. 13–17) [60].
Euplotes woodruffi differs from E. aspergilliformis sp. nov. in (1) the shape of the adoral zone (conspicuous collar positioned at anterior end vs. evenly bending, nearly C-shaped), (2) the shape of the macronucleus (T-shaped vs. C-shaped), (3) larger cell size (83–200 × 58–130 vs. 60–85 × 40–60), (4) greater number of adoral membranelles (40–99 vs. 32–37), (5) higher number of dorsal kineties (8–11, normally nine or ten vs. eight), and (6) greater number of basal bodies in the middle kinety (17–35 vs. 13–17) [22,48,49,56,59,61,62,63,64,65,66].
Euplotes aediculatus can be distinguished from E. aspergilliformis sp. nov. due to (1) the appearance of the adoral zone (triangular-shaped vs. evenly bending, nearly C-shaped), (2) the shape of the macronucleus (3-shaped vs. C-shaped), (3) larger cell size (105–170 × 60–120 vs. 60–85 × 40–60), (4) a higher number of adoral membranelles (40–70 vs. 32–37), and (5) a higher number of basal bodies in the middle kinety (18–30 vs. 13–17) [22,49,59,62,67,68,69,70,71].
Euplotes indica differs from E. aspergilliformis sp. nov. in (1) the location of marginal cirri (post-orally located, clearly separated from two caudal cirri vs. more subcaudally located, near the caudal cirri), (2) smaller body size in vivo (49–52 × 40–46 vs. 60–85 × 40–60), (3) lower number of adoral membranelles (20–25 vs. 32–37), and (4) lower number of dorsal kineties (seven vs. eight) [22].
4.2. Euplotes borealis sp. nov.
Comparison with related congeners. In terms of the double-eurystomus type silverline system, nine frontoventral cirri, and one marginal cirrus, seven Euplotes species are considered relevant for comparison with E. borealis sp. nov., namely E. mazeii Lian et al., 2023, E. dogieli Agamaliev, 1967, E. bisulcatus Kahl, 1932, E. affinis (Dujardin, 1841) Perty, 1852, E. foissneri Valbonesi et al., 2021, E. warreni Valbonesi et al., 2021, and E. nana Jones & Owen, 1974 (Table 3).
E. mazeii can be clearly distinguished from E. borealis sp. nov. by (1) the appearance of the adoral zone (evenly bending, nearly C-shaped vs. bent almost 90° at the posterior end), (2) a slightly lower number of adoral membranelles (18–26 vs. 25–33), and (3) a fewer number of dorsal kineties (seven vs. nine) [16].
Euplotes dogieli differs from E. borealis sp. nov. in the following respects: (1) body shape (right side rectilinear and left side convex vs. generally oval-shaped with both left and right side convex), (2) location of the marginal circus (more subcaudal, near the caudal cirri vs. post-oral, clearly separated from two caudal cirri), (3) body length (ca. 65 vs. 45–55), (4) number of adoral membranelles (35–38 vs. 25–33), (5) number of dorsal kineties (seven vs. nine) and (6) basal bodies in the middle kinety (ca. 13 vs. 8–10) [72].
Euplotes bisulcatus can be clearly distinguished from E. borealis sp. nov. by: (1) biotope (marine vs. freshwater), (2) the number of adoral membranelles (ca.17 vs. 25–33), (3) a fewer number of dorsal kineties (eight vs. nine), (4) a lower number of dikinetids in middle dorsal kinety (five to seven vs. 8–10), and (5) a lower number of dikinetids in the left dorsal kinety (ca. two vs. four to six) [73,74].
Euplotes affinis was first described by Dujardin from a freshwater population. Subsequently, Kahl reported E. affinis forma tricirratus, which is characterized by a smaller body size and the presence of only one marginal cirrus. Later, Curds provided a redescription of E. affinis based on a British population, which closely resembled the population reported by Kahl and supplemented the illustrative diagrams of the silverline system. Given its possession of nine frontoventral, one marginal cirri, and a double-eurystomus type silverline system, E. affinis forma tricirratus also should be compared with E. borealis sp. nov. Euplotes affinis forma tricirratus can be clearly separated from E. borealis sp. nov. by (1) the shape of the macronucleus (3-shaped vs. C-shaped), (2) a lower number of adoral membranelles (18–20 vs. 25–33), and (3) a lower number of dorsal kineties (seven vs. nine) [73,75,76].
Euplotes foissneri can be clearly distinguished from E. borealis sp. nov. by: (1) the arrangement of caudal cirri (aggregated vs. separated), (2) the biotope (brackish water vs. freshwater), (3) the number of dorsal kineties (eight vs. nine), and (4) dikinetids in the middle dorsal kinety (12–16 vs. 8–10) [19].
Euplotes warreni can be separated from E. borealis sp. nov. due to its (1) body shape (D-shape vs. ovoid), (2) biotope (marine vs. freshwater), (3) a lower number of adoral membranelles (23–25 vs. 25–33), and (4) a lower number of dorsal kineties (six vs. nine) [19].
Euplotes nana Jone & Owen, 1974 can be distinguished from E. borealis sp. nov. by: (1) the position of the contractive vacuole (situated medially near the posterior body end vs. located posteriorly near right body margin), (2) the biotope (marine vs. freshwater), (3) the location of the marginal circus (more subcaudally, near the caudal cirri vs. post-orally located, clearly separated from the caudal cirri), (4) a lower number of dorsal kineties (ca. eight vs. nine), and (5) a lower number of dikinetids in the middle dorsal kinety (ca. five vs. 8–10) [77].
4.3. Euplotes verebkovi sp. nov.
Comparison with related congeners. In terms of morphological and molecular data, five Euplotes species are considered relevant for comparison with E. verebkovi sp. nov., namely E. elegans Kahl, 1932, E. nobilii Valbonesi & Luporini, 1990, E. qatarensis Fotedar et al., 2016, E. curdsi Syberg-Olsen et al., 2016, and E. daqingensis Lian et al., 2026 (Table 4).
Euplotes elegans can be separated from E. verebkovi by its larger body size (87–118 × 43–59 vs. 40–54 × 22–31), more membranelles in the adoral zone (47–64 vs. 27–34), more dorsolateral kineties (nine or ten vs. seven), more bristles in the central row (15–20 vs. eight or nine), and the type of habitat (marine vs. freshwater). Also, the authors mention that E. elegans has nine frontoventral cirri and one reduced cirrus. Euplotes verebkovi sp. nov. has ten normal-sized frontoventral cirri [9].
Euplotes nobilii differs from E. verebkovi sp. nov. in the cell shape (spindle-shaped vs. elongated oval), number of membranelles in the adoral zone (18–22 vs. 27–34), dargyrome type (double-patella vs. double-eurystomus), number of dorsal ridges (six vs. seven), number of dorsolateral kineties (eight vs. seven), and the type of habitat (marine vs. freshwater) [29].
The newly described species can be distinguished from Euplotes qatarensis based on the following differences: number of dorsolateral kineties (seven vs. 10), number of marginal cirri (one vs. two), and habitat type (freshwater vs. marine) [78].
Euplotes curdsi can be separated from E. verebkovi sp. nov. in the number of dorsal ridges (five or six vs. seven), the number of bristles in the central row (10–12 vs. eight or nine), the number of marginal cirri (two vs. one), and the type of habitat (brackish and marine vs. freshwater) [79].
Euplotes daqingensis, discovered by Lian et al., despite its close phylogenetic position to E. verebkovi sp. nov., has a number of significant morphological differences. Firstly, the number of frontoventral cirri (nine vs. ten). Secondly, the number of dorsal kineties (eight vs. seven) and the number of dorsal ridges (five vs. seven) [80].
4.4. Phylogenetic Analyses
The newly sequenced Euplotes aspergilliformis is closely related to E. patella and E. octocarinatus in our ML and BI trees; this corresponds well with the similar morphological characteristics between these three species. The sequence of E. aspergilliformis differs in only two nucleotides from the sequence of E. patella (compared with EF094964, isolated from Japan rather than the type locality) [81]. Meanwhile, E. patella and E. aspergilliformis are very similar in cell size and shape, biotope, number of adoral membranelles, the shape of the adoral zone, and the general infraciliature on the ventral side. The most significant difference between E. aspergilliformis and E. patella is the number of dorsal kineties (eight in E. aspergilliformis vs. nine in E. patella) and the type of silverline system (double-eurystomus type in E. aspergilliformis vs. double-patella type in E. patella), which can be detected only after protargol impregnation and silver nitrate staining. Furthermore, no morphological information is available regarding the sequence of E. patella (EF094964), so misidentification cannot be excluded. In the meantime, E. aspergilliformis also differs from E. octocarinatus in that the new species has an evenly bending adoral zone (vs. nearly triangular in shape) and a double-eurystomus type silverline system (vs. double-patella type), and the 19 bp sequence differences between them further support the separation of these two species.
Euplotes borealis is closely related to E. elegans, E. daqingensis, E. verebkovi, and Euplotes sp. (LN870038) in both trees. Among these species, the newly obtained E. borealis is most closely related to Euplotes sp. (LN870038), sharing 99.57% sequence similarity with only 8 nucleotide differences. Since neither a detailed description of a living organism nor morphometric data was reported for the sequence of Euplotes sp. (LN870038), their conspecificity cannot be excluded. In contrast, the remaining species show distinct morphological differences both in vivo and after protargol impregnation, as well as sequence variations exceeding 23 bp, thereby supporting the identification of the new species.
The new species Euplotes verebkovi in the present study shows high sequence similarity to E. daqingensis, with a genetic divergence of only four nucleotides. Despite this close genetic relationship, they are considered distinct species due to clear morphological differences [80]. The other molecular closely related species, Euplotes elegans (DQ309868), exhibits an 18 bp nucleotide difference and clear morphological distinctions, that is, the presence of nine (vs. ten) frontoventral cirri and only one (vs. two) marginal cirrus. Therefore, the phylogenetic analyses further support the establishment of Euplotes verebkovi as a new and distinct species.
Recently, many studies have inferred the evolutionary trajectories among Euplotes species using various morphological characteristics and genetic markers [82,83]. However, the key morphological evolutionary lineages remain to be clearly identified. Currently accepted morphological evolutionary clues include the pattern of the silverline system, habitat, and infraciliature. In the present studies, all three species examined are freshwater Euplotes possessing double-eurystomus silverline type; nevertheless, only E. borealis and E. verebkovi exhibit a close phylogenetic relationship. This finding seems consistent with the fact that these two species have only one marginal cirrus. However, other species with one marginal cirrus are scattered throughout the phylogenetic tree, indicating that this morphological appearance alone is not evolutionarily informative. Therefore, a single morphological feature is not a reliable indicator for reconstructing the evolutionary pathway of the genus Euplotes. Further attempts incorporating additional evidence may be necessary in future research.
5. Conclusions
As a result of historical research deficiencies, our understanding of euplotid ciliate diversity in high-latitude regions still lags considerably behind that of mid- to low-latitude areas. Although the scientific significance of these regions has now been increasingly recognized, filling this knowledge void will require sustained and long-term efforts. The present study contributes to this ongoing endeavor by providing evidence for the underestimated species diversity of protists in high-latitude habitats and underscoring the necessity for further exploration in these extreme environments.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Song W. Wilbert N. Chen Z. Shi X. Considerations on the systematic position of Uronychia and related euplotids based on the data of ontogeny and 18S r RNA gene sequence analyses, with morphogenetic redescription of Uronychia setigera Calkins, 1902 (Ciliophora: Euplotida)Acta Protozool.200443313328
- 2Shen Z. Yi Z. Warren A. The morphology, ontogeny, and small subunit r RNA gene sequence analysis of Diophrys parappendiculata n. sp. (Protozoa, Ciliophora, Euplotida), a new marine ciliate from coastal waters of southern China J. Eukaryot. Microbiol.20115824224810.1111/j.1550-7408.2011.00536.x 21449957 · doi ↗ · pubmed ↗
- 3Shen Z. Huang J. Lin X. Yi Z. Li J. Song W. Morphological and molecular characterization of Aspidisca hongkongensis spec. nov. (Ciliophora, Euplotida) from the South China Sea Eur. J. Protistol.20104620421110.1016/j.ejop.2010.01.00420303717 · doi ↗ · pubmed ↗
- 4Park M. Kim S. Min G. First record of two Euplotes ciliates (Ciliophora: Spirotrichea: Euplotida) from Korea Korean J. Syst. Zool.201026212710.5635/KJSZ.2010.26.1.021 · doi ↗
- 5Modeo L. Petroni G. Lobban C.S. Verni F. Vannini C. Morphological, ultrastructural, and molecular characterization of Euplotidium rosati n. sp. (Ciliophora, Euplotida) from Guam J. Eukaryot. Microbiol.201360253610.1111/jeu.1200323194274 · doi ↗ · pubmed ↗
- 6La Terza A. Papa G. Miceli C. Luporini P. Divergence between two Antarctic species of the ciliate Euplotes, E. focardii and E. nobilii, in the expression of heat-shock protein 70 genes Mol. Ecol.2001101061106710.1046/j.1365-294X.2001.01242.x 11348511 · doi ↗ · pubmed ↗
- 7Kwon C. Kang Y. Shin M. Two newly recorded estuarine ciliates, Euplotes vannus and E. parawoodruffi (Ciliophora: Spirotrichea: Euplotida) from Korea Korean J. Syst. Zool.20072322923510.5635/KJSZ.2007.23.2.229 · doi ↗
- 8Kim E. Lee W.J. Redescriptions of Euplotes encysticus and E. rariseta (Protist: Ciliophora: Euplotida)J. Species Res.20198128135