Pathogenicity and Aggressiveness of Corticioid Basidiomycetes Associated with Stem and Branch Rot of Avocado
José Julio Rodríguez-Aguilar, Juan Mendoza-Churape, Erwin Saúl Navarrete-Saldaña, Yurixhi Atenea Raya-Montaño, Margarita Vargas-Sandoval

TL;DR
This study identifies corticioid fungi as key pathogens causing white rot in avocado trees, challenging previous assumptions about their role.
Contribution
The study demonstrates that corticioid basidiomycetes are primary pathogens in avocado white rot, not just secondary saprotrophs.
Findings
Dentocorticium portoricense showed the highest pathogenicity and virulence in avocado white rot.
Grammothele spp. caused slower, moderate disease progression compared to D. portoricense.
Phylogenetic analysis confirmed D. portoricense as a distinct pathogen while Grammothele remained unresolved at species level.
Abstract
Woody tissue diseases of avocado (Persea americana Mill. var. Hass) pose a major phytosanitary threat due to their chronic progression, late symptom expression, and severe impact on tree stability and productivity. Although white rot has traditionally been attributed to saprobic basidiomycetes, increasing evidence suggests corticioid fungi may act as facultative pathogens in agricultural systems. This study examined corticioid basidiomycetes associated with white rot in stems and branches of avocado in Michoacán, Mexico. Field surveys revealed consistent symptoms of structural weakening, branch dieback, and wood decay. Fungal isolates obtained from symptomatic tissues and sporomes were characterized morphologically and identified through ITS-based phylogenetic analyses. Representative isolates of Grammothele spp. and Dentocorticium portoricense were evaluated in pathogenicity assays…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3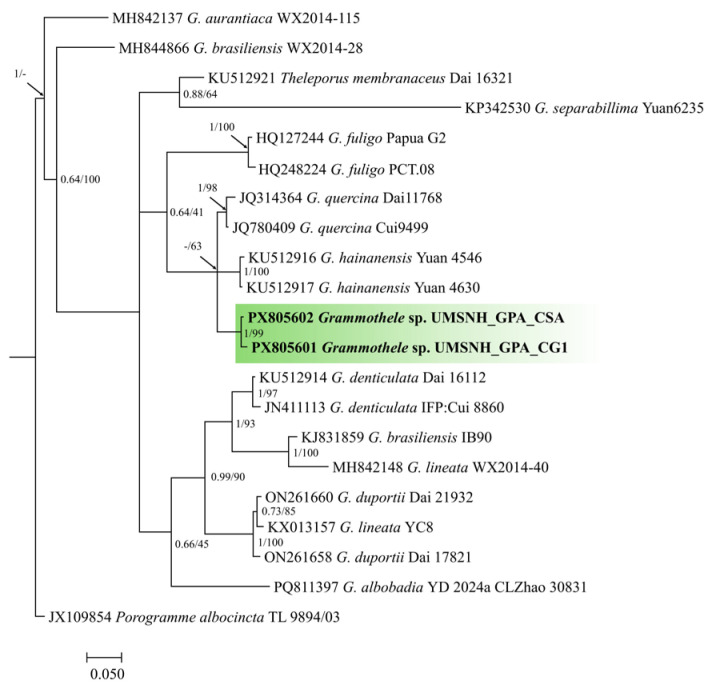 Figure 4
Figure 4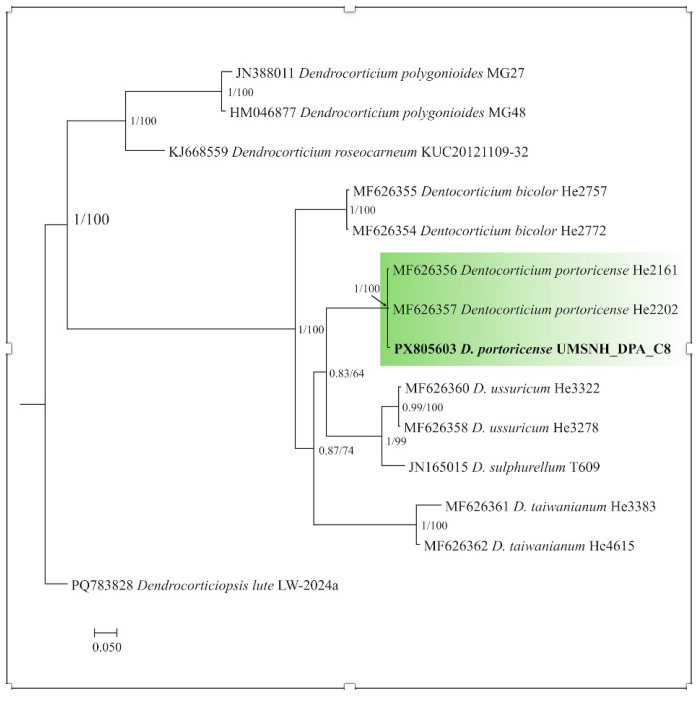 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8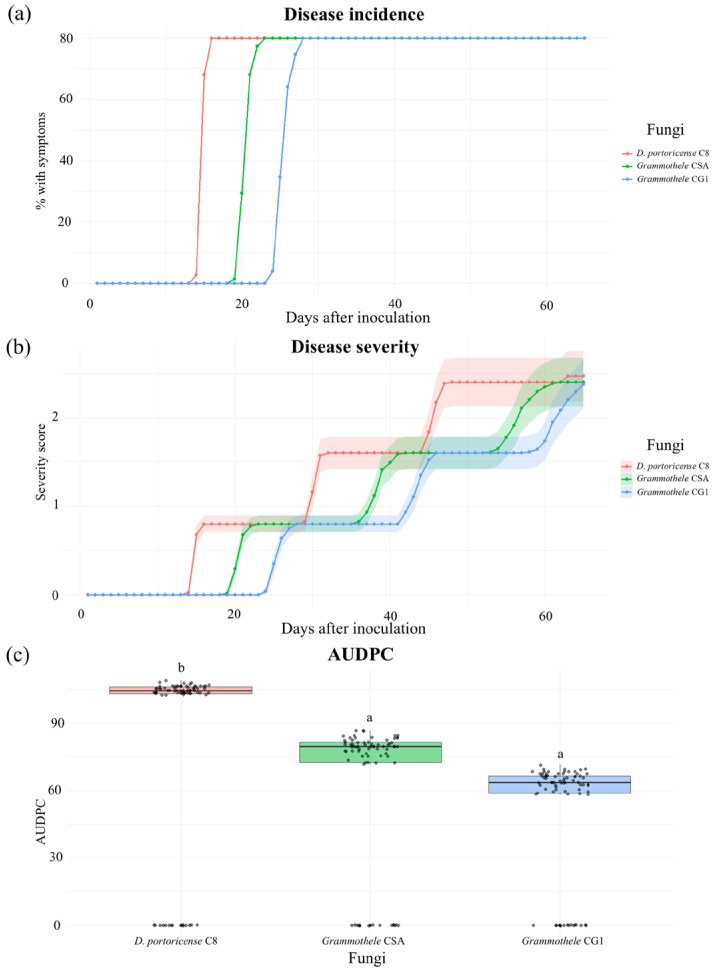 Figure 9
Figure 9- —Universidad Michoacana de San Nicolás de Hidalgo (UMSNH)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Plant Pathogens and Fungal Diseases · Cocoa and Sweet Potato Agronomy
1. Introduction
Stem and branch diseases represent one of the major phytosanitary constraints in avocado (Persea americana) production systems, as they compromise vascular integrity, structural stability, and long-term tree productivity [1,2]. Symptoms affecting branches and trunks commonly include cankers, tissue necrosis, wood decay, and progressive wilting, leading to the loss of productive limbs and, in severe cases, the death of the entire tree [3,4]. Traditionally, these symptoms have been mainly associated with a limited group of phytopathogenic fungi, particularly members of the family Botryosphaeriaceae, as well as species of Colletotrichum, Fusarium, and other ascomycetes commonly linked to trunk and branch diseases in avocado [5,6].
However, in recent years, members of Basidiomycota have been increasingly recognized as playing a relevant role in stem and branch diseases, contrary to previous assumptions [7,8]. Within this phylum, corticioid fungi constitute a morphologically and ecologically diverse group characterized by resupinate basidiomata and a close association with woody substrates. Nevertheless, they have been scarcely studied from a phytopathological perspective [9]. For decades, these organisms were regarded exclusively as saprophytes, restricted to the decomposition of dead wood and organic residues. This historical perception led to their systematic exclusion from phytopathological studies and from routine disease diagnostics in agricultural crops [10,11].
Nonetheless, several studies have demonstrated that certain corticioid fungi are capable of colonizing living woody tissues, causing structural degradation, vascular dysfunction, and tissue necrosis in a range of host plants [7,12]. These findings challenge the traditional view of corticioid fungi as strictly saprophytic organisms and suggest a broader ecological amplitude that includes facultative pathogenic behavior. Despite this evidence, in agricultural systems these fungi are still commonly interpreted as secondary or opportunistic colonizers, particularly when isolated from infected or decaying tissues [13].
As a consequence, the pathogenic potential, epidemiological relevance, and contribution of corticioid fungi to the severity of stem and branch diseases in woody crops such as avocado remain poorly documented. This lack of recognition limits diagnostic accuracy and hinders the development of effective phytosanitary management strategies, particularly in complex pathosystems where multiple fungal agents coexist [4,14].
Recent observations in avocado orchards exhibiting symptoms of stem and branch rot have revealed the recurrent presence of corticioid fungi associated with colonized and degraded tissues. Preliminary evaluations suggest that some of these fungi are capable of actively colonizing healthy tissues and inducing disease symptoms under favorable conditions; however, systematic studies assessing their pathogenicity and aggressiveness, supported by quantitative analyses of disease progression, are still limited [15,16].
In this context, the objective of the present study was to characterize corticioid fungi associated with stem and branch rot symptoms in avocado and to evaluate their pathogenicity under controlled conditions. Disease development was quantified using incidence, severity, and the area under the disease progress curve (AUDPC), allowing a comparative assessment of isolate aggressiveness over time. The results obtained contribute to a better understanding of the role of corticioid fungi in stem and branch diseases of avocado and highlight their relevance as emerging or previously underestimated pathogens in woody crop systems.
2. Materials and Methods
2.1. Study Area and Field Sampling
Sampling was conducted in commercial avocado orchards located within the avocado-producing belt of Michoacán State, Mexico (Table 1). Trees exhibiting visible symptoms of stem and branch diseases were selected, including white rot, cortical necrosis, cankers, branch dieback, and the presence of corticioid-type basidiomata. Sampling was directed toward symptomatic trees, following methodologies commonly employed in studies of woody tissue diseases and lignicolous fungi [17].
Six representative municipalities within the region were surveyed (Table 1), with two to four orchards visited per municipality. In each orchard, three to six affected trees were selected, resulting in a total of 72 sampled trees and approximately 95 woody tissue samples and associated basidiomata.
2.2. Isolation and Establishment of Pure Cultures
Woody tissue samples were surface-disinfested by immersion in 70% ethanol for 30 s, followed by immersion in 1% sodium hypochlorite for 1 min, and rinsed three times with sterile distilled water. Subsequently, 5 mm tissue fragments were plated onto potato dextrose agar (PDA) and incubated at 25 ± 2 °C for 36–48 h. Isolates exhibiting morphological characteristics consistent with corticioid fungi were subcultured through hyphal tip transfer until pure cultures were obtained.
2.3. DNA Extraction and Identification Through Phylogenetic Reconstruction
Genomic DNA was extracted from mycelium grown for 10 days using the MLO protocol described by Osuji et al. [18], combined with 2% CTAB. DNA quality and concentration were assessed spectrophotometrically using a NanoDrop™ instrument (Thermo Scientific, Waltham, MA, USA). Amplification of the ribosomal DNA internal transcribed spacer (ITS) region was performed by PCR using the primers ITS5 and ITS4 [19]. Amplification products were verified by electrophoresis on 1.5% agarose gels. A PCR cleanup step was perfomed to remove remaining primers using ExoSAP and commercially sequenced. The resulting sequences were compared with those available in GenBank using the BLAST tool (BLAST+ 2.17.0) of the National Center for Biotechnology Information (NCBI).
For phylogenetic analyses, a reference sequence dataset was constructed using sequences retrieved from NCBI, with taxonomic nomenclature verified through the MycoBank online platform. Sequence alignments were performed using MAFFT v7 and manually edited. The best-fitting evolutionary model was selected using jModelTest v2.1.10. Phylogenetic analyses were conducted using Bayesian inference in MrBayes v3.2 and maximum likelihood in RAxMLGUI. Phylogenetic trees were visualized and edited using MEGA v7 and FigTree v1.4.5.
2.4. Strain Preservation and Inoculum Preparation
Fungal isolates were preserved on sterilized sorghum grains autoclaved at 121 °C (15 psi) for 2 h and stored at 4 °C until use. Inoculum was prepared by inoculating actively growing mycelial plugs (7-day-old cultures) onto hydrated sorghum grains, which were then incubated at 28 °C until complete colonization.
2.5. Pathogenicity Tests
Experimental avocado plants were grown under greenhouse conditions. All plants originated from seeds collected from a single mother tree and were subsequently grafted with scions obtained from a single Hass donor tree, in order to minimize genetic variability. At the time of inoculation, plants were 11 months old, showed uniform growth, and did not exhibit visible symptoms of trunk or branch diseases.
Artificial inoculations were conducted on both trunks and branches. Prior to inoculation, the bark surface was disinfected with 70% ethanol. Circular wounds approximately 5 mm in diameter and ~2 mm deep was made using a sterile cork borer, carefully removing the bark to expose the xylem tissue without causing extensive mechanical damage.
Inoculation sites on the trunk were established on the main stem, while branch inoculations were performed on secondary branches of comparable diameter. All wounds were made on healthy tissue, avoiding natural cracks, lenticels, or pre-existing injuries.
Agar plugs (5 mm diameter) taken from the actively growing margin of 7-day-old fungal cultures were placed mycelium-side down onto the exposed xylem. The inoculation sites were covered with sterile moist cotton and sealed with Parafilm^®^ to prevent desiccation and external contamination. Control plants were treated identically, using sterile PDA plugs.
Inoculated plants were maintained under greenhouse conditions and monitored periodically for symptom development. At the end of the evaluation period, tissues from inoculated sites were collected, and the fungi were re-isolated to fulfill Koch’s postulates.
Disease development was monitored every 48 h for 125 days after inoculation. At each evaluation, lesion size was quantified by measuring lesion length (L, cm) along the main longitudinal axis of the stem or branch and maximum lesion width (W, cm) perpendicular to L at the widest point, using a ruler or digital caliper.
Lesion area (A, cm^2^) was estimated using an elliptical approximation according to the formula:
where L corresponds to lesion length and W to maximum lesion width. This approach was used to provide a consistent and reproducible estimation of lesion expansion over time.
In addition to lesion measurements, fungal signs were recorded as qualitative observations throughout the evaluation period. The presence of visible mycelial growth was assessed both superficially on the bark and internally, after careful removal of the bark to expose the underlying xylem tissue. Internal colonization was identified by the presence of white to cream mycelial growth within discolored wood. These observations were documented photographically and used as supporting evidence of active fungal colonization, but not as primary criteria for severity scoring.
2.6. Disease Assessment and Statistical Analysis
Disease severity was assessed using an ordinal scale from 0 to 5, primarily based on lesion length and associated tissue alterations: 0 = no visible symptoms; 1 = very slight lesion (L ≤ 1.0 cm) limited to the wound margin, without evident xylem discoloration; 2 = mild lesion (L > 1.0–2.5 cm) with initial xylem discoloration; 3 = moderate lesion (L > 2.5–4.5 cm) with evident xylem browning around the inoculation point; 4 = severe lesion (L > 4.5–6.5 cm) with pronounced xylem discoloration extending beyond the inoculation site and incipient tissue softening; 5 = very severe symptoms (L > 6.5–8.0 cm), characterized by extensive necrosis, marked xylem discoloration, and clear degradation of woody tissue, frequently accompanied by visible superficial and/or internal mycelial growth.
Disease progress over time was summarized by calculating the Area Under the Disease Progress Curve (AUDPC) for lesion length (and lesion area when applicable) using the trapezoidal integration method.
Aggressiveness data were analyzed using one-way analysis of variance (ANOVA), followed by Tukey’s test (p ≤ 0.05). Statistical analyses were performed using the R software (R version 4.5.2) environment, employing the agricolae package.
3. Results
3.1. Symptoms Observed in the Field
In the evaluated avocado orchards, trees exhibiting consistent symptoms of white rot affecting woody tissues were observed during the annual cycles from 2022 to 2025, primarily on stems and branches. Symptoms included structural weakening, mechanical fractures, bark fissures, and exposure of degraded wood. In both transverse and longitudinal sections, affected tissues exhibited a whitish to creamy-white discoloration, a marked loss of hardness, a fibrous to spongy texture. Structural alteration of the xylem, with diffuse boundaries between healthy and diseased tissues. No signs associated with brown rot, such as generalized darkening or cubical cracking of the wood, were observed.
In trees showing a higher degree of damage, branch dieback, reduced vegetative vigor, and foliar chlorosis were recorded. In addition, resupinate basidiomata firmly attached to the surface of woody tissues were detected on affected trunks and branches (Figure 1, Figure 2 and Figure 3).
3.2. Morphological Characterization of Basidiomata
Basidiomata collected in the field exhibited macromorphological characteristics consistent with corticioid fungi. Basidiomata were resupinate or effused, firmly adherent to the woody substrate, corky in texture when fresh and brittle when dry, with white to pale yellowish margins and smooth to slightly rough surfaces, ranging in color from white to light brown (Figure 1, Figure 2 and Figure 3).
3.2.1. Basidiomata of Grammothele sp.
Basidiomata attributed to Grammothele sp. corresponded to thin, resupinate basidiomata with a membranous to coriaceous consistency, forming irregular crusts on trunks and branches. The hymenophore exhibited a poroid to irpicoid surface, with shallow pores arranged in angular to sinuous patterns. Hymenial coloration ranged from white to creamy white, without dark brown tones. In longitudinal section, a thin subiculum and short tubes were observed, with the hymenium mainly restricted to the base of the tubes (Figure 1 and Figure 2).
3.2.2. Basidiomata of Dentocorticium portoricense
Basidiomata of Dentocorticium portoricense developed as extensive resupinate crusts on trunks and branches, mainly in areas exhibiting cortical damage. They displayed a white to creamy-white coloration and a well-defined odontioid hymenophore, with short, densely arranged tooth-like projections oriented perpendicular to the substrate. In adjacent tissues, xylem whitening and a loss of wood consistency were observed, with no evidence of brown rot symptoms (Figure 3).
3.3. Recovery of Fungal Isolates
From surveys conducted in 15 commercial orchards within the avocado-producing belt of Michoacán State, a total of 28 pure fungal isolates were obtained from woody tissues and associated basidiomata collected from trees exhibiting white rot symptoms. All isolates showed active mycelial growth on PDA medium, with noticeable variation in growth rate, texture, and mycelial coloration, allowing preliminary differentiation into distinct morphotypes.
Based on the morphological diversity observed both in the field and in culture, ten representative isolates were selected for the first experimental block. These isolates originated from different municipalities within the region (Ziracuaretiro, Salvador Escalante, Puruarán, Parácuaro, Tancítaro, and Uruapan), in order to capture the geographical and morphological variability of the recovered fungi. Isolates associated with the corticioid morphological group were selected based on their delimitation using the BLAST tool (BLAST+ 2.17.0) of the NCBI, retaining three isolates (CSA, CG1, and C8) for identification through phylogenetic reconstruction (Figure 4 and Figure 5) and for subsequent pathogenicity (Figure 6, Figure 7 and Figure 8) and aggressiveness assays (Figure 9).
3.4. Morphological Characterization of Isolates in Culture
On PDA medium, the selected isolates (CSA, CG1, and C8) exhibited slow to moderate growth, with predominantly white to creamy-white mycelium in isolates CSA and CG1 (Figure 6 and Figure 7), and in some cases light brown tones in isolate C8 (Figure 6). Colonies showed a cottony, velvety, or slightly flattened texture and were firmly adherent to the medium. Colony margins were regular to slightly irregular, with no evident production of diffusible pigments or exudates. Micromorphological analysis revealed hyaline, septate hyphae with clearly visible clamp connections in most isolates. Hyphae exhibited thin to moderately thickened walls, frequent branching, and variable diameters. No anamorphic structures or conidial production were observed under in vitro conditions. In some isolates, generative and skeletal hyphae were identified, suggesting a dimitic hyphal system, characteristic of white-rot-associated basidiomycetes.
In Grammothele isolates, colonies developed on culture media with uniform radial growth, white to creamy-white cottony mycelium (Figure 6a), and concentric growth patterns (Figure 7a,b). No macroscopic reproductive structures were observed during the evaluation period.
In the case of Dentocorticium portoricense, colonies grown in culture exhibited dense white mycelium, with homogeneous growth and continuous radial expansion, uniformly covering the surface of the culture medium (Figure 8a).
3.5. Identification Through Phylogenetic Reconstruction
Molecular analysis based on the ITS region enabled the identification of three corticioid fungal isolates: two belonging to the genus Grammothele (CG1 and CSA) and one to the genus Dentocorticium (C8). In the phylogenetic analysis of Grammothele, both isolates clustered within a well-supported clade (PP/BS = 1.0/99), clearly differentiated from the outgroup Porogramme albocincta (Figure 4).
In the case of Dentocorticium, isolate C8 was consistently grouped within the clade of Dentocorticium portoricense, showing maximum support in both Bayesian inference and maximum likelihood analyses (PP/BS = 1.0/100), and was clearly separated from other lineages of the genus included in the analysis (Figure 5).
3.6. Pathogenicity Test Results
Inoculated avocado seedlings developed the first symptoms at 82 days post inoculation, initially manifested as foliar chlorosis and progressive apical necrosis. Subsequently, from 125 days onward, creamy-white mycelial growth with a corky appearance was observed on the substrate, collar region, and cotyledons of the inoculated plants.
Pathogenicity assays conducted on avocado seedlings inoculated with corticioid fungi revealed the progressive development of foliar and stem symptoms, alterations of woody tissues, and the presence of signs associated with mycelial growth, both under in vitro conditions and in plants inoculated under controlled conditions (Figure 6, Figure 7 and Figure 8).
In seedlings inoculated with Grammothele, initial symptoms appeared as irregular chlorotic mottling distributed across the leaf lamina in both young and mature leaves (Figure 6c,d). As the evaluation period progressed, these symptoms advanced to partial foliar wilting (Figure 6e,g), characterized by loss of turgor and leaf curvature, followed by foliar necrosis, mainly affecting leaf apices and margins (Figure 6f,g and Figure 7d). At advanced stages, shoot dieback was observed, with progressive desiccation of lateral branches and a visible reduction in seedling vegetative vigor (Figure 6f and Figure 7e).
At the stem level, plants inoculated with Grammothele exhibited localized signs characterized by the presence of visible resupinate basidiomata on affected stem areas (Figure 6h). Longitudinal stem sections revealed alterations in woody tissues, including loss of xylem consistency and changes in internal coloration, without evidence of generalized darkening or cubical cracking of the wood.
In plants inoculated with isolate C8, more severe foliar symptoms were observed, beginning with pronounced chlorotic mottling and generalized chlorosis of the leaf lamina (Figure 6b,c). Subsequently, advanced wilting was recorded, characterized by collapse of foliar tissues, followed by branch necrosis, foliage desiccation, and evident shoot dieback (Figure 6d,e). In addition, clear signs of the pathogen were detected on stems inoculated with D. portoricense, including abundant external colonization characterized by firmly adherent mycelial growth on the cortical surface, particularly around the inoculation point (Figure 6f). Longitudinal stem sections revealed whitening of woody tissues, structural alteration of the xylem, and a marked loss of tissue consistency, evidencing progressive wood degradation associated with fungal colonization (Figure 6g).
Taken together, Figure 6, Figure 7 and Figure 8 visually document the sequence of symptom onset and progression in leaves, stems, and woody tissues, as well as the presence of mycelial signs, thereby supporting the quantitative results of incidence, severity, and aggressiveness obtained in the pathogenicity evaluations.
3.7. Incidence, Severity, and Aggressiveness
The isolate Dentocorticium portoricense (C8) exhibited an earlier and more advanced temporal incidence compared with the Grammothele isolates (CSA and CG1), which began to show symptoms from 20 days after inoculation (dai) (Figure 9a).
Regarding disease severity, D. portoricense C8 recorded the highest level of tissue damage (84.0 ± 3.96), followed by Grammothele CSA (63.8 ± 3.96) and Grammothele CG1 (51.8 ± 3.96) (Figure 9b).
Statistically significant differences were detected among the evaluated isolates in terms of aggressiveness (p < 0.05). Aggressiveness analysis, determined through the calculation of the area under the disease progress curve (AUDPC), showed a similar pattern consistent with incidence and severity, with significantly higher aggressiveness values for D. portoricense C8 compared with the Grammothele isolates (CSA and CG1) (Figure 9c).
4. Discussion
The evidence obtained in this study demonstrates that corticioid fungi associated with white rot of woody tissues act as active and differential pathogenic role in the progressive deterioration of stems and branches of Persea americana var. Hass within the avocado-producing belt of Michoacán, Mexico.
The consistent presence of structural signs and symptoms observed under field conditions, together with pathogenicity, confirmation under controlled conditions and phylogenetic support of the isolates, allows us to establish that these basidiomycetes should not be regarded solely as secondary saprophytes, but rather as functional phytopathogenic agents within intensive agricultural systems, particularly in avocado production systems.
In the evaluated orchards, the observed symptoms included structural weakening of branches, mechanical fractures, foliar chlorosis, shoot dieback, and exposure of whitish wood with a fibrous to spongy texture, which unequivocally correspond to white rot processes. This type of deterioration is characterized by the preferential degradation of lignin and the subsequent disorganization of xylem structural components, leading to a significant loss of mechanical resistance of the tree [7,11,20]. The absence of generalized darkening or cubical cracking in affected wood allowed brown rot processes to be ruled out and reinforced the association with ligninolytic basidiomycetes [21].
A relevant aspect observed under field conditions was the discrepancy between the severity of internal damage and the external expression of symptoms, a phenomenon widely documented in chronic woody tissue diseases. In such pathosystems, internal colonization and degradation may progress over prolonged periods before visible symptoms become evident in the aerial parts of the tree, which explains the delayed appearance of dieback, defoliation, or structural collapse in apparently functional trees [17]. In this context, the detection of resupinate basidiomata firmly attached to stems and branches acquires high diagnostic value, as it represents one of the few visible indicators of advanced colonization of woody tissues by corticioid fungi.
Morphological characterization of basidiomata and cultures allowed the recognition of features consistent with the genera Grammothele and Dentocorticium, including resupinate basidiomata firmly attached to the substrate, pale hymenial surfaces, and distinctive hymenial patterns. Nevertheless, the high morphological plasticity of corticioid fungi, together with the scarcity of highly diagnostic macroscopic characters, limits species-level identification based solely on morphology [22,23]. In this regard, the results of the present study reinforce the need to integrate molecular and phylogenetic approaches to achieve reliable taxonomic delimitation in phytopathological studies involving basidiomycetes with corticioid morphology.
Pathogenicity tests confirmed that the evaluated isolates are capable of inducing symptoms consistent with those observed under field conditions, strengthening the causal relationship between the isolated fungi and the woody tissue decline syndrome in avocado. However, quantitative analyses revealed significant differences in incidence, severity, and aggressiveness among the evaluated taxa. In particular, the isolate identified as Dentocorticium portoricense (C8) exhibited the highest severity and AUDPC values, indicating greater aggressiveness, understood as the combination of damage magnitude and the rate of disease progression over time [15]. In contrast, Grammothele sp. isolates (CSA and CG1) showed a slower and more moderate development of disease.
These differences in aggressiveness may be related to functional variation in the ligninolytic capacity of the isolates. White-rot basidiomycetes possess highly specialized enzymatic complexes, mainly composed of laccases, manganese-dependent peroxidases, and lignin peroxidases, which enable the efficient degradation of lignin and other structural polymers of the plant cell wall [24,25,26]. Although enzymatic activity was not directly evaluated in the present study, the observed severity and AUDPC patterns are consistent with a higher ligninolytic efficiency of isolate C8 compared with Grammothele isolates.
From a molecular perspective, ITS-based identification allowed robust phylogenetic assignment of isolate C8 as Dentocorticium portoricense, with maximum support in both Bayesian inference and maximum likelihood analyses. In the case of Grammothele, isolates CSA and CG1 clustered consistently within the genus but lacked unequivocal species-level resolution. This limitation is consistent with reports for several Basidiomycota groups, in which the ITS marker exhibits reduced interspecific variability, hindering fine taxonomic delimitation [27,28]. Consequently, the present results suggest that future studies aimed at species-level resolution in Grammothele should incorporate additional molecular markers or multilocus approaches, as well as more detailed morphofunctional characterization.
From a phytosanitary perspective, the identification of corticioid fungi with differing pathogenic aggressiveness has relevant implications for avocado orchard management. The chronic and slow-progressing nature of white rot hampers early detection, favoring scenarios in which internal damage advances unnoticed until severe structural failure or significant productivity losses occur. In this context, the recognition of D. portoricense as a highly aggressive agent and Grammothele spp. as more slowly progressing pathogens provides a scientific basis for the design of monitoring strategies, early diagnostic tools, and preventive management approaches targeting woody tissue diseases in avocado.
Overall, the results obtained support the notion that corticioid fungi, traditionally regarded as strictly saprophytic organisms, may modify their ecological behavior and act as facultative pathogens in intensive agricultural systems. The expansion of avocado cultivation in regions with high diversity of lignicolous fungi, combined with management practices and climatic factors that generate wounds in woody tissues, may favor this functional shift. Corticioid fungi as emerging or previously underestimated pathogens in avocado phytosanitary systems.
5. Conclusions
This study provides strong evidence that corticioid fungi associated with white rot of avocado woody tissues act as active pathogens rather than secondary saprophytes. Field observations, morphological and molecular analyses, and pathogenicity tests consistently confirmed their role in stem and branch decline. Among the evaluated taxa, Dentocorticium portoricense exhibited significantly higher incidence, severity, and aggressiveness compared to Grammothele spp., underscoring its relevance as a major phytopathogenic agent in avocado orchards. These findings highlight the importance of incorporating corticioid fungi into diagnostic protocols, monitoring systems, and management strategies for woody tissue diseases. Recognizing their pathogenic potential will improve early detection and strengthen phytosanitary measures, ultimately reducing structural damage and productivity losses in avocado production systems.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hardy G.E.S. Barrett S. Shearer B.L. The future of phosphite as a fungicide to control the soilborne plant pathogen Phytophthora cinnamomi in natural ecosystems Australas. Plant Pathol.20013013313910.1071/AP 01012 · doi ↗
- 2Shearer B.L. Crane C.E. Cochrane A. Quantification of the susceptibility of the native flora of the South-West Botanical Province, Western Australia, to Phytophthora cinnamomi Aust. J. Bot.20045243544310.1071/BT 03131 · doi ↗
- 3Eskalen A. Mc Donald V. Lynch S.C. First report of Neofusicoccum parvum causing branch canker of avocado in California Plant Dis.20139799310.1094/PDIS-11-12-1032-PDN 30754556 · doi ↗ · pubmed ↗
- 4Slippers B. Wingfield M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact Fungal Biol. Rev.2007219010610.1016/j.fbr.2007.06.002 · doi ↗
- 5Slippers B. Crous P.W. Denman S. Coutinho T.A. Wingfield B.D. Wingfield M.J. Combined multiple gene genealogies and phenotypic characters differentiate several species previously identified as Botryosphaeria dothidea Mycologia 2005968310110.1080/15572536.2005.1183300021148832 · doi ↗ · pubmed ↗
- 6Guarnaccia V. Crous P.W. Emerging citrus diseases in Europe caused by species of Diaporthe IMA Fungus 2017831733410.5598/imafungus.2017.08.02.0729242778 PMC 5729715 · doi ↗ · pubmed ↗
- 7Blanchette R.A. Delignification by wood-decay fungi Annu. Rev. Phytopathol.19912938140310.1146/annurev.py.29.090191.002121 · doi ↗
- 8OlsonÅ. Aerts A. Asiegbu F.O. Belbahri L. Bouzid O. Broberg A. Canbäck B. Coutinho P.M. Cullen D. Dalman K. Insight into trade-off between wood decay and parasitism from the genome of a fungal plant pathogen New Phytol.20121941001101310.1111/j.1469-8137.2012.04128.x 22463738 · doi ↗ · pubmed ↗