Arginine Decarboxylase Gene (adc) Is Essential for Vibrio anguillarum Virulence and Physiological Phenotypes
Binghong Liu, Haichuan Li, Jinyuan Che, Baolong Bao

TL;DR
This study shows that the adc gene is crucial for the virulence of Vibrio anguillarum, a harmful bacteria in aquaculture.
Contribution
The study identifies the adc gene as essential for L-arginine metabolism and virulence in Vibrio anguillarum.
Findings
The adc gene is critical for hemolytic activity and virulence in Vibrio anguillarum.
Δadc mutants show reduced hemolysis, biofilm formation, and anti-phagocytic ability.
Transcriptomic analysis reveals 704 differentially expressed genes linked to virulence pathways.
Abstract
Vibrio anguillarum is a major pathogenic bacterium causing vibriosis in aquatic animals, leading to substantial economic losses in the global aquaculture industry. Previous studies have indicated that L-arginine modulates the virulence of the pathogen, but the underlying molecular mechanisms remain elusive. The present study aimed to clarify the regulatory role of L-arginine metabolism in V. anguillarum virulence. We first evaluated the effects of L-arginine and its major metabolites (agmatine, putrescine, spermine) on the hemolytic activity of V. anguillarum. Results showed that L-arginine and its metabolites regulated hemolytic activity in a concentration-dependent biphasic manner, with agmatine exerting the most potent promoting effect. To identify the critical metabolic branch involved, four isogenic mutants were constructed targeting key genes in arginine metabolism (adc, astA,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
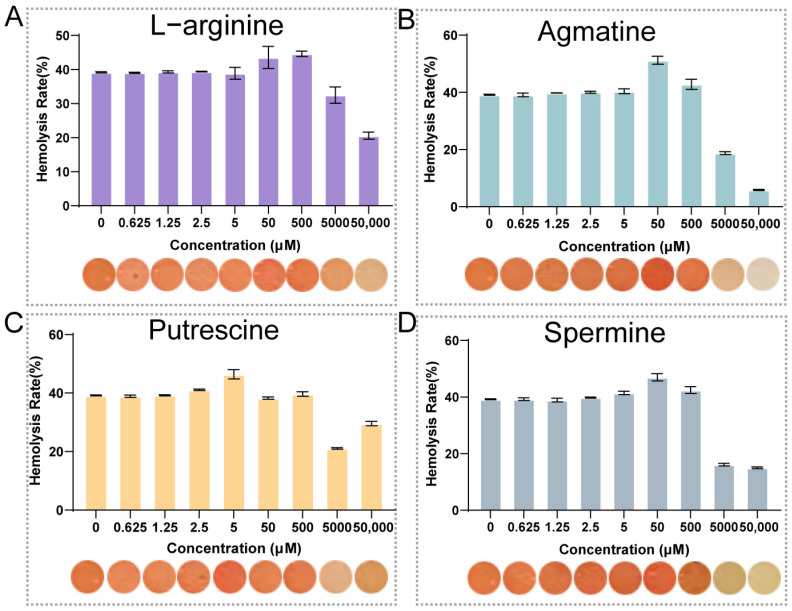 Figure 1
Figure 1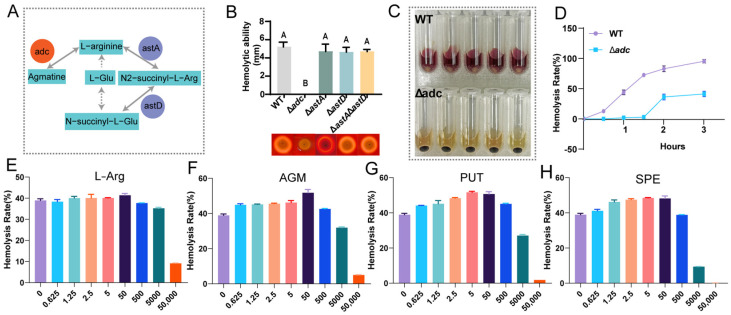 Figure 2
Figure 2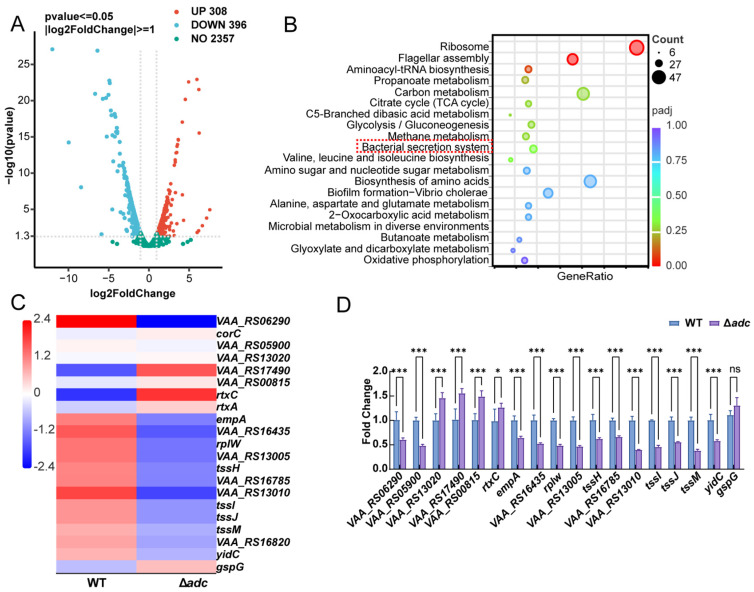 Figure 3
Figure 3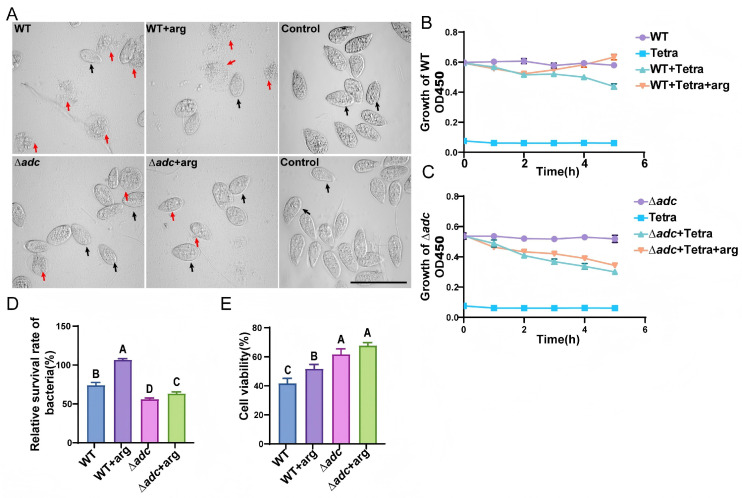 Figure 4
Figure 4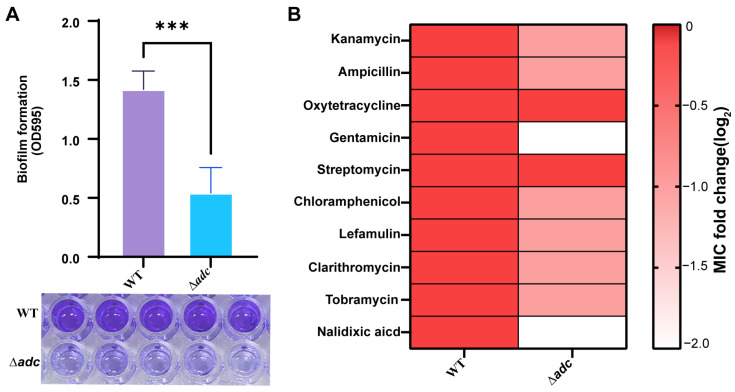 Figure 5
Figure 5- —National Key R&D Program of China
- —Fisheries High Quality Development Project of Fujian
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAquaculture disease management and microbiota · Vibrio bacteria research studies · Invertebrate Immune Response Mechanisms
1. Introduction
Vibrio anguillarum is a major bacterial pathogen in aquaculture and poses a serious threat to the global aquaculture industry. Infections caused by V. anguillarum can result in large-scale mortality of fish and crustaceans, leading to substantial economic losses worldwide [1,2]. The virulence of V. anguillarum is multifactorial and involves a broad array of virulence genes and determinants, including flagella, biofilm formation, iron acquisition systems, lipopolysaccharides (LPSs), and various extracellular products with proteolytic or hemolytic activities. Together, these factors facilitate bacterial colonization, infection, and disease progression within the host [3,4,5]. For instance, biofilm formation enhances bacterial tolerance to host immune defenses and antibiotic treatment, rendering infections more difficult to eradicate and often resulting in chronic infections and therapeutic failure
V. anguillarum is the etiological agent of vibriosis, an acute to subacute hemorrhagic septicemia that has been reported in a broad range of marine, brackish-water, and freshwater finfish. Susceptible and economically important hosts include salmonids (e.g., Atlantic salmon, rainbow trout), sea bass, turbot, striped bass, cod, eel, and red sea bream, among others, and outbreaks frequently cause rapid high mortality and major economic losses in both hatchery and grow-out systems [3,6].
In recent years, amino acid metabolism, particularly arginine (L-arginine) metabolism, has attracted increasing attention for its role in the regulation of bacterial virulence [7,8,9]. Arginine is a multifunctional amino acid that not only serves as a fundamental building block for protein synthesis but also plays crucial roles in microbial metabolism, survival, interspecies communication, and the expression of virulence traits [8,10]. Accumulating evidence indicates that arginine can significantly influence the pathogenic potential of a wide range of bacterial species [7,9].
Specifically, arginine-derived metabolites, such as the polyamines putrescine and agmatine, have been shown to be closely associated with bacterial virulence. Previous studies have demonstrated that microorganisms are capable of producing bioactive polyamines through complex biosynthetic pathways, and these molecules exert profound effects on host physiology and immune responses [11,12]. In Streptococcus pneumoniae, the SP_0916 gene encodes an arginine decarboxylase responsible for the production of agmatine, which is essential for capsular polysaccharide biosynthesis. The capsule is a critical virulence factor that enables S. pneumoniae to evade host immune surveillance [13]. Moreover, in Vibrio fluvialis, the arginine deiminase (ADI) pathway, under the regulation of ArgR, promotes bacterial survival under acidic conditions, highlighting the importance of arginine metabolism in bacterial environmental adaptation and virulence [14]. Notably, bacterial infection can also induce enhanced metabolic flux from arginine to spermidine in host cells, thereby suppressing NLRP3 inflammasome activation, further underscoring the pivotal role of arginine metabolism in host–pathogen interactions [15].
Despite the well-documented involvement of arginine and its metabolites in virulence regulation across diverse bacterial pathogens in humans, the role of arginine in V. anguillarum remains poorly understood. In particular, how arginine modulates its metabolic network to influence virulence and pathogenicity in V. anguillarum has not yet been systematically investigated. In aquaculture, arginine is routinely supplemented as a functional amino acid to support growth and immune competence. In parallel, the concept of prebiotics has expanded beyond non-digestible carbohydrates; ISAPP defines a prebiotic as “a substrate that is selectively utilized by host microorganisms conferring a health benefit” [16]. Emerging evidence suggests that some amino acids can act as “aminobiotics” with prebiotic-like effects by shaping the gut microbiota and its metabolites [17], and dietary L-arginine has been reported to alter intestinal microbiota profiles in fish (e.g., Nile tilapia) [18]. Given the increasing interest in prebiotics as antibiotic-sparing feed additives in aquaculture [19], it is important to clarify whether arginine supplementation could also influence opportunistic pathogens such as V. anguillarum, either directly through bacterial arginine metabolism or indirectly via microbiome-mediated interactions. Current knowledge gaps include the lack of mechanistic insight into the arginine metabolic network of V. anguillarum and its direct linkage to virulence-associated phenotypes, such as hemolysis and biofilm formation. Notably, the specific function and regulatory mechanisms of ADC, a key enzyme in arginine metabolism, during V. anguillarum pathogenesis remain largely unexplored.
This study aimed to systematically explore the regulatory role of L-arg and its metabolites in the hemolytic activity of V. anguillarum and clarify the underlying molecular mechanisms. We first investigated the effects of L-arg and its downstream metabolites (agmatine, putrescine, spermine) on the hemolytic activity of wild-type V. anguillarum. Then, we constructed four isogenic mutants targeting key genes in L-arg metabolism (∆adc, ∆astA, ∆astD, ∆astA∆astD) to identify the critical metabolic branch involved in hemolysis regulation. Additionally, we performed metabolite supplementation assays, transcriptomic analysis, and phenotypic validation to elucidate the regulatory role of the adc gene in V. anguillarum virulence. This study is the first to report the concentration-dependent biphasic regulation of V. anguillarum hemolysis by L-arg and its metabolites, providing novel insights into the molecular mechanisms of L-arg-mediated virulence regulation in aquatic pathogenic vibrios and a potential target for the prevention and control of V. anguillarum-associated vibriosis.
2. Materials and Methods
2.1. Strains, Media, and Experimental Animals
All bacterial strains and plasmids employed in this study are summarized in Table 1. The gene deletion mutant was generated and characterized in a Vibrio anguillarum background strain that was originally isolated from diseased Larimichthys crocea and is maintained in our laboratory collection. This strain was obtained previously from routine diagnostic samples and stored in our laboratory collection; no new animal sampling or bacterial isolation was performed and no identifiable patient data were involved, so ethics approval was not required under Shanghai Ocean University guidelines. Both the wild-type strain and its derivative mutants were routinely propagated in Luria–Bertani (LB) medium supplemented with 1% (w/v) NaCl at 30 °C with agitation at 150 rpm. For virulence assays, Tetrahymena, kindly provided by Prof. Shan Gao (Ocean University of China), was used as the infection model organism. Tetrahymena cultures were maintained in sterile SPP medium containing 2% proteose peptone, 0.1% yeast extract, 0.2% glucose, and 0.003% sequestrene, and incubated at 30 °C.
2.2. Construction of the adc, astA, astD, astAastD Deletion Mutant Strain
Four adc, astA, astD, astAastD gene deletion mutant strains (∆adc, ∆astA, ∆astD, ∆astAastD) were constructed via homologous recombination [20]. The primers used are listed in Table 2. The upstream and downstream flanking regions of adc, astA, astD, astAastD were amplified using the primers P1/P2 and P3/P4, respectively. These fragments were then ligated into the linearized suicide plasmid pSR47S, which had been digested with Sac I and Sal I, using the ClonExpress II One Step Cloning Kit (C112-01, Vazyme, Nanjing, China). The resulting plasmid was transformed into CC118 λpir, then conjugated into the wild-type (WT) strain. Mutants were selected on LB agar with kanamycin (Kan) and ampicillin (Amp), and screened on LB agar with 10% sucrose. Mutation was confirmed by PCR and sequencing using the primers T1/T2. Growth characteristics were evaluated by incubating cultures at 30 °C with shaking at 150 rpm until OD_600_ reached 1.0. The cultures were then diluted 1:100 and grown in a temperature-controlled incubator, with samples collected hourly.
2.3. Biofilm Formation
Biofilm formation ability was evaluated via a modified protocol. Briefly, 20 µL of overnight cultures of the wild-type (WT), ∆adc, ∆astA, ∆astD, and ∆astAastD strains were diluted 1:100 into fresh LB medium in 96-well plates, followed by incubation at 30 °C for 48 h. After incubation, the biofilms were gently washed to remove planktonic bacteria, stained with 0.1% crystal violet solution for 10 min, thoroughly rinsed with sterile water to eliminate excess stain, and then allowed to air dry. Subsequently, 100 µL of 95% ethanol was added to each well to dissolve the bound crystal violet, and the absorbance at 595 nm was measured using a microplate reader (Agilent, Cheadle, UK). All experiments were conducted in triplicate.
2.4. Hemolytic Activity Assay
All bacterial strains were grown overnight, after which 500 μL of bacterial culture was mixed with 50 μL of sterile, defibrinated sheep blood. L-arginine and its metabolic products, including agmatine, putrescine, and spermidine, were added to the mixtures at final concentrations of 0.625, 1.25, 2.5, 5, 50, 500, 5000, and 50,000 μM to generate hemolysis curves. The samples were incubated at 30 °C with shaking at 150 rpm for 3 h. Following incubation, the mixtures were centrifuged at 6000× g for 2 min, and 100 μL of the resulting supernatant was transferred to a 96-well microtiter plate. Absorbance was measured at 540 nm (OD_540_) using a microplate reader. Blank controls containing sterile, defibrinated sheep blood and the corresponding additives without bacterial inoculation were included, and background absorbance was subtracted accordingly. Each experiment was performed in triplicate. The hemolysis rate (%) was calculated according to the formula hemolysis rate (%) = [(A_540_,sample − A_540_,blank)/(A_540_,total − A_540_,blank)] × 100, where A_540_, sample represents the absorbance of samples incubated with bacterial culture and the indicated compounds; A_540_,blank represents the absorbance of sterile, defibrinated sheep blood with the corresponding additives but without bacterial inoculation; and A_540_,total represents the absorbance of sheep blood subjected to complete hemolysis.
To further evaluate hemolytic activity under solid conditions, bacterial cultures grown overnight were adjusted to the same cell density (OD_600_ = 1.0) using fresh LB medium. Aliquots (2 μL) of the standardized suspensions were carefully spotted onto Columbia blood agar plates supplemented with 5% (v/v) defibrinated sheep blood. The inoculated plates were incubated at 30 °C for 24 h. Hemolytic activity was assessed by visually examining the formation of transparent hemolytic zones surrounding the bacterial colonies. The diameters of hemolysis zones were measured using ImageJ 1.53, and hemolytic capacity was expressed as the mean zone diameter from three independent biological replicates.
2.5. Determination of Antibiotic Susceptibility
Antibiotic susceptibility of the strains was evaluated using kanamycin, 3ampicillin, spectinomycin, gentamicin, streptomycin, chloramphenicol, Rifampicin, clarithromycin, tobramycin and nalidixic acid. For each antibiotic, the half-maximal inhibitory concentration (MIC) was determined based on gradient treatments of the wild-type (WT) strain. Initially, all antibiotics were tested over a concentration range of 5–50 μg/mL with increments of 5 μg/mL. For kanamycin, gentamicin, chloramphenicol, rifampicin, and clarithromycin, tobramycin and tetracycline whose minimal inhibitory concentrations (MICs) were below 5 μg/mL, MIC values were further determined using a refined concentration gradient of 0.5–5 μg/mL with 0.5 μg/mL increments. In contrast, for ampicillin, whose MIC exceeded 50 μg/mL, an expanded concentration range of 55–155 μg/mL with 10 μg/mL increments was applied to determine the MIC. Overnight cultures of the WT and knockout strains were inoculated at 1% (v/v) into 96-well plates containing 200 μL of LB medium supplemented with the corresponding antibiotic concentrations. Cultures were incubated at 30 °C for 12 h, after which optical density was measured at 600 nm (OD_600_). All experiments were performed in triplicate.
2.6. Assessment of Strains’ Virulence Using Tetrahymena Model
The virulence of the ∆adc mutant was evaluated with a Tetrahymena infection model by measuring the relative survival of both bacteria and Tetrahymena after co-culture, as previously reported [21]. Briefly, Tetrahymena was grown in sterile SPP medium at 28 °C for 48 h, starting with an initial density of 10^3^ cells/mL. The cells were then collected by centrifugation at 2000× g for 10 min at 10 °C, washed twice with sterile SPP medium, and adjusted to a concentration of 1 × 10^5^ cells/mL.
Each bacterial strain was cultured for 12 h, harvested, washed twice with SPP medium, and resuspended to 3 × 10^9^ CFU/mL. Co-cultures were prepared by mixing V. anguillarum (3 × 10^9^ CFU/mL) with Tetrahymena (1 × 10^5^ cells/mL) at a ratio of 5000:1. The co-culture assays were divided into two experimental sets, including bacterial growth controls and Tetrahymena co-culture assays, resulting in a total of eight samples: WT, Δadc, WT + ARG, and Δadc + ARG for each set. For the co-culture assays, 100 μL of bacterial suspension was mixed with 100 μL of Tetrahymena culture, whereas for the growth control assays, 100 μL of bacterial suspension was mixed with 100 μL of LB medium. All mixtures were transferred into 96-well plates. For arginine-treated groups, L-arginine was added during the mixing process to a final concentration of 500 μM, followed by incubation at 30 °C for 5 h. followed by incubation at 30 °C for 5 h. Bacterial growth in the mixed suspension was monitored hourly by measuring the absorbance at 600 nm. Control groups contained bacterial strains mixed with an equal volume of SPP medium instead of Tetrahymena, and sterile SPP medium was used as the blank.
2.7. RNAseq and qRT-PCR Analysis
Wild-type (WT) and ∆adc mutant strains were cultured in LB medium at 30 °C overnight and then diluted 1:100 into fresh LB for 12 more hours. Six samples from each strain were collected by centrifugation, and total RNA was extracted using the RNAprep pure cell/bacteria kit (TianGen, Beijing, China). For RNA sequencing, six RNA samples from each strain were mixed proportionally to their molar mass and assessed with an Agilent 2100 bioanalyzer. Libraries were constructed with the NEBNext^®^ Ultra™ II Directional RNA Library Prep Kit (NEB, Ipswich, MA, USA) for paired-end 150 bp reads and sequenced on the Illumina NovaSeq 6000 platform by Novogene biotech (Beijing, China). Reads were aligned to the V. anguillarum 775 reference genome using Bowtie 2.3.4.3, and gene expression levels were quantified as RPKM. Differential expression analysis was performed with the edgeR package (3.24.3), setting a corrected p-value of 0.05 and a log2(Fold change) of 1 as thresholds for significance. DEGs related to key KEGG pathways were validated by qRT-PCR. Primers are shown in Table 3. Reverse transcription was done using a cDNA reverse transcription kit (Yeasen, Shanghai, China), and qRT-PCR was performed with an initial denaturation at 95 °C for 30 s, followed by 40 cycles of 95 °C for 10 s and 60 °C for 30 s. The relative expression of each gene in the mutant was quantified relative to WT strain after normalization to the 16S rRNA gene using the 2^−∆∆Ct^ method. The experiment was conducted in triplicate.
2.8. Statistical Analysis
All statistical analyses were performed using GraphPad Prism version 9 (GraphPad Software, San Diego, CA, USA). Differences among groups were evaluated by one-way or two-way analysis of variance (ANOVA), followed by Duncan’s multiple range post hoc test when appropriate. All experiments were conducted with at least three independent biological replicates, and data are presented as the mean ± standard deviation (SD). Statistical significance was defined as p < 0.05, with significance levels indicated as * p < 0.05, ** p < 0.01, *** p < 0.001, different uppercase letters indicate significant differences at p < 0.01.
3. Results
3.1. Effects of L-arginine and Its Metabolites on Hemolytic Activity of Vibrio Anguillarum
To investigate the regulatory effects of L-arginine (ARG) and its metabolites, namely agmatine (AGM), putrescine (PUT), and spermine (SPE), on the hemolytic activity of the WT strain, we treated the strain with these substances at concentrations ranging from 0.625 to 5 × 10^4^ μM and determined the corresponding hemolysis rates. L-arginine (ARG) showed a stable hemolysis rate ranging from 39% to 40% at 0.625–5 μM, then increased to 46.4% (50 μM) and 45.4% (500 μM) at medium concentrations, and decreased to 32.5% and 21.2% at 5000 and 5 × 10^4^ μM, respectively (Figure 1A). Agmatine (AGM) displayed the most prominent promoting effect: its hemolysis rate remained at ~39–40% at 0.625–2.5 μM, peaked at 52.7% at 50 μM (the highest among all tested substances), and then dropped sharply to 19.2% and 6.1% at 5000 and 5 × 10^4^ μM, respectively. For putrescine (PUT), the hemolysis rate was sustained at ~39% at 0.625–2.5 μM, reached a peak of ~44.6% at 50 μM, and was significantly inhibited at concentrations ≥5000 μM. Similarly, spermine (SPE) exhibited a hemolysis rate ranging from 39% to 41% at 0.625–5 μM, peaked at 48.3% at 50 μM, and decreased to 16.6% and 15.1% at 5000 and 5 × 10^4^ μM, respectively.
In conclusion, L-arginine and its metabolites (AGM, PUT, SPE) all regulate the hemolytic activity of the WT strain in a concentration-dependent biphasic manner, with AGM showing the most significant promoting effect at 50 μM. These results elucidate the distinct regulatory roles of L-arginine and its metabolites in bacterial hemolytic activity, providing a clear basis for further mechanistic explorations.
3.2. Construction of Mutant Strains and Mechanistic Elucidation of Hemolytic Ability
To dissect the mechanistic basis by which L-arginine and its metabolites modulate hemolysis in Vibrio anguillarum, we interrogated the metabolic network of L-arginine and its derivatives in this pathogen. The arginine metabolic pathway is evolutionarily conserved, and in V. anguillarum, arginine is channeled into two distinct metabolic branches: one catalyzed by arginine decarboxylase (adc, encoded by adc), directing arginine towards agmatine biosynthesis; the other initiated by arginine N-succinyltransferase (AstA, encoded by astA), with the resultant N_2_-succinyl-L-arginine further metabolized by succinylglutamic semialdehyde dehydrogenase (AstD, encoded by astD) to yield N(2)-succinyl-glutamate, a precursor for glutamate biosynthesis (Figure 2A). We generated a panel of isogenic mutants in V. anguillarum via homologous recombination: adc deletion mutant (∆adc), astA deletion mutant (∆astA), astD deletion mutant (∆astD), astA/astD double deletion mutant (∆astA∆astD), and deletion mutant (encodes argininosuccinate lyase, a rate-limiting enzyme in de novo arginine biosynthesis). To quantify hemolytic potential, we assessed hemolysis zone diameters on Columbia blood agar plates. Strikingly, only ∆adc exhibited a dramatic reduction in hemolytic activity, displaying near-abolished hemolysis. In contrast, ∆astA, ∆astD, and ∆astA∆astD showed hemolysis zones indistinguishable from the wild type (WT), indicating that disruption of the agmatine biosynthesis branch, but not the succinylative branch or de novo arginine biosynthesis, abrogates hemolysis (Figure 2B; p < 0.001, ns = not significant).
To corroborate these findings, we performed a time-course liquid hemolysis assay. Consistent with plate-based assays, ∆adc failed to induce significant hemolysis within the first 2 h of incubation with sheep erythrocytes, whereas WT achieved nearly complete erythrocyte lysis. Notably, ∆adc exhibited delayed hemolysis at later time points, with a hemolysis rate of ~36% at 3 h, whereas WT maintained a steadily increasing hemolysis rate, reaching near 100% at 3 h (Figure 2C,D).
To delineate the metabolite(s) mediating hemolysis in the adc pathway, we supplemented ∆adc cultures with graded concentrations of L-arginine (L-arg), agmatine (AGM), putrescine (PUT), and spermine (SPE), and measured hemolysis after 2 h. L-arg supplementation exerted a biphasic effect: low concentrations (≤5 μM) had no discernible impact, a moderate increase was observed at 50 μM, and higher concentrations (≥5 × 10^3^ μM) significantly inhibited hemolysis. In contrast, AGM, PUT, and SPE dose-dependently restored hemolytic activity in ∆adc. AGM at 0.625 μM elicited a 6% increase in hemolysis relative to the control without supplementation, and maximal recovery (51.8%, a 13% increase vs. control) was observed at 50 μM. Similarly, PUT and SPE showed concentration-dependent restoration, with peak hemolysis at 1 μM (51.6%) and 10 μM (48.5%), respectively (Figure 2E–H). These data collectively demonstrate that the agmatine biosynthesis branch, and its downstream polyamines, are pivotal for V. anguillarum hemolysis.
3.3. Imapct of the adc Mutant on V. anguillarum
Based on the above results, the deletion of adc critically impairs the hemolytic virulence of V. anguillarum. To explore the genome-wide regulatory effect of adc deficiency, we performed transcriptomic sequencing. A total of 704 significantly differentially expressed genes (DEGs) were identified between the ∆adc mutant and WT strains, including 308 up-regulated and 396 down-regulated genes (Figure 3A). KEGG pathway enrichment analysis of the down-regulated DEGs revealed the enrichment of multiple pathways associated with bacterial virulence and metabolism (Figure 3B). The most significantly enriched pathways included flagellar assembly; bacterial secretion system; biofilm formation—Vibrio cholerae; and glycolysis/gluconeogenesis, alongside basic metabolic pathways such as carbon metabolism and ribosome. These results indicate that adc deletion not only affects hemolysis-related virulence pathways but also broadly regulates bacterial secretion systems, motility, and energy metabolism. We further identified 21 DEGs closely associated with hemolysin synthesis and secretion, of which 2 were significantly up-regulated and 14 were significantly down-regulated. Hierarchical clustering heatmap analysis (Figure 3C) of these hemolysis and secretion system-related genes revealed that key hemolytic genes (e.g., VAA_RS06290(vah3), empA, rtxC) and Type VI secretion system (T6SS) genes (e.g., tssH, tssM, tssJ) were significantly down-regulated in the ∆adc mutant, with only a few genes showing up-regulation. This suggests that adc modulates hemolytic activity by regulating the expression of these virulence-associated genes. To confirm the transcriptomic findings, we performed qRT-PCR on a subset of key genes involved in hemolysis and the bacterial secretion system (Figure 3D). The results demonstrated that the expression of vah3 (VAA_RS06290), VAA_RS05900, empA, rtxC, and other genes was significantly down-regulated in the ∆adc mutant, which was highly consistent with the transcriptomic data. This validation confirms that adc deletion negatively regulates the expression of these virulence-related genes.
3.4. Virulence Assessment of the ∆adc Mutant Using a Tetrahymena Co-Culture Model
To further evaluate the virulence of the ∆adc mutant, a series of co-culture assays were performed with the ciliated protozoan Tetrahymena, a widely used model for investigating bacterial anti-phagocytic capacity and cytotoxicity. In the bacteria-free control group, Tetrahymena cells maintained normal cellular morphology and vigorous proliferative activity. Following co-incubation with the wild-type (WT) V. anguillarum strain, Tetrahymena exhibited severe morphological abnormalities, including marked cellular shrinkage and deformation, accompanied by significant growth suppression. In contrast, co-culture with the ∆adc mutant resulted in markedly alleviated cytotoxic effects: Tetrahymena displayed fewer aberrant cells and higher viable cell counts, indicative of attenuated virulence (Figure 4A). Bacterial growth dynamics during co-culture were monitored by measuring the optical density at 600 nm (OD_600_). After 6 h of co-incubation, the OD_600_ value of the ∆adc mutant was significantly lower than that of the WT strain (Figure 4B,C), demonstrating that the ∆adc mutant was phagocytosed at a substantially higher rate and exhibited impaired resistance to predation by Tetrahymena.
To further assess cytotoxicity, bacterial cell lysates of the WT and ∆adc strains were used to challenge Tetrahymena, and cell viability was quantified via the Cell Counting Kit-8 (CCK-8) assay. Tetrahymena cells treated with ∆adc lysates showed significantly higher viability than those exposed to WT lysates (Figure 4D), confirming that deletion of adc also reduced the cytotoxic potential of V. anguillarum. Collectively, these results demonstrate that loss of adc significantly compromises both the anti-phagocytic ability and cytotoxicity of V. anguillarum, leading to an obvious reduction in bacterial virulence.
3.5. Biofilm and Antimicrobial Susceptibility Defects in the ∆adc Mutant
The ∆adc mutant exhibited a profound reduction in biofilm formation, with OD595 values decreasing from 1.4 ± 0.2 in the WT to 0.5 ± 0.1 in the mutant. Crystal violet staining confirmed impaired biofilm development (Figure 5A). To assess antimicrobial susceptibility, minimal inhibitory concentrations (MICs) of 10 clinically relevant antibiotics were determined, with results presented as log_2_ fold changes (∆adc MIC/WT MIC) (Figure 5B). The ∆adc mutant exhibited a 2-fold reduction in MICs for β-lactam antibiotics (cefotaxime, ampicillin), quinolones (nalidixic acid), macrolides (clarithromycin), and aminoglycosides (streptomycin, tobramycin), indicating increased susceptibility to these classes of antibiotics. In contrast, MICs for tetracycline, and chloramphenicol remained unchanged between the ∆adc mutant and WT strains (log_2_ fold change = 0). The heatmap in Figure 5B visually represents these differences, with red-to-white gradients indicating decreasing MIC ratios (∆adc/WT).
4. Discussion
This study investigated the regulatory function of arginine in Vibrio anguillarum by constructing a series of arginine metabolism-related deletion mutants (∆adc, ∆astA, ∆astD, ∆astAastD) and performing metabolite supplementation, transcriptomic sequencing and multiple phenotypic assays. Our results demonstrated that adc mediates the agmatine synthesis branch of L-arginine metabolism, and deletion of adc significantly abolished the hemolytic activity of V. anguillarum, downregulated the transcription of multiple virulence-related genes, reduced bacterial anti-phagocytic capacity and cytotoxicity, impaired biofilm formation, and increased susceptibility to multiple classes of antibiotics.
L-arginine (L-arg) exhibits remarkable metabolic and regulatory diversity across diverse organisms, being ubiquitous in all domains of life and playing a pivotal role in host–microbe interactions [8,9]. As such, L-arg metabolism is centrally important for numerous biological processes, as well as the crosstalk between mammals, microbes, and plants [8,22,23]. Beyond serving as a precursor for polyamine biosynthesis and a building block for protein synthesis, L-arg is critical for microbial growth, differentiation, and energy metabolism [8,22]. Consequently, the availability and metabolism of L-arg in microbes are tightly regulated by multiple anabolic and catabolic enzymatic pathways. Previous studies have demonstrated that exogenous L-arg supplementation induces virulence gene expression in enterohemorrhagic Escherichia coli (EHEC) in vitro, while arginine transport-deficient mutants (∆artP) show reduced virulence gene expression [6]. In 2023, Björn Nüse et al. further highlighted the enzymatic pathways of L-arg metabolism in microbes and mammalian cells, as well as their significant roles in immune function, luminal metabolism, colonization resistance, and the pathogenic mechanisms of intestinal microbes [7]. However, limited studies have addressed the effects of L-arg and its metabolism on bacterial virulence in aquatic pathogenic vibrios, particularly Vibrio anguillarum-a major pathogen threatening both marine and freshwater cultured animals. In this study, we are the first to report that L-arginine and its downstream metabolites (agmatine, putrescine, spermine) regulate the hemolytic activity of wild-type V. anguillarum in a concentration-dependent biphasic manner. Notably, although high concentrations of L-arg inhibit bacterial hemolysis and significantly reduce virulence, the natural concentration of L-arg in aquatic environments is generally very low. Thus, the inhibitory effect of high concentrations is physiologically irrelevant under natural conditions, and L-arg typically exerts a stimulatory effect at environmentally relevant low concentrations. Among the tested metabolites, agmatine exhibited the most potent stimulatory effect on hemolysis, further emphasizing its key role in this regulatory axis.
From a translational perspective, our findings highlight the need to validate arginine-based interventions in fish. Dietary arginine can benefit host growth/immune function and may exert prebiotic-like effects on gut microbiota [17,18,19]; however, our in vitro data indicate that low micromolar arginine and polyamines can enhance V. anguillarum hemolysis. Therefore, controlled fish challenge experiments and, ultimately, pond/net-pen field trials are required to determine the net outcome of arginine supplementation under realistic farming conditions (e.g., temperature/salinity, stocking density, background microbiota). Such studies should quantify survival, bacterial loads, and host immune/barrier responses, while monitoring gut microbiome and metabolite changes and testing dose/timing regimens to minimize any unintended promotion of pathogen virulence [24].
While a limited number of studies have reported that L-arg modulates bacterial virulence, the underlying mechanisms by which L-arg promotes virulence and regulates hemolysin secretion remain unclear in V. anguillarum. In this pathogen, L-arg metabolism primarily proceeds through two distinct pathways: one catalyzed by arginine decarboxylase (encoded by adc) that directs L-arg toward agmatine biosynthesis, and the other mediated by arginine N-succinyltransferase (encoded by astA) and succinylglutamate desuccinylase (encoded by astD) that metabolizes L-arg to L-glutamate. To dissect the mechanism by which L-arg influences virulence, we constructed four isogenic mutants targeting key genes in these pathways (∆adc, ∆astA, ∆astD, ∆astA∆astD). Phenotypic analysis revealed that only the ∆adc strain exhibited near-complete loss of hemolytic activity, whereas ∆astA, ∆astD, and ∆astA∆astD showed hemolytic phenotypes indistinguishable from the wild-type strain. Moreover, exogenous supplementation with agmatine, putrescine, or spermine dose-dependently restored the hemolytic activity of ∆adc, confirming that these polyamines are the critical effectors of the adc-mediated pathway.
Previous studies have confirmed that polyamines act as important signaling molecules regulating the virulence of pathogenic Vibrio species and other pathogens [12,25,26,27]. Our results further clarify that the adc-dominated agmatine biosynthesis branch—rather than the succinylative catabolic branch or de novo arginine synthesis pathway—serves as the key metabolic node for L-arginine-mediated regulation of V. anguillarum hemolysis. This finding supplements the regulatory mechanism of arginine metabolism on core virulence phenotypes in aquatic vibrios. The adc gene and its mediated arginine decarboxylation pathway have been widely studied in Gram-negative bacteria, primarily focusing on bacterial acid stress tolerance and polyamine synthesis (e.g., Escherichia coli, Vibrio cholerae, Pseudomonas aeruginosa) [1,2,3]. However, the specific role of adc in regulating the virulence and physiological phenotypes of V. anguillarum—a dominant aquatic pathogen with significant impacts on aquaculture—has not been systematically elucidated prior to this study. Our findings thus fill this knowledge gap, highlighting the species-specific regulatory function of adc in linking arginine metabolism to virulence in V. anguillarum.
Transcriptomic analysis showed that 704 differentially expressed genes were identified between the ∆adc mutant and the wild-type strain, and the down-regulated genes were significantly enriched in flagellar assembly, bacterial secretion system, biofilm formation and glycolysis/gluconeogenesis pathways. Key hemolysin genes such as vah3, empA, rtxC and Type VI secretion system (T6SS) genes including tssH, tssM, tssJ were significantly down-regulated, and the qRT-PCR results were highly consistent with the transcriptomic data. It has been documented that the virulence of pathogenic Vibrio is a complex trait regulated by multiple gene networks, and hemolysin, secretion system and flagellar system are usually co-regulated to jointly mediate bacterial adhesion, invasion and cytotoxicity [3,28]. The widespread down-regulation of virulence and metabolic pathways in the ∆adc mutant indicates that adc is not only a structural gene involved in arginine metabolism, but also a global regulatory factor affecting the physiological and pathogenic processes of V. anguillarum.
In addition, the ∆adc mutant exhibited reduced biofilm formation and lower MICs to several antibiotic classes. Importantly, these MIC shifts in an isogenic, laboratory-constructed mutant should not be interpreted as antimicrobial resistance (AMR) in the epidemiological sense; AMR is driven by genetic change and selection under antimicrobial exposure and is accelerated by misuse and overuse of antibiotics [29,30]. Instead, our data most likely reflect altered intrinsic susceptibility/tolerance linked to metabolic homeostasis, envelope permeability, and/or biofilm-associated protection [31,32]. Biofilms can promote environmental persistence and antimicrobial tolerance by restricting penetration, altering metabolic states, and enabling stress responses [31]. Intracellular amino acid metabolism and polyamines are also known to regulate biofilm maturation in Gram-negative bacteria [33,34]. Combined with transcriptomic data, the impaired biofilm formation in ∆adc may stem from metabolic imbalance induced by blocked arginine decarboxylation, though the specific polyamine-mediated mechanism requires further validation. Overall, the increased antibiotic susceptibility of ∆adc suggests that adc-dependent polyamine metabolism contributes to baseline stress tolerance and cell envelope homeostasis rather than acquired AMR.
In the Tetrahymena co-culture model, the ∆adc mutant displayed significantly reduced growth and diminished cytotoxicity of its cell lysates toward Tetrahymena, directly demonstrating that deletion of adc attenuates the anti-phagocytic ability and cytotoxicity of V. anguillarum—two key virulence traits underlying bacterial pathogenicity. As a ubiquitous predator in aquatic ecosystems, Tetrahymena exerts strong selective pressure on the virulence evolution of aquatic pathogenic bacteria, and bacterial strains with compromised anti-phagocytic capacity typically exhibit attenuated pathogenic potential in hosts [21,35]. Our results confirm that adc is essential for mediating V. anguillarum’s environmental adaptability and pathogenicity by regulating anti-phagocytic capacity. Notably, the attenuated virulence of ∆adc is not limited to anti-phagocytosis and cytotoxicity: the mutant also shows drastically reduced hemolytic activity, impaired biofilm formation, and increased susceptibility to multiple classes of antibiotics. Collectively, these multi-faceted virulence defects align with the notion that gene deletion strains with disrupted key metabolic pathways and attenuated virulence are promising candidates for attenuated live vaccines [15,20]. Thus, the ∆adc strain exhibits comprehensive virulence attenuation mediated by a core metabolic pathway, offering a novel and effective strategy for the control of vibriosis in aquaculture.
5. Conclusions
In conclusion, this study systematically clarified the pivotal regulatory role of arginine in V. anguillarum using a panel of arginine metabolism-related deletion mutants, with a focus on the adc deletion mutant (∆adc). Results showed that adc deletion abolished hemolytic activity, impaired anti-phagocytic ability and biofilm formation and increased susceptibility to multiple antibiotics, while exogenous agmatine, putrescine or spermine restored hemolysis. These findings align with studies in other Gram-negative bacteria, highlighting adc’s conserved role in linking arginine metabolism to bacterial virulence. Notably, adc deletion attenuated V. anguillarum pathogenicity by disrupting agmatine biosynthesis, suggesting ∆adc as a potential candidate for attenuated vaccines against vibriosis. However, the study was limited by the lack of long-term in vivo fish infection models and unclear downstream molecular targets of adc. Future studies should address these limitations to evaluate the long-term protective efficacy of ∆adc strains and clarify the detailed regulatory mechanisms of adc, facilitating the development of effective strategies to control V. anguillarum infections.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Urku C. Secer F.S. Onalan S. Akayli T. Investigation of vibriosis caused by Vibrio anguillarum in rainbow trout (Oncorhynchus mykiss)Cell. Mol. Biol. Noisy Grand 202470323810.14715/cmb/2024.70.8.439262265 · doi ↗ · pubmed ↗
- 2Pazos F. Santos Y. Magarinos B. Bandin I. Nunez S. Toranzo A.E. Phenotypic Characteristics and Virulence of Vibrio anguillarum-Related Organisms Appl. Environ. Microbiol.1993592969297610.1128/aem.59.9.2969-2976.199316349042 PMC 182394 · doi ↗ · pubmed ↗
- 3Frans I. Michiels C.W. Bossier P. Willems K.A. Lievens B. Rediers H. Vibrio anguillarum as a fish pathogen: Virulence factors, diagnosis and prevention J. Fish Dis.20113464366110.1111/j.1365-2761.2011.01279.x 21838709 · doi ↗ · pubmed ↗
- 4Xie H. Huang Y. Wang S. Che J. Luo T. Li L. Bao B. Deletion of spe A and aro C genes impacts the pathogenicity of Vibrio anguillarum in spotted sea bass Microb. Pathog.202418910659710.1016/j.micpath.2024.10659738395316 · doi ↗ · pubmed ↗
- 5Li Z. Sun Y. Tan R. Gao Y. Identification, characterization and complete genome analysis of a Vibrio anguillarum isolated from Sebastes schlegelii Microb. Pathog.202419010661110.1016/j.micpath.2024.10661138467165 · doi ↗ · pubmed ↗
- 6Irshath A.A. Rajan A.P. Vimal S. Prabhakaran V.-S. Ganesan R. Bacterial Pathogenesis in Various Fish Diseases: Recent Advances and Specific Challenges in Vaccine Development Vaccines 20231147010.3390/vaccines 1102047036851346 PMC 9968037 · doi ↗ · pubmed ↗
- 7Menezes-Garcia Z. Kumar A. Zhu W. Winter S.E. Sperandio V. l-Arginine sensing regulates virulence gene expression and disease progression in enteric pathogens Proc. Natl. Acad. Sci. USA 2020117123871239310.1073/pnas.191968311732409599 PMC 7275701 · doi ↗ · pubmed ↗
- 8Nuse B. Holland T. Rauh M. Gerlach R.G. Mattner J. L-arginine metabolism as pivotal interface of mutual host-microbe interactions in the gut Gut Microbes 202315222296110.1080/19490976.2023.222296137358082 PMC 10294761 · doi ↗ · pubmed ↗