PurK, N5-Carboxyaminoimidazole Ribonucleotide Synthetase, an Exocrine Protein Induced by Potato Plants, Influences the Virulence Through Motility Modulation in Pectobacterium brasiliense NJAU180
Lingyan Xia, Yuanxu Zhuo, Nanqiao Lin, Na Yu, Shu Che, Chunting Wang, Liping Yang, Baishi Hu, Yanli Tian, Jiaqin Fan

TL;DR
A protein called PurK, produced by a potato pathogen, helps the bacteria infect plants by boosting movement and suppressing plant defenses.
Contribution
PurK is identified as a novel extracellular protein in Pectobacterium brasiliense that modulates virulence through motility and immune suppression, independent of its role in purine biosynthesis.
Findings
PurK is a plant-induced extracellular protein in Pectobacterium brasiliense that significantly reduces virulence when deleted.
PurK promotes bacterial motility by modulating flagellar gene transcription and suppresses plant callose deposition.
PurK triggers hypersensitive responses and upregulates PTI marker genes in host plants, but its effects are independent of purine biosynthesis.
Abstract
Bacterial pathogens secrete effector proteins that suppress plant immune responses and facilitate infection. This study focuses on Pectobacterium brasiliense NJAU180, a bacterial pathogen causing severe blackleg disease in potato plants in Inner Mongolia, China. Using exoproteomic analysis, plant-induced extracellular proteins were identified by comparing culture supernatants from P. brasiliense NJAU180 grown in minimal medium (MM) alone and in the presence of aseptically grown potato plantlets at an early growth stage (OD600 ≈ 0.5). The results reveal PurK as a novel plant-induced extracellular protein, and deletion of purK markedly reduces virulence. PurK, N5-carboxyaminoimidazole ribonucleotide synthetase, is a key enzyme in de novo purine biosynthesis. Its impact on virulence is distinct from the conventional production of plant cell wall–degrading enzymes: PurK promotes motility by…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
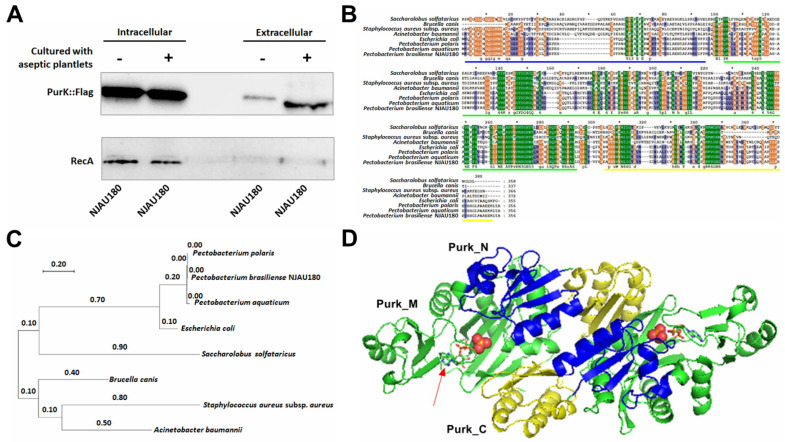 Figure 1
Figure 1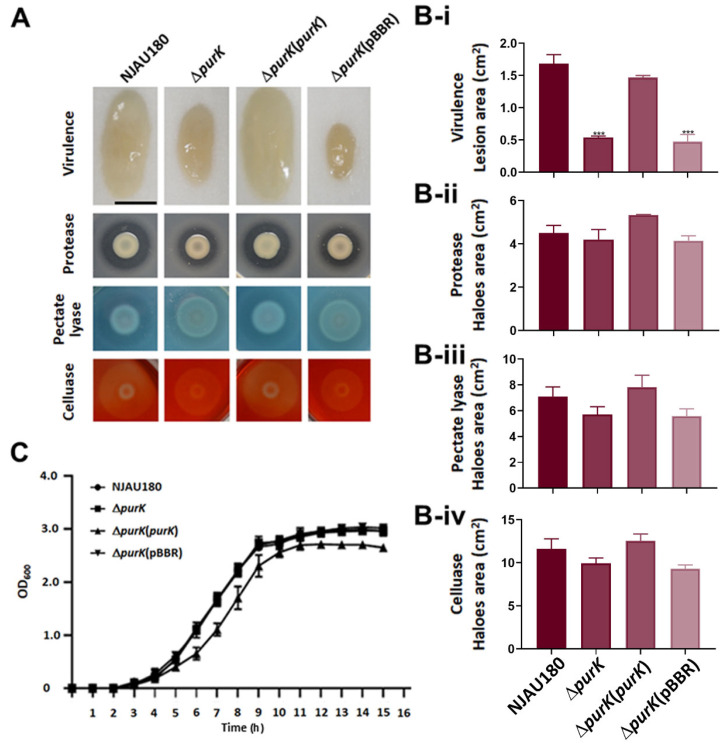 Figure 2
Figure 2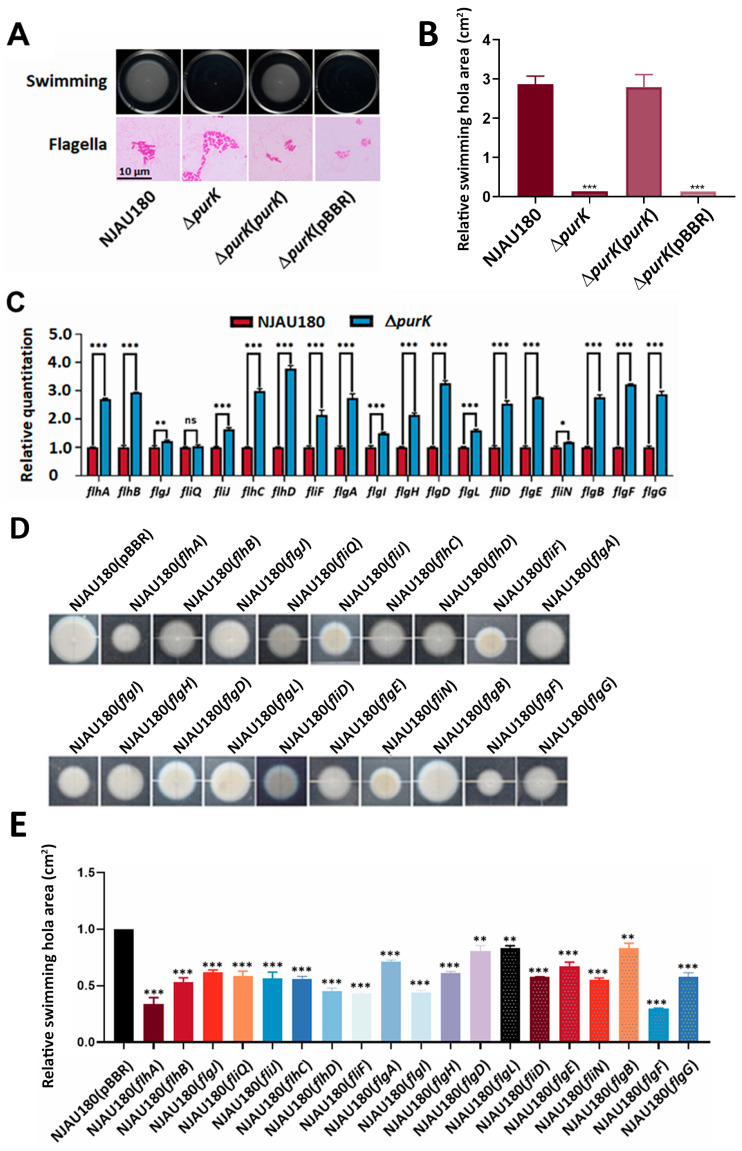 Figure 3
Figure 3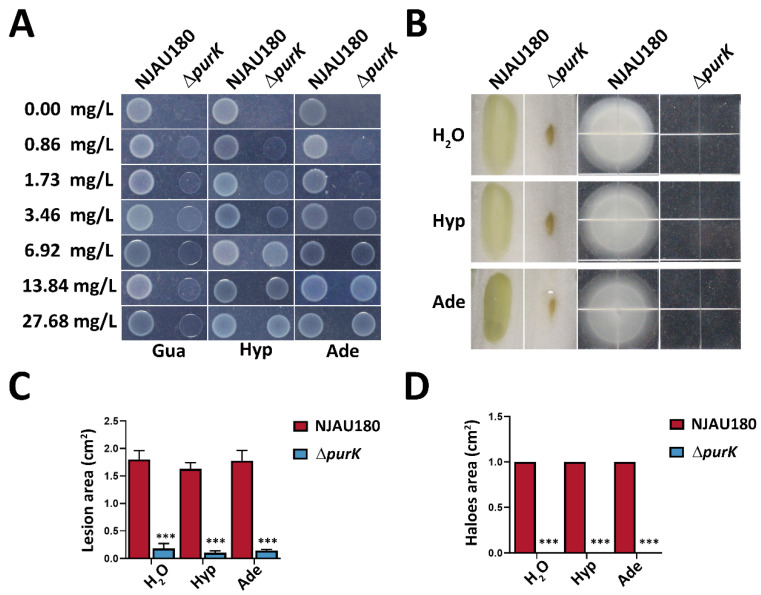 Figure 4
Figure 4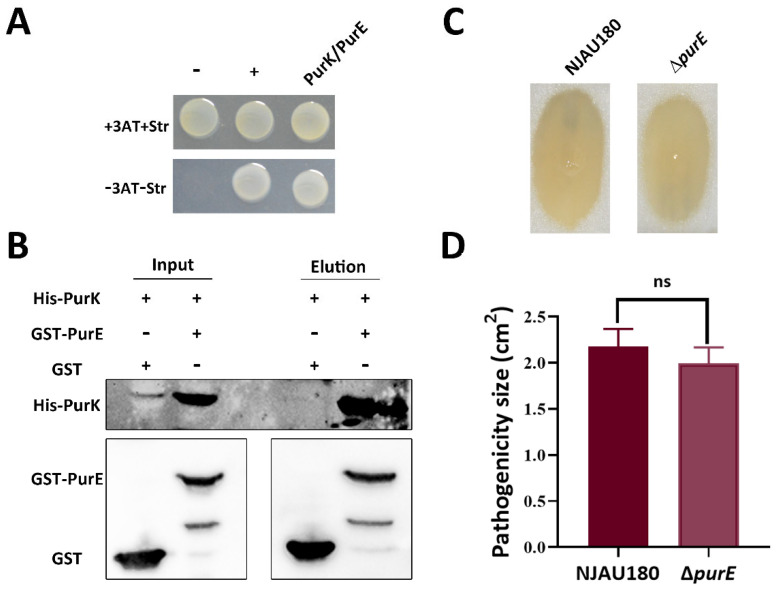 Figure 5
Figure 5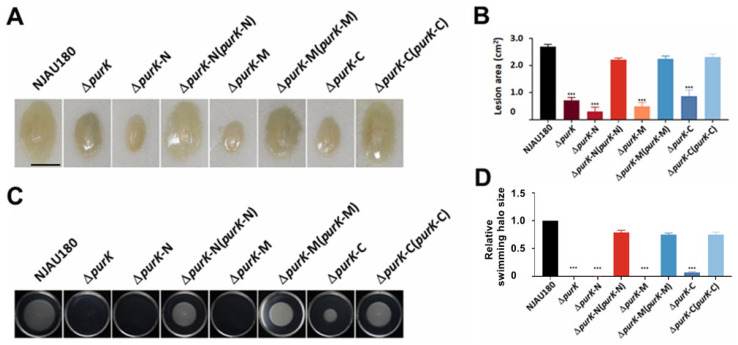 Figure 6
Figure 6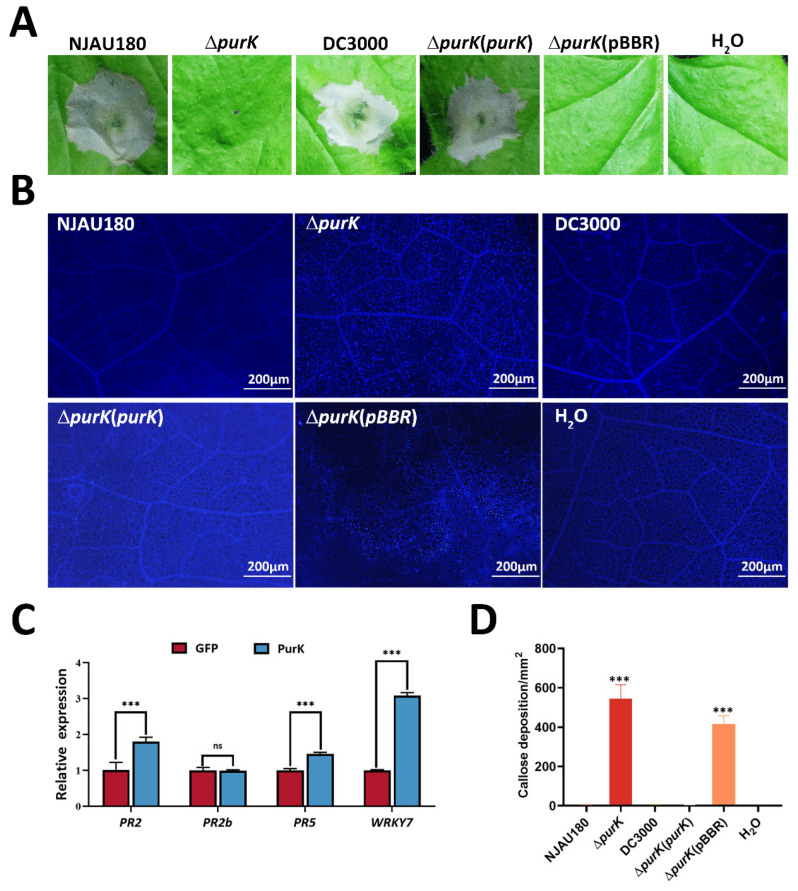 Figure 7
Figure 7- —the National Key Research and Development Program of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogenic Bacteria Studies · Plant-Microbe Interactions and Immunity · Bacterial biofilms and quorum sensing
1. Introduction
The strains from the genus of Pectobacterium are important plant pathogenic bacteria that infect various crops and cause severe soft rot disease. These bacteria produce a variety of enzymes that degrade cell walls, including pectinases, cellulases, and proteases. These enzymes disrupt the structure of the plant cell wall, particularly the middle lamella, resulting in tissue maceration and softening, and ultimately develop a viscous and unpleasantly odorous rot [1,2]. They have a broad host range, including many economically important crops from the Brassicaceae, Solanaceae, and Cucurbitaceae families [3]. These strains occasionally exist in a latent state within seemingly healthy plant organs, including potato tubers, taro corms, and the fruits of tomatoes and peppers [3,4]. Under conditions of high temperature and humidity during harvest, transportation, or storage, they proliferate quickly and spread widely, leading to extensive decay and substantial economic losses postharvest [5]. Bacteria–plant interaction is a dynamic and complex process of attack and defense. During infection, pathogens have evolved various virulence strategies to enhance their ability to colonize and infect host tissues. For instance, by forming biofilms either on the surface or within plant tissues, pathogens improve their adhesion, resist environmental stresses, and achieve persistent survival [6]. Some pathogens can produce phytotoxins that interfere with the host’s physiological metabolism and disrupt normal cellular functions [7].
In response to pathogen attacks, host plants have evolved a highly sophisticated innate immune system. The first line of defense is known as pathogen-associated molecular patterns (PAMP) triggered immunity (PTI). This system is activated when pattern recognition receptors located on the plant cell surface detect conserved molecular patterns associated with pathogens [8]. Early events in this immune response include a burst of reactive oxygen species and oscillations in cytosolic calcium concentrations [9], followed by the activation of mitogen-activated protein kinase (MAPK) signaling cascades. These pathways regulate the expression of defense-related genes and promote the synthesis of phytoalexins and pathogenesis-related proteins [10,11]. Simultaneously, plants reinforce physical barriers by depositing callose at the cell wall, effectively impeding further pathogen invasion [12].
Bacterial motility is one of the key virulence factors enabling the pathogens such as Pectobacterium to achieve successful infection [1]. This motility primarily relies on the flagellum—a highly conserved molecular machine that not only provides propulsive force but also participates in sensing environmental chemical signals, thereby regulating chemotactic behavior and guiding the bacterium toward specific host sites for targeted migration [13]. Structurally, the flagellum can be divided into three main parts: the basal body, the hook, and the filament. The basal body is embedded in the cell membrane and functions as a rotary motor driven by proton or sodium ion gradients. The filament, formed by the polymerization of flagellin, acts as a propeller, propelling the bacterium through liquid or semi-solid media [14,15,16]. The assembly of this sophisticated structure is under strict genetic regulation, involving multiple hierarchically expressed operons, with master regulators such as flhDC and fliA playing central roles in this hierarchical network [17]. Through flagellum-mediated motility, P. carotovorum can effectively traverse physical barriers on the plant surface, locate infection sites, and enhance its ability to migrate, colonize, and spread within host tissues, thereby increasing its pathogenic efficiency.
Purines are essential nitrogenous bases that constitute fundamental biomolecules such as DNA, RNA, and ATP [18]. In living organisms, the purine ring structure is primarily assembled stepwise via the de novo biosynthesis pathway [19]. A key enzyme in this pathway, PurK, catalyzes the fifth-step carboxylation reaction, converting the substrate CAIR (5-aminoimidazole-4-carboxamide ribonucleotide) into N^5^-CAIR. This step not only serves as a critical energy checkpoint in purine biosynthesis but also plays an important regulatory role in maintaining metabolic flux [20,21]. Researches have shown that mutations in the pur operon encoding PurK significantly attenuate bacterial virulence. For example, in Brucella species, the integrity of the purine biosynthesis pathway has been demonstrated to be essential for persistent infection in vivo, marking the first direct evidence linking purine synthesis and pathogenicity [22]. However, how PurK influences virulence remains unknown.
In this study, a secreted protein, PurK, was identified by comparing the exoproteome in the supernatant of pathogenic strain P. brasiliense NJAU180, which was grown in the presence and absence of aseptically grown potato plantlets, and a purK deletion mutant was created to systematically evaluate the impact of this gene on the growth and pathogenicity of NJAU180. The results demonstrate that PurK primarily influences virulence by modulating motility in NJAU180, unlike other strains of Pectobacterium, which rely on potent cell wall-degrading enzymes. It elucidates new functional roles of PurK in the pathogenesis of NJAU180 focusing on its secreted proteins.
2. Materials and Methods
2.1. Bacterial Strains and Plasmid Construction
The purK gene deletion mutant (ΔpurK) and the domain-specific deletion mutants (ΔpurK-N, ΔpurK-M, and ΔpurK-C) from the wild-type P. brasiliense strain NJAU180 were constructed using homologous recombination-mediated knockout technology. For functional complementation assays, the pBBR plasmid harboring the fragment of purK gene was introduced into ΔpurK to generate the complemented strain ΔpurK (purK). Similarly, pBBR plasmids carrying the respective domain fragments (purK-N, purK-M, or purK-C) were transformed respectively into ΔpurK-N, ΔpurK-M, and ΔpurK-C to generate the corresponding complemented strains ΔpurK-N (purK-N), ΔpurK-M (purK-M), and ΔpurK-C (purK-C). Meanwhile, the empty pBBR vector was transformed into ΔpurK to obtain the control strain ΔpurK (pBBR), which was used to assess the potential influence of the vector alone on subsequent phenotypic assays. All constructs were verified by PCR amplification and DNA sequencing.
2.2. Bacterial Strains, Plant Materials and Growth Conditions
The bacterial strains and plasmids used in this study are summarized in Supplementary Table S1. The wild-type strain P. brasiliense NJAU180 and its derivate strains were cultured in Luria-Bertani (LB) [23] or minimal medium (MM) [24] at 28 °C, and the derivate strains from E. coli were cultured in LB at 37 °C. The final antibiotic concentrations as shown below: kanamycin (Km) at 50 μg·mL^−1^, rifampin (Rif) at 100 μg·mL^−1^ and gentamicin (Gm) at 50 μg·mL^−1^. The optical density (OD) of the culture was measured by a BioPhotometer (Eppendorf, Hamburg, Germany) at 600 nm. Potato (Solanum tuberosum DM1-3) aseptic cultured seedlings were grown in 1/2 MS medium in an illumination incubator at 26 °C under a photoperiod at 16 h light/8 h dark. The plants of Nicotiana benthamiana were cultured in a greenhouse (temperature varying from 22 to 28, 16 h day length and an approximately 80% relative humidity).
2.3. RNA Isolation, cDNA Synthesis, qRT-PCR
Total RNA was extracted from P. brasiliense NJAU180 and its derived strains using an RNA extraction kit (Cat. No. AF505B; Proteinssci, Shanghai, China). To eliminate potential genomic DNA contamination, the extracted RNA was treated with DNase I. First-strand cDNA was then synthesized using Prime Script Reverse Transcriptase (Vazyme, Nanjing, China). Quantitative real-time PCR (qRT-PCR) was performed using the SYBR Prime-Script RT-PCR Kit (Cat. No. R323-01; Vazyme, Nanjing, China) on an ABI Prism 7500 Fast Real-Time PCR System (Applied Biosystems Inc., Foster City, CA, USA). The relative expression levels were calculated using the 2^−ΔΔCT^ method. All primers used in this study are listed in Supplemental Table S2, with the recA gene serving as an internal control as previously described [24].
2.4. Protein Extraction and Western Blotting
The wild-type strain P. brasiliense NJAU180 and its derived strains were complemented with a plasmid carrying the target gene fused with an epitope tagging fragment (DYKDDDDK) at the C-terminus as previously described [25]. Intracellular proteins were extracted using SDS lysis buffer (20 mM Tris-HCl, pH 7.0, 4% SDS, 20% glycerol), and extracellular proteins were collected as described previously [25,26]. The protein samples were separated by SDS-PAGE and transferred to a PVDF membrane for immunoblotting analysis. For detection of the FLAG-tagged target protein, the membrane was first incubated with a mouse anti-FLAG monoclonal primary antibody (1:3000 dilution; Cat. No. F1804, Merck KGaA, Darmstadt, Germany) at 4 °C overnight. After thorough washing, the membrane was then incubated with a horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG secondary antibody (1:5000 dilution; Cat. No. 31430, Thermo Fisher Scientific Inc., Waltham, MA, USA) at room temperature for 1 h. Protein bands were finally visualized using an Ultra High Sensitivity ECL Kit (Cat. No. E412-01/02; Vazyme, Nanjing, China).
2.5. Extracellular Proteome Analysis
The wild type P. brasiliense NJAU180 and its derivative strains were co-cultured with 0.6 g aseptic, whole-plant potato grown plantlets in 20 mL MM medium, respectively. To avoid interference from intracellular proteins resulting from cell lysis, extracellular proteins were obtained from the supernatant of the strains after rapid interaction with the plantlets for 3 h at an initial concentration of OD_600_ = 0.1. Three independent biological replicates were prepared for each strain.
Protein samples were reduced, alkylated, and digested with Trypsin at a 1:50 (w/w) enzyme-to-substrate ratio. The resulting peptides were separated using an EASY nLC-1200 UHPLC system coupled online to a timsTOF Pro2 mass spectrometer. Following separation on a C18 column with an acetonitrile gradient, peptides were ionized at 1.5 kV and analyzed using the PASEF method [27]. The acquired MS/MS spectra were searched against the UniProt database using MaxQuant, with a false discovery rate (FDR) of <1% for peptide identification.
For label-free quantification, normalization based on total peptide intensity was performed to correct for sample loading variations and technical fluctuations. Specifically, peptide intensities in each sample were divided by the sum of all peptide intensities in that sample and multiplied by the average total intensity across all samples. Normalized intensities were used for subsequent statistical analysis.
To identify differentially expressed proteins (DEPs), a two-step screening strategy was employed. For initial screening, proteins with |fold change| > 1.2 and p < 0.05 were considered as potentially differential to ensure no candidates were missed. To obtain a more stringent list of candidates for downstream functional validation, the threshold was further raised to |fold change| > 2.0, combined with multiple testing correction.
Statistical analysis was performed using R software (version 4.2.1, R Foundation for Statistical Computing, Vienna, Austria) packages. Protein abundance differences between groups were compared using Student’s t-test. To control for multiple hypothesis testing, p-values were adjusted using the Benjamini–Hochberg method to control the false discovery rate (FDR). Proteins with |fold change| > 2.0 and adjusted p-value < 0.05 were defined as significantly differentially expressed.
Functional annotation of all identified proteins was performed using Gene Ontology (GO, http://geneontology.org/, accessed on 15 March 2025) and KEGG (http://www.genome.jp/kegg/, accessed on 15 March 2025) databases. Significantly, DEPs were further subjected to GO functional enrichment analysis and KEGG pathway enrichment analysis [28].
2.6. PurK Auxotroph Phenotype Validation Under Purine Supplementation
To evaluate the purine auxotrophy of the wild-type strain P. brasiliense NJAU180 and the purK mutant, the following procedure was employed. Both strains were initially cultured overnight at 28 °C in LB medium, then subcultured at a 1:100 ratio into fresh LB and grown to an OD_600_ of 1.0. Cells were harvested, washed, and resuspended in sterile water. Stock solutions of adenine, guanine, and hypoxanthine were prepared at a concentration of 3.46 g/L each. These stocks were incorporated into M9 minimal agar plates to achieve final purine concentrations of 27.68, 13.84, 6.92, 3.46, 1.73, and 0.86 mg/L, respectively. An M9 plate without any purine supplement served as the negative control. Resuspended cell suspensions were spotted onto the series of plates and incubated at 28 °C for 48 h, after which growth was visually assessed to compare the purine-dependent growth capacities of the two strains.
2.7. Pull-Down Analysis
Pull-down assays were performed as previously described [27]. Briefly, the His-purK expression plasmid was constructed by cloning the fragment of purK into the pET30a vector, while the fragment of purE was fused with a GST tag using the pGEX-6p plasmid. All recombinant plasmids were transformed into E. coli BL21 (DE3) competent cells. The transformed strains were cultured overnight at 37 °C, then diluted 1:100 into fresh LB medium and grown at 37 °C until the OD_600_ reached 0.4. Protein expression was induced with 0.1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) at 28 °C for 4 h. After centrifugation, the cell pellets were resuspended and lysed in 0.01 M PBS buffer (pH 7.4), and the supernatant was collected and concentrated. The concentrated tagged protein samples were incubated with 50 μL of GST affinity beads (GE Healthcare, Shanghai, China) in a final volume of 1 mL adjusted with 0.01 M PBS (pH 7.4) and incubated overnight at 16 °C. The beads were then washed six times with PBS buffer containing 1% Triton X-100 to remove non-specifically bound proteins. Finally, the bound proteins were eluted using glutathione elution buffer and analyzed by Western blotting. His-tag monoclonal antibody (M30111L) and GST-tag monoclonal antibody (M20007L) were used for detection.
2.8. Protein Structure Simulation
Three-dimensional homology models of full-length PurK and its N-terminal, M-terminal, and C-terminal domains were generated using the SWISS-MODEL server (https://swissmodel.expasy.org, accessed on 15 March 2025). The optimal model for each target sequence was selected based on the Global Model Quality Estimate score. Structural comparisons and visualizations of the modeled structures were performed using PyMOL software (version 2.5.0, Schrödinger, LLC, New York, NY, USA) [26].
2.9. Phylogenetic Tree Analysis
The amino acid sequences of homologous proteins from P. brasiliense NJAU180 and other representative strains, including P. polaris, P. aquaticum, E. coli, Saccharolobus solfataricus, Brucella canis, Staphylococcus aureus subsp. aureus, and Acinetobacter baumannii, were selected for phylogenetic analysis. Multiple sequence alignments were performed using ClustalX software (version 2.1, University College Dublin, Dublin, Ireland), and the resulting alignments were manually refined where necessary before being exported in FASTA format. The phylogenetic tree was constructed using the neighbor-joining method based on the neighbor-joining model with MEGA software (version 11.0, Pennsylvania State University, University Park, PA, USA). Visualization and layout optimization of the resulting phylogenetic tree were also conducted using MEGA software (version 11.0, Pennsylvania State University, University Park, PA, USA) [26].
2.10. Motility Assay
Swimming motility assays were performed as previously described [23]. Briefly, bacterial strains were cultured to logarithmic phase (OD_600_ = 1.0). A total of 2 μL aliquot of the bacterial suspension was spot-inoculated onto the center of MM plates containing 0.3% agar. After incubation at 28 °C for 18 h, the migration zone diameter was determined and photographed. For comparative purposes, all measured data were normalized to the mean of the control group.
2.11. The Activity of Plant Cell Wall Degrading Enzymes (PCWDEs) Assay
PCWDE activity of wild-type NJAU180 and its derivate strains was analyzed as previously described [25]. Bacterial cultures were grown to an OD_600_ of 1.0, spot-inoculated onto PCWDE activity assay plates independently, and incubated at 28 °C for 48 h. PCWDE production was evaluated by measuring the halos formed around the bacterial colonies.
2.12. Bacterial Two-Hybrid Analysis
The bacterial two-hybrid assay was performed following established methods [26]. The coding sequences of PurL, GuaA (NJAU180_1200), PurE, PurF, PurN, PurH, PurC, PurD, PurM, GuaA (NJAU180_1433), ThiC, and GuaA (NJAU180_1471) were cloned into the pTRG vector as previously described, while the fragment of purK was constructed into the pBT vector. The recombinant pTRG plasmids and purK-pBT were co-transformed into E. coli XL1-Blue competent cells. To validate protein–protein interactions, transformed cells were plated on selective medium containing 3-amino-1,2,4-triazole (3AT) and streptomycin. Protein interactions between PurK and each target protein were assessed based on bacterial growth.
2.13. Callose Deposition Assay
Callose deposition assays in Nicotiana benthamiana leaves were performed as previously described [23,29]. Wild-type strain P. brasiliense NJAU180 and its derivate strains were cultured to an OD_600_ of 1.2 and inoculated into leaves of 6-week-old greenhouse-grown N. benthamiana plants via syringe infiltration. After 18 h of incubation, the leaves were harvested and decolorized completely with 95% ethanol at 37 °C, followed by two washes with 70% ethanol and two washes with sterile water. The decolorized leaves were stained in 0.01% (w/v) aniline blue solution prepared in 150 mM K_2_HPO_4_ (pH 9.5) for 5 h in darkness. The stained samples were observed and imaged using an epifluorescence microscope (Olympus Corporation, Center Valley, PA, USA). Sterile water and Pseudomonas syringae pv. tomato DC3000 were used as controls.
2.14. Hypersensitive Response (HR) Assay
The hypersensitive response (HR) assay was performed as previously described [29]. Briefly, wild-type strain P. brasiliense NJAU180 and its derivate strains were cultured to an OD_600_ of 1.0, and the bacterial suspensions were infiltrated into the leaves of N. benthamiana using a sterile syringe. After inoculation, plants were maintained at 24 °C for 16 h, and the diameters of necrotic lesions were measured and photographed.
2.15. Agrobacterium-Mediated Transient Expression Assays
Agrobacterium-mediated transient expression assays were performed as previously described [30] with minor modification. Briefly, the fragment of target gene purK was cloned into the pBinGFP vector, and the resulting recombinant plasmid was transformed into Agrobacterium tumefaciens strain GV3101. After incubation, bacterial cells were harvested and washed twice with 10 mM MgCl_2_, then resuspended in infiltration buffer (10 mM MgCl_2_, 100 mM MES, 200 μM acetosyringone). The bacterial suspension was adjusted to an OD_600_ of 0.5 and incubated in the dark at 28 °C for 2 h without shaking. The prepared suspension was infiltrated into the leaves of N. benthamiana plants using a needleless syringe. Inoculated plants were maintained under 24 °C for 24 h before qRT PCR experimental analyses.
2.16. Flagella Staining
Flagellar staining was performed to observe the flagellar morphology of different strains. The tested strains were inoculated into LB liquid medium and cultured at 28 °C with shaking until the logarithmic growth phase. An appropriate volume of fresh bacterial culture was collected by centrifugation, and the cell pellets were washed twice with sterile water before being resuspended to a final concentration of approximately 10^8^ CFU/mL. A total of 10 μL aliquot of the bacterial suspension was applied to one end of a clean glass slide and air-dried naturally. The smear was then covered with flagellar staining solution and incubated at room temperature for 5–10 min. The staining solution was gently rinsed off with distilled water, and the slide was air-dried naturally. Flagellar morphology was observed under an oil immersion lens using an upright fluorescence microscope, and images were captured. At least 20 random fields were examined for each strain, and representative images were selected to illustrate flagellar characteristics.
2.17. Statistical Analysis
Each assay described above was repeated at least three times with 3–5 replicates in each.
Data on extracellular enzyme activities, pathogenicity, motility, and callose deposition are presented as the mean ± standard deviation (SD) (n = 3). Statistical analysis was performed using GraphPad Prism (version 9.0, GraphPad Software, Inc., San Diego, CA, USA) with one-way ANOVA followed by Dunnett’s post hoc test, with the wild-type strain NJAU180 serving as the control group, and differences were considered statistically significant at p < 0.05. Data from qRT-PCR and purine validation assays are presented as the mean ± standard deviation (SD) (n = 3). Statistical analysis was performed using GraphPad Prism (version 9.0, GraphPad Software, Inc., San Diego, CA, USA) with Student’s t-test (two-tailed). Data were confirmed to meet the assumptions of normality before analysis. Differences were considered statistically significant at p < 0.05.
3. Results
3.1. Inducement of Aseptic Grown Potato Plantlets on the Proteins Pectobacterium brasiliense NJAU180 Secreted Outside Membrane
Previous studies [31,32] have shown that pathogenic bacteria secrete many small molecule compounds into host plants in the course of infection. Our previous research demonstrated that plant extracts significantly induce the expression of virulence genes in P. aroidearum PccS1 [32].
In the present work, P. brasiliense NJAU180, isolated from potato soft rot samples in Inner Mongolia, China, was selected to analyze the exo-secreted proteins in the supernatant of NJAU180 cultured with aseptic grown potato plantlets.
Using a threshold of a fold change greater than 2.0 (i.e., log_2_FC > 1.0), compared to the wild-type strain cultured in minimal medium (MM), the supernatant of P. brasiliense NJAU180 cultured in MM supplemented with aseptic grown potato plants (NJAU180^plant^) showed a significant accumulation increase in 54 proteins and a decrease in 1268 proteins, indicating that the secretion of these proteins was markedly induced or suppressed during interaction with potato (Table S3).
To further focus on the exocrine proteins induced by the host, we implemented a stricter threshold of log_2_FC > 4.0. Under this criterion, compared to the sample of P. brasiliense NJAU180 cultured in MM only, the group from NJAU180^plant^ exhibited significantly enhanced secretion of 31 proteins and suppressed secretion of 694 proteins (Table S3).
3.2. PurK, a Potato Plant-Promoted Secretion Protein Conserved in Bacteria and Containing an ATP-Grasp Domain, Is Crucial to Pectobacterium brasiliense NJAU180 Virulence
Among the 31 plant-promoting exoproteins showing upregulated secretion (Table S3), there are seven proteins that exhibited significantly higher extracellular abundance compared to their counterparts in P. brasiliense NJAU180 cultures grown exclusively in MM. We selected 24 proteins which were detected only in the supernatant of NJAU180^plant^ rather than in that of NJAU180 cultured solely in MM; the later ones will be dealt with in the future works. Here, we selected these seven candidate proteins to investigate their potential roles in mediating the plant-enhanced exocrine protein effects on P. brasiliense NJAU180 virulence. Among the genes encoding these seven proteins, only deletion of purK resulted in a significant reduction in bacterial virulence, whereas deletion of any of the other genes did not affect the virulence of P. brasiliense NJAU180 (Figure S1).
Therefore, PurK was selected for further analysis to validate the proteomic findings and to establish a foundation for more detailed investigations into its pathogenicity. The 3 × FLAG tag, characterized by its small size and high hydrophilicity, minimizes interference with protein function, secretion, or localization while preserving native expression regulatory features, and has been widely used in bacterial studies [33]. In parallel, Western blot analysis of TCA-precipitated culture supernatants is a well-established technique for investigating protein secretion in bacterial systems [34,35,36]. Based on this, we complemented the strains with a plasmid carrying the purK gene fused to a C-terminal 3 × FLAG tag (DYKDDDDK) and employed the aforementioned method to assess the intra- and extracellular distribution of PurK.
The results showed that PurK could be detected both inside and outside the wild type P. brasiliense NJAU180 cultured solely in MM, albeit a significantly lower proportion showed in extracellular. Interestingly, in the sample of NJAU180^plant^, there is a notable increase in extracellular accumulation of PurK, despite the intracellular levels remaining comparable to those observed in that of NJAU180 cultured in MM only (Figure 1A). These observations confirm that PurK is one of the exocrine proteins promoted by the plants.
Bioinformatic prediction using SignalP-5.0 revealed that PurK lacks a canonical N-terminal signal peptide (Sec/SPI probability = 0.0085; OTHER probability = 0.9913), suggesting it is not secreted via the classical Sec or Tat pathways. Consistent with this, SecretomeP 2.0 prediction for non-classical secretion yielded a low score (0.0001, below the threshold of 0.5), indicating that PurK does not conform to the defined model of constitutive non-classical secretion. However, Western blot analysis detected low but clear PurK signals in the culture supernatant under standard laboratory conditions (Figure 1A), contrasting with the bioinformatic predictions. This discrepancy suggests that PurK may be exported via an unconventional mechanism not captured by current prediction algorithms, which are primarily trained on constitutively secreted proteins. Notably, extracellular PurK abundance significantly increased upon co-culture with potato plantlets (Figure 1A), indicating that this unconventional secretion pathway is responsive to host-derived signals. These findings suggest that PurK may utilize a previously uncharacterized, host-inducible secretion mechanism that maintains basal activity even in the absence of host stimulation. Furthermore, it also reinforces the notion that plant-induced factors predominantly influence the secretion of PurK in Pectobacterium NJAU180.
PurK of P. brasiliense NJAU180 is a highly conserved protein, the deduced amino acid sequence exhibits 97–99% identity with those in the other two species of Pectobacterium, approximately 80% homology with that of E. coli, and lower with the others (Figure 1B,C). Among these proteins, the N-termini (highlighted with a blue underline) and ATP-grasp domains (indicated by a green underline) exhibit a high degree of conservation, while their C-termini (marked with a yellow underline) display significant variation (Figure 1B). Structural simulations suggest that PurK functions as a dimer, composed of two homodimers that band ADP through the ATP-grasp domain present in each unit (as indicated by the red arrow in Figure 1D).
3.3. The Gene purK Affects the Virulence by Influencing Motility Rather than Plant Cell Degrading Enzymes in Pectobacterium brasiliense NJAU180
The virulence assay indicated that the maceration ability of the strain ΔpurK was significantly lower than that of the wild type P. brasiliense NJAU180, and the decreased virulence of ΔpurK could be restored by complementing the strain with the vector that carries purK fragment, rather with an empty vector (Figure 2A,B-i and Figure S1), though the multiplication of ΔpurK in LB medium showed no difference with the wild type (Figure 2C), indicating that the gene purK is associated with virulence.
Plant cell wall degrading enzymes (PCWDEs) are widely recognized as the primary virulence factors in Pectobacterium [23]. However, the assays of PCWDEs activity showed that the derivate strains from NJAU180, including ΔpurK, ΔpurK (purK), and ΔpurK (pBBR), exhibited similar levels of pectate lyase (Pel), celluase (Cel), and protease (Prt) activities compared to the wild-type NJAU180 (Figure 2A,B-ii–B-iv). The findings suggest that the virulence gene purK may employ a distinct mechanism in the modulation of virulence in P. brasiliense NJAU180.
How does purK influence the virulence? Previous studies reveal that motility acts as an auxiliary virulence factor in many pathogenic bacteria [23,31]. Thus, the swimming motility of P. brasiliense NJAU180 and its derivates was assessed on MM containing 0.3% agar. The results demonstrated that ΔpurK nearly completely lost its motility, and this ability can be restored in the ΔpurK strain when complemented with a vector carrying the purK fragment, whereas the strain with the empty vector remained unchanged (Figure 3A,B).
The bacterial flagellum is a sophisticated organelle that plays a crucial role in bacterial movement, allowing them to swim actively towards host plants. Through chemotactic regulation, bacteria can navigate effectively, directing their movement towards nutrients and chemical signals released by the hosts. We first investigated flagellar morphology through staining, as flagellar assembly is essential for motility. Our observations revealed that the deletion of purK did not interfere with normal flagellation (Figure 3A). To understand how the purK gene influences bacterial motility, we measured the expression levels of 19 flagellar genes in both wild-type P. brasiliense NJAU180 and ΔpurK. The results revealed that in ΔpurK, expression of fliQ did not differ significantly from the wild-type NJAU180, whereas the other 18 genes were markedly upregulated, suggesting that purK acts as a negative regulator of flagellar gene expression at the transcriptional level (Figure 3C).
To assess the impact of these upregulated genes on bacterial motility, we overexpressed them in the wild-type P. brasiliense NJAU180 and analyzed their motility on the MM plates containing 0.3% agar. The results revealed that these strains overexpressing the purK gene significantly reduced bacterial swimming capacity (Figure 3D,E). This finding reveals a mechanism by which purK negatively regulates the expression of flagellar genes, thereby enhancing bacterial motility and promoting successful infection.
3.4. PurK Modulates the Virulence Through a Mechanism Operating Independently of Purine Biosynthesis
Purine is an essential precursor for the synthesis of key biomolecules such as nucleic acids and ATP in microbial cells. The de novo synthesis of purine nucleotides is a highly conserved multi-step metabolic pathway, in which the purK gene encodes phosphoribosylaminoimidazole carboxylase, an enzyme responsible for catalyzing the early rate-limiting step of converting CAIR to N^5^-CAIR, representing a critical node in the IMP biosynthesis pathway [19]. To elucidate the metabolic function of purK and its association with pathogenic phenotypes in NJAU180, this study systematically validated its role through exogenous purine supplementation experiments.
First, we assessed the growth capacity of the purK mutant in a chemically defined M9 minimal medium. Compared to the wild-type strain, the purK mutant completely failed to grow on M9 agar plates without exogenous purine supplementation, indicating a severe disruption in its purine synthesis pathway, consistent with a typical purine auxotrophic phenotype. To further dissect the specific step of this metabolic defect, we prepared a series of solid agar plates supplemented with varying concentrations of adenine (Ade), guanine (Gua), or hypoxanthine (Hyp). The results demonstrated that the purK mutant could not restore growth at any concentration of guanine, whereas on plates containing higher concentrations (13.84 mg/L and 27.68 mg/L) of either adenine or hypoxanthine, the mutant regained growth comparable to that of the wild type P. brasiliense NJAU180 (Figure 4A). These findings clearly indicate that the absence of purK disrupts IMP synthesis, leading to a complete loss of de novo purine synthesis capability in the strain, thereby confirming the indispensable role of purK in purine metabolism in NJAU180.
Having established the metabolic function of purK, we further investigated whether its influence on bacterial pathogenicity is mediated through the provision of purine precursors. The purK mutant was treated with 13.84 mg/L adenine and hypoxanthine, restoring its basic metabolic capacity to a level equivalent to that of the wild type, and its virulence and motility were subsequently evaluated. The results showed that although exogenous purine supplementation fully rescued the growth defect of the purK mutant on M9 medium, it did not restore its pathogenic capability toward the host or its motility (Figure 4B). This finding suggests that the role of purK in the pathogenicity of NJAU180 extends beyond merely supplying purine precursors for bacterial proliferation. Its contribution to virulence and motility appears to be independent of its metabolic synthesis function.
Studies have demonstrated that purK and purE are two functionally synergistic key genes in the bacterial purine biosynthesis pathway [37]. They jointly encode an enzyme system that catalyzes the same critical step: the carboxylation of AIR (5-aminoimidazole ribonucleotide) to CAIR (5-aminoimidazole-4-carboxylate ribonucleotide) [37]. To determine whether purK and purE have analogous roles during P. brasiliense NJAU80 infection, we initially carried out bacterial two-hybrid and pull-down assays. The results confirmed that PurK specifically interacts with PurE (Figure 5A,B).
To investigate the functional relationship between purK and purE in pathogenicity, a purE gene knockout mutant was constructed for virulence assay; the results showed that DpurE exhibited no significant difference in virulence compared to the wild type (Figure 5C,D), indicating that purE is not essential for virulence in P. brasiliense NJAU180. This aligns with previous observations that the loss of pathogenicity in ΔpurE likely arises from an independent role of PurK itself, rather than from its canonical function in purine biosynthesis.
The phenomenon of a metabolic enzyme possessing additional, non-metabolic functions is well established in bacteriology. Pancholi and Fischetti [38] first demonstrated that the glycolytic enzyme GAPDH functions as a surface adhesin and virulence factor in Streptococcus. Subsequent reviews have shown that such “moonlighting proteins”—including GAPDH, enolase, and chaperonins—are employed by over 90 bacterial species as virulence determinants [39,40]. Our finding that PurK plays dual roles in purine metabolism and virulence regulation, with its extracellular accumulation induced by plants, is consistent with the emerging paradigm of moonlighting proteins, raising the possibility that PurK may function as a novel moonlighting protein in P. brasiliense NJAU180.
3.5. Every Domain of PurK Is Indispensable for Virulence and Motility
Our structural simulation results suggest that PurK functions as a dimer, comprising two homodimer units. These units interact with ADP through the ATP-grasp domain located at the center. Notably, each unit possesses a conserved N-terminus and a variable C-terminus. (Figure 1B,C). To identify the key regions responsible for pathogenicity and motility, we constructed single domain knockout mutants targeting the C-terminal (purK-C), N-terminal (purK-N), and ATP-grasp (purK-M) regions, respectively. The virulence assays showed that removing any domain significantly reduced the virulence of P. brasiliense NJAU180, while complementation with the corresponding fragments restored the maceration ability and motility of the strains (Figure 6A,B). This functional recovery indicates that each of the constructed domains is properly and stably expressed [41,42,43]. However, compared with the other two mutants, the strain with a mutation in the C-terminal domain fragment exhibited a lesser reduction in virulence (Figure 6A,B).
Meanwhile, the motility assays revealed that the deletion of purK-C reduced bacterial motility, and the strains of both purK-N and purK-M impaired completely abolished motility, and the defects of motility were fully restored upon genetic complementation with the corresponding domains (Figure 6C,D). These results demonstrate that the structural integrity of purK is essential for the full pathogenicity and motility of P. brasiliense NJAU180. The N-terminal, C-terminal, and ATP-grasp domains function synergistically as an integrated functional unit in regulating virulence and motility, rather than serving redundant roles. Notably, the N-terminal and ATP-grasp domains are crucial in these processes. Their loss results in more pronounced phenotypic defects, suggesting that they serve essential functions in critical biochemical processes, such as purine metabolism or energy supply.
3.6. Extracellular PurK Suppresses Plant PTI Responses
Previous studies have shown that pathogenic bacteria secrete effectors into host cells to target and suppress plant basal immunity, known as PTI, facilitating their infection and colonization of the plant [1,8,31]. In response, plants have evolved a multi-layered immune system to recognize pathogenic signals and activate defense mechanisms, including PTI mediated by pattern recognition receptors, as well as effector-triggered immunity (ETI), mediated by intracellular resistance proteins. To explore the role of the host-induced exocrine protein PurK in enhancing plant immunity, the observations of hypersensitive response (HR) and callose deposition were carried out with the wild type P. brasiliense NJAU180 and its derived strains, the results showed that ΔpurK failed to elicit HR, but induced significant callose deposition in the N. benthamiana leaves (Figure 7A,B). The phenotypes of ΔpurK (purK) closely resembled those of the wild-type, suggesting that PurK is essential for inhibiting callose deposition and triggering HR (Figure 7B). To confirm whether PurK activates PTI responses, we transiently expressed PurK in tobacco leaves and detected the transcript levels of PTI downstream marker genes (PR2, PR2b, PR5, and WRKY7) 18 h after inoculation. The results of qRT-PCR demonstrated that the expression of these genes was significantly upregulated (Figure 7C), suggesting that PurK strongly induces PTI-associated immune responses.
4. Discussion
PurK, A Plant-Induced Extracellular Protein, Suppresses Plant Immune Responses and Enhances Virulence by Promoting Motility with Its Three Domains
In this study, the supernatant of P. brasiliense NJAU180 co-cultured with aseptically grown potato plantlets in MM (NJAU180^plant^) was collected during the early growth stage for exoproteomic analysis. These profiles were subsequently compared with those derived from NJAU180 cultures grown solely in MM. The results of these analyses indicated that the supplementation of host plants in the medium significantly enhanced both the quality (Figure 1A) and diversity (Table S3) of exocrine proteins. It was found that only the strain without the fragment of purK significantly lost the maceration ability (Figure S1). In the previous study, PurK, N^5^-carboxyaminoimidazole ribonucleotide synthetase, has been demonstrated to play a crucial role in the de novo purine biosynthesis pathway [37,44], and it is a virulence-associated gene coding for factors involved in global bacterial physiology in Yersinia pestis [45]. Our findings reveal that PurK is a novel plant-induced secreted protein (Figure 1A). It suppresses host callose deposition while simultaneously triggering a hypersensitive response and upregulating the expression of plant immunity marker genes such as PR2 and WRKY7 (Figure 6). PurK contributes to the virulence of NJAU180 by enhancing bacterial motility (Figure 3A) modulated through its three domains as an integrated unit to infect successfully (Figure 5). These observations provide new insights into the pathogenic mechanism of P. brasiliense NJAU180 and establish an experimental system for studying microbe–plant interactions.
The Experimental Model of Pathogen Interaction with Aseptic Grown Plantlets Established in This Study Provides A Critical Framework for Analysing Bacterial Extracellular Proteins.
In natural environments, plants form complex symbiotic relationships with rhizospheric and phyllospheric microbiomes, which significantly influence pathogen infection processes and plant immune responses through mechanisms such as competition, antagonism, and immune modulation [46]. To investigate the key responses required for successful pathogen colonization under undisturbed conditions, in this study a simplified interaction system was established using aseptic grown potato plants (Solanum tuberosum DM1-3) co-cultured with the pathogen, and the exocrine proteins were collected at their early growth phase to avoid the influence of intracellular proteins resulting from pathogenic cell autolysis. This system not only effectively eliminates interference from environmental microbiota, ensuring that observed protein expression changes can be directly attributed to the interaction between pathogen P. brasiliense NJAU180 and the host as showed in the previous work [47], but also preserves the host’s tissue structure and fundamental physiological processes, thereby more accurately simulating natural infection scenarios—particularly suitable for focusing on molecular events during early infection stages [48].
Furthermore, we found that P. brasiliense NJAU180 exhibits slow growth in minimal medium (MM), and prolonged cultivation can easily induce cell autolysis. Concurrently, aseptic cultured seedlings may develop minor damage, both of which could introduce non-specific protein interference and compromise the accuracy of subsequent proteomic analyses. Therefore, this study focused on the early stage of pathogen–host interaction (3 h) to minimize background noise caused by plant tissue degradation and bacterial autolysis, ensuring that the detected extracellular protein profile authentically reflects biological processes related to infection [31], which is different from the inducement of plant-extract that was thought of as a nutrient [49], and avoids the contamination of the plant extracts from field products that might contain pesticides widely used in agriculture [50].
Inducement of Aseptic Grown Potato Plantlets on the Proteins Pectobacterium brasiliense NJAU180 Secreted outside Membrane/Outer Membrane Proteins as Pioneer Signals in Microbiota–Host Interactions.
Through exoproteomic analysis, this study revealed that during the early stages of pathogen infection, the host plant significantly suppresses the expression of multiple bacterial extracellular proteins. These suppressed proteins are primarily enriched in key pathways, including carbohydrate metabolism, amino acid metabolism, and membrane transport systems, as well as cofactor and vitamin metabolism (Table S3). These findings suggest that the host plant may actively interfere with the fundamental metabolism and transport capacity of the pathogen during early interaction, thereby limiting its pathogenicity.
Specifically, the host’s suppression of key enzymes involved in sugar and amino acid metabolism directly impairs the pathogen’s energy supply and biosynthesis of precursors, effectively disrupting its “metabolic engine”. This leads to energy deficiency and insufficient building blocks, hindering the synthesis of virulence-associated proteins and consequently inhibiting pathogen growth and reproduction [51]. Concurrently, the inhibition of membrane transport-related proteins severs the pathogen’s channels for nutrient uptake, toxin efflux, and signal communication, isolating it in a “metabolic island” within the host environment and impairing its ability to coordinate group behaviors and infection progression [52]. Furthermore, interference with cofactor and vitamin metabolism pathways affects multiple core biological processes, including energy metabolism, amino acid synthesis, and DNA replication, thereby establishing a multi-target defense network at the systemic level [53].
On the other hand, a total of 31 proteins were significantly upregulated under host induction, with functions mainly related to ABC transporters, purine synthesis, and partial carbohydrate metabolism pathways. This indicates that, in response to host-imposed nutritional immunity pressure, the pathogen enhances its active uptake of critical nutrients such as sugars, amino acids, and metal ions to sustain viability and virulence [54]. Notably, PurK, a key enzyme in the purine synthesis pathway, was markedly upregulated under host induction, suggesting its potential important role in the pathogen’s adaptation to the host environment and the establishment of a successful infection.
PurK Modulates the Flagellar Gene Expression to Control Motility for Successful Infection, rather than Stimulates PCWDEs in the Cells of Pectobacterium brasiliense NJAU180.
Previous studies have generally established that for soft rot pathogens such as Pectobacterium spp., extracellular enzymes (e.g., pectinases and cellulases) are key virulence factors, directly degrading plant cell wall structures and leading to tissue maceration and rot symptoms [55]. In this classical model, motility is often regarded as an accessory factor, primarily facilitating the later stages of pathogen spread and colonization within host tissues [56]. However, through systematic phenotypic analysis of the purK mutant, this study reveals a distinct pathogenic mechanism in P. brasiliense NJAU180. Extracellular enzymes are not the primary route through which purK regulates virulence. Instead, PurK significantly restrains bacterial motility by negatively regulating the expression of flagellar synthesis and related genes at the transcriptional level, and this impairment in motility is the key factor responsible for the marked reduction in pathogenicity. These findings suggest that, in NJAU180, efficient motility is not merely an accessory factor for colonization but may serve as an essential prerequisite for the pathogen to successfully locate and breach host physical barriers, thereby establishing initial infection.
Notably, the majority of flagellar genes were significantly upregulated at the transcriptional level in the ΔpurK mutant, yet its swimming motility was markedly impaired. This apparent “transcription-up—phenotype-down” paradox suggests the existence of complex post-transcriptional or post-translational regulatory mechanisms intervening between flagellar gene expression and functional motility apparatus assembly. This seemingly paradoxical phenomenon has been previously documented. Dasgupta et al. [57] reported that inactivation of fleN in Pseudomonas aeruginosa resulted in upregulation of multiple flagellar promoters (e.g., flgBCDE by 27-fold, fliLMNOPQ by 5.6-fold) and loss of directional motility, although this was accompanied by hyper-flagellation (3–6 flagella per cell). In the present study, flagellar staining revealed no significant difference in flagellar number or morphology between the purK mutant and wild-type strains. This indicates that the motility defect in our mutant is not due to gross structural abnormalities in flagellar assembly, but rather to functional impairment of otherwise morphologically normal flagella—a finding that extends the observations of Dasgupta et al.
Based on these findings, we hypothesize that PurK may affect flagellar function through one or more of the following mechanisms: (i) altering translation efficiency or stability of flagellin proteins; (ii) impairing flagellar motor complex assembly or rotation; or (iii) disrupting chemotactic signaling. These potential mechanisms, which appear to operate independently of transcriptional regulation or flagellar assembly, warrant further investigation
The generality of this mechanism across other Pectobacterium strains or soft rot pathogens, and whether motility consistently contributes more significantly to virulence than extracellular enzymes in NJAU180, requires further investigation across broader genetic backgrounds and host environments.
PurK in Pectobacterium brasiliense NJAU180 May Function as An Immune Modulator when Present Extracellularly under Plant Inducement.
Traditionally, PurK has been widely recognized as a highly conserved metabolic enzyme dedicated to the de novo purine biosynthesis pathway. In model organisms such as E. coli, the purK gene is typically located within the pur operon and encodes an intracellular carboxylase that catalyzes an ATP-dependent carboxylation reaction, serving as a key component in the biosynthesis of inosine monophosphate (IMP). However, this study is the first to reveal a significant pathogenic role of PurK beyond its conventional metabolic function in a plant pathogenic bacterium.
Contrary to previous understanding, we found that PurK in P. brasiliense NJAU180 can be secreted extracellularly, a process significantly induced by plant signals, suggesting that it may possess biological activities independent of its metabolic role during pathogen–host interactions.
The detection of extracellular PurK under both basal and host-induced conditions, despite the absence of recognizable secretion signals, points to an unconventional export mechanism. The low SignalP-5.0 and SecretomeP 2.0 scores rule out classical Sec/Tat-dependent secretion and constitutive non-classical pathways as defined by current models [58]. Yet, the clear immunoblotting signals in culture supernatants suggest that PurK employs an export route that falls outside the training space of existing prediction algorithms. This host-responsive secretion pattern is reminiscent of stimulus-coupled export systems described in other bacterial pathogens, in which basal secretion maintains a ready-to-respond pool while host contact triggers rapid mobilization [59]. The identification of PurK as a protein that defies conventional prediction underscores the limitations of relying solely on bioinformatic tools for secretome analysis and highlights the value of empirical approaches, particularly under infection-mimicking conditions. Elucidating the precise mechanism—whether involving outer membrane vesicles, a dedicated secretion system, or other pathways—will require further investigation.
Furthermore, extracellular PurK was shown to play a dual role in plant immune regulation: on one hand, it suppresses plant PTI responses such as callose deposition, while on the other hand, it elicits a strong hypersensitive response (HR) in tobacco and significantly induces the expression of PTI marker genes. These findings imply that secreted PurK may function as a novel effector protein, participating in the complex network of plant immune signaling.
In summary, findings from this study raise the possibility that PurK could have dual functions in bacterial pathogenesis: beyond its canonical role in purine biosynthesis, it may function as a plant-induced secreted protein that influences bacterial motility and virulence through modulation of flagellar gene expression. Additionally, PurK exhibits biphasic immunomodulatory activity, suppressing early PTI responses while later triggering HR and defense gene upregulation. While these findings provide new insights into the complexity of bacterial pathogenicity, we acknowledge that several questions remain—including the precise secretion mechanism, the role of protein stability in domain function, and the plant recognition components involved. Future studies addressing these limitations will further elucidate the multifaceted roles of PurK in pathogen–host interactions. These observations also suggest that other conventional metabolic proteins may possess yet-unexplored non-canonical functions, providing new theoretical insights and potential targets for developing control strategies against plant bacterial diseases.
5. Conclusions
This study, through secretory proteomic analysis, revealed that Pectobacterium brasiliense NJAU180 secretes a plant-induced protein, PurK, during early interaction with its potato host. While PurK is a key enzyme in the purine biosynthesis pathway, we demonstrate that in P. brasiliense NJAU180, it retains its canonical enzymatic function and significantly influences bacterial virulence. Unlike traditional Pectobacterium that primarily rely on extracellular enzymes for pathogenicity, PurK promotes bacterial motility by modulating flagellar gene expression, functioning as an integrated unit through its three distinct domains. Furthermore, extracellularly detected PurK suppresses callose deposition in plants, simultaneously eliciting a hypersensitive response and upregulating the expression of PTI marker genes.
We acknowledge that several questions remain, including the precise secretion mechanism, the role of protein stability in domain function, and the plant recognition components involved. Future studies addressing these limitations will further elucidate the multifaceted roles of PurK in pathogen–host interactions. Nevertheless, this study systematically elucidates the multiple functions of PurK in the pathogenesis of NJAU180, offering novel potential molecular targets for developing disease control strategies against this pathogen.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Charkowski A.O. The changing face of bacterial soft-rot diseases Annu. Rev. Phytopathol.20185626928810.1146/annurev-phyto-080417-04590629958075 · doi ↗ · pubmed ↗
- 2Motyka A. Zoledowska S. Sledz W. Lojkowska E. Molecular methods as tools to control plant diseases caused by Dickeya and Pectobacterium spp: A minireview New Biotechnol.20173918118910.1016/j.nbt.2017.08.01028847714 · doi ↗ · pubmed ↗
- 3Duarte V. de Boer S.H. Ward L.J. de Oliveira A.M. Characterization of atypical Erwinia carotovora strains causing blackleg of potato in Brazil J. Appl. Microbiol.20049653554510.1111/j.1365-2672.2004.02173.x 14962133 · doi ↗ · pubmed ↗
- 4Nabhan S. De Boer S.H. Maiss E. Wydra K. Pectobacterium aroidearum sp. nov., a soft rot pathogen with preference for monocotyledonous plants Int. J. Syst. Evol. Microbiol.2013632520252510.1099/ijs.0.046011-023223819 · doi ↗ · pubmed ↗
- 5Ma B. Hibbing M.E. Kim H.S. Reedy R.M. Yedidia I. Breuer J. Breuer J. Glasner J.D. Perna N.T. Kelman A. Host range and molecular phylogenies of the soft rot enterobacterial genera Pectobacterium and Dickeya Phytopathol.2007971150116310.1094/PHYTO-97-9-115018944180 · doi ↗ · pubmed ↗
- 6Bogino P.C. Oliva M.d.l.M. Sorroche F.G. Giordano W. The role of bacterial biofilms and surface components in plant-bacterial associations Int. J. Mol. Sci.201314158381585910.3390/ijms 14081583823903045 PMC 3759889 · doi ↗ · pubmed ↗
- 7Zheng X.Y. Spivey N.W. Zeng W. Liu P.P. Fu Z.Q. Klessig D.F. He S.Y. Dong X. Coronatine promotes Pseudomonas syringae virulence in plants by activating a signaling cascade that inhibits salicylic acid accumulation Cell Host Microbe 20121158759610.1016/j.chom.2012.04.01422704619 PMC 3404825 · doi ↗ · pubmed ↗
- 8Desaki Y. Miyata K. Suzuki M. Shibuya N. Kaku H. Plant immunity and symbiosis signaling mediated by Lys M receptors Innate Immun.2018249210010.1177/175342591773888529105533 PMC 6830897 · doi ↗ · pubmed ↗