Mazdutide Ameliorates Metabolic Dysfunction-Associated Steatotic Liver Disease by Modulating Endoplasmic Reticulum Stress, Improving Lipid Metabolism and Alleviating Inflammation
Liangyu Gan, Lengxin Duan, Xueyi Zheng

TL;DR
Mazdutide helps treat liver disease by reducing stress in liver cells, improving fat metabolism, and reducing inflammation.
Contribution
Mazdutide's novel therapeutic mechanism in treating MASLD through ER stress modulation and lipid metabolism improvement is identified.
Findings
Mazdutide reduces liver fat and inflammation in MASLD mouse models.
Mazdutide modulates the PERK pathway to alleviate endoplasmic reticulum stress.
Mazdutide downregulates key lipogenic regulators like SREBP-1 and PPARγ.
Abstract
Background: Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD) is the most prevalent chronic liver disorder globally. Mazdutide has shown clinical benefits in weight management and metabolic regulation, indicating its potential as a therapeutic agent for MASLD. This study aimed to investigate the efficacy and mechanism of action of Mazdutide against early-stage MASLD. Methods: A MASLD mouse model was induced by a 12-week high-fat diet, followed by a 4-week treatment with subcutaneous Mazdutide (100, 200, or 400 μg/kg). In vitro, a cellular MASLD model was established by treating hepatocytes with 1 mM free fatty acids for 24 h, followed by co-treatment with Mazdutide (10, 20, or 50 nM) or the endoplasmic reticulum (ER) stress inhibitor 4-phenylbutyric acid (4-PBA). Serum and hepatic lipid profiles, liver injury markers, and pro-inflammatory cytokines were quantified. Liver…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
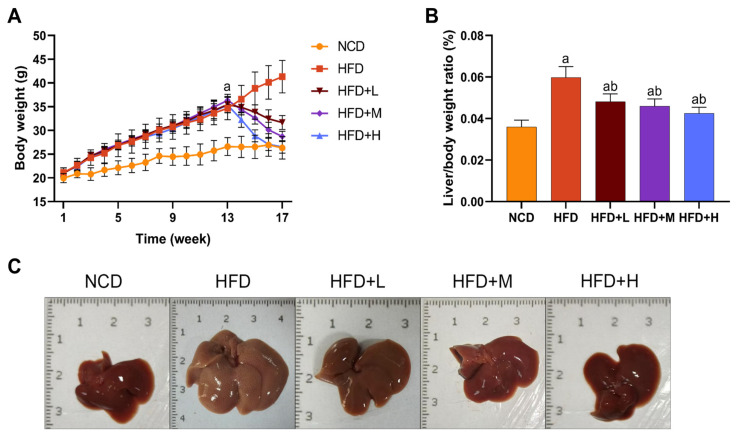 Figure 1
Figure 1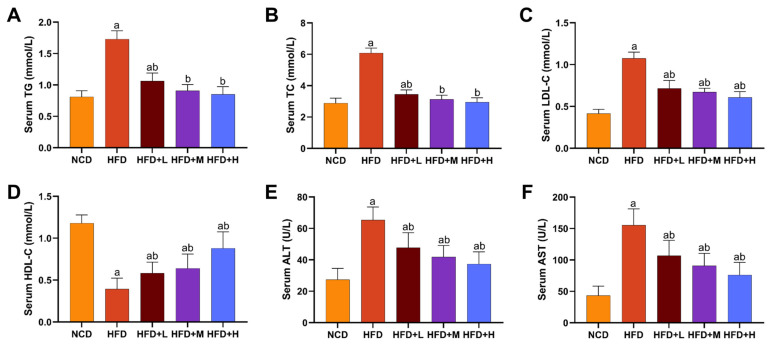 Figure 2
Figure 2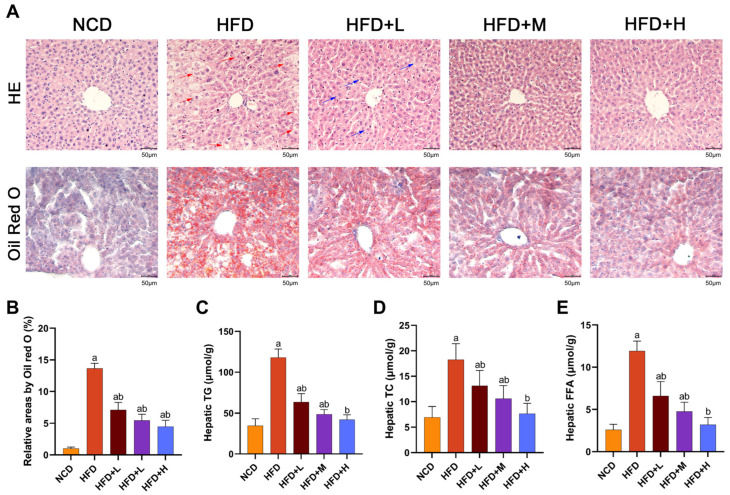 Figure 3
Figure 3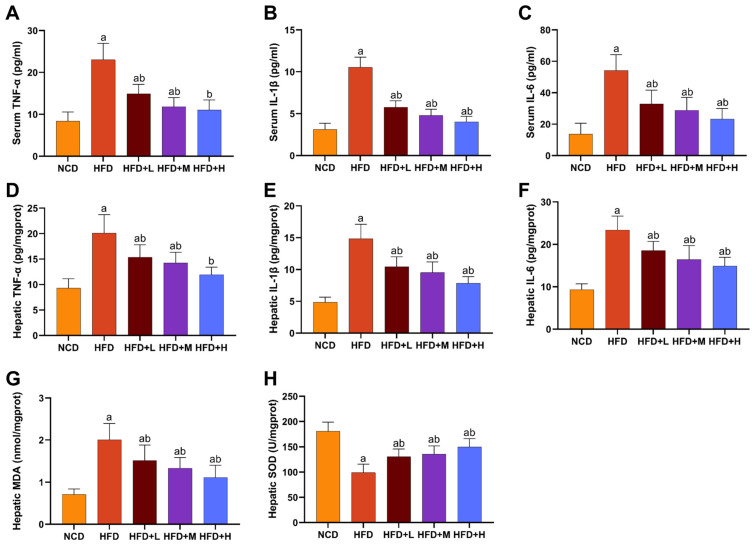 Figure 4
Figure 4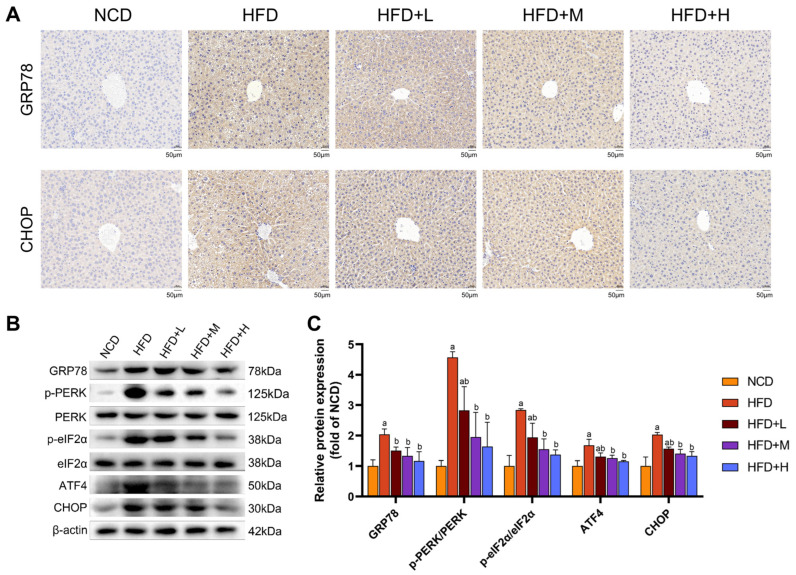 Figure 5
Figure 5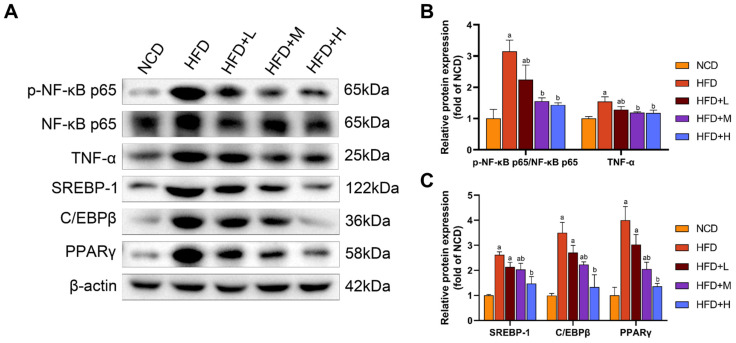 Figure 6
Figure 6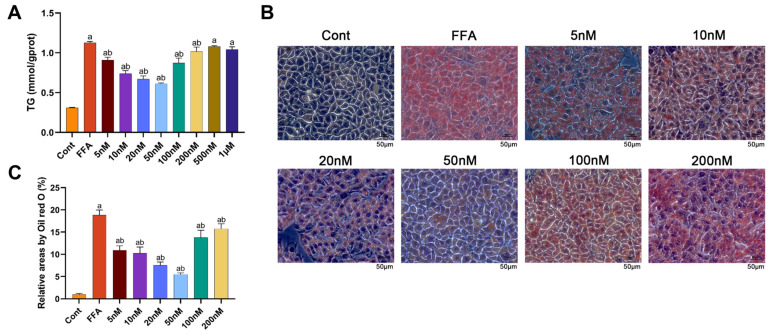 Figure 7
Figure 7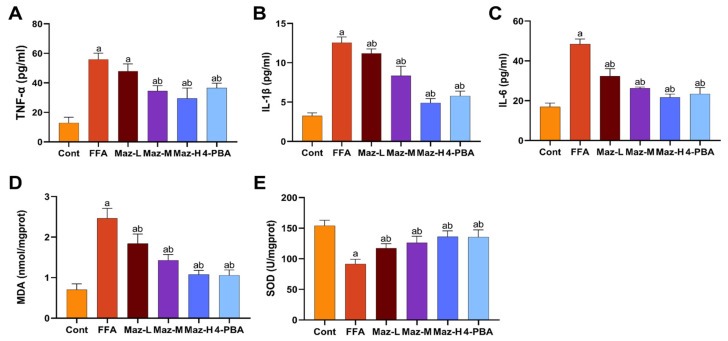 Figure 8
Figure 8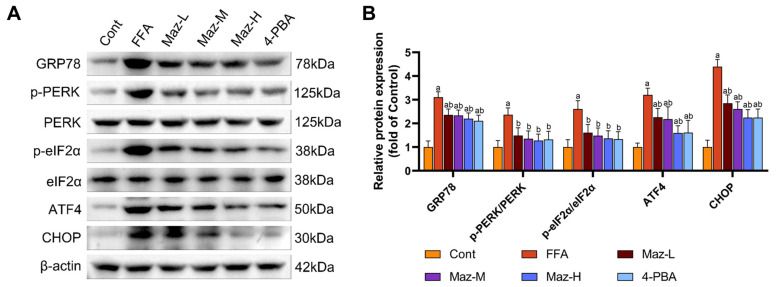 Figure 9
Figure 9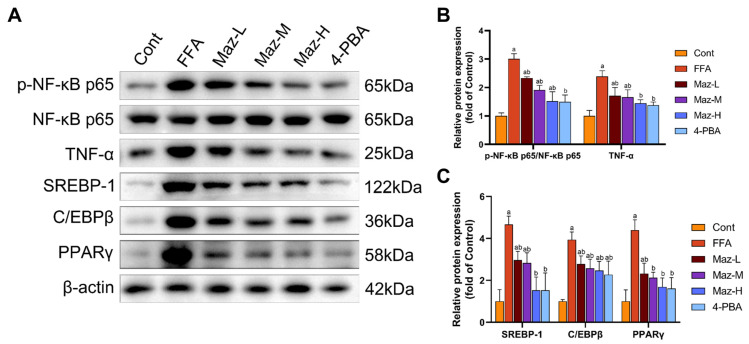 Figure 10
Figure 10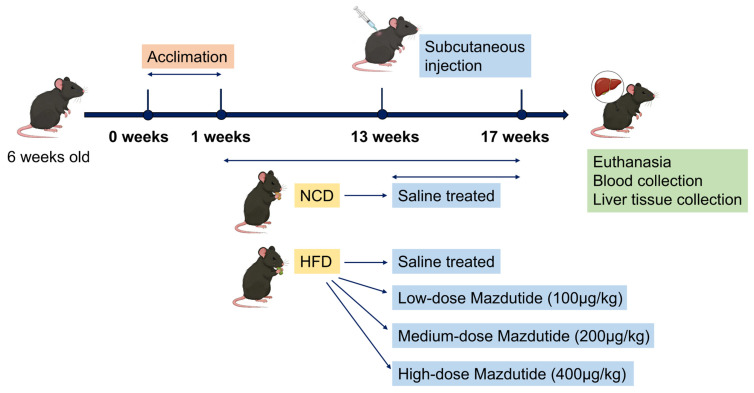 Figure 11
Figure 11- —National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLiver Disease Diagnosis and Treatment · Silymarin and Mushroom Poisoning · Drug-Induced Hepatotoxicity and Protection
1. Introduction
Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD), formerly known as non-alcoholic fatty liver disease (NAFLD), is a chronic liver disease closely associated with metabolic dysfunction, with a global prevalence of approximately 25% [1], making it the most common chronic liver disease worldwide [2]. This terminology update was proposed to more accurately reflect the central role of metabolic dysfunction in the pathogenesis of the disease [3,4]. The MASLD spectrum progresses from simple hepatic steatosis to metabolic dysfunction-associated steatohepatitis (MASH), and further to liver fibrosis, cirrhosis, and even hepatocellular carcinoma [5,6]. Notably, MASLD not only increases the risk of liver-related diseases but is also associated with cardiovascular disease, chronic kidney disease, and certain extrahepatic cancers [7]. Consequently, MASLD has emerged as a major global health issue, imposing a significant socioeconomic burden [3].
In MASLD, excessive lipid accumulation within hepatocytes leads to lipotoxicity, which disrupts endoplasmic reticulum (ER) homeostasis and triggers ER stress. This stress, through sustained activation of the unfolded protein response (UPR), causes dysregulation of hepatic lipid metabolism and exacerbates hepatic fat accumulation [8]. ER stress activates pro-inflammatory signaling and apoptotic pathways through the UPR pathway, suppresses the expression of antioxidant enzymes, and exacerbates hepatic inflammation and oxidative stress [9,10]. The protein kinase R-like endoplasmic reticulum kinase (PERK) signaling pathway is one of the three core pathways of the UPR. In MASLD, excessive lipid accumulation activates PERK, leading to phosphorylation of its downstream signaling molecule eukaryotic initiation factor 2 alpha (eIF2α) and subsequent upregulation of activating transcription factor 4 (ATF4) and C/EBP homologous protein (CHOP) expression. Studies indicate that inhibiting the PERK pathway not only reduces ER stress but also modulates lipid metabolism, alleviates inflammation and oxidative stress [11,12,13], thereby multidimensionally interfering with the pathological progression of MASLD.
Currently, approved drugs for MASLD are limited, necessitating a deeper understanding of disease mechanisms to develop effective therapies. Mazdutide is indicated for the treatment of obesity and type 2 diabetes mellitus (T2DM), demonstrating clinical benefits in weight management and metabolic regulation. Notably, obesity and T2DM are the most prevalent and critical risk factors for MASLD: the global prevalence of MASLD exceeds 70% in obese populations and approximately 50–70% in T2DM patients, and these three diseases share the core pathological mechanisms of insulin resistance, glycolipid metabolic disorder and chronic low-grade inflammation [14,15]. Unlike existing single-target Glucagon-like peptide-1 (GLP-1) receptor agonists such as semaglutide, Mazdutide exerts synergistic metabolic regulation through dual receptor activation: GLP-1 receptor activation enhances insulin sensitivity, suppresses appetite, improves systemic glucose and lipid metabolism, and reduces free fatty acids and de novo lipogenesis in the liver. Glucagon (GCG) receptor activation directly modulates hepatic metabolism by promoting hepatic gluconeogenesis and accelerating lipolysis in adipose tissue, thereby reducing hepatic lipid accumulation directly [16,17]. Mazdutide’s dual-target pharmacology confers superior efficacy compared to single-target GLP-1 agonists in weight reduction, insulin resistance improvement, and lipid metabolism normalization [18,19,20]. Its mechanism directly addresses the pathophysiological mechanisms of MASLD, positioning it as a more rational and promising therapeutic candidate for this disease.
Beyond its well-validated efficacy in weight management and glycemic control, Mazdutide is currently under active clinical evaluation for the treatment of MASLD [21]—a Phase II clinical trial (NCT06937749) is investigating its efficacy and safety in patients with MASH, and a Phase III clinical trial (NCT06884293) is focused on com-paring the efficacy and safety of IBI362 and semaglutide in patients with MASLD among overweight or obese adults in China. However, the specific therapeutic effects of Mazdutide on MASLD and its underlying molecular mechanisms associated with its metabolic regulatory properties remain largely unelucidated, particularly whether its hepatoprotective effects are achieved by modulating endoplasmic reticulum stress pathways and improving lipid metabolism and alleviating inflammation. This study aims to preliminarily elucidate the therapeutic effects and potential mechanisms of Mazdutide on MASLD through in vitro and in vivo experiments, so as to provide an experimental basis and theoretical support for the clinical application of Mazdutide and similar drugs in the treatment of MASLD.
2. Results
2.1. Effects of Mazdutide on Body Weight, Liver-to-Body Weight Ratio, and Liver Morphology in MASLD Mice
The effects of Mazdutide on MASLD progression were assessed by monitoring weekly body weight, terminal liver-to-body weight ratio, and liver morphology. After a 1-week acclimation, initial body weights were comparable across all groups. After 13 weeks, mice on a high-fat diet (HFD) exhibited significantly higher body weight gain compared to those on a normal chow diet (NCD) (Figure 1A). Following 4 weeks of drug treatment, the liver-to-body weight ratio was markedly elevated in the HFD group compared with the NCD group, and this increase was significantly reversed by Mazdutide treatment at all doses compared with the HFD group (Figure 1B). Macroscopically, livers from the NCD group displayed a normal deep-red color and texture. In stark contrast, HFD-fed mice developed pale yellow, enlarged, and friable livers with a granular surface, indicative of severe steatosis. Mazdutide treatment improved these gross morphological alterations (Figure 1C).
2.2. Effects of Mazdutide on Serum Lipids and Liver Enzymes in MASLD Mice
Mazdutide’s effects on lipid metabolism and hepatoprotection were evaluated by measuring serum lipid profiles and liver enzyme activities. The HFD group displayed a dyslipidemic profile, characterized by significantly elevated levels of total cholesterol (TC), triglycerides (TG), and low-density lipoprotein cholesterol (LDL-C), alongside reduced high-density lipoprotein cholesterol (HDL-C), compared to the NCD group. The HFD+L/M/H group significantly reversed these HFD-induced alterations, lowering TC, TG, and LDL-C and increasing HDL-C relative to the HFD group (Figure 2A–D). Serum TG and TC levels in the HFD+M and HFD+H groups returned to the levels of the NCD group (Figure 2A,B). Consistently, serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels were markedly increased in the HFD group compared to the NCD group, indicating hepatic injury. All doses of Mazdutide treatment significantly reduced ALT and AST levels compared to the HFD group (Figure 2E,F).
2.3. Mazdutide Attenuates Hepatic Steatosis and Pathological Injury in MASLD Mice
Histopathology and hepatic lipid accumulation were assessed. Histological analysis by Hematoxylin and Eosin (HE) staining showed normal liver architecture in the NCD group. The HFD group exhibited disorganized hepatocyte arrangement, hepatic steatosis, ballooning degeneration, and hepatocyte edema. Compared with the HFD group, Mazdutide treatment significantly alleviated these pathological changes. Specifically, the HFD+L group showed only mild hepatocyte edema, while the hepatic morphology in the HFD+M and HFD+H groups was close to that of the NCD group. Oil Red O staining consistently revealed markedly increased lipid droplet accumulation in the HFD group compared with the NCD group. Treatment with Mazdutide significantly reduced the Oil Red O-stained area of lipid droplets compared with the HFD group (Figure 3A,B). Liver tissue levels of TG, TC, and free fatty acid (FFA) were significantly higher in the HFD group than in the NCD group. Compared with the HFD group, lipid levels were reduced in the HFD+L/M/H group. There was no significant difference in hepatic TG, TC, and FFA levels between the HFD+H group and the NCD group, this indicates that TG, TC, and FFA levels in the liver of the HFD+H group returned to normal (Figure 3C–E).
2.4. Mazdutide Alleviates Inflammation and Hepatic Oxidative Stress in MASLD Mice
Serum and hepatic levels of the pro-inflammatory cytokines tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), and interleukin-1 beta (IL-1β) were significantly elevated in the HFD group compared to the NCD group. Mazdutide treatment significantly reduced the levels of all three cytokines compared with the HFD group (Figure 4A–F). Consistently, hepatic oxidative stress was markedly induced by HFD, as evidenced by increased malondialdehyde (MDA) content and decreased superoxide dismutase (SOD) activity compared with the NCD group. Mazdutide administration reversed these alterations, significantly lowering MDA levels and enhancing SOD activity compared to the HFD group (Figure 4G,H).
2.5. Mazdutide Suppresses ER Stress via the PERK Pathway in MASLD Mouse Liver Tissue
To assess ER stress, we analyzed key markers by immunohistochemistry and Western blot. Immunohistochemistry revealed strong positive staining for GRP78 and CHOP in the HFD group, which was visibly attenuated in all Mazdutide-treated groups compared to the HFD group (Figure 5A). Western blot analysis confirmed these findings. Protein levels of the ER chaperone GRP78 were significantly elevated in the HFD group compared to the NCD group. Furthermore, key components of the PERK-eIF2α-ATF4-CHOP branch were markedly activated in the HFD group, as evidenced by increased phosphorylation levels of PERK (p-PERK/PERK ratio) and eIF2α (p-eIF2α/eIF2α ratio), along with upregulated protein expression of ATF4 and its downstream target CHOP compared with the NCD group. Mazdutide treatment reversed these HFD-induced increases, significantly reducing the phosphorylation of PERK and eIF2α and downregulating the protein levels of GRP78, ATF4, and CHOP compared to the HFD group (Figure 5B,C).
2.6. Mazdutide Downregulates Hepatic Inflammation and Lipogenesis in a Mouse Model of MASLD
Hepatic expression of key inflammatory and lipogenic proteins was assessed by Western blot (Figure 6). Compared to the NCD group, the HFD group showed a significant increase in the ratio of phosphorylated nuclear factor kappa-B p65 (p-NF-κB p65) to total NF-κB p65 (p-NF-κB p65/NF-κB p65) and the protein level of TNF-α, indicating activation of the NF-κB pathway. Concurrently, the expression of lipogenic transcription factors—sterol regulatory element-binding protein 1 (SREBP-1), CCAAT/enhancer-binding protein beta (C/EBPβ), and peroxisome proliferator-activated receptor gamma (PPARγ)—was also significantly elevated in the HFD group. Treatment with Mazdutide significantly reversed these HFD-induced changes. Compared to the HFD group, the p-NF-κB p65/NF-κB p65 ratio and the expression levels of TNF-α, SREBP-1, C/EBPβ, and PPARγ were all significantly downregulated in the liver.
2.7. Mazdutide Reduces Lipid Accumulation in an MASLD Cell Model and Informs Concentration Selection
An in vitro MASLD model was established in Alpha Mouse Liver-12 (AML-12) cells using 1 mM FFA (FFA group), and the control group (Cont) was treated with medium containing the same concentration of bovine serum albumin (BSA) solvent as the FFA group. Cells were co-treated with 1 mM FFA and varying concentrations of Mazdutide (5 nM–1 μM). Compared to the Cont group, intracellular TG levels were significantly increased by FFA, but were markedly reduced by Mazdutide at concentrations ranging from 5 nM to 200 nM (Figure 7A), with the most pronounced reductions observed at 10, 20, and 50 nM. Similarly, Oil Red O staining demonstrated that lipid droplet accumulation induced by FFA was visibly diminished by Mazdutide at 5 nM to 200 nM compared to the FFA group (Figure 7B,C), most notably at 10, 20, and 50 nM. The 50 nM treatment restored cell morphology most closely to that of the Cont group. Based on these results, 10 nM (low), 20 nM (medium), and 50 nM (high) Mazdutide were selected for subsequent experiments.
2.8. Mazdutide Mitigates Inflammation and Oxidative Stress in MASLD Cells
Inflammatory and oxidative stress markers were quantified in the MASLD cell model. Compared to the Cont group, the FFA group exhibited significantly elevated levels of TNF-α, IL-6, and IL-1β, alongside decreased SOD activity and increased MDA content. These FFA-induced alterations were significantly reversed by co-treatment with low-dose Mazdutide (Maz-L), medium-dose Mazdutide (Maz-M), high-dose Mazdutide (Maz-H), or ER stress inhibitor 4-phenylbutyric acid (4-PBA) (Figure 8).
2.9. Mazdutide Inhibits the PERK-eIF2α-ATF4-CHOP Pathway in MASLD Cells
Western blot analysis of key ER stress markers demonstrated that FFA challenge robustly activated the PERK-eIF2α-ATF4-CHOP pathway in hepatocytes. This activation was evidenced by a significant upregulation of GRP78, ATF4, and CHOP, alongside increased phosphorylation of PERK and eIF2α, relative to the Cont group. Notably, co-treatment with Mazdutide (at low, medium, or high doses) or the chemical chaperone 4-PBA effectively mitigated this stress response, with all intervention groups exhibiting markedly attenuated expression of these proteins compared to the FFA model group (Figure 9).
2.10. Mazdutide Reduces the Expression of Inflammation and Lipid Metabolism-Related Proteins in MASLD Cells
Western blot analysis revealed that FFA treatment simultaneously activated pro-inflammatory and lipogenic pathways in hepatocytes. Specifically, it significantly enhanced NF-κB pathway activity, evidenced by an increased p-NF-κB p65/NF-κB p65 ratio, and upregulated the expression of the downstream cytokine TNF-α and the key lipogenic regulators SREBP-1, C/EBPβ, and PPARγ, compared to the Cont group. Importantly, co-administration of Mazdutide (at low, medium, or high doses) or the ER stress inhibitor 4-PBA effectively countered these changes, significantly attenuating NF-κB activation and reducing the expression of TNF-α, SREBP-1, C/EBPβ, and PPARγ compared to the FFA model group (Figure 10).
3. Discussion
MASLD, formerly known as NAFLD, represents the most prevalent chronic liver disorder globally, posing a significant public health burden. The current lack of targeted pharmacotherapies underscores an urgent need for effective and safe intervention strategies. Mazdutide, a dual GLP-1 and GCG receptor agonist, has shown efficacy in weight reduction and glycemic control. However, its therapeutic potential and precise molecular mechanisms in MASLD remain insufficiently defined [21]. This study utilized an HFD-induced murine MASLD model to replicate common unhealthy dietary patterns. Concurrently, an in vitro MASLD model was established by treating AML-12 cells with FFA at a final concentration of 1 mM, composed of oleic acid (OA) and palmitic acid (PA) in a 2:1 molar ratio, mimicking the imbalanced fatty acid milieu observed in patients. We systematically evaluated the therapeutic effects of Mazdutide on MASLD and explored its underlying mechanisms, thereby providing preclinical evidence to support its clinical translation.
Previous studies [22,23] have demonstrated that the progression of MASLD typically involves hepatic lipid accumulation, hepatocyte injury, and impaired liver function, often accompanied by chronic inflammation and oxidative stress—pathological features all reproduced in our model. Our findings indicate that Mazdutide exerts protective effects against MASLD at both the animal and cellular levels. Mice fed a high-fat diet developed severe dyslipidemia and hepatic dysfunction, which were significantly ameliorated following Mazdutide treatment. Histopathological analysis further revealed that Mazdutide markedly reduced hepatic steatosis and lipid droplet accumulation. Consistent results were observed in both in vivo and in vitro experiments: Mazdutide decreased oxidative stress levels and reduced proinflammatory cytokine secretion in the MASLD model. Collectively, these findings align with previous reports on similar agents (e.g., semaglutide), indicating that GLP-1-based therapies can mitigate MASLD progression [24,25,26]. By demonstrating Mazdutide’s protective effects against MASLD, this study expands existing knowledge and further validates the therapeutic potential of dual GLP-1/GCG agonists for MASLD.
Notably, during in vitro studies investigating Mazdutide’s lipid-lowering effects and determining its optimal concentration range, we observed that Mazdutide significantly reduced TG levels at lower doses (10 nM–50 nM). While at higher doses (≥100 nM), TG levels increased. This suggests Mazdutide may exhibit a “Low concentrations reduce lipid accumulation, while high concentrations diminish the effect” profile at the cellular level. Considering Mazdutide’s pharmacological properties as a dual GLP-1/GCG receptor agonist and established literature patterns, we hypothesize that its potential mechanism may involve dual receptor signaling imbalance: at concentrations exceeding 100 nM, the GLP-1 receptor may undergo phosphorylation-dependent endocytosis due to excessive stimulation, reducing the number of effective receptors on the cell membrane surface and consequently weakening the signal inhibiting lipogenesis [27]. In contrast, β-arrestin recruitment at the GCG receptor is phosphorylation-independent, as previously reported [28]; this regulatory feature may confer greater signaling persistence at high concentrations compared to the GLP-1 receptor. This disparity may cause receptor functional imbalance: the GCG receptor continuously drives lipolysis, producing large amounts of free fatty acids, while the GLP-1 receptor-mediated inhibition of lipogenesis is significantly weakened. Fatty acid oxidation capacity reaches saturation, and free fatty acids not promptly oxidized are passively re-esterified into TG and accumulate [29,30]. This ultimately manifests as diminished lipid-lowering effects at high concentrations.
Extensive evidence indicates that ER stress plays a pivotal role in the pathogenesis and progression of MASLD [31,32]. Specifically, excessive lipid accumulation disrupts ER homeostasis, triggering abnormal activation of the UPR. Under stress conditions, expression of the stress sensor GRP78 is upregulated, subsequently activating downstream pathways such as PERK-eIF2α-ATF4-CHOP [8]. Concurrently, chronic ER stress drives lipid synthesis and inflammatory responses, thereby accelerating MASLD progression. This creates a vicious cycle of mutual causation among ER stress, abnormal lipid accumulation, and inflammatory responses within hepatocytes. As previously demonstrated, the lipogenic transcription factor SREBP-1 promotes hepatic lipid accumulation by regulating its downstream lipogenic enzymes [33,34], while ER stress modulates SREBP-1 maturation via the PERK pathway [35,36]. C/EBPβ participates in hepatic metabolic disorders by regulating the expression of lipogenic and inflammation-related genes [37,38]. Its transcriptional activity increases under ER stress, potentially exacerbating lipid abnormalities [39]. PPARγ, a key regulator of lipid storage, is closely associated with hepatic steatosis [40,41]. Under conditions of nutritional excess, PERK activation promotes nuclear translocation of PPARγ [42,43]. Furthermore, PERK-mediated ER stress activates the NF-κB signaling pathway [44], and sustained NF-κB activation may form a vicious cycle exacerbating metabolic dysfunction and ER stress [45,46]. This study found that Mazdutide significantly downregulated GRP78 and key proteins in the PERK-eIF2α-ATF4-CHOP pathway both in vivo and in vitro, thereby alleviating hepatic ER stress. Simultaneously, it downregulated expression of lipid synthesis transcription factors SREBP-1, C/EBPβ, and PPARγ, reduced NF-κB p65 phosphorylation and TNF-α protein expression, thereby mitigating de novo lipogenesis and hepatic inflammation. These multidimensional synergistic regulatory effects resemble those of the classic ER stress inhibitor 4-PBA, supporting a close association between inhibition of the PERK-eIF2α-ATF4-CHOP pathway and improved lipid metabolism alongside reduced inflammation. This suggests that Mazdutide exerts protective effects by alleviating endoplasmic reticulum stress and mitigating associated lipogenesis and inflammatory responses.
In summary, this study confirms Mazdutide’s significant hepatocellular protective effects against MASLD. At the molecular level, Mazdutide regulates the expression of lipogenic and inflammatory molecules and the PERK-eIF2α-ATF4-CHOP signaling pathway, breaking the vicious cycle linking ER stress, abnormal lipid accumulation, and inflammation. This demonstrates its multi-target synergistic protective characteristics. Given the current lack of clinical therapeutic options for MASLD, this study provides preclinical evidence supporting the clinical translation of Mazdutide for this disease and offers novel research perspectives for multi-targeted interventions in metabolic liver diseases.
4. Materials and Methods
4.1. Reagents
Mazdutide acetate was purchased from Abmole Bioscience Inc. (Houston, TX, USA). TG, TC, LDL-C, HDL-C, ALT, AST, MDA, and SOD assay kits were purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing, China; Catalog Nos.: A110-1-1, A111-1-1, A113-1-1, A112-1-1, C009-2-1, C010-2-1, A003-1, A001-3, respectively). The FFA assay kit was purchased from Abbkine Scientific Co., Ltd. (Wuhan, China; Catalog No.: KTB2230). Mouse TNF-α, mouse IL-1β, and mouse IL-6 ELISA kits were purchased from Jonln Bio (Shanghai, China; Catalog Nos.: JLW10484, JLW18442, JLW20268, respectively). Oleic acid (OA), fatty acid- free and IgG-free bovine serum albumin (BSA), and BCA protein concentration assay kits were purchased from Beyotime Biotechnology (Shanghai, China). Palmitic acid (PA) was purchased from Sigma-Aldrich (St. Louis, MO, USA). 4-PBA was purchased from MedChemExpress (Monmouth Junction, NJ, USA). Primary antibodies for GRP78, ATF4, CHOP, and β-actin were purchased from Proteintech Group, Inc (Wuhan, China). Antibodies for SREBP-1, C/EBPβ, and PPARγ were purchased from ZEN-BIOSCIENCE Co., Ltd. (Chengdu, China). Antibodies for eIF2α and p-eIF2α (Ser51) were purchased from Cell Signaling Technology (Danvers, MA, USA). Antibodies for PERK, p-PERK (T892), NF-κBp65, p-NF-κBp65 (Ser536), and TNF-α were purchased from Wanlei Life Sciences (Shenyang) Co., Ltd. (Shenyang, China). Secondary antibodies were purchased from Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd. (Beijing, China).
4.2. Animals and Treatment
Six-week-old male C57BL/6J mice [47] were purchased from Henan Skobes Biotechnology Co., Ltd. (Anyang, China) and housed in a constant temperature (22 ± 2 °C) and humidity (40–60%) environment under a 12 h light/dark cycle. Mice had free access to chow and drinking water. Mice were randomly divided into five groups (n = 15 per group). Following a 1-week acclimation, one normal chow diet (NCD) group and four high-fat diet (HFD, 60% kcal from fat) groups were established. The normal chow used in this study was commercially available maintenance chow for mice, purchased from Huanyu Biotechnology Co., Ltd. (Beijing, China). The high-fat diet was custom-produced by Henan Skobes Biotechnology Co., Ltd. (Anyang, China) based on the publicly available D12492 formulation from Research Diets, Inc. (New Brunswick, NJ, USA). For details, the composition of NCD and HFD is shown in Table S1 in the Supplementary Materials.
Prior to the formal drug administration treatment, a single high dose of Mazdutide (600 μg/kg) was subcutaneously injected into 5 naive healthy SPF male C57BL/6J mice (not included in the formal experimental groups). The general condition, body weight and obvious abnormal reactions were monitored daily for 14 days, with no toxic effects observed in all mice. Based on this safety verification and referring to the effective dose range of Mazdutide in metabolic models [16], 100, 200 and 400 μg/kg were set as the gradient doses for the subsequent formal study. After 12 weeks of HFD feeding, Mazdutide was administered subcutaneously every 3 days for 4 weeks to three HFD-fed groups: the high-fat-diet low-dose group (HFD+L, 100 μg/kg), the high-fat-diet medium-dose group (HFD+M, 200 μg/kg), and the high-fat-diet high-dose group (HFD+H, 400 μg/kg). Mazdutide was dissolved in 0.9% sterile normal saline for subcutaneous injection at respective doses; equal volumes of the same saline were subcutaneously administered to NCD and HFD control mice. At the end of the treatment, all mice were euthanized for the collection of blood and liver samples. A schematic diagram of the animal experiment design was drawn (Figure 11).
4.3. Preparation of FFA
PA and OA were individually dissolved in sodium hydroxide (NAOH) solution at a molar ratio of 1:1 (fatty acid:NaOH). Solubilization was facilitated by incubation in a 70 °C water bath, followed by dilution of each fatty acid into 10% fatty-acid-free and IgG-free BSA solution. The resultant PA and OA stock solutions were sterile-filtered through 0.22 μm membranes, aliquoted, and stored at −80 °C in the dark. For experimental use, the working FFA solution was freshly prepared by mixing PA and OA stocks at a molar ratio of 1:2 [48].
4.4. Cell Culture and Grouping
AML-12 cell line, an immortalized and non-transformed hepatocyte line, was purchased from Zhongqiaoxinzhou Biotechnology Co., Ltd. (Shanghai, China). AML-12 cells were cultured in DMEM/Ham’s F12 medium supplemented with 10% fetal bovine serum, insulin-transferrin-selenium mixture, 0.1 mM dexamethasone, penicillin, and streptomycin. Cells were cultured at 37 °C in a humidified environment with 5% CO_2_. TG content was measured after co-treating AML-12 cells with varying Mazdutide concentrations (5 nM–1 μM) and 1 mM FFA for 24 h. Lipid droplet formation was visualized via Oil Red O staining. The optimal Mazdutide concentrations for improving lipid accumulation—10 nM, 20 nM, and 50 nM—were selected as low, medium, and high doses, respectively. AML-12 cells were divided into six experimental groups: the Cont group, treated with basal medium supplemented with an equal concentration of BSA as in FFA-treated groups but without FFA; the FFA group, treated with basal medium containing 1 mM free fatty acids to establish a high-fat cell model; three Mazdutide intervention groups co-treated with 1 mM FFA and Mazdutide at low (10 nM), medium (20 nM), or high (50 nM) doses (Maz-L, Maz-M, and Maz-H, respectively); and the 4-PBA group, co-treated with 1 mM FFA and 1 mM 4-PBA. Mazdutide and 4-PBA were first dissolved in basal medium to prepare stock solutions.
4.5. Biochemical Parameter Detection
Serum levels of TG, TC, LDL-C, HDL-C, ALT, and AST were measured using commercially available biochemical assay kits according to the manufacturers’ instructions, with TG quantified via the glycerol-3-phosphate oxidase-p-aminophenol (GPO-PAP) enzymatic method (Limit of Detection, LOD: 0.035 mmol/L; Limit of Quantification, LOQ: 0.3 mmol/L), TC via the cholesterol oxidase-p-aminophenol (COD-PAP) method (LOD: 0.28 mmol/L; LOQ: 0.42 mmol/L), LDL-C and HDL-C via a direct enzymatic approach with surfactant-mediated clearance (LOD: 0.075 mmol/L and 0.072 mmol/L; LOQ: 0.61 mmol/L and 0.09 mmol/L, respectively), and ALT and AST via colorimetric detection using 2,4-dinitrophenylhydrazine (LOD: 0.35 U/L and 0.3 U/L; LOQ: 0.62 U/L and 0.74 U/L, respectively). The levels of TG, TC, FFA, MDA, and SOD in 10% (w/v) mouse liver tissue homogenate were detected according to the kit manufacturer’s instructions, where FFA was assayed by a copper-based microplate colorimetric method (LOD: 0.0156 mM; LOQ: 0.0313 mM), MDA via the thiobarbituric acid reactive substances assay (TBARS) (LOD: 0.36 nmol/mL; LOQ: 0.5 nmol/mL), and SOD activity via the water-soluble tetrazolium salt-1 (WST-1) reduction method (LOD: 2.89 U/mL; LOQ: 5 U/mL). Similarly, the levels of TG, MDA, and SOD in AML-12 cell lysate were detected according to the kit manufacturer’s instructions, utilizing the same reagents and analytical workflows as applied to liver homogenates to maintain assay uniformity.
4.6. Inflammatory Cytokine Assay
Inflammatory markers in mouse serum and AML-12 cell culture supernatants, including TNF-α (LOD: 0.59 pg/mL; LOQ: 1.25 pg/mL), IL-1β (LOD: 0.32 pg/mL; LOQ: 0.78 pg/mL), and IL-6 (LOD: 0.74 pg/mL; LOQ: 1.56 pg/mL), were detected using sandwich ELISA kits according to the manufacturers’ instructions. The concentrations of these three inflammatory cytokines in 10% (w/v) mouse liver tissue homogenates were also quantified via the same protocol.
4.7. HE Staining
Liver tissues from corresponding sites in each group of mice were fixed in 4% paraformaldehyde for 24 h, then routinely processed and embedded in paraffin. The paraffin-embedded tissue blocks were sectioned at 5 μm thickness using a rotary microtome. For HE staining, the sections were dewaxed in xylene and rehydrated through a graded ethanol series. Subsequently, the sections were stained with hematoxylin for 10 min, differentiated in 1% acid ethanol for 30 s, and blued in running water for 10 min. This was followed by eosin staining for 5 min. Finally, the stained sections were dehydrated through a graded ethanol series, cleared in xylene, mounted with a neutral resin, and examined under an upright transmitted light microscope (Olympus BX41, Olympus Corporation, Tokyo, Japan) for morphological assessment.
4.8. Oil Red O Staining
4.8.1. Tissue Staining
Mouse liver tissues from corresponding sites were embedded in O.C.T. compound, cryopreserved at −80 °C, and sectioned at 10 μm using a cryostat. After fixation with 4% paraformaldehyde and washing, sections were stained with freshly filtered Oil Red O working solution (3:2 stock:water) for 20 min in the dark. Sections were then differentiated in 60% isopropanol, rinsed, and counterstained with hematoxylin. Following bluing in water, sections were mounted with glycerol-gelatin medium and examined by the Olympus BX41 microscope. Lipid droplets within hepatocytes were identified as orange-red deposits.
4.8.2. Cell Staining
AML-12 cells were seeded in 6-well plates. Upon reaching 60–70% confluence, the cells were co-treated with an FFA solution and various concentrations of Mazdutide for 24 h. After treatment, the cells were washed three times with PBS, fixed with 4% paraformaldehyde for 10 min, and then stained using the same Oil Red O staining protocol as described for tissue sections (Section 4.8.1). Finally, the stained cells were washed, and images were acquired using an inverted microscope (Zeiss Axio Observer 3, Carl Zeiss AG, Oberkochen, Germany) in bright-field mode.
4.9. Immunohistochemical Staining
Paraffin-embedded mouse liver sections were dewaxed, rehydrated, and subjected to antigen retrieval in citrate buffer (pH 6.0). After blocking endogenous peroxidase with 3% H_2_O_2_ and nonspecific sites with 5% BSA, sections were incubated overnight at 4 °C with primary antibodies against GRP78 or CHOP (1:500). Following incubation with an HRP-conjugated goat anti-rabbit secondary antibody (1:200), immunoreactivity was visualized using a DAB kit. Nuclei were counterstained with hematoxylin. Sections were then dehydrated, cleared, mounted, and examined under an upright light microscope (Nikon ECLIPSE E100, Nikon Corporation, Tokyo, Japan). Positive expression was identified by yellow-brown cytoplasmic or nuclear granules.
4.10. Western Blotting
Proteins were extracted from liver tissues and hepatocytes using RIPA buffer with phosphatase and protease inhibitors. Concentrations were determined by BCA assay. Equal protein amounts were denatured, separated by SDS-PAGE, and transferred to PVDF membranes. After blocking with 5% non-fat milk in TBST, membranes were incubated overnight at 4 °C with primary antibodies, followed by HRP-conjugated secondary antibodies for 1 h at room temperature. Following TBST washes, bands were visualized using an enhanced chemiluminescence (ECL) detection system.
4.11. Statistical Analysis
Statistical analyses were performed using GraphPad Prism 9.0. Data from ≥3 independent experiments are expressed as mean ± SD. Multiple group comparisons used one-way ANOVA followed by Tukey’s test. p < 0.05 was statistically significant. Western blot band intensities were quantified via ImageJ 1.54p densitometry and normalized to β-actin.
5. Conclusions
In summary, this study demonstrates that Mazdutide significantly improves multiple abnormal indicators in high-fat-diet-induced MASLD mice, such as alleviating dyslipidemia, improving liver function, reducing hepatic steatosis, and simultaneously lowering oxidative stress levels and the release of inflammation-related factors. Its hepatocyte-protective effects were further validated in an FFA-induced MASLD cellular model. The mechanism may involve Mazdutide inhibiting endoplasmic reticulum stress (via the PERK-eIF2α-ATF4-CHOP axis) and its associated inflammatory pathways (NF-κB/TNF-α), as well as regulating lipid metabolism-related molecules (SREBP-1, C/EBPβ, PPARγ), thereby improving and delaying MASLD progression. This finding provides new experimental evidence for the molecular mechanism of Mazdutide in improving MASLD and its clinical application in related disease treatment.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Drummer C.IV Saaoud F. Jhala N.C. Cueto R. Sun Y. Xu K. Shao Y. Lu Y. Shen H. Yang L. Caspase-11 promotes high-fat diet-induced NAFLD by increasing glycolysis, OXPHOS, and pyroptosis in macrophages Front. Immunol.202314111388310.3389/fimmu.2023.111388336776889 PMC 9909353 · doi ↗ · pubmed ↗
- 2Turkseven S. Turato C. Villano G. Ruvoletto M. Guido M. Bolognesi M. Pontisso P. Di Pascoli M. Low-Dose Acetylsalicylic Acid and Mitochondria-Targeted Antioxidant Mitoquinone Attenuate Non-Alcoholic Steatohepatitis in Mice Antioxidants 20231297110.3390/antiox 1204097137107346 PMC 10135482 · doi ↗ · pubmed ↗
- 3Rinella M.E. Lazarus J.V. Ratziu V. Francque S.M. Sanyal A.J. Kanwal F. Romero D. Abdelmalek M.F. Anstee Q.M. Arab J.P. A multisociety Delphi consensus statement on new fatty liver disease nomenclature Hepatology 2023781966198610.1097/HEP.000000000000052037363821 PMC 10653297 · doi ↗ · pubmed ↗
- 4Bae J. Han E. Lee H.W. Park C.Y. Chung C.H. Lee D.H. Cho E.H. Rhee E.J. Yu J.H. Park J.H. Metabolic Dysfunction-Associated Steatotic Liver Disease in Type 2 Diabetes Mellitus: A Review and Position Statement of the Fatty Liver Research Group of the Korean Diabetes Association Diabetes Metab. J.2024481015102810.4093/dmj.2024.054139610131 PMC 11621661 · doi ↗ · pubmed ↗
- 5Li S. Mehal W.Z. Ouyang X. RNA modifications in the progression of liver diseases: From fatty liver to cancer Sci. China Life Sci.2024672105211910.1007/s 11427-023-2494-x 38809498 PMC 11545962 · doi ↗ · pubmed ↗
- 6Peiseler M. Schwabe R. Hampe J. Kubes P. Heikenwälder M. Tacke F. Immune mechanisms linking metabolic injury to inflammation and fibrosis in fatty liver disease—Novel insights into cellular communication circuits J. Hepatol.2022771136116010.1016/j.jhep.2022.06.01235750137 · doi ↗ · pubmed ↗
- 7Manikat R. Nguyen M.H. Nonalcoholic fatty liver disease and non-liver comorbidities Clin. Mol. Hepatol.202329 s 86s 10210.3350/cmh.2022.044236603574 PMC 10029963 · doi ↗ · pubmed ↗
- 8Karin M. Kim J.Y. Endoplasmic reticulum stress at the forefront of fatty liver diseases and cancer Pharmacol. Rev.20257710009610.1016/j.pharmr.2025.10009641197470 · doi ↗ · pubmed ↗