Something Old Brings New Insights: Activity of Cerivastatin Against Thermally Dimorphic Fungi and Its Potential as an Antifungal Scaffold
Luana P. Borba-Santos, Sonia Rozental

TL;DR
Cerivastatin, a known drug, shows strong antifungal activity against certain fungi and works better when combined with itraconazole.
Contribution
Cerivastatin's antifungal potential and its scaffold as a basis for new antifungal drugs are newly identified.
Findings
Cerivastatin inhibited Sporothrix species growth at low concentrations.
Combining cerivastatin with itraconazole enhanced antifungal effects.
Cellular changes were more pronounced with combination treatment.
Abstract
Background/Objectives: Species of the Sporothrix genus were used as a biological model to identify potential antifungal compounds within the NIH Clinical Collection library. Methods: A total of 707 compounds were screened for antifungal activity using in vitro susceptibility assays. Compounds exhibiting significant inhibitory effects (growth inhibitions greater than 80%) were further evaluated by determining their minimum inhibitory concentrations (MICs). As cerivastatin demonstrated the highest activity after itraconazole, it was selected for further evaluation, either alone or in combination with itraconazole, using susceptibility assays, electron microscopy, and flow cytometry analyses. Results: Among the screened compounds, twenty-six showed significant inhibition of yeast growth (≥80%). Compounds with previously reported antifungal activity or not used as oral treatment were…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
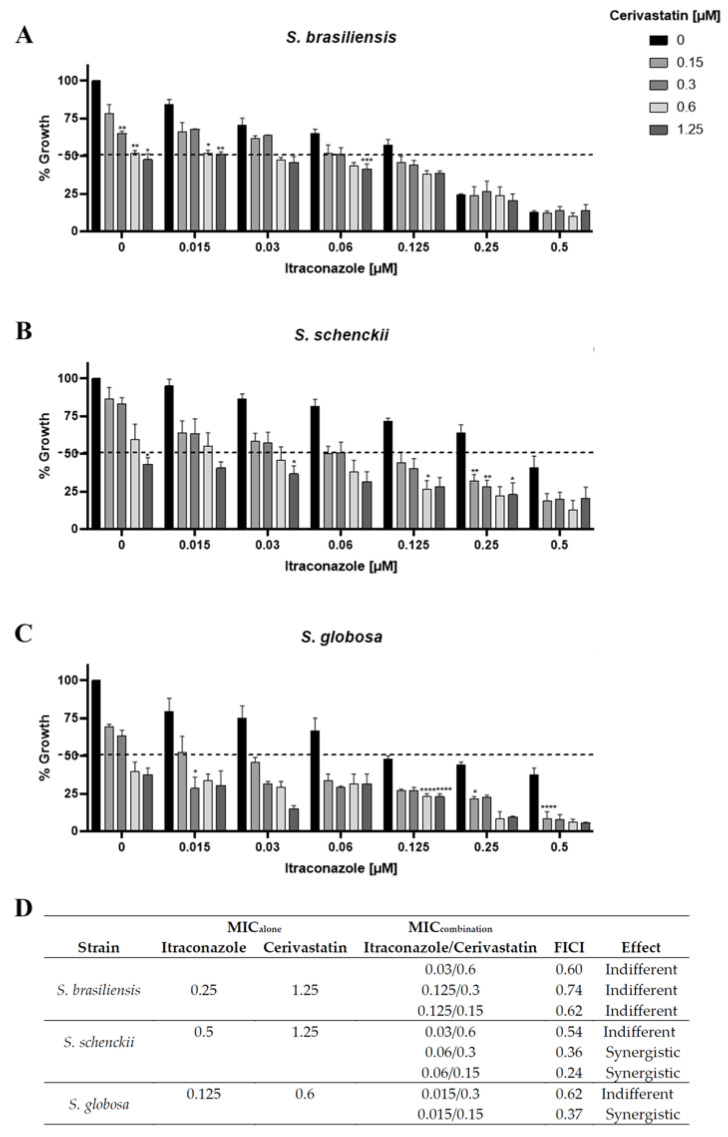 Figure 1
Figure 1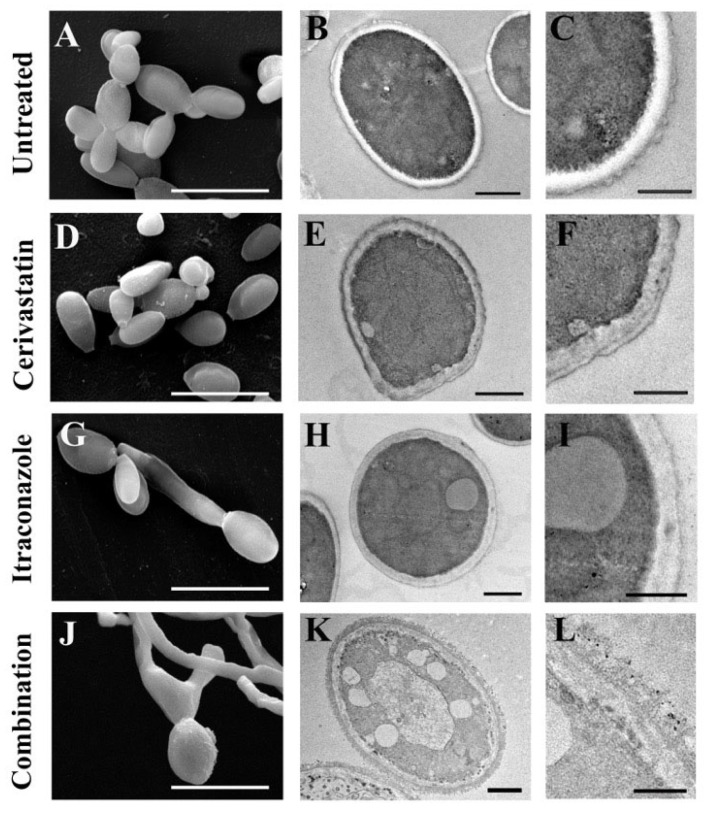 Figure 2
Figure 2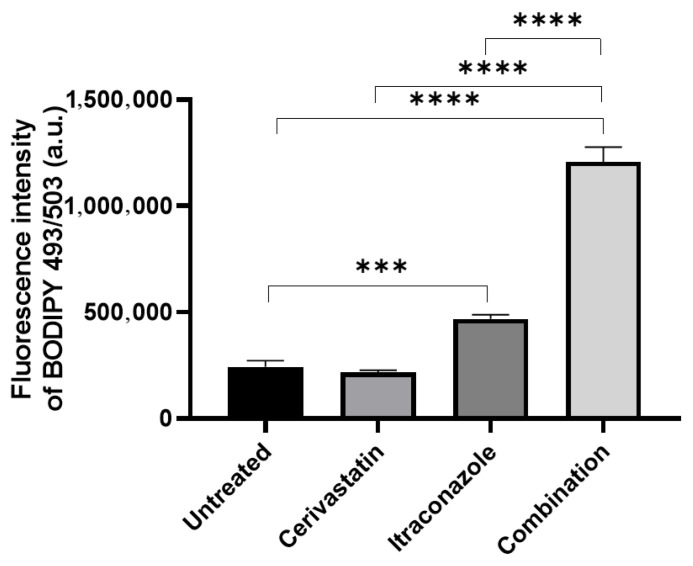 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9 Figure 10
Figure 10- —the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
- —Fundacão Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Natural Products and Biosynthesis · Tannin, Tannase and Anticancer Activities · Sirtuins and Resveratrol in Medicine
1. Introduction
Fungal infections are one of the leading causes of infectious disease-related deaths worldwide [1,2]. The main antifungal classes used to treat these infections are limited by suboptimal efficacy, toxicity, drug interactions, and poor activity against resistant species [3]. Therefore, the study of more effective compounds that act more selectively on fungal cells and require shorter treatment times is essential.
The evaluation of libraries containing hundreds of molecules is a useful tool for discovering promising chemical groups with antifungal activity [4]. This approach enables the study of a wide range of molecules with diverse physicochemical characteristics, which can serve as a good scaffold for the development of new antifungals.
In the present work, we used pathogenic species of the Sporothrix genus as a biological model for medically relevant fungi: Sporothrix brasiliensis, Sporothrix schenckii, and Sporothrix globosa. These species cause sporotrichosis, a neglected tropical disease that is the most common subcutaneous mycosis worldwide [5,6]. The main sporotricosis manifestation is lymphocutaneous disease, followed by cutaneous form; however, sporotrichosis can assume ocular, pulmonary, osteoarticular, meningitis, and other manifestations [5].
Sporothrix spp. are thermodimorphic fungi, present in filamentous form in nature (28 °C) and or in yeast form when infected the mammalian host (37 °C). The most virulent species is S. brasiliensis [5], which is also the most frequent species observed in Brazil. Sporotrichosis is hyperendemic in various regions of the country, with zoonotic transmission by infected cats being the primary mode of disease acquisition [5].
Reports of therapeutic failures and the identification of isolates with reduced sensitivity to itraconazole (the reference antifungal used to treat sporotrichosis) have become more frequent in recent years [5]. Here, we evaluated the NIH Clinical Collection library against Sporothrix yeasts to discover promising chemical groups with antifungal activity, and studied the effects of the most promising compound combined with itraconazole. We showed that cerivastatin inhibited Sporothrix growth and enhanced itraconazole activity.
2. Results
2.1. NIH Clinical Collection Screening Reveals Antifungal Activity of Cerivastatin
Initially, the in vitro susceptibility of S. brasiliensis yeasts cells to compounds from the NIH Clinical Collection library was evaluated at a concentration of 10 µM. Among the 707 molecules tested, 26 inhibited more than 80% of yeast growth (Table 1 and Table S1). From these group, fifteen compounds with previously reported antifungal activity as well as compounds with previously antifungal descriptions and not used as oral treatment were excluded from further analysis [5,6,7,8].
Minimum inhibitory concentrations (MICs) were determined using the broth microdilution method for the remaining eleven compounds against yeast forms of Sporothrix brasiliensis, Sporothrix schenckii, and Sporothrix globosa (Table 2). Itraconazole was included as the reference antifungal agent. Among these eleven compounds, cerivastatin exhibited the most potent antifungal activity after itraconazole, inhibiting the growth of all three Sporothrix species at concentrations as low as 1.25 µM (Table 2).
In addition to cerivastatin, other statins present in the NIH Clinical Collection library (fluvastatin, pitavastatin, lovastatin, mevastatin, pravastatin, and simvastatin) were also evaluated in the growth inhibition assay. In contrast to cerivastatin, which induced marked growth inhibition, none of the other statins exhibited significant inhibitory activity against Sporothrix brasiliensis at a concentration of 10 µM (Table 3).
2.2. Cerivastatin Enhances the Anti-Sporothrix Activity of Itraconazole
Cerivastatin, like other statins, inhibits the enzyme HMG-CoA reductase (3-hydroxy-3-methylglutaryl-coenzyme A reductase), a key component of the sterol biosynthesis pathway [11]. Itraconazole also targets sterol biosynthesis but acts at a distinct step by inhibiting C14-α-lanosterol demethylase. To evaluate the potential synergistic interaction between cerivastatin and itraconazole, a checkerboard assay was performed. The results indicated that the combination of the two compounds produced a greater inhibitory of Sporothrix growth than either agent alone (Figure 1A–C). Analysis of the fractional inhibitory concentration index (FICI) revealed synergistic interactions for S. schenckii and S. globosa (FICI ≤ 0.5) when the drugs were combined (Figure 1D). In S. brasiliensis, lower concentrations of the combined drugs were effective to inhibit fungal growth, reducing MIC values by more than twofold; however, no synergistic effect was observed according to the FICI criteria (Figure 1D).
To compare the cellular effects of cerivastatin in combination with itraconazole, S. brasiliensis cells were treated with 0.125 µM itraconazole and 0.15 µM cerivastatin for 48 h and subsequently analyzed by scanning electron microscopy (SEM), transmission electron microscopy (TEM), and flow cytometry. SEM analysis showed that untreated cells displayed an elongated morphology, which was also observed in cells treated with cerivastatin alone (Figure 2A,C). In contrast, yeast-to-hyphae conversion was observed following exposure to itraconazole and to the combination treatment (Figure 2E,G). TEM analysis of untreated cells revealed an electron-dense cytoplasm surrounded by an intact cell wall and plasma membrane (Figure 2B). Treatment with either cerivastatin or itraconazole led to the formation of electron-lucent vacuoles within the cytoplasm, with cerivastatin additionally inducing marked alterations in the cell wall and plasma membrane (Figure 2D,F). Cells exposed to the combination therapy exhibited extensive cytoplasmic disorganization, pronounced accumulation of electron-lucent vacuoles, fragmentation of the plasma membrane, and severe structural disruption of the cell wall (Figure 2H).
We investigated whether treatment with the combination of cerivastatin and itraconazole increased neutral lipid accumulation using the fluorescent probe BODIPY 496/503 to stain fungal cells (Figure 3). Cerivastatin alone did not induce neutral lipid accumulation, whereas itraconazole treatment resulted in a statistically significant increase in neutral lipid levels. Notably, this effect was further amplified when the two drugs were used in combination (Figure 3). Taken together, these findings indicate that the combined treatment with cerivastatin and itraconazole induces more pronounced morphophysiological alterations in fungal cells than either drug alone.
3. Discussion
The limitations of current therapies for fungal infections highlight the need to explore alternative molecular scaffolds and cellular targets. In this study, we screened 707 compounds from the NIH Clinical Collection library, which contains molecules with a documented history of use in human clinical trials, to identify those with significant antifungal activity against pathogenic species of the genus Sporothrix. Our results identify cerivastatin as the most promising candidate in this group.
Cerivastatin stood out for its ability to inhibit the growth of three clinically relevant Sporothrix species (S. brasiliensis, S. schenckii, and S. globosa) at low concentrations (<1.25 µM). Moreover, cerivastatin exhibited synergistic activity when combined with itraconazole, the first-line antifungal therapy for sporotrichosis. Cerivastatin is a statin originally developed in the 1990s to treat hyperlipidemia and prevent cardiovascular disease, and was withdrawn from the market in 2001 due to an increased incidence of rhabdomyolysis [12]. Curiously, other statins evaluated in this study did not display a significant inhibition of fungal growth (Table 3). The distinctive structural features of cerivastatin may account for its enhanced affinity for fungal HMG-CoA reductase (3-hydroxy-3-methylglutaryl-coenzyme A reductase), resulting in a stronger inhibitory effect on fungal growth compared with other statins. HMG-CoA reductase is the primary target of statins and plays a central role in metabolic pathways responsible for the synthesis of essential cellular components, particularly sterols, cholesterol in humans, and ergosterol in fungal cells [11,13].
Statins are widely prescribed as first-line agents for the control of hypercholesterolemia and prevention of cardiovascular disease [14]. In addition to their lipid-lowering properties, particularly the reduction in LDL cholesterol, these drugs have been increasingly investigated for pleiotropic effects, including anti-inflammatory and immunoregulatory activities [14]. Moreover, accumulating evidence indicates that statins possess diverse biological actions, with reported antimicrobial effects that extend to antiparasitic, antibacterial, antiviral, and antifungal activity [14]. The antifungal potential of statins appears to be multifactorial. One of the primary mechanisms involves the disruption of sterol biosynthesis in fungi through the inhibition of HMG-CoA reductase, a key enzyme in the mevalonate pathway. This interference not only compromises ergosterol production but also alters other processes such as protein prenylation, mitochondrial integrity, and cellular signaling pathways, which may culminate in apoptotic cell death. Additionally, statins have been associated with disturbances in fungal morphogenesis and cell cycle progression [13]. By targeting HMG-CoA reductase, these compounds decrease the synthesis of mevalonate-derived intermediates, including farnesyl pyrophosphate. The resulting impairment of isoprenoid formation limits protein isoprenylation, a post-translational modification essential for the proper localization and function of regulatory proteins [13]. Consequently, processes governing cell proliferation, differentiation, programmed cell death, respiration, and iron homeostasis may be adversely affected in fungal cells [13].
The antifungal activity of cerivastatin has been previously demonstrated against filamentous fungi, with minimum inhibitory concentrations (MICs) ranging from 0.04 to 0.14 µM against Trichophyton species, 0.03 µM against Aspergillus fumigatus, and 1.61 µM and 0.4 µM against the hyphal and conidial forms of Exserohilum rostratum, respectively [15,16,17]. In contrast, the in vitro inhibitory activity of other statins, including lovastatin, simvastatin, pravastatin, and fluvastatin, has been reported against a broad range of pathogenic fungi, such as species of Candida, Cryptococcus, Aspergillus, dermatophytes, and mucormycetes [13,18]. However, antifungal effects are generally observed only at relatively high drug concentrations [13,18], which may explain the limited activity of these statins when tested at 10 µM against S. brasiliensis in the present study.
Cholesterol is the predominant sterol in the plasma membrane of mammalian cells, whereas ergosterol is the principal sterol in the plasma membrane of most pathogenic fungi [11,13]. Inhibition of ergosterol biosynthesis represents one of the main mechanisms exploited by clinically available antifungal agents. The azole class, the most widely used antifungals, targets C14-α-lanosterol demethylase, an enzyme that catalyzes a later step in sterol biosynthesis than HMG-CoA reductase [11,13]. Disruption of ergosterol synthesis compromises membrane integrity, fluidity, and permeability, thereby increasing fungal susceptibility to osmotic, oxidative, and cell wall stresses. Moreover, the accumulation of alternative sterol intermediates can exert toxic effects on fungal cells [11,13,19].
Targeting distinct steps of the sterol biosynthesis pathway through drug combinations represents a promising antifungal strategy. Consistent with previous reports showing that statins can potentiate the activity of azoles against diverse fungal pathogens [13], we investigated whether cerivastatin could exert synergistic effects in vitro when combined with itraconazole against Sporothrix species, and demonstrated that cerivastatin significantly enhanced the antifungal activity of itraconazole against Sporothrix yeasts (Figure 1).
Based on these findings, we selected the lowest concentration of cerivastatin that produced significant growth inhibition in combination with itraconazole against S. brasiliensis yeasts to further assess the morphophysiological changes. S. brasiliensis was chosen due to its major epidemiological relevance in Brazil [5]. Scanning and transmission electron microscopy revealed profound ultrastructural alterations after the combined treatment, indicating that simultaneous inhibition of two distinct steps in the ergosterol biosynthesis pathway markedly increased fungal cell damage (Figure 2).
Inhibition of ergosterol biosynthesis can result in the intracellular accumulation of neutral lipids, which correspond to sterol intermediates generated during incomplete ergosterol synthesis [11]. Flow cytometric analysis revealed significantly higher levels of neutral lipid accumulation following treatment with the cerivastatin–itraconazole combination compared with either cerivastatin or itraconazole alone (Figure 3). These findings support the notion that the enhanced antifungal effect of the combined treatment is primarily associated with the intensified inhibition of ergosterol biosynthesis.
In recent years, several studies have highlighted the potential of statins as antifungal agents when repurposed in combination with azoles [13]. Nevertheless, additional in vivo studies and clinical evidence are still required to substantiate their therapeutic applicability. From this perspective, accumulating data on the antifungal activity of statins suggest that their chemical scaffolds could be exploited for the development of compounds with increased selectivity toward fungal cells. Notably, comparative sequence analysis indicates limited similarity between fungal and human HMG-CoA reductase (approximately 55% identity), as determined by BLASTp alignment of the human and S. brasiliensis enzymes (XP_011541659.1 and XP_040622855.1, respectively) [20]. This divergence may provide a molecular basis for the rational design of fungal-selective inhibitors.
It is important to emphasize that the other compounds that demonstrated anti-Sporothrix activity and were not explored in this study are potential subjects for future investigations, including studies involving other pathogenic fungi. In the present work, only the compounds capable of markedly inhibiting S. brasiliensis at 10 micromolar, whose antifungal activity had not been previously described and which are used in oral treatments, were selected for the determination of inhibitory concentrations, with cerivastatin being the most promising compound.
In conclusion, our findings highlight the potential of cerivastatin, particularly in combination with itraconazole, as a promising strategy against agents of sporotrichosis. Moreover, the chemical structure of cerivastatin represents a valuable scaffold for the development of novel statins with antifungal properties. We anticipate that the insights generated by this study will contribute to the identification of new therapeutic targets and foster the development of more effective antifungal agents.
4. Materials and Methods
4.1. Fungal Strains and Growth Conditions
Three reference isolates, S. brasiliensis ATCC MYA 4823, S. schenckii ATCC 32286, and S. globosa CBS 130104, were used in this study. To obtain filamentous growth, strains were maintained on Sabouraud agar (Kasvi, São José dos Pinhais, PR, Brazil) at 25 °C for seven days. Thermal dimorphism was subsequently induced by transferring the cultures to brain heart infusion agar (BD Difco, Franklin Lakes, NJ, USA) and incubating them for an additional seven-day period at 35 °C in an atmosphere containing 5% CO_2_. After this phase transition, cells in the yeast (parasitic) form were collected and used throughout all experimental procedures.
4.2. Chemical Library and Reference Drug
A collection of clinically characterized small molecules from the NIH Clinical Collection, provided by the National Center for Advancing Translational Sciences, was screened in this work. Each compound was solubilized in dimethyl sulfoxide (DMSO) to generate 10 mM stock solutions, which were stored at −20 °C until required. Itraconazole (1 mM; Sigma-Aldrich^®^, MO, USA) prepared in DMSO was included as a positive antifungal control.
4.3. Screening of the NIH Clinical Collection Library
To identify molecules with antifungal activity, S. brasiliensis ATCC MYA 4823 was selected as the screening model. Test compounds were prepared at 10 μM in RPMI 1640 medium (Sigma-Aldrich^®^, St. Louis, MO, USA) supplemented with 2% glucose and buffered with 0.165 M MOPS (pH 7.2). Aliquots were dispensed into flat-bottom 96-well microplates (Kasvi, São José dos Pinhais, PR, Brazil). Yeast cells were then inoculated at a final concentration of 1 × 10^5^ CFU/mL per well, and plates were incubated for 48 h at 35 °C in a 5% CO_2_ environment. After incubation, fungal proliferation was qualitatively examined under an inverted microscope and quantitatively determined by optical density measurement at 492 nm. Absorbance readings were corrected by subtracting background values obtained from wells containing only supplemented RPMI. Growth inhibition percentages were calculated relative to the untreated controls according to the equation: I = 100 − (A × 100/C), where A represents treated samples and C corresponds to untreated controls. Wells containing 0.1% DMSO were included to exclude solvent-related effects. Compounds producing more than 80% inhibition were selected as hits, consistent with marked suppression of visible growth. The screening data reflect two independent assays performed with four technical replicates each.
4.4. Determination of Minimum Inhibitory Concentration
The antifungal potency of selected hits was further characterized by determining the minimum inhibitory concentrations using a broth microdilution protocol adapted for Sporothrix spp. [21]. Compounds were serially diluted in supplemented RPMI to yield final concentrations between 0.02 and 10 μM in 96-well plates. Yeast suspensions were added to achieve a final density of 1 × 10^5^ CFU/mL and incubated under the same temperature and CO_2_ conditions described above. Post-incubation assessment included microscopic evaluation and absorbance measurement at 492 nm. The MIC was defined as the lowest drug concentration resulting in at least 50% inhibition of fungal growth, determined using the same inhibition formula described above. Values are reported as the average of three independent experiments conducted in duplicate.
4.5. Evaluation of the Interaction Between Cerivastatin and Itraconazole
Itraconazole–cerivastatin interactions were investigated using a checkerboard microdilution assay as previously described [22]. S. brasiliensis yeast suspensions (1 × 10^5^ CFU/mL) were distributed into microplates containing serial dilutions of cerivastatin (0.15–10 μM) along one axis and itraconazole (0.001–1 μM) along the other. Following incubation for 48 h at 35 °C under 5% CO_2_, fungal growth was assessed and minimum inhibitory concentrations (MICs) were established for each compound alone and in combination. The interaction profile was determined by calculating the fractional inhibitory concentration index (FICI) according to the formula: FICI = (MIC of cerivastatin in combination/MIC of cerivastatin alone) + (MIC of itraconazole in combination/MIC of itraconazole alone) [22]. The drug combination effects were considered synergistic if FICI ≤ 0.5, indifferent if FICI > 0.5 and ≤4, and antagonist if FICI > 4 [23]. The most effective combinations were identified as those yielding the lowest FICI scores. All findings represent results obtained from at least three independent experiments.
4.6. Scanning Electron Microscopy (SEM)
To examine surface ultrastructural alterations, S. brasiliensis yeast cells were exposed for 48 h to 0.125 μM itraconazole and 0.15 μM cerivastatin, either individually or together. After treatment, cells were washed with phosphate-buffered saline (PBS) and chemically fixed for 1 h in a solution containing 2.5% glutaraldehyde and 4% formaldehyde prepared in 0.1 M cacodylate buffer. Samples were subsequently washed, adhered to poly-L-lysine–coated glass coverslips (Sigma-Aldrich^®^), and post-fixed for 30 min in 1% osmium tetroxide supplemented with 1.25% potassium ferrocyanide in cacodylate buffer. A graded ethanol dehydration series (30–100%, 20 min per step) was performed before critical-point drying in CO_2_. Specimens were then sputter-coated with gold and visualized using an FEI Quanta 250 scanning electron microscope (FEI Company, Hillsboro, OR, USA). Digital images were processed with Adobe Photoshop CS6 software.
4.7. Transmission Electron Microscopy (TEM)
For internal ultrastructural analysis, yeast cells were treated under the same drug conditions described above. After washing with PBS, fixation was carried out in 2.5% glutaraldehyde and 4% formaldehyde in 0.1 M cacodylate buffer for 48 h. Cells were washed and post-fixed in 1% osmium tetroxide containing 1.25% potassium ferrocyanide for 1 h, followed by dehydration in ascending acetone concentrations (15–100%, 20 min per step). Dehydrated samples were embedded in Spurr resin, sectioned into ultrathin slices, and contrasted with uranyl acetate. Imaging was obtained using a Hitachi HT 7800 transmission electron microscope (Hitachi High-Technologies Corporation, Tokyo, Japan), and micrographs were digitally processed using Adobe Photoshop.
4.8. Flow Cytometry Analysis
To assess lipid-related alterations, yeast cells subjected to the same treatment conditions described above were processed for flow cytometry. Cells were washed three times with PBS and incubated with 20 μM BODIPY 496/503 (Thermo Fisher Scientific, Waltham, MA, USA) for 30 min at room temperature in the absence of light. Following staining, cells were washed, fixed in 2% formaldehyde in PBS, and washed again. Data acquisition was conducted using a BD Accuri™ C6 flow cytometer (BD Biosciences, San Jose, CA, USA), with 10,000 events recorded per sample. Analyses were carried out using BD Accuri C6 1.0 software. Experiments were independently repeated three times. Statistical comparisons were performed using one-way ANOVA followed by Tukey’s post hoc test, adopting a significance threshold of p < 0.05 (GraphPad Prism 8.4, GraphPad Software Inc., San Diego, CA, USA).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bongomin F. Gago S. Oladele R.O. Denning D.W. Global and Multi-National Prevalence of Fungal Diseases-Estimate Precision J. Fungi 201735710.3390/jof 304005729371573 PMC 5753159 · doi ↗ · pubmed ↗
- 2Denning D.W. Global incidence and mortality of severe fungal disease Lancet Infect. Dis.202424 e 428e 43810.1016/S 1473-3099(23)00692-838224705 · doi ↗ · pubmed ↗
- 3Rauseo A.M. Coler-Reilly A. Larson L. Spec A. Hope on the Horizon: Novel Fungal Treatments in Development Open Forum Infect. Dis.20207 ofaa 01610.1093/ofid/ofaa 01632099843 PMC 7031074 · doi ↗ · pubmed ↗
- 4Liu N. Wang C. Su H. Zhang W. Sheng C. Strategies in the discovery of novel antifungal scaffolds Future Med. Chem.201681435145410.4155/fmc-2016-002027463376 · doi ↗ · pubmed ↗
- 5Rodrigues A.M. Gonçalves S.S. de Carvalho J.A. Borba-Santos L.P. Rozental S. de Camargo Z.P. Current Progress on Epidemiology, Diagnosis, and Treatment of Sporotrichosis and Their Future Trends J. Fungi 2022877610.3390/jof 808077635893145 PMC 9331723 · doi ↗ · pubmed ↗
- 6World Health Organization WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action WHO Geneva, Switzerland 2022
- 7Borba-Santos L.P. Reis de SáL.F. Ramos J.A. Rodrigues A.M. de Camargo Z.P. Rozental S. Ferreira-Pereira A. Tacrolimus Increases the Effectiveness of Itraconazole and Fluconazole against Sporothrix spp.Front. Microbiol.20178175910.3389/fmicb.2017.0175928966608 PMC 5605639 · doi ↗ · pubmed ↗
- 8Benelli J.L. Poester V.R. Munhoz L.S. Melo A.M. Trápaga M.R. Stevens D.A. Xavier M.O. Ebselen and diphenyl diselenide against fungal pathogens: A systematic review Med. Mycol.20215940942110.1093/mmy/myaa 11533421963 · doi ↗ · pubmed ↗