Bovine-Derived Acinetobacter indicus Co-Harboring Chromosomal tet(X3) and Plasmid-Located tet(X4) Isolated from Henan, China
Qing Wang, Guonian Dai, Yanhua Qiu, Yaxin Zhou, Jing Xu, Weiwei Wang, Jiyu Zhang

TL;DR
A new strain of Acinetobacter indicus from a cattle farm in China was found to carry two tigecycline resistance genes, raising concerns about antibiotic resistance spread.
Contribution
First report of a strain co-harboring chromosomal tet(X3) and plasmid-borne tet(X4) in Acinetobacter.
Findings
The strain showed resistance to multiple antibiotics including tigecycline and omadacycline.
Genomic analysis revealed tet(X3) on the chromosome and tet(X4) on a plasmid with multiple resistance genes.
The tet(X4)-positive plasmid could not be transferred to other bacteria in conjugation experiments.
Abstract
The coexistence of antibiotic resistance genes (ARGs), particularly those conferring resistance to last-resort antibiotics, is increasingly present in lesser-studied bacterial species. Tigecycline is currently one of the last important barriers in the treatment of carbapenem-resistant bacterial infections, whose resistance gene, tet(X), is prevalent across multiple bacterial genera, but the coexistence of tet(X3) and tet(X4) in Acinetobacter sp. is rarely observed. Here, we report a strain co-harboring the chromosomal tet(X3) and plasmid-borne tet(X4) isolated from a commercial beef cattle farm in Henan province, China. The strain exhibited resistance to ampicillin, gentamicin, chloramphenicol, sulfamethoxazole, tetracycline, doxycycline, tigecycline, and omadacycline. Based on whole-genome sequencing (WGS), the strain was identified as A. indicus using Average Nucleotide Identity (ANI)…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
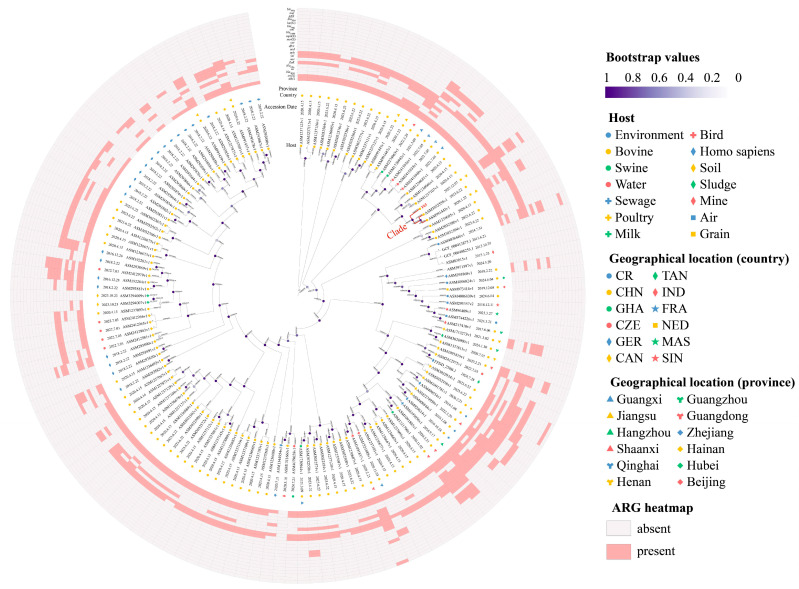 Figure 1
Figure 1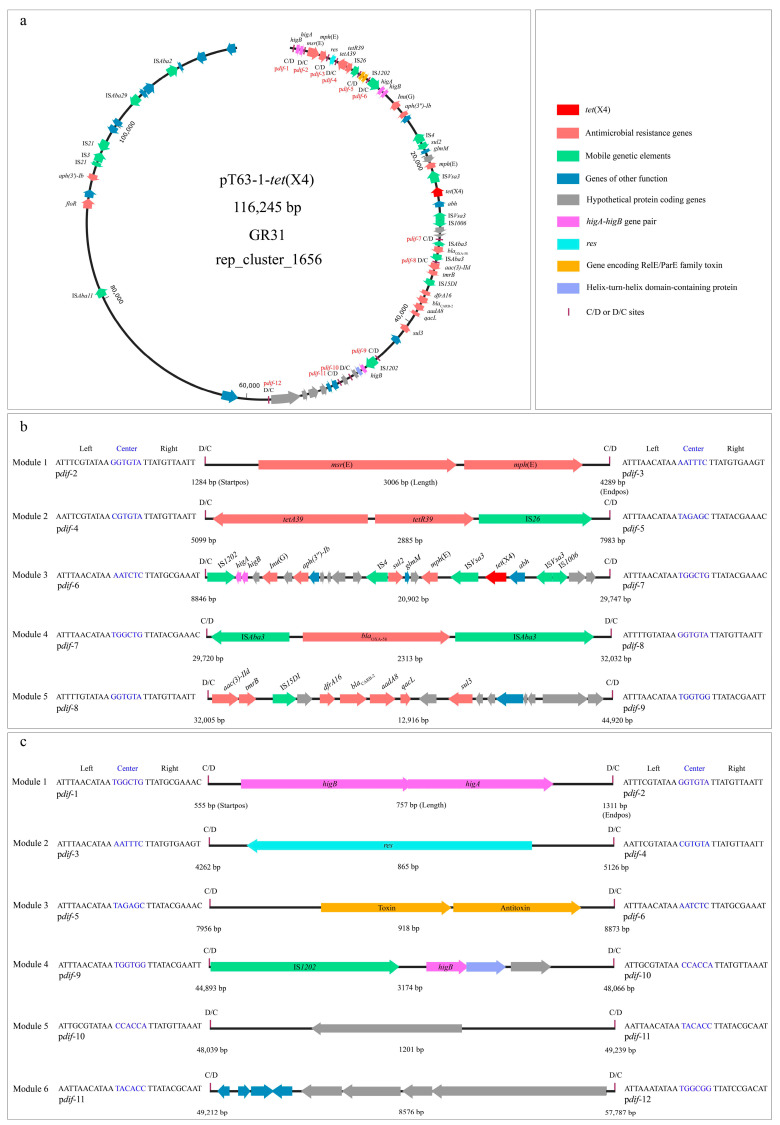 Figure 2
Figure 2 Figure 3
Figure 3- —National Natural Science Foundation of China
- —Provincial Talent Project of Gansu Province
- —China Agriculture Research System (CARS)
- —National Key Research and Development Program of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Pharmaceutical and Antibiotic Environmental Impacts · Antibiotic Use and Resistance
1. Introduction
Tigecycline, a broad-spectrum modified minocycline derivative, is considered a drug of last resort against multidrug-resistant bacteria, especially carbapenem-resistant bacteria (CRB) [1]. However, the emergence of plasmid-mediated tigecycline resistance gene tet(X) undermined this key line of defense. In particular, the novel tigecycline resistance genes tet(X3) and tet(X4) have recently been identified in Acinetobacter and Enterobacteriaceae from animals and humans [2]. They exhibit a more potent tigecycline resistance phenotype than the earlier-detected variants (such as tet(X) and tet(X2)) and have rapidly become the most epidemic variant within the tet(X) family [3]. These genes were first discovered in China in 2018 [1] and have been detected as widely distributed across dozens of provinces, including Liaoning, Hebei, Henan, Shanxi, Jiangsu, Zhejiang, Shaanxi, Gansu, Sichuan, Hunan, Hubei, Guangxi, and Guangdong [4,5]. In addition to China, this gene has been widely distributed across dozens of countries worldwide, including Canada, Thailand, the Republic of Korea, Iran, South Africa, Turkey, Iraq, the United Kingdom, Pakistan, Norway, and Singapore [6,7]. So far, human-, animal-, and environment-derived tet(X3) and tet(X4) have been reported in hospitals [8], farms [4,5], slaughterhouses [9], and supermarkets [10].
Acinetobacter comprises a complex, heterogeneous group of bacteria and can colonize the skin, throat, respiratory, and digestive tracts of humans and animals [11]. Acinetobacter can spread through food production, processing, and storage; through the hands of healthcare personnel; and through cross-contamination of medical devices, making it a significant contributor to nosocomial infections [12]. In recent years, Acinetobacter has attracted increasing public attention due to its frequent association with genes conferring resistance to last-resort antibiotics [13,14,15]. Furthermore, a new transfer mechanism of antibiotic resistance, associated with intracellular mobilization, has been identified in Acinetobacter [16,17]. This mechanism involves the pdif-ARG module flanked by XerCD site-specific recombination sites, which is mediated by the XerCD site-specific recombination (XerCD SSR) system [18]. XerC and XerD are encoded by numerous bacteria, usually in pairs. They are homologous recombinases (tyrosine recombinase family), which catalyze the cleavage of two consecutive pairs of DNA strands and exchange with a restriction site, dif, located in the terminus region of the chromosome [18]. Typically, the dif site is a 28 bp site consisting of two inverted repeat 11 bp Xer-binding motifs (the left and right regions of C/D and D/C), separated by a six bp interval called the central region. A monomer of XerC and XerD each binds to an 11 bp semi-binding site [19]. Usually, a single dif site is involved in dimeric chromosome resolution [19]. The dif sites in plasmids are called pdif sites, appear multiple times in a plasmid, and are considered to be associated with the transfer of resistance genes [20].
Here, we exhibit the multidrug-resistant A. indicus co-harboring chromosomal tet(X3) and plasmid-borne tet(X4) isolated from a commercial beef cattle farm in the Henan province of China.
2. Methods and Materials
Sampling and microbial identification. Cattle feces samples were collected from a commercial beef cattle farm in Henan province, China, in 2023. Specimens were preserved and transported in Brain Heart Infusion (BHI) broth (HuanKai Microbial, Guangzhou, China) to maintain microbial viability. Strain was isolated from MacConkey (HuanKai Microbial, Guangzhou, China) plates supplemented with tigecycline (2 mg/L, Solarbio, Beijing, China) with incubation for 16 h at 37 °C. A universal primer pair [1] was used to amplify the tet(X) gene in presumptive positive strains, followed by Sanger sequencing confirmation.
Whole-genome sequencing. Long-read sequencing was performed on the Oxford Nanopore platform. Briefly, the genome was sequenced using the Oxford Nanopore and DNBSEQ (short-read sequencing) platforms (Beijing Genomics Institution, Beijing, China). The corrected reads were generated by hybrid assembly in combination with DNBSEQ short reads. The assembled genome was checked for completeness and contamination using CheckM (v 1.2.3). The draft genome was assembled using SPAdes (v 3.9.0) with default parameters. Genome assembly and plasmid circularization detection were performed using Canu (v 2.2) with default parameters.
Bacterial species identification. Bacterial species identification was carried out on the JSpeciesWS and Deutsche Sammlung von Mikroorganismen und Zellkulturen (DSMZ) platforms using tetra correlation search (TCS) and type strain genome server (TYGS). The Average Nucleotide Identity (ANI) and digital DNA–DNA hybridization (dDDH) were calculated on the EZBioCloud and the DSMZ platforms using the OrthoANIu algorithm and genome-to-genome distance calculator (GGDC).
Antimicrobial susceptibility testing. The minimum inhibitory concentration (MIC) of 18 antibiotics (meropenem, aztreonam, ampicillin, ceftazidime, cefepime, gentamicin, chloramphenicol, colistin, kanamycin, fosfomycin, ciprofloxacin, sulfamethoxazole, azithromycin, tetracycline, doxycycline, tigecycline, rifampin, and omadacycline) was determined using microdilution of Mueller-Hinton broth (Huan Kai Microbial, Guangzhou, China). Microdilution was performed according to the method recommended by Clinical and Laboratory Standards Institute (CLSI). The testing concentration range is 2 mg/L to 1024 mg/L. Results were interpreted according to CLSI documents M100-S34 (2024) [5]. Since there is no established breakpoint for the tigecycline and omadacycline resistance of Acinetobacter, their breakpoints were determined by FDA-defined interpretive criteria for Enterobacteriaceae (R ≥ 8 mg/L for tigecycline and R ≥ 16 mg/L for omadacycline). E. coli ATCC25922 was used as a quality control for antimicrobial susceptibility testing, and the quality control range for ATCC25922 was based on the CLSI criterion. An independent colony was picked for testing, and MIC determinations were performed in triplicate.
Conjugation assay. Conjugation assays were performed using the filter mating method. E. coli J53 (sodium azide-resistant) and Salmonella LGJ2 (rifampicin-resistant) were used as recipients to observe the transfer of tet(X) to Enterobacteriaceae. Bacteria solution with a donor-to-recipient ratio of 10:1 (tet(X)-positive strain to J53 or LGJ2) was incubated on a 0.22 μm filter membrane at 35 °C for 16 h, then moved to MacConkey plates supplemented with antibiotics for which the donor (4 mg/L tigecycline) and recipient (100 mg/L sodium azide or 200 mg/L rifampicin) are resistant. The donor-to-recipient ratios of 5:1 and 15:1 were supplemented to test. At least three attempts were made for each parameter, and three parallel experiments were made for each attempt.
Phylogenetic tree construction. A maximum-likelihood (ML) phylogenetic tree based on core-genome single-nucleotide polymorphisms (cgSNP) was constructed using the A. indicus genome T63 and all A. indicus genomes stored in the NCBI Genome Database. SNPs were yielded using Snippy (v 4.6.0) with default parameters, and strain T63 was used as the reference genome. Each genome produced a core genome of about 60 kb. Snippy-core (v 4.6.0) was used to combine multiple Snippy outputs into a core SNP alignment. SNP distance matrix (v 0.8.2) was used to compute distance in SNPs between all sequences. Recombination filtering was not applied because it prevented a sufficient number of core genomes from being yielded. The ML phylogenetic tree was built with FastTree (v 2.2) and further visualized in iTOL (v 7) to highlight the features of each strain.
Bioinformatics analysis. ResFinder (v 4.6.0) was used to screen all known acquired antibiotic resistance genes (ARGs). The ARG identification threshold was set to 90%, and the minimum length was set to 80%. MOB-typer was used to search replicon, oriT, relaxase, T4CP, and T4SS, as well as to type plasmid. Furthermore, another plasmid typing based on a Rep-based strategy (designed for A. baumannii) [21,22] was performed using SnapGene (GR BLASTn). Pdif sites were searched on the pdifFinder platform. Genome annotation was performed using the RAST genome annotation service, with further manual correction using ORFfinder, UniProt, and ISFinder. The linear genomic comparison was created using Easyfig (v 2.2.5).
3. Results
In August 2023, we isolated a tigecycline-resistant strain, T63, from a cattle fecal sample collected at a commercial beef cattle farm in Henan province, China. PCR amplification and Sanger sequencing verified that the strain carried the tet(X) gene.
The identification of bacterial species. The strain was sequenced using WGS on the Oxford Nanopore platform (long-read sequencing technology). The assembled genome was confirmed to be 99.89% completeness and 0.68% contamination. Current standards require ANI > 95% and dDDH > 70% to robustly assign a novel strain to a species. Thus, the genome was searched on the JSpeciesWS and DSMZ platforms using TCS and TYGS. Results revealed that strain T63 best matched GCF_000369465.1, GCF_000488255.1, GCF_000830155.1, and GCF_000413875.1. These genomes registered as Acinetobacter indicus. Subsequently, the ANI and dDDH values between genome T63 and these genomes were analyzed on the EZBioCloud and DSMZ platforms using the OrthoANIu algorithm and DDGC. Results found that genome T63 exhibited > 95% ANI and >70% dDDH with each of these genomes, with the highest ANI and dDDH values with GCF_000369465.1 (ANI: 97.98%, dDDH: 82.1%). Based on the above analyses, strain T63 was identified as A. indicus within the Acinetobacter genus under family Moraxellaceae (NCBI taxonomy database).
Genetic diversity analysis. The 408X coverage was obtained for short-read sequencing, and the 344X coverage was obtained for long-read sequencing. The WGS of strain T63 yielded one circular chromosome (2,986,742 bp, 45.9% GC content) and three circular plasmids (pT63-1: 116,245 bp, 41.8%; pT63-2: 13,463, 38.7%; pT63-3: 11,175, 35.1%) (Table 1). The GC contents of pT63-2 and pT63-3 were significantly lower than that of the chromosome, hinting that they originated from different hosts. GC content can serve as a marker to assess the origin of plasmids, indicating potential horizontal transfer [23]. Tigecycline resistance genes, tet(X3) and tet(X4), were located in the chromosome and plasmid (pT63-1) of strain T63, respectively (Table 1). A total of 19 resistance genes were identified in strain T63, among which 4/19 (23.53%) genes, including tet(X3), aph(6)-Id, aph(3″)-Ib and sul2, were located in the chromosome, and 2/19 (11.76%) genes, including msr(E) and mph(E) were located in the pT63-2 (Table 1). Notably, 17/19 (89.47%) genes, including aac(3)-IId, aph(3′)-Ib, aph(3″)-Ib, aadA8, blaCARB-2, blaOXA-58, msr(E), mph(E), lnu(G), floR, sul2, sul3, tet(X4), tet(39), dfrA16, tmrB, and qacL were located in the pT63-1 (Table 1). No resistance gene was identified in the pT63-3 (Table 1). The tet(X4)-positive plasmid (pT63-1) accounted for 89.47% of the resistant genes carried by the strain, reflecting co-selection-driven plasmid persistence.
Antibiotic resistance phenotype. The antimicrobial susceptibility testing showed that strain T63 was resistant to ampicillin (256 mg/L), gentamicin (8 mg/L), chloramphenicol (64 mg/L), sulfamethoxazole (1024 mg/L), tetracycline (64 mg/L), doxycycline (16 mg/L), tigecycline (8 mg/L), and omadacycline (32 mg/L) (Table 2). However, it was susceptible to meropenem, aztreonam, ceftazidime, cefepime, colistin, kanamycin, fosfomycin, ciprofloxacin, azithromycin, and rifampin (Table 2).
Phylogenetic analysis. There were only 138 A. indicus genomes stored in the NCBI genome database. These genomes and T63 were used to build an ML circular phylogenetic tree based on cgSNPs to understand the genetic similarity of strain T63. The analysis found that strain T63 exhibited genetic similarity with ASM3032558v1 and ASM991443v1, which was characterized by the low level of core genome diversity that was reflected by a limited number of single-nucleotide polymorphisms (SNPs ≤ 340) (Figure 1). These strains showed similar isolation sources (bovine) and geographical locations (China) to strain T63 (Figure 1). Notably, these strains and strain T63 exhibited a similar resistance gene profile. They carried tet(X3), sul2, aph(3″)-Ib and aph(6)-Id, particularly ASM991443v1 and T63 additionally carrying aac(3)-IId, blaOXA58, floR, msr(E) and mph(E).
Plasmidome analysis. The plasmid typing based on MOB-Typer revealed that pT63-1-tet(X4) was typed to rep_cluster_1656, pT63-2 to rep_cluster_1254, and pT63-3 to an unidentified type (Table 1). The oriT of these plasmids could not be identified. The relaxases of pT63-1-tet(X4) and pT63-2 were identified as MOBP and MOBQ, which belonged to the MOB family (Table 1). The pT63-1-tet(X4) and pT63-2 were predicted as mobilizable, and pT63-3 was predicted as non-mobilizable (Table 1). Subsequently, plasmid typing based on a Rep-based strategy (designed for A. baumannii) revealed that pT63-1-tet(X4) best matched GR31 (99.66% nucleotide identity and 100% coverage), a RepB-family plasmid replication initiator protein. The remaining plasmids did not best match the identified GRs of the Rep-based strategy (<50% nucleotide identity and coverage). Conjugation assay demonstrated that the tet(X4)-positive plasmid could not be transferred to recipients, E. coli J53 or Salmonella LGJ2, despite multiple experimental parameters and repeated attempts employed.
Pdif-ARG module analysis. The analysis based on pdifFinder revealed a pdif module island composed of 12 pdif sites (Figure 2a–c), containing 11 consecutively arranged pdif modules (Figure 2a). This special configuration inevitably leads to sharing the internal C/D or D/C sites to form two types of pdif modules, one flanked by a C/D and a D/C site, and the other type flanked by a D/C and a C/D site (Figure 2a). These modules included five pdif-ARG modules, pdif-msr(E)-mph(E) (length: 3006 bp), pdif-tetA39-tetR39-IS26 (2885 bp), pdif-Inu(G)-aph(3″)-Ib-sul2-tet(X4) (20,902 bp), pdif-blaOXA-58 (2313 bp) and pdif-aac(3)-IId-dfrA16-blaCARB-2-aadA8-sul3 (12,916 bp) (Figure 2b). Among these modules, pdif-Inu(G)-aph(3″)-Ib-sul2-tet(X4) and the pdif-aac(3)-IId-dfrA16-blaCARB-2-aadA8-sul3 modules have not been reported, with no similar segments (<70% identity and coverage) identified in the NCBI database. Pdif-msr(E)-mph(E), pdif-blaOXA-58, and pdif-tetA39-tetR39 have been described in previous studies, and similar segments (>99% identity and coverage) could be identified in the NCBI database. Thus, based on the high similarity (>99% identity and coverage), multiple segments containing these modules from chromosomes and plasmids of Acinetobacter were used to compare with those in the tet(X4)-positive plasmid (Figure 3a). The analysis found that pdif-msr(E)-mph(E) and pdif-blaOXA-58 modules were highly conserved (>99% identity and coverage) in the chromosomes and plasmids of various Acinetobacter (A. pittii, A. baumannii, and A. towneri) (Figure 3a). However, the pdif-tetA39-tetR39-IS26 module in pT63-1-tet(X4) observably differed from the typical pdif-tetA39-tetR39 module [20] due to the IS26 downstream of tetR39 (Figure 3a). A similar characteristic was identified in the pM2-blaNDM-1 (CP183241.1) [15], in which the pdif-ISAjo2-higA-higB-msr(E)-mph(E) module was significantly different from the typical module only formed by msr(E)-mph(E) (Figure 3a).
In addition to the pdif-ARG module, six pdif modules carrying genes encoding other functional proteins or hypothetical proteins were identified. Module one (757 bp) harbored a gene pair of higA-higB, which belongs to Type II toxin–antitoxin systems. Module two (865 bp) carried the res gene encoding resolvase (Figure 2c). Module three (918 bp) contained genes encoding the RelE/ParE toxin and RelB/ParD antitoxin (Figure 2c). Notably, the above modules typically appear as independent units and are closely adjacent to the Xer-ARG module (Figure 3a). Module four (3174 bp) carried IS1202, higB, and the genes encoding helix-turn-helix domain-containing protein and hypothetical protein (Figure 2c). The orphaned toxin (higB) may interact with the antitoxin located at another site on the plasmid. Module five (1201 bp) only contained a gene encoding a hypothetical protein (Figure 2c). Module six (8576 bp) contained genes encoding recombinase, DUF1778 domain-containing protein, GNAT family N-acetyltransferase, and hypothetical proteins (Figure 2c).
The genetic contexts of chromosomal tet(X3). Tet(X3) was located within a typical genetic context (5949 bp), ISVsa3-XerD-tet(X3)-res-ISVsa3, in the chromosome of strain T63 (Figure 3b). BLASTn analysis of the center region, XerD-tet(X3)-res, in the NCBI database found that this region was generally located on chromosomes and plasmids of various Acinetobacter species. Based on the high similarity (>99.00% identity and coverage) of this region, multiple segments (long-read sequencing) containing the upstream and downstream regions of the tet(X3) genetic context from Acinetobacter plasmids (A. baumannii, A. gandensis, A. variabilis, A. schindleri, and A. portensis) were used to compare with those in the tet(X3)-positive chromosome (Figure 3b). Among these plasmids, 4/7 plasmids could not be typed using the MOB-Typer and Rep-based strategy (Figure 3b). The remaining plasmids (3/7) were typed as Rep_cluster_1656, Rep_cluster_1481, and Rep_cluster_481 using MOB-Typer (Figure 3b). Only one plasmid was typed as GR26 (>99.00% identity and coverage) using the Rep-based strategy (Figure 3b). The typical tet(X3) genetic context was two copies of ISVsa3 flanking XerD-tet(X3)-res, which were invariably in the same orientation (Figure 3b). Usually, in this context, upstream ISVsa3 was either intact (1494 bp) or truncated (216–845 bp), but downstream ISVsa3 was commonly intact (1494 bp) (Figure 3b). Furthermore, various other genetic contexts could also be observed, such as a single ISVsa3 (intact and truncated) located upstream or downstream of tet(X3) (pXM9F202-2 and pJNE5); the upstream or downstream ISVsa3 replaced by other IS elements like IS26 or IS1008 (pHZE30-1-1 and pD1) (Figure 3b). Notably, a structure change in the center region, XerD-tet(X3)-res, was observed in pD1, in which XerD shifted to downstream of intact ISVsa3, and only tet(X3)-res was conserved (Figure 3b). The upstream and downstream regions of the tet(X3) genetic context exhibited significant differences, but multiple IS elements and resistance genes were commonly adjacent to the tet(X3) genetic context (Figure 3b).
The genetic contexts of plasmid-mediated tet(X4). Tet(X4) was located in a genetic context (5440 bp) with the gene arrangement, ISVsa3-abh-tet(X4)-ISVsa3, in the plasmid of A. indicus T63 (Figure 2a). An IS1006 in the opposite direction to ISVsa3 was closely adjacent to the upstream ISVsa3 (Figure 2a). Mph(E)-glmM-sul2-IS4 was located in the region adjacent to downstream ISVsa3 (Figure 2a). Analysis of the center region, abh-tet(X4), in the NCBI database found that it was typically carried by chromosomes and plasmids of Enterobacteriaceae, as well as the chromosomes of Acinetobacter. However, it was rarely located in the Acinetobacter plasmids. Thus, based on the high similarity (>99.00% identity and coverage) of this region, multiple segments (long-read sequencing) containing the upstream and downstream regions of tet(X4) genetic context from these typical bacterial vehicles (chromosomes: A. indicus, A. towneri, and Aeromonas caviae; plasmids: E. coli (IncX1, IncQ1, IncFIA), Klebsiella aerogenes (IncR), and Salmonella enterica (IncHI2A-IncN) were used to compare with those in the tet(X4)-positive Acinetobacter plasmid (Figure 3b). The analysis revealed that the center region, abh-tet(X4), was highly conserved in Acinetobacter and Enterobacteriaceae (Figure 3b). Similarly to the tet(X3) genetic context, the diversity of tet(X4) genetic context was mainly mediated by the upstream and downstream ISVsa3 of tet(X4), with the following configurations: two intact copies of ISVsa3 flanked tet(X4); an intact and an truncated ISVsa3 flanked tet(X4); a replacement of the upstream or downstream ISVsa3 by other IS elements, such as IS1R or IS15DI; the absence of the upstream or downstream ISVsa3 (Figure 3b). The genes located within the upstream and downstream regions of the tet(X4) genetic context in Enterobacteriaceae plasmids were complex and versatile. In contrast, the genes located within the upstream and downstream regions of the tet(X4) genetic context in Acinetobacter were relatively stable and conserved, such as the high occurrence frequency of sulP and IS1006 in the upstream region; the high conservation of mph(E)-glmM-sul2-IS4 in the downstream region (Figure 3b). This hints that the transmission of tet(X4) in Enterobacteriaceae may be more frequent than that in Acinetobacter. Furthermore, the gene pair, glmM-sul2, emerged frequently in the downstream regions of genetic contexts for tet(X3) and tet(X4) and was conserved across Acinetobacter chromosomes and plasmids (Figure 3b).
4. Discussion
Livestock is recognized as a critical reservoir for tigecycline-resistant bacteria [4,5], yet investigations into bovine production remain limited. The current study reveals that bovine-derived A. indicus co-harbors the chromosomal tet(X3) and the plasmid-located tet(X4). Tigecycline is regarded as a crucial last-resort antibiotic in human medicine, but its use in food-producing animals is not authorized worldwide [4]. It is noteworthy that the detection rate of the tet(X) correlates positively with the frequency of tetracycline antibiotic use, indicating that tetracycline utilization is a driving force promoting the enrichment and dissemination of the tet(X) [4,24]. In the livestock industry, older tetracycline antibiotics, such as chlortetracycline and oxytetracycline, remain extensively employed as feed additives or therapeutic drugs, which may drive the co-occurrence of tet(X3) and tet(X4). The tet(X3) is initially detected in the A. baumanii plasmid (p34AB), and tet(X4) is first identified in the E. coli plasmid (p47EC) [1]. Although the rapid transmission of these genes, tet(X4), exhibits a more formidable dissemination capability [25]. Compared with the typical location of tet(X3) in the chromosomes and plasmids of Acinetobacter [26,27], tet(X4) is rarely carried by the Acinetobacter plasmid. The tet(X4) can be identified in the Acinetobacter chromosome, but it is commonly located in the chromosomes and plasmids of Enterobacteriaceae, such as Escherichia [4,5], Klebsiella [28], and Enterobacter [29]. Enterobacteriaceae constitute typical flora within animal gut systems and environmental habitats, thereby subjecting them to more direct and persistent exposure to the selective pressure exerted by antibiotics. Although Acinetobacter, as a commensal bacterium, can colonize the digestive system together with Enterobacteriaceae, its colonization level is significantly lower than that of Enterobacteriaceae [11]. Moreover, current research on antibiotic resistance has focused on Enterobacteriaceae. However, Acinetobacter, a heterogeneous bacterial genus widely distributed in human and animal communities with robust environmental adaptability and survival capabilities [30], has been neglected (reflected in a limited number of genomes in the NCBI database), which may lead to an underestimation of the spread of tet(X3) and tet(X4) in this genus.
Tet(X4) is typically located in a wide variety of Inc plasmids, including single replicon and multireplicon plasmids, such as IncX1, IncFIA, IncHI1A, IncHI1B, IncR, IncN, IncHI1A-IncR, IncX1-IncN, IncX1-IncR, etc. [5]. Inc plasmids are considered the dominant vector for the spread of tet(X4) and contribute to the global epidemic of tet(X4) [24,31]. These plasmids are usually seen in Enterobacteriaceae and emphasize the plasmid replication and the coexistence of multiple replicons, with a diversified host range. They can carry resistance genes and facilitate the horizontal transfer of resistance genes among different bacterial genera via conjugation [24]. Inc plasmids commonly exhibit optimal replication and maintenance fitness within Enterobacteriaceae. In contrast to Inc plasmids, Acinetobacter plasmids exhibit genus-specificity and primarily adapt, persist, and evolve within the genus Acinetobacter, with a narrow host range [32]. This study failed to observe the transfer of the tet(X4)-positive Acinetobacter plasmid via conjugation assays. Meanwhile, the MOB-Typer analysis indicated that this plasmid lacked oriT and was predicted to be mobilizable rather than conjugative. Compared to the commonly observed transferability of Inc plasmids, Acinetobacter plasmids were rarely reported to be conjugative [33]. However, this is inconsistent with established biological principles: (1) conjugative plasmids serve as fundamental drivers of bacterial evolution by facilitating the dissemination of antibiotic resistance genes, heavy metal tolerance determinants, and virulence factors, which enables bacterial adaptation to diverse environmental and selective pressures [34]; (2) a wide variety of plasmids, whether conjugative or not, have been observed in Acinetobacter genomes [35]. Laboratory systems may lack key microbial ecological drivers, such as nutrient gradients, multispecies competitive dynamics, and stress-induced epigenetic regulatory pathways, resulting in the rarely observed transferability of Acinetobacter plasmids. Furthermore, recent studies revealed that natural transformation, a horizontal gene transfer mechanism highlighted in A. baumannii, enables highly efficient interbacterial transfer of genetic elements conferring resistance to last-line carbapenem antibiotics. This should be considered for the other gene conferring resistance to last-resort antibiotics and the other species of Acinetobacter [36].
The dissemination of tet(X3) and tet(X4) critically depends on the flanking insertion sequence, ISVsa3. This element can mobilize adjacent genes via a rolling-circle transposition mechanism [1]. Most IS elements typically require two copies (one of which must be intact) to complete transposition, such as the transposition of mcr-1 mediated by ISApl1 [37], whereas ISVsa3 can achieve transposition of adjacent DNA sequences using a single copy [38]. In addition to the tet(X) family, ISVsa3 is associated with multiple ARGs, such as floR, tet(A), aph(6)-Id, aph(3″)-Ib, and sul2 [39]. Thus, monitoring ISVsa3 is critical for understanding the distribution and spread of ARGs, particularly tet(X). The various tet(X3) and tet(X4) genetic contexts have been found in Enterobacteriaceae and Acinetobacter, which may be attributed to the dynamic IS elements adjacent to ISVsa3 and from recurrent rearrangement events. The upstream and downstream regions of the tet(X3) genetic context in Acinetobacter exhibit more complexity and diversity than that of the tet(X4) genetic context, suggesting that the transmission of tet(X3) in this genus may be more dynamic than that of tet(X4). Furthermore, the glmM-sul2 gene pair is potentially associated with the transfer of tet(X3) and tet(X4) in Acinetobacter, as it is frequently located in the downstream regions of the genetic contexts of tet(X3) and tet(X4) within the chromosomes and plasmids of this genus.
A total of 15 resistance genes were identified in the tet(X4)-positive plasmid, of which 13/15 genes were located in the pdif modules. Previous studies [40] indicated that the pdif modules identified in Acinetobacter have a genus-specific characteristic. Among these modules, pdif-msr(E)-mph(E) and pdif-blaOXA-58 modules were highly conserved (>99% identity and coverage) with the Xer-ARG modules identified in the chromosomes and plasmids of Acinetobacter from the GenBank database, suggesting their potential mobilization. Pdif-tetA39-tetR39-IS26 module exhibited a difference with the typical module only formed by tetA39-tetR39 due to the presence of IS26. A similar characteristic is also observed in pdif-ISAjo2-higA-higB-msr(E)-mph(E) module [15], indicating that the elements located in the pdif module are variable and dynamic. Pdif-Inu(G)-aph(3″)-Ib-sul2-tet(X4) module has not been matched in the NCBI database. However, an intact tet(X4) genetic context was identified in this module, which may contribute to the effective expression of tigecycline resistance and the maintenance of tet(X4) transfer capability [15]. The genes for which resistance phenotypes were tested exhibited resistance to the corresponding antibiotics. Among these, some genes are located in pdif modules, suggesting that these modules can perform their functions in antibiotic resistance.
Despite the findings, this study has several limitations. First, although antibiotic resistance phenotypes and genotypes were tested, it was not determined which gene was responsible when multiple genes were associated with the resistance phenotype. Second, the current inadequate plasmid typing methods constrain the study of Acinetobacter resistance. Inc testing is only suitable for Enterobacteriaceae plasmids, not for Acinetobacter plasmids [32]. In this study, we employed the MOB-typer and Rep-based strategy (designed for A. baumannii plasmids) [21,22], but these methods only classify plasmids into distinct Rep clusters and are not precise enough to cover the full range of plasmids. Furthermore, MOB-typer is insufficient to cover various elements, such as Rep, oriT, and relaxase, in Acinetobacter plasmids. Therefore, the predicted mobility should only be considered as a reference. Although a novel typing method for the Acinetobacter plasmid is needed, the extremely limited genomic data may delay its development. Third, phenotypic resistance research in Acinetobacter was constrained by the lack of interpretive breakpoints for most antibiotics in standard guidelines (CLSI, EUCAST, FDA). Fourth, FastTree is acceptable for rapid phylogenetic inference but is less accurate for fine-scale analyses. Fifth, the toxin–antitoxin systems observed are identical to those previously described in other sources, or are they only homologues, in which case the function or mode of action can only be inferred.
5. Conclusions
The current study exhibited that A. indicus co-carried the chromosomal tet(X3) and the plasmid-borne tet(X4). Genome analysis and antimicrobial susceptibility testing confirm the association between these genes and resistant phenotypes. The continuous emergence of lesser-studied bacterial species carrying genes conferring resistance to last-resort antibiotics, particularly those co-harboring these genes, is a global public health issue that cannot be ignored and, therefore, warrants more urgent attention.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1He T. Wang R. Liu D. Walsh T.R. Zhang R. Lv Y. Ke Y. Ji Q. Wei R. Liu Z. Emergence of Plasmid-Mediated High-Level Tigecycline Resistance Genes in Animals and Humans Nat. Microbiol.201941450145610.1038/s 41564-019-0445-231133751 · doi ↗ · pubmed ↗
- 2Sun C. Cui M. Zhang S. Wang H. Song L. Zhang C. Zhao Q. Liu D. Wang Y. Shen J. Plasmid-Mediated Tigecycline-Resistant Gene Tet (X 4) in Escherichia Coli from Food-Producing Animals, China, 2008–2018 Emerg. Microbes Infect.201981524152710.1080/22221751.2019.167836731631781 PMC 6818123 · doi ↗ · pubmed ↗
- 3Cui C.-Y. He Q. Jia Q.-L. Li C. Chen C. Wu X.-T. Zhang X.-J. Lin Z.-Y. Zheng Z.-J. Liao X.-P. Evolutionary Trajectory of the Tet(X) Family: Critical Residue Changes towards High-Level Tigecycline Resistancem Systems 20216 e 00050-2110.1128/m Systems.00050-2134006624 PMC 8269203 · doi ↗ · pubmed ↗
- 4Sun J. Chen C. Cui C.-Y. Zhang Y. Liu X. Cui Z.-H. Ma X.-Y. Feng Y. Fang L.-X. Lian X.-L. Plasmid-Encoded Tet(X) Genes That Confer High-Level Tigecycline Resistance in Escherichia coli Nat. Microbiol.201941457146410.1038/s 41564-019-0496-431235960 PMC 6707864 · doi ↗ · pubmed ↗
- 5Sun C. Cui M. Zhang S. Liu D. Fu B. Li Z. Bai R. Wang Y. Wang H. Song L. Genomic Epidemiology of Animal-Derived Tigecycline-Resistant Escherichia Coli across China Reveals Recent Endemic Plasmid-Encoded Tet(X 4) Gene Commun. Biol.2020341210.1038/s 42003-020-01148-032737421 PMC 7395754 · doi ↗ · pubmed ↗
- 6Pan Y. Awan F. Zhenbao M. Zhang X. Zeng J. Zeng Z. Xiong W. Preliminary View of the Global Distribution and Spread of the Tet(X) Family of Tigecycline Resistance Genes J. Antimicrob. Chemother.2020752797280310.1093/jac/dkaa 28432766786 · doi ↗ · pubmed ↗
- 7Xu Y. Liu L. Sun J. Feng Y. Limited Distribution and Mechanism of the Tet X 4 Tetracycline Resistance Enzyme Sci. Bull.2019641478148110.1016/j.scib.2019.08.02436659553 · doi ↗ · pubmed ↗
- 8Ma L. Xie M. Yang Y. Ding X. Li Y. Yan Z. Chan E.W.-C. Chen S. Chen G. Zhang R. Prevalence and Genomic Characterization of Clinical Escherichia coli Strains That Harbor the Plasmid-Borne Tet(X 4) Gene in China Microbiol. Res.202428512773010.1016/j.micres.2024.12773038805981 · doi ↗ · pubmed ↗