Endocytosis Mediated by Candida albicans END3 Is Required for Its In Vivo Virulence as an Opportunistic Fungal Pathogen
Miranda Yu, Cameron Gilmore, Elena Dos Santos, Susan Eszterhas, Samuel A. Lee

TL;DR
The study shows that a gene called END3 in Candida albicans is essential for its ability to cause disease in a mouse model.
Contribution
The novel finding is that END3-mediated endocytosis is required for the in vivo virulence of Candida albicans.
Findings
The end3 null mutant of Candida albicans is hypovirulent in a murine model of disseminated candidiasis.
END3 is crucial for processes like filamentation, biofilm formation, and extracellular protease secretion in C. albicans.
Loss of END3 leads to impaired actin patch formation and reduced host-cell damage in vitro.
Abstract
Endocytic trafficking in Candida albicans is a fundamental cellular process that is crucial for its secretion, filamentation, and virulence-related processes. We have previously demonstrated that loss of the key endocytosis-related C. albicans gene END3 disrupts clathrin-mediated endocytosis, leading to impairments in actin patch formation, filamentation, biofilm formation, cell wall integrity, and extracellular protease secretion. The end3 null mutant also exhibits altered antifungal susceptibility and reduced host-cell damage in an in vitro keratinocyte infection model. To ascertain whether endocytosis is required for virulence in vivo, we assessed virulence of the C. albicans end3 null mutant in a murine model of disseminated candidiasis. After infection via the tail vein, and analysis of host survival over 28 days, the end3 null mutant was markedly hypovirulent compared to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
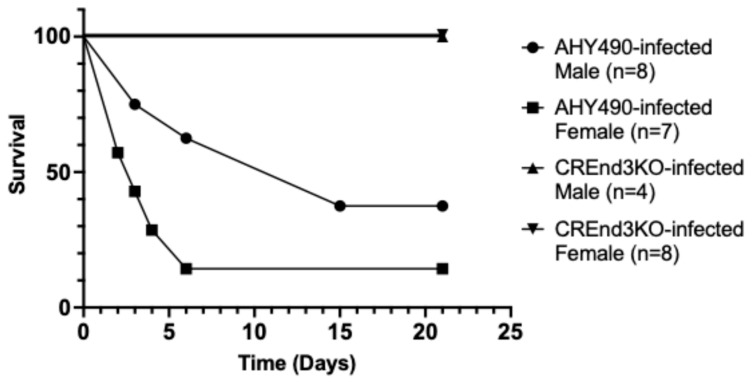 Figure 1
Figure 1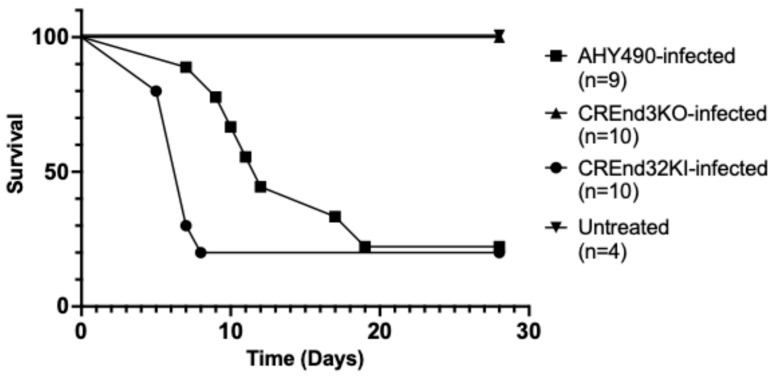 Figure 2
Figure 2- —Department of Veterans Affairs
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntifungal resistance and susceptibility · Fungal Infections and Studies · Fungal and yeast genetics research
1. Introduction
The opportunistic fungal pathogen Candida albicans is a common cause of hospital-acquired bloodstream infections (BSIs), and is a major cause of catheter-associated infections and invasive disease. Secretory pathways in C. albicans enable virulence by conducting secretion of virulence-related proteins, facilitating the highly orchestrated process of polarized secretion during filamentation, and supporting biofilm formation which in turn promotes intravascular infection and dissemination [1,2,3]. However, there is still an incomplete understanding of the molecular mechanisms of trafficking and secretion in C. albicans, and of their roles in virulence-related processes and pathogenesis.
In prior work, we demonstrated that pre-vacuolar secretion, mediated by VPS1, VPS4, and PEP12, is required for secretion of Saps and lipases, and also for filamentation and biofilm formation [1,4,5,6]. We also demonstrated that pre-vacuolar secretion is required for virulence through both in vitro and in vivo models of infection. Specifically, we demonstrated that the pre-vacuolar secretory gene VPS4 is required for virulence in a mouse tail vein model of infection [4]. Similarly, the pre-vacuolar secretory gene PEP12 is also required for in vivo virulence [5]. We next determined that late secretion plays a key role in C. albicans polarized secretion and filamentation; specifically, we determined that the exocyst-related SNARE proteins Sso2p and Sec9p are essential for viability in C. albicans, and are required for Sap and lipase secretion and filamentation [7]. Interestingly, many of these anterograde secretory pathways also appear to contribute to retrograde or endocytic pathways [1]. However, despite its importance as a fundamental cellular process, endocytosis has only been studied to a limited extent in C. albicans.
Prior studies have shown that the early endocytic coat protein encoded by EDE1 is non-essential in C. albicans. In [8], its null mutant was found to display normal growth and filamentation, though endocytosis was not specifically assayed. We studied the early endocytosis gene PAL1 and found that mutants lacking PAL1 were defective in filamentation and biofilm formation, instead forming pseudohyphal cells in filamentation-inducing conditions [9]. Additionally, the C. albicans middle coat protein encoded by SLA2 contributes significantly to filamentous growth [10]. C. albicans sla2 null mutants are defective in fluid-phase endocytosis and polarized secretion [11]. Lipid rafts normally trafficked to the hyphal tip are mis-localized. C. albicans sla2 mutants are also defective in Rbt5p-mediated uptake of iron that is based upon endocytosis [12]. Furthermore, they exhibit reduced rates of growth and form enlarged, globular mother cells because of delays in cell cycle progression mediated by the morphogenesis checkpoint kinase Swe1p [13]. In addition, we studied the C. albicans ortholog of ENT1/2 and found that C. albicans ent2 mutants lacking the middle coat protein encoded by ENT2 are defective in endocytosis, filamentation, biofilm formation, and tissue invasion in vitro [14].
The key late-coat gene SLA1 has also been investigated. C. albicans sla1 mutants were found to exhibit altered filamentation under hyphal-inducing conditions; they produce shorter filaments, along with cortical actin patches that are reduced in number and no longer concentrated at the hyphal tip [15]. Interestingly, in [16], C. albicans sla1 mutants displayed minimal defects in Lucifer Yellow uptake, but marked defects in FM4-64 internalization to the vacuole and receptor-mediated endocytosis. Another study showed that the sla1 null mutant strain was significantly attenuated in pathogenicity in a murine infection model [17]. Colonization by the sla1 mutant was also found to be significantly reduced, compared to colonization by the wild-type strain. Furthermore, the endocytosis gene PAN1, which appears to be an essential gene in C. albicans, has been studied minimally [8,18]. A conditional C. albicans MET3p-PAN1/pan1 mutant in repressing conditions developed thick, swollen cells with abnormal filamentation and failure to endocytose FM4-64 [8]. The C. albicans homolog of LAS17, WAL1, has also been studied. In [19], C. albicans wal1 null mutants were found to be defective in endocytosis, vacuolar morphology, and hyphal development. Although wal1 mutants were able to conduct polarized morphogenesis, most mutants formed pseudohyphae rather than hyphae.
Our most recent experiments focused on the endocytosis gene END3, which encodes a key late-coat endocytosis protein thought to play an important role in Sla1 recruitment and in a Pan1/End3/Sla1 complex [20]. We found that C. albicans end3 null mutants displayed abnormal actin patch formation, filamentous growth, biofilm formation, cell wall integrity, and extracellular protease secretion [21]. In an in vitro vaginal epithelial model of infection, both C. albicans ENT2 and END3 were found to be necessary for tissue invasion, thus revealing a clear link between endocytosis and key virulence-related processes [14,21]. To further define the role of endocytic pathways in pathogenesis, we now seek to define the contribution of the endocytosis gene END3 to virulence and murine survival in a mouse tail vein model of disseminated candidiasis.
2. Materials and Methods
2.1. Strains and Media
We previously generated the end3 mutant and re-integrant strains, and characterized the resultant phenotypes, in earlier work [21]. The parental C. albicans strain AHY490 was obtained directly from A. Hernday [22], while the re-integrant strain had been established through homologous recombination of alleles of END3 into the end3 null mutant. C. albicans strain AHY490, end3 null mutant (named CREnd3KO), and END3 re-integrant (named CREnd32KI) were grown at 30 °C in YPD (1% yeast extract, 2% peptone, 2% glucose), or in minimal glucose (0.67% yeast nitrogen base without amino acids [YNB], 2% glucose). Solid media were prepared by adding 2% agar.
2.2. Murine Model of Disseminated Candidiasis
All animal experiments were conducted in accordance with institutional guidelines and approved by the White River Junction VA Medical Center Institutional Animal Care and Use Committee (Approval No. 18-A262). BALB/c mice (Charles River Laboratories, Boston, MA, USA) were housed four per sterile rodent microisolator ventilated cage, with wood chip bedding and environmental enrichment (nestlets, plastic huts, and/or tubes). Mice were kept under a 12 h light/dark cycle, humidity levels between 30% and 70%, and temperatures between 68 °F and 79 °F. Mice were also provided with standard mouse chow (Teklad) and plain water ad lib. Throughout the study, mice were weighed and monitored twice daily, and were euthanized if they developed any signs of morbidity according to a multi-point scoring system based on clinical observation (Supplemental Figure S1). Mice showing signs of distress such as, but not limited to, weight loss, ruffled fur, excessive huddling, lethargy, shivering and labored breathing were euthanized immediately to minimize pain and suffering, as were mice exhibiting any condition clearly indicating that the animal would not survive. Animals surviving 30 days were also euthanized. All mice were humanely euthanized in an inhalation chamber via inhalation of 100% gaseous CO_2_ from a compressed gas tank introduced at a rate resulting in 30–70% displacement of the air per minute in the chamber. This procedure resulted in rapid induction of unconsciousness (2–3 min) with minimal distress, followed rapidly by cessation of respiration. Mice were maintained under 100% CO_2_ for an additional two minutes prior to cervical dislocation.
In order to assess virulence in a standard mouse tail vein model of invasive candidiasis, the C. albicans mutant strains and controls were grown to mid-log phase in YPD at 30 °C. Yeast-phase cells from these strains–parental strain AHY490 from A. Hernday [22], end3 null mutant (CREnd3KO), and END3 re-integrant (CREnd3KI) from our lab [21]–were harvested, washed, counted, and re-suspended in sterile 0.9% (w/v) NaCl. Groups of up to 10 BALB/c male or female mice (Charles River Laboratories, Boston, MA, USA) were injected intravenously with 0.2 mL of each cell suspension at 1.0 × 10^6^ cells per animal, and survival over 28 days was assessed.
We conducted a two-proportion power analysis to estimate the required size of group samples as being 10, achieving 82% power to detect a difference of 0.60000 between the null hypothesis that both group proportions were 0.95000 and the alternative hypothesis that the proportion in group 2 was 0.35000, using a one-sided test with a significance level of 0.05000. We chose to initially over-represent each test condition by using 12 mice in each of the WT-, KO-, and KI-infected groups, in order to account for any failed injections via the lateral tail vein. We confirmed successful intravenous inoculation at time of injection by using established visual and tactile criteria, specifically, visualization of flashback and vein clearing (blanching of the semi-transparent vein during injection), indicating intravascular placement, absence of resistance consistent with intraluminal delivery, and lack of subcutaneous bleb formation. The tail was examined immediately for evidence of extravasation or leakage. Any mouse not successfully inoculated was excluded from the study, resulting in rejection of 3, 2, and 2 mice in each group, respectively.
2.3. Statistical Analyses
Kaplan–Meier survival curves were analyzed using the Mantel–Cox test (GraphPad Prism v. 6.0). Data were assessed visually to ensure the test’s assumption of proportional hazards was true. A result was considered statistically significant when p < 0.05 compared to other survival curves. Initial weights of mice were analyzed using Student’s t test (GraphPad Prism v. 6.0), with significant differences indicated by p < 0.01.
3. Results
The end3 Null Mutant Attenuated Virulence in a Murine Model of Infection
We first assayed the ability of the end3 null mutant, compared to the wild-type control strain, to infect mice in a tail vein model of disseminated candidiasis as a pilot experiment, comparing male and female mice aged approximately 12 weeks (Figure 1). The mice were infected with either the wild-type strain or the null mutant. The wild-type strain caused significant mortality, with six of seven female mice succumbing to infection over 21 days (median survival 3 days) while five out of the eight male mice did not survive (median survival 15 days). This slight difference in survival between wild-type strain-infected female and male mice–with mortality rates of 85.7% compared to 62.5%—was not statistically significant (Mantel–Cox test, p = 0.08). In contrast, all mice infected with the end3 null mutant survived the infection, as none of the seven females or five males died (Mantel–Cox test, p = 0.0002). In this pilot experiment, conducted in a single trial, we did not test the re-integrant strain. The positive control was the parental strain AHY490. A saline-only negative control was not used in the initial pilot study to minimize the number of animals used.
There was a difference in median survival time between male and female mice. However, the initial weights of the male and female mice were also significantly different (Supplemental Figure S2). The male mice were substantially heavier, as determined by a Student’s t test (p < 0.0001). In contrast, there was no significant difference in starting weights between WT- and KO-infected mice of each sex. The median weights of WT- and KO- infected male mice were 27.8 g and 27.75 g, respectively. For the WT- and KO-infected female mice, the median starting weights were 21.5 g and 21.2 g, respectively.
Based on these initial results, we performed another experiment in a single trial to compare the end3 mutant, wild-type (parental), and END3 re-integrant strains using only female mice (Figure 2) aged approximately 10 weeks. Female mice infected with the wild-type strain, as expected, were vulnerable to infection, with seven of nine mice dying over 28 days (median survival 12 days). The group of 10 mice infected with the end3 mutant again exhibited no mortality (Mantel–Cox test, p < 0.0001). Infection with the re-integrant strain resulted in eight of ten mice dying (median survival 7 days). The slight difference in survival between the wild-type strain and re-integrant strain was not statistically significant (Mantel–Cox test, p = 0.92). All four untreated control mice survived.
Additionally, in a post hoc analysis, we combined data from all female mice in both experiments infected with the wild-type strain and compared them to the wild-type-strain-infected male cohort from the first experiment. There was no significant difference in survival outcomes (Mantel–Cox test, p = 0.33), a finding which supports our rationale for utilizing a single sex of mice in our main experiment to analyze virulence (Figure 2). In all cases, the difference in survival between the end3 null mutant and both the wild-type and re-integrant strains was clinically and statistically significant, as none of the end3 mutant-infected mice died. We observed that both groups of CREND3KO-infected female mice across the two experiments survived completely, along with all CREND3KO-infected male mice in the pilot experiment. These findings demonstrate that the endocytosis gene END3 is required for wild-type virulence in a tail vein model of disseminated candidiasis.
4. Discussion
In prior studies, we demonstrated that the C. albicans pre-vacuolar secretory pathway mediated by the vacuolar protein sorting gene VPS4 is required for secretion of the extracellular proteases Sap2 and Saps4–6 in vitro, and that mutants lacking VPS4 are dramatically reduced in virulence in a mouse tail vein model of disseminated infection [5]. Interestingly, however, we found that C. albicans VPS4 is dispensable for virulence in a mouse vaginal model of candidiasis, thus revealing a differential role for the pre-vacuolar secretory pathway in virulence, depending on the infected tissue [23]. We also demonstrated that PEP12, which encodes a t-SNARE involved in pre-vacuolar trafficking, is also required for normal biofilm integrity in vitro, and for virulence in a mouse tail vein model in vivo [6].
Because of the intersection of the endocytic pathway with pre-vacuolar and vacuolar trafficking pathways, we subsequently undertook an analysis of the role of endocytosis in virulence-related phenotypes in C. albicans. We focused on the key early endocytic genes ENT2 and END3. ENT2 encodes an epsin-like protein involved in clathrin binding in early endocytosis [24]. In C. albicans, we demonstrated that the ent2 null mutant was defective in endocytosis: unable to form proper hyphae and biofilms, and unable to invade vaginal keratinocytes in vitro [14]. It was subsequently demonstrated that a C. albicans tetO-regulated ent2 null mutant strain, in the presence of doxycycline, was avirulent in a mouse model of systemic candidiasis (via retro-orbital injection), compared to controls or in the absence of doxycycline [25]. This study did not specifically examine organ fungal burden or histopathology. In this work, we further demonstrate a key role of early endocytosis mediated by END3 in virulence, using a similar mouse model of disseminated candidiasis (via tail vein injection).
In our initial pilot study, we sought to first determine if there was a difference in survival between female and male mice. We found that there was no statistically significant difference in survival, thus providing reassurance that there were no major biological differences in survival from disseminated infection with C. albicans in this model. Of note, the male mice of the same age were larger (by weight) than the female mice, which is likely responsible for the longer median survival of the male mice in this experiment. Moreover, in a large meta-analysis of the clinical literature, sex was not identified as a risk for development of invasive candidiasis in critically ill patients [26]. Taking all these findings together, we believe that use of a single sex of mice is acceptable for conducting virulence studies such as these, and that performance of such experiments using separate male and female mice is unnecessary, particularly in light of the need to limit the overall number of mice used in these experiments in light of ethical considerations.
We found a complete, consistent absence of mortality across all groups of end3 KO-infected mice, which included both groups of KO-infected female mice, whereas most WT-infected and KI-infected mice did not survive. Collectively, this and preceding studies indicate that early endocytosis in C. albicans plays a central role in key virulence-related processes including filamentation and biofilm formation. Additional molecular studies of the role of endocytosis in C. albicans pathogenesis are warranted, in order to better understand how this common and dangerous opportunistic pathogen causes morbidity and mortality.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Rollenhagen C. Mamtani S. Ma D. Dixit R. Eszterhas S. Lee S.A. The Role of Secretory Pathways in Candida albicans Pathogenesis J. Fungi 202062610.3390/jof 6010026 PMC 715105832102426 · doi ↗ · pubmed ↗
- 2Schaller M. Korting H.C. Schäfer W. Bastert J. Chen W. Hube B. Secreted aspartic proteinase (Sap) activity contributes to tissue damage in a model of human oral candidosis Mol. Microbiol.19993416918010.1046/j.1365-2958.1999.01590.x 10540295 · doi ↗ · pubmed ↗
- 3Tsui C. Kong E.F. Jabra-Rizk M.A. Pathogenesis of Candida albicans biofilm Pathog. Dis.201674 ftw 01810.1093/femspd/ftw 01826960943 PMC 5975230 · doi ↗ · pubmed ↗
- 4Lee S.A. Jones J. Hardison S. Kot J. Khalique Z. Bernardo S.M. Lazzell A. Monteagudo C. Lopez-Ribot J. Candida albicans VPS 4 is Required for Secretion of Aspartyl Proteases and In Vivo Virulence Mycopathologia 2009167556310.1007/s 11046-008-9155-718814053 PMC 5898969 · doi ↗ · pubmed ↗
- 5Palanisamy S.K. Ramirez M.A. Lorenz M. Lee S.A. Candida albicans PEP 12 Is Required for Biofilm Integrity and In Vivo Virulence Eukaryot. Cell 2010926627710.1128/EC.00295-0920023068 PMC 2823007 · doi ↗ · pubmed ↗
- 6Lv Q. Yan L. Jiang Y. The Importance of Vacuolar Ion Homeostasis and Trafficking in Hyphal Development and Virulence in Candida albicans Front. Microbiol.20211277917610.3389/fmicb.2021.77917634956142 PMC 8696117 · doi ↗ · pubmed ↗
- 7Bernardo S.M. Rane H.S. Chavez-Dozal A. Lee S.A. Secretion and filamentation are mediated by the Candida albicans t-SNAR Es Sso 2p and Sec 9p FEMS Yeast Res.20141476277510.1111/1567-1364.1216524911595 PMC 4126854 · doi ↗ · pubmed ↗
- 8Martin R. Hellwig D. Schaub Y. Bauer J. Walther A. Wendland J. Functional analysis of Candida albicans genes whose Saccharomyces cerevisiae homologues are involved in endocytosis Yeast 20072451152210.1002/yea.148917431925 · doi ↗ · pubmed ↗