Nematicidal Potential of Purpureocillium takamizusanense PMEPF27 Against Motile Bursaphelenchus rainulfi In Vitro
Yuh Tzean, Elena Gamboa Chen, Xiao-Yu Wei, I-En Shih, Hui-Yu Hsu, Ya-Zhen Xu, Ying-Hong Lin, Meng-Ling Wu, Tai-Yuan Chen, Yung-Yu Yang, Jen-Chih Chen

TL;DR
This study shows that the fungus Purpureocillium takamizusanense PMEPF27 can effectively kill the motile nematode Bursaphelenchus rainulfi in lab conditions.
Contribution
The study expands the known biocontrol capabilities of P. takamizusanense to include motile nematode stages.
Findings
PMEPF27 caused ~80% nematode mortality within 7 days in vitro.
The fungus caused surface disruption of nematode cuticles.
B. rainulfi is validated as a suitable motile species for bioassays involving pine nematodes.
Abstract
Bursaphelenchus rainulfi is a pine-associated, non-pathogenic nematode that serves as a motile comparative species for evaluating nematophagous fungi. We investigated the in vitro biocontrol activity of Purpureocillium takamizusanense strain PMEPF27, originally isolated from insect cadavers in Taiwan, against mixed motile stages of B. rainulfi. Identity of the fungus was confirmed by morphology and ITS/EF-1α sequencing. Nematodes were exposed in liquid suspension to PMEPF27 conidia, with sterile water as the negative control and fluopyram as the positive control. Mortality was monitored over 7 days, and scanning electron microscopy was used to observe fungus–nematode interactions. PMEPF27 caused significantly higher mortality than the water control, reaching ~80% by day 7, and showed surface disruption of nematode cuticles, although no direct spore penetration was observed. These…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
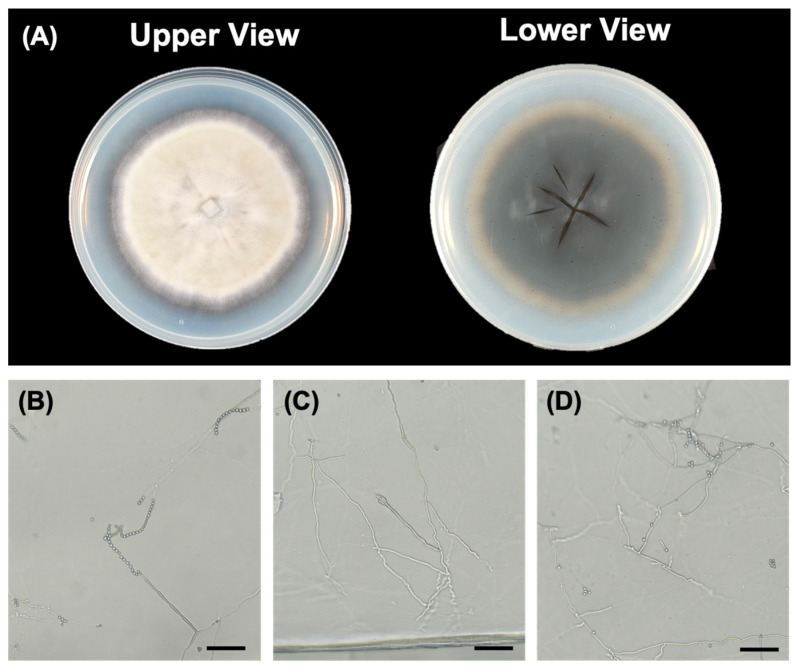 Figure 1
Figure 1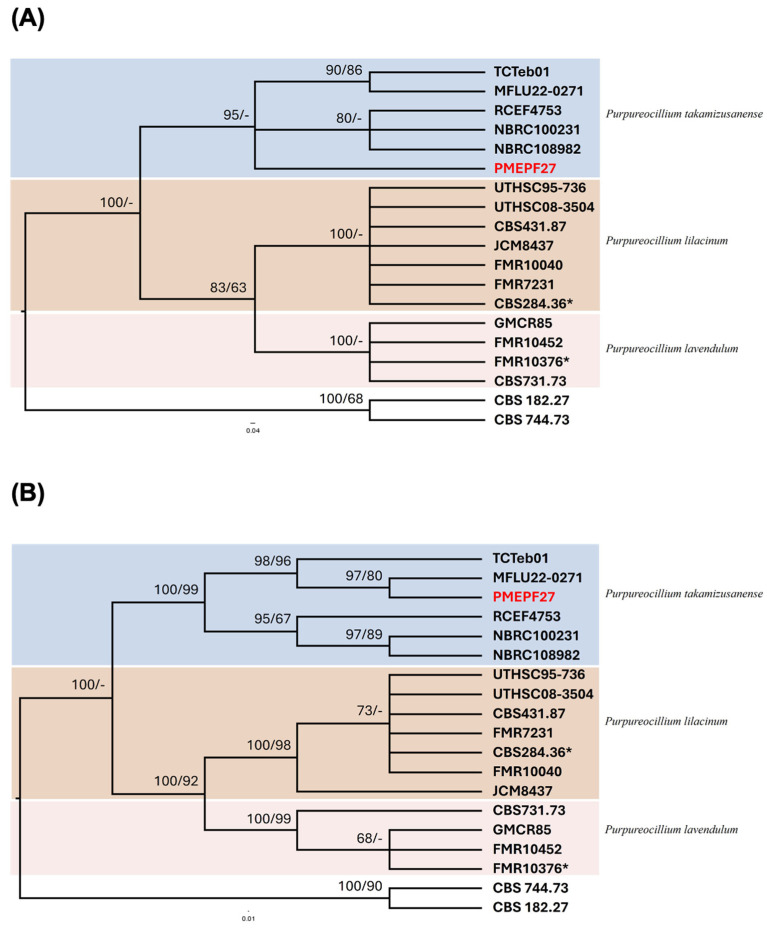 Figure 2
Figure 2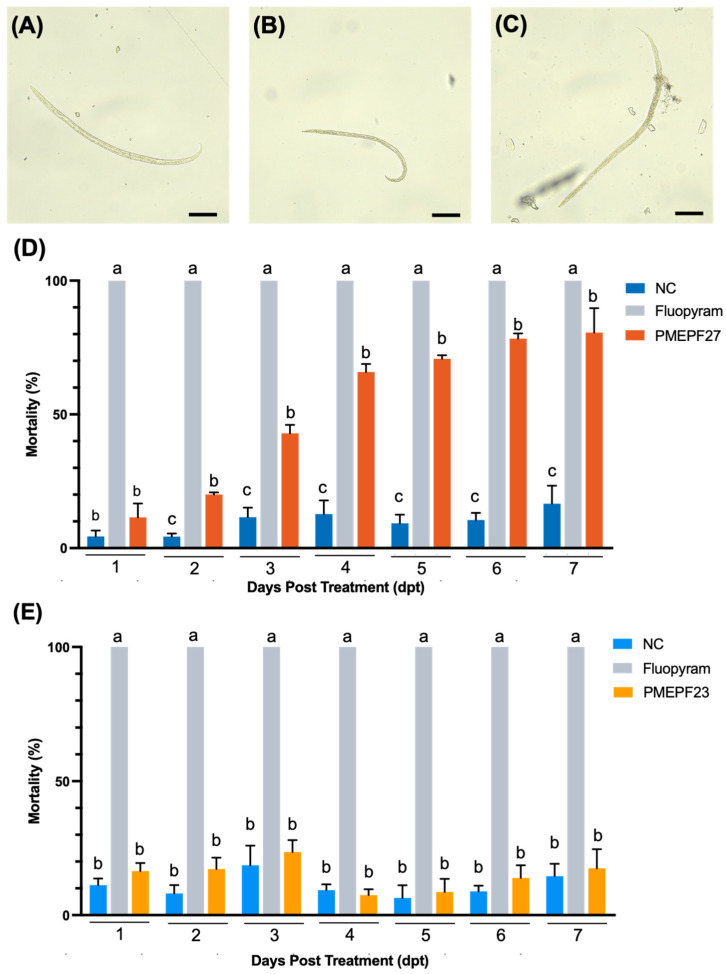 Figure 3
Figure 3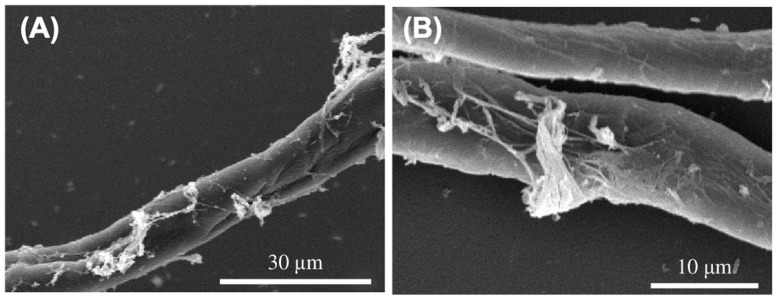 Figure 4
Figure 4- —Ministry of Agriculture, Executive Yuan, Taiwan
- —National Taiwan University, Taiwan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNematode management and characterization studies · Entomopathogenic Microorganisms in Pest Control · Plant Pathogens and Fungal Diseases
1. Introduction
Plant-parasitic nematodes (PPNs) are among the most destructive pests in agriculture and forestry, causing an estimated 12–13% reduction in global crop yields annually [1,2]. The pine wood nematode (PWN), Bursaphelenchus xylophilus, is particularly notorious as the causal agent of pine wilt disease (PWD) and is considered one of the most damaging forest pathogens worldwide [3]. Initially believed to be native to North America, the devastating impact of PWN was first documented in Japan during the early 20th century [4,5], and the nematode later made its way to Europe, first impacting Portugal and later spreading to Spain [6]. Because B. xylophilus is a quarantine organism in many countries, work with this pathogen is restricted and requires strict phytosanitary measures [7].
Current control measures for PWN and other PPNs rely heavily on chemical nematicides and phytosanitary interventions, like fumigation or burning of infested wood [8,9,10,11]. However, these approaches raise environmental and safety concerns, and their effectiveness is limited by cost, labor, and the potential for nematicide resistance [12]. There is an urgent need for sustainable, ecologically friendly nematode management strategies [13]. Consequently, there is a strong need for alternative, sustainable approaches and for laboratory models that allow mechanistic study of nematode–microbe interactions without the regulatory burden of working directly with quarantine nematodes [14].
Biological control has emerged as a promising pillar of integrated pest management (IPM), utilizing the natural antagonism between microorganisms and pests to achieve suppression [15]. Among the various microorganisms explored, nematophagous fungi have received considerable attention due to their diverse mechanisms of action, which include parasitism and the secretion of cuticle-degrading enzymes [16,17]. The genus Purpureocillium, formerly categorized under Paecilomyces, contains several species recognized for their robust nematophagous properties [18,19]. Purpureocillium lilacinum is perhaps the most well-known member, having been extensively commercialized for its ability to parasitize the eggs and sedentary stages of various nematodes [20,21]. The mechanisms of nematode inhibition by P. lilacinum include the production of cuticle-degrading enzymes such as proteases, chitinases, and lipases that facilitate hyphal penetration of nematode eggs and cuticles [22,23]. Additionally, P. lilacinum produces secondary metabolites with nematicidal and egg-hatching inhibitory effects [24,25].
In contrast, P. takamizusanense has been much less studied. First described as the anamorph of a Cordyceps teleomorph (Isaria takamizusanensis), it is recognized as an emerging entomopathogenic fungus in Asia [26,27]. Recent reports indicate that P. takamizusanense can infect diverse insect hosts and may exhibit plant growth-promoting traits, such as phosphate solubilization [26]. The genome of P. takamizusanense has been sequenced, revealing 11,855 protein-coding genes and 36 biosynthetic gene clusters that may contribute to bioinsecticidal activity [28]. Whether this species also possesses nematophagous capabilities remains largely unexplored.
Previous research on Purpureocillium biocontrol has largely concentrated on nematode eggs or sedentary stages, whereas comparatively less is known about activity toward motile vermiform stages. Evaluating pathogenicity against actively moving nematodes is essential for assessing the true biocontrol potential of these fungi. In the present study, B. rainulfi was selected as a model organism to examine the nematicidal capacity of P. takamizusanense strain PMEPF27. B. rainulfi is a pine-associated species originally described from decaying pine wood in Malaysia and subsequently reported in China and Taiwan [29,30]. Pathogenicity assays have demonstrated that it does not induce pine wilt disease [30], yet it co-occurs with other pine-associated nematodes and can serve as a reliable surrogate in laboratory bioassays. Furthermore, interactions of B. rainulfi with nematophagous fungi, including susceptibility to Esteya vermicola infection in vitro [31], reinforce its suitability as a model for fungus–nematode studies.
Here, we use B. rainulfi as a non-pathogenic motile model to investigate the nematophagous potential of P. takamizusanense PMEPF27 under controlled in vitro conditions. The objectives of this study were to (i) confirm the identity of strain PMEPF27 using morphological and molecular approaches, (ii) quantify its effect on motile B. rainulfi survival over time in comparison with a chemical nematicide control, and (iii) characterize fungus–nematode surface interactions by scanning electron microscopy (SEM) to generate hypotheses about possible killing mechanisms. To contextualize whether the observed activity reflects strain-specific or broader EPF-associated effects, we additionally included Beauveria bassiana PMEPF23 as a taxonomically distinct entomopathogenic fungal comparative control. By combining a non-pathogenic pine-associated nematode with an entomopathogenic fungus that shows nematicidal activity, we establish a tractable model system for dissecting fungal factors involved in nematode killing without the need to manipulate quarantine nematodes in the laboratory.
This study establishes a safe and tractable experimental model for investigating fungus–nematode interactions using a non-pathogenic, pine-associated motile nematode under controlled in vitro conditions. By demonstrating that P. takamizusanense can increase the mortality of motile nematodes, our findings expand the currently limited understanding of nematode-targeting activity in this species. These results contribute to a deeper understanding of fungal-nematode interactions and may inform the development of sustainable, biologically based strategies for nematode management.
2. Materials and Methods
2.1. Preparation of Bursaphelenchus rainulfi
Bursaphelenchus rainulfi was obtained from Dr. Pei-Chen Chen (Department of Plant Pathology, National Chung Hsing University, Taiwan). The nematodes were maintained on Alternaria sp. colonies cultured on potato dextrose agar (PDA), as described for this species [30]. Alternaria sp. colonies were cut using a circular puncher (diameter 1 cm) and subcultured on PDA, then incubated at 28 °C in the dark for 7 days. After 7 days, B. rainulfi were transferred onto Alternaria sp. colonies and cultured in the dark at 28 °C for 14 days. The B. rainulfi culture medium was washed with 3 mL of sterile water to prepare a nematode suspension. Each biological replicate in the experimental set contained approximately 50 nematodes.
2.2. Cultivation of Entomopathogenic Fungi (EPF)
The entomopathogenic fungal (EPF) strains used in the experiments were PMEPF27 (Purpureocillium takamizusanense) (PMEPF27) and PMEPF23 (Beauveria bassiana). PMEPF23 was isolated from a lepidopteran larva collected in Heping District, Taichung City, Taiwan. PMEPF27 was isolated from Tessaratoma papillosa collected in Laopi Village, Neipu Township, Pingtung County, Taiwan.
For conidial production, each EPF strain was cultured on PDA at 28 °C in the dark for 14 days. After incubation, the EPF culture medium was washed with 3 mL of sterile water, and a triangular glass rod was used to prepare spore suspensions. Spore concentrations were determined using a hemocytometer and adjusted to 1 × 10^6^ conidia mL^−1^ for bioassays.
2.3. Morphological Observation of PMEPF27
For microscopy observation of sporulation structures of PMEPF27, the fungal strain was cultured on water agar (WA) at 28 °C in the dark for 14 days. A circular piece (diameter 1 cm) of the mycelial medium was cut and placed on a slide, and the structures were observed using a microscope (Zeiss Axio Scope, Jena, Germany). Colony characteristics were documented on PDA after 7 days of incubation at 28 °C, including colony morphology, pigmentation, texture, and radial growth rate.
2.4. Molecular Identification of Fungal Isolate
PMEPF27 and PMEPF23 colonies were cultured on WA medium for 14 days. The mycelium was ground to a powder with liquid nitrogen, and 100 mg of the powdered sample was transferred to a 1.5 mL microcentrifuge tube. Fungal genomic DNA was isolated using the Plant DNA Mini Kit (LabPrep, Taipei, Taiwan) according to the manufacturer’s instructions. Molecular identification of fungi was conducted using ITS and EF-1α gene primers (Supplementary Table S1). The ITS region was amplified using primers ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) [32]. The EF-1α gene was amplified using primers EF1-983F (5′-GCYCCYGGHCAYCGTGAYTTYAT-3′) and EF1-2218R (5′-ATGACACCRACRGCRACRGTYTG-3′) [33]. PCR was performed in 25 μL reactions containing 1× PCR buffer, 2.5 mM MgCl_2_, 0.2 mM dNTPs, 0.4 μM of each primer, 1 U Taq polymerase, and approximately 50 ng of template DNA. PCR cycling conditions were: initial denaturation at 95 °C for 5 min; 35 cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 1 min; and final extension at 72 °C for 10 min. PCR products were purified using the GEL/PCR Purification mini Kit (FavorPrep™, Ping-Tung, Taiwan) and sequenced by Tri-I Biotech Inc., New Taipei City, Taiwan. Sequences were deposited in GenBank (PMEFP27 ITS: PZ092910; EF-1α: PZ111818).
2.5. Phylogenetic Analyses
Phylogenetic analyses of PMEPF27 were performed using the ITS and EF-1α sequences, along with reference strains, using Bayesian inference and maximum likelihood approaches. The accession numbers for the reference stains are listed in Supplementary Table S2. MrBayes inference analysis was conducted using MrBayes V3.2.7 [34,35,36]. Markov Chain Monte Carlo (MCMC) sampling was employed for 1,000,000 generations with lset nst = 6, while other parameters remained unchanged. Evolutionary analysis was performed using the Maximum Likelihood method and Tamura-Nei model [37]. The evolutionary history was inferred utilizing MEGA X [38].
2.6. In Vitro Nematode Mortality Assay of Entomopathogenic Fungi Against Bursaphelenchus rainulfi
A 6 cm plastic culture dish containing approximately 50 individual B. rainulfi in 3 mL sterile water was mixed with 3 mL of the corresponding conidial suspension (PMEPF27 or PMEPF23) adjusted to 1 × 10^6^ conidia mL^−1^, resulting in a final concentration of 5 × 10^5^ conidia mL^−1^. The culture dishes were tightly sealed with Parafilm to prevent evaporation and observed under a dissecting microscope at 24 h intervals for 7 days. The number of motile (living) and immotile (dead) nematodes was recorded. Nematodes were considered dead when they showed no movement after gentle agitation or mechanical stimulation with a fine probe. The 7-day observation period was selected to capture delayed mortality effects and to characterize the full time course of fungal activity, which can develop over multiple days.
In the negative control group, the spore suspension of either PMEPF27 or PMEPF23 was replaced with sterile water. In the positive control group, fluopyram (Velum^®^ Prime, Bayer CropScience, St. Louis, MO, USA), a succinate dehydrogenase inhibitor nematicide, was used. A 1500-fold dilution of fluopyram was mixed with an equal volume of nematode suspension, resulting in the manufacturer’s recommended concentration for nematode control (approximately 0.5 mg L^−1^ active ingredient). Each treatment group consisted of three biological replicates (n = 3). The experiment was repeated twice to confirm reproducibility (experimental sets 1 and 2). For each time point, mortality data were analyzed by one-way ANOVA followed by Tukey’s multiple-comparison test to evaluate differences among treatments (negative control, fluopyram, and fungal treatment). Statistical significance was defined at p < 0.05.
2.7. Scanning Electron Microscopy
The fungal-infected B. rainulfi were first prepared as described previously for the in vitro assay. Nematodes were collected by centrifugation (300 rpm, 3 min, 25 °C) and deposited at the bottom, which were then transferred onto the glass coverslips using 200 µL and 1000 µL pipettes. The samples were fixed in glutaraldehyde and stored at 4 °C for 1–2 h. Sample preparation for SEM, including dehydration and critical point drying, was conducted similarly according to the standard biological specimen protocols described previously [39]. Following incubation, samples were washed twice with PBS buffer for 15 min each. A graded series of alcohol concentrations (30%, 50%, 70%, 80%, 90%) was used for dehydration, 10 min each step, followed by two 15 min treatments with 100% alcohol, and then two 15 min treatments with acetone. Samples were then dried using a critical point dryer (CPD) (73 atm, 1 h 10 min, 37 °C). After drying, the coverslips were attached to metal stubs coated with carbon tape using forceps and sputter-coated using an ion sputter coater (7 atm, 180 s, 30–40 °C). Finally, the surface morphology of the nematodes and the fungal parasitism were examined using a scanning electron microscope (Hitachi, Tokyo, Japan).
3. Results
3.1. Growth Characteristics and Morphology
Strain PMEPF27 grew readily on PDA, exhibiting a dense, velvety texture with a radially folded surface and white to purplish pigmentation at the colony edge (Figure 1A). The radial growth reached 1.81 ± 0.28 cm after 7 days of incubation at 28 °C. The reverse side of colonies showed cream to pale purple coloration.
Microscopic examination of PMEPF27 showed smooth-walled, single-celled conidia that were elliptical to cylindrical in shape, and arranged in long chains (Figure 1B–D). Conidia measured 2.5–4.0 μm in length and 1.5–2.5 μm in width. Hyphae were septate and branched. Conidiophores were erect and often formed clusters with a series of phialides (Figure 1B–D). These morphological characteristics were consistent with the description of Purpureocillium species [40,41].
3.2. Molecular Characterization of PMEPF27
To confirm the taxonomic identity of PMEPF27, the internal transcribed spacer (ITS) region and elongation factor-1α (EF-1α) gene regions were amplified and sequenced. The ITS sequence was 532 bp (GenBank: PZ092910) and the EF-1α sequence was 936 bp (GenBank: PZ11818). BLAST 2.17.0 analysis showed high sequence similarity of PMEFP27 to P. takamizusanense reference strains for both ITS and EF-1α (Supplementary Table S2).
Phylogenetic analysis using Bayesian inference (MrBayes) and maximum likelihood (ML) methods showed three main distinct clades comprising P. takamizusanense, P. lilacinum, and P. lavendulum (Figure 2). PMEPF27 clustered within the clade comprising P. takamizusanense with high statistical support in both trees. In the ITS-based tree, PMEPF27 grouped with P. takamizusanense reference strains with 100% Bayesian posterior probability and 98% ML bootstrap support (Figure 2A). Similarly, in the EF-1α-based tree, PMEPF27 formed a well-supported clade with P. takamizusanense strains, showing 100% Bayesian posterior probability and 97% ML bootstrap support (Figure 2B). These results identified PMEPF27 as P. takamizusanense.
3.3. Pathogenicity of PMEPF27 Against Bursaphelenchus rainulfi
Mortality of motile B. rainulfi was quantified following exposure to conidial suspensions of PMEPF27 (P. takamizusanense) (Figure 3A–D). To contextualize whether the observed mortality patterns were specific to PMEPF27 or could also be observed with another EPF taxon under the same assay format, PMEPF23 (B. bassiana) was evaluated in a parallel assay as a taxonomically distinct entomopathogenic fungal control (Figure 4). In each assay, sterile water served as the negative control (NC) and fluopyram served as the chemical positive control. Fluopyram resulted in 100% mortality by 1 day post-treatment (dpt), the first observation time point, whereas mortality in NC remained low throughout the observation period (Figure 3D and Figure 4).
PMEPF27 exposure produced a clear time-dependent increase in mortality relative to NC (Figure 3D). Mortality increased progressively over the observation period and reached high levels at later time points (5–7 dpt). The independent repeat (Experimental Set 2) showed a similar temporal pattern (Supplementary Figure S1A), supporting reproducibility. One-way ANOVA followed by Tukey’s multiple-comparison test indicated significantly higher mortality in the PMEPF27 treatment than in NC at multiple time points (p < 0.05; Figure 3D).
In the PMEPF23 assay, mortality remained comparatively low over time (Figure 4). Although immotile nematodes were observed in the PMEPF23 treatment, the overall mortality profile did not show a sustained increase relative to NC under the multiple-comparison analysis. The independent repeat (Experimental Set 2) is provided in Supplementary Figure S1B. Together, these results indicate that EPF strains differed in the magnitude and temporal dynamics of mortality against motile B. rainulfi under the assay conditions, with PMEPF27 showing the stronger time-dependent effect.
3.4. Pathogenicity of PMEPF27 Against Bursaphelenchus rainulfi as Revealed by Scanning Electron Microscopy (SEM)
To further analyze the interaction of PMEPF27 and B. rainulfi, scanning electron microscopy (SEM) was used to observed B. rainulfi that was exposed to PMEPF27. Based on the observation under SEM, by 72 h post-exposure, marked degradation of the nematode cuticle was observed (Figure 4). Although several nematodes displayed eroded, disrupted, and breached cuticles, no PMEPF27 spores were detected adhering to the nematodes at the examined time point.
4. Discussion
This study demonstrates the strong in vitro biocontrol activity of P. takamizusanense PMEPF27 against motile B. rainulfi, a pine-associated nematode that serves as a surrogate species for laboratory evaluation of fungal effects relevant to pinewood nematode systems. Previous research on this fungus has focused predominantly on its entomopathogenic properties and potential plant growth-promoting capabilities [26,27,42]. In Taiwan, P. takamizusanense was first documented as an entomopathogenic fungus infecting Tessaratoma papillosa adults [41], and the species has been reported to exhibit activity against agricultural pests from multiple insect orders [26]. Our findings expand the currently recognized functional scope of P. takamizusanense and motivate further examination of the interactions with nematodes.
P. lilacinum has been extensively studied for nematode control over past decades [22,23,43,44,45,46]. A number of studies emphasize egg parasitism where fungi penetrate egg shells or cuticles and then consume the nematode contents [47]. The fungus produces an array of hydrolytic enzymes, including serine proteases, subtilisins, chitinases, and lipases, that degrade the chitin-rich egg shells and protein-rich cuticles of nematodes [22,23,48,49]. Whole-genome analysis has confirmed that P. lilacinum possesses large sets of these hydrolytic enzymes, with serine proteases specifically up-regulated in the presence of nematode eggs [19,28]. For example, overexpression of a serine protease gene in P. lilacinum significantly enhanced virulence against Meloidogyne incognita eggs [50]. In the case of motile nematodes, P. lilacinum can also infect second-stage juveniles (J2) by enzymatic digestion of the cuticle [51,52], though many studies have concentrated on egg infection since eggs are stationary targets.
In our assay, PMEPF27 caused a time-dependent increase in mortality of motile nematodes over 5–7 days, whereas fluopyram produced complete mortality by the first observation time point. Although PMEPF27 acted more slowly than the chemical control, it produced substantial mortality by later time points. The time course observed here is consistent with report for other Purpureocillium spp. affecting motile juveniles over multi-day intervals, including P. lavendulum against M. incognita J2 [53], and P. lilacinum, which typically kills Meloidogyne eggs and females within 5–7 days [19,49,54]. While chemicals like fluopyram offer rapid nematicidal effect, biological agents like P. lilacinum are increasingly recommended for sequential application with chemical nematicides to provide sustained control and improved yields [54].
In addition, culture-derived compounds from P. lavendulum have been reported to exhibit nematicidal or egg-hatching inhibitory effects [53,55]. Recent chemo-profiling has identified specific metabolites responsible for this activity, including 5-methoxymethyl-1H-pyrrole-2-carboxaldehyde from P. lavendulum [53], as well as hydroxylated fatty acids and steroids [55]. In P. lilacinum, organic acids, phenolic compounds, and fatty acids such as myristic and lauric acid have been correlated with high J2 mortality [24,25]. Together, these findings support the view that Purpureocillium spp. may affect nematodes through a combination of processes, although the relative contributions are likely strain- and context dependent.
The SEM observations documented cuticle damage in PMEPF27-treated nematodes; however, we did not conidia directly attached to nematode surfaces or appressoria-like penetration structures at the examined time points. These observations are compatible with, but do not demonstrate, a role extracellular enzymes and metabolites. Secondary metabolite diversity and secreted enzymes, including leucinostatins, have been documented in Purpureocillium spp. and are linked to nematode antagonisms in other systems [18,22,23,24,25,56]. Genomic resources available for P. takamizusanense also suggest the potential for substantial repertoire of secreted proteins and secondary-metabolite biosynthetic gene clusters [28]. Accordingly, an important next step will be to couple time-resolved microscopy with assays of extracellular enzyme activities and culture-filtrate bioactivity, followed by chemical profiling and gene-level interrogation, to identify which factors are responsible for PMEPF27-induced mortality.
Our study also supports B. rainulfi as a potential comparative model organism for evaluating fungal effects on pine-associated nematodes. B. rainulfi is phylogenetically related to the pathogenic B. xylophilus but does not cause pine wilt disease [30], making it a safer alternative for laboratory studies that do not require quarantine facilities. In Taiwan, pathogenicity assays on Pinus thunbergii indicate that B. rainulfi did not induce wilting symptoms under tested conditions [30]. The species has been reported from pine wood in Malaysia, China, and Taiwan and co-occurs with other Bursaphelenchus spp. in pine ecosystems [29,30,57]. Previous studies also showed that B. rainulfi is susceptible to infection by the nematophagous fungus Esteya vermicola [31], demonstrating its utility for fungus–nematode interaction studies. Future studies could directly compare PMEFP27 effects on B. rainulfi and B. xylophilus to PMEPF27 to determine whether the biocontrol efficacy extends to the pathogenic species.
Our comparison of P. takamizusanense PMEPF27 and B. bassiana PMEPF23 revealed marked differences in nematicidal activity. Including PMEPF23 as a taxonomically distinct entomopathogenic fungal control provided context for EPF-associated effects under the same assay format. Under our conditions, PMEPF23 produced comparatively low mortality and did not show a sustained increase relative to the negative control, whereas PMEPF27 produced a stronger time-dependent effect. Although Beauveria species have been reported to exhibit nematicidal activity against B. xylophilus in vitro and B. mucronatus in vivo [58,59], nematode-antagonistic effects appear to be strain- and context-dependent rather than universal within the genus. In contrast, the stronger activity observed for PMEPF27 is consistent with the biology of Purpureocillium species, which commonly inhabit soil environments where nematode interactions are frequent [18,22]. Furthermore, the ability of Purpureocillium isolates to tolerate water stress and varying temperature ranges supports their capacity to function effectively under variable agroecological conditions [12,18]. Together, these findings highlight the importance of standardized assays and empirical screening when selecting fungal candidates for nematode biocontrol.
This study provides evidence that P. takamizusanense PMEPF27 can affect motile pine-associated nematodes in vitro. These findings expand the currently recognized functional breadth of B. rainulfi and establish a tractable platform for future mechanistic studies. Further work should prioritize identification of causal factors underlying mortality, evaluation against B. xylophilus under appropriate containment, and assessment of formulation and delivery strategies relevant to pine systems.
5. Conclusions
P. takamizusanense PMEPF27 increased the mortality of motile B. rainulfi under controlled in vitro conditions and was associated with surface cuticle damage observable by SEM. Molecular and phylogenetic analyses identified PMEPF27 as P. takamizusanense. The use of B. rainulfi as a non-quarantine, pine-associated motile model provides a tractable platform for screening and hypothesis generation relevant to pinewood nematode systems. Future work should determine the causal factors underlying PMEPF27-associated mortality through targeted assays of culture filtrates, extracellular enzyme activities, and secreted metabolites, and should evaluate efficacy against B. xylophilus under appropriate containment.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Singh S. Singh B. Singh A. Nematodes: A threat to sustainability of agriculture Proc. Environ. Sci.20152921521610.1016/j.proenv.2015.07.270 · doi ↗
- 2Decraemer W. Hunt D.J. Structure and classification Plant Nematology CABI Wallingford UK Wallingford, UK 2006332
- 3Pires D. Vicente C.S. Inácio M.L. Mota M. The Potential of Esteya spp. for the Biocontrol of the Pinewood Nematode, Bursaphelenchus xylophilus Microorganisms 20221016810.3390/microorganisms 1001016835056617 PMC 8781088 · doi ↗ · pubmed ↗
- 4Linit M. Nemtaode-vector relationships in the pine wilt disease system J. Nematol.19882022719290206 PMC 2618795 · pubmed ↗
- 5Kikuchi T. Cotton J.A. Dalzell J.J. Hasegawa K. Kanzaki N. Mc Veigh P. Takanashi T. Tsai I.J. Assefa S.A. Cock P.J. Genomic insights into the origin of parasitism in the emerging plant pathogen Bursaphelenchus xylophilus P Lo S Pathog.20117 e 100221910.1371/journal.ppat.100221921909270 PMC 3164644 · doi ↗ · pubmed ↗
- 6Mota M.M. Braasch H. Bravo M.A. Penas A.C. Burgermeister W. Metge K. Sousa E. First report of Bursaphelenchus xylophilus in Portugal and in Europe Nematology 1999172773410.1163/156854199508757 · doi ↗
- 7Dwinell L.D. The pinewood nematode: Regulation and mitigation Ann. Rev. Phytopathol.19973515316610.1146/annurev.phyto.35.1.15315012519 · doi ↗ · pubmed ↗
- 8Takai K. Suzuki T. Kawazu K. Development and preventative effect against pine wilt disease of a novel liquid formulation of emamectin benzoate Pest Manag. Sci.20035936537010.1002/ps.65112639056 · doi ↗ · pubmed ↗