Chemical Composition and Biological Activities of Diverse Products from Commiphora gileadensis: A Comparative Review
Fawaz K. Alanazi, Nashwa Hashad, Asmaa A. Ahmed, Haitham A. Ibrahim, Reham R. Ibrahim, Mohamed I. S. Abdelhady, Eman G. Haggag, Fatma M. Abdel Bar

TL;DR
This paper reviews the chemical makeup and health benefits of Commiphora gileadensis, a traditional medicinal plant, to support its potential use in modern medicine.
Contribution
The study provides a comprehensive and comparative analysis of the phytochemical composition and biological activities of Commiphora gileadensis products.
Findings
The plant contains ten chemical groups, including terpenoids and flavonoids, with essential oil profiles varying by geography and preparation.
Pharmacological studies show antioxidant, anti-inflammatory, antimicrobial, anticancer, antidiabetic, and wound-healing properties.
Toxicological data indicate low toxicity, but clinical and pharmacokinetic studies are lacking.
Abstract
Background/Objectives: Commiphora gileadensis (Balm of Gilead) is an aromatic medicinal plant with a history of traditional use in ancient and Arabic medicine. It has been used traditionally to treat inflammation, infections, and wounds. Despite its long-standing cultural and economic importance, modern pharmacological validation requires a comprehensive synthesis of current scientific data. This review aims to provide a thorough comparative summary of the phytochemical composition and biological activities of its diverse products. Methods: An updated literature search was conducted using databases such as ScienceDirect, PubMed, Scopus, and Google Scholar, covering publications from approximately 2000 to 2025. The review included English-language peer-reviewed articles, books, and reports providing phytochemical analyses or biological evaluations. Data were manually extracted and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1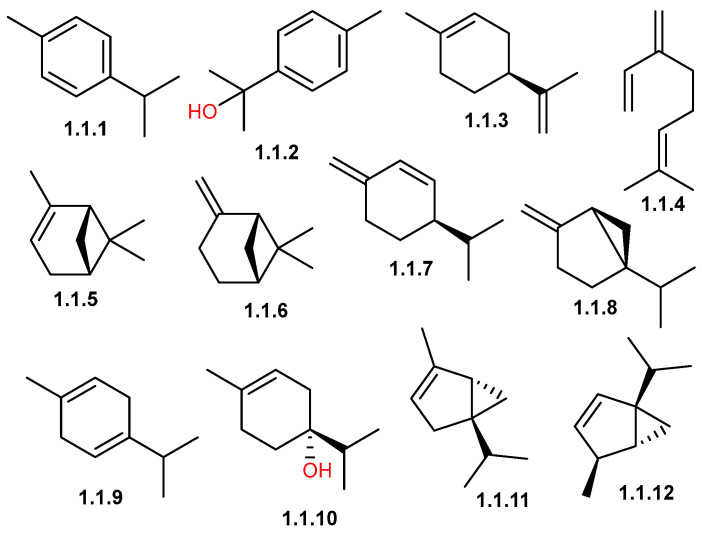 Figure 2
Figure 2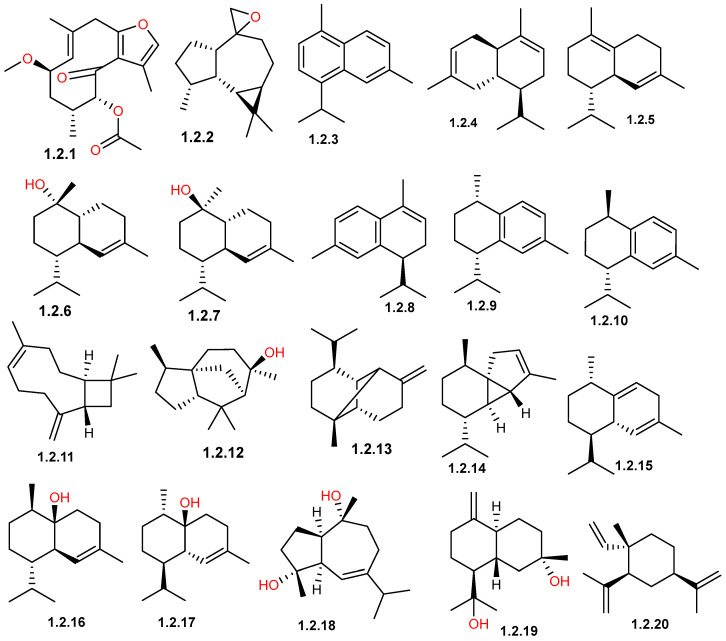 Figure 3
Figure 3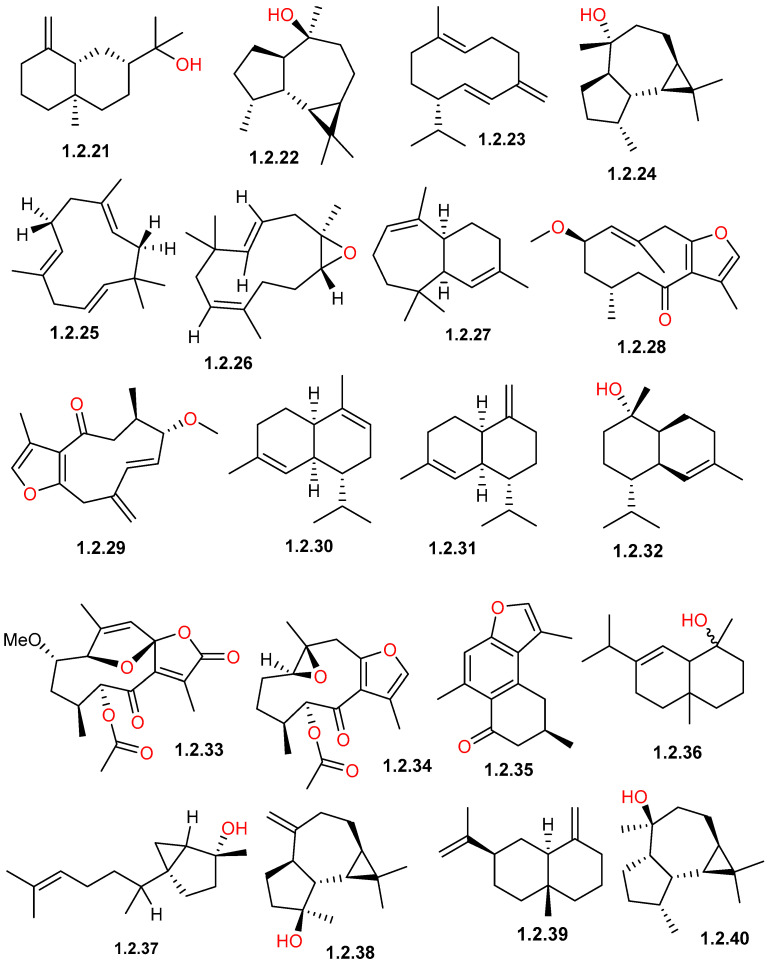 Figure 4
Figure 4 Figure 5
Figure 5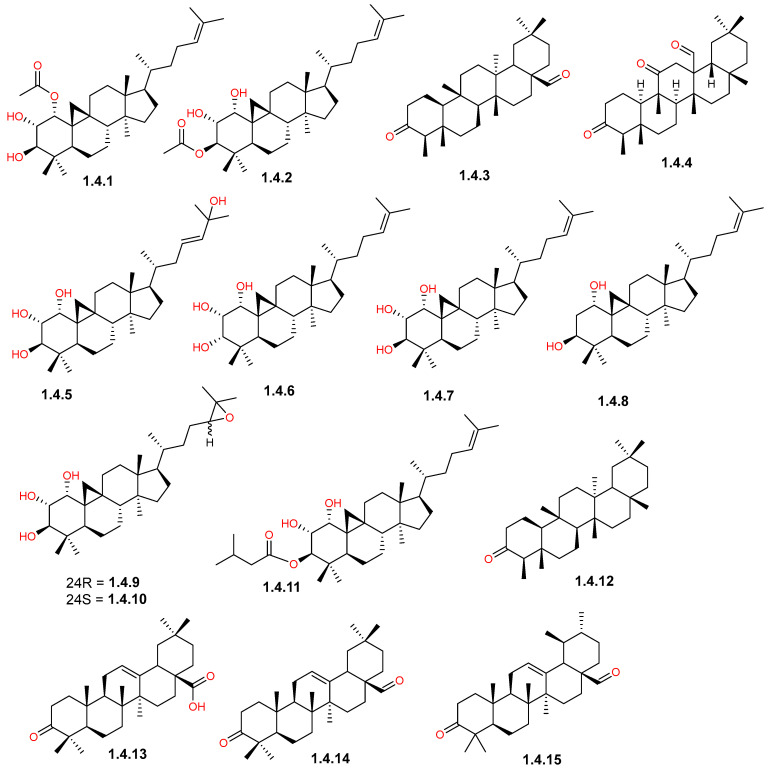 Figure 6
Figure 6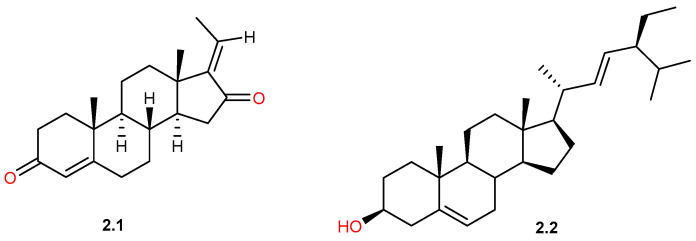 Figure 7
Figure 7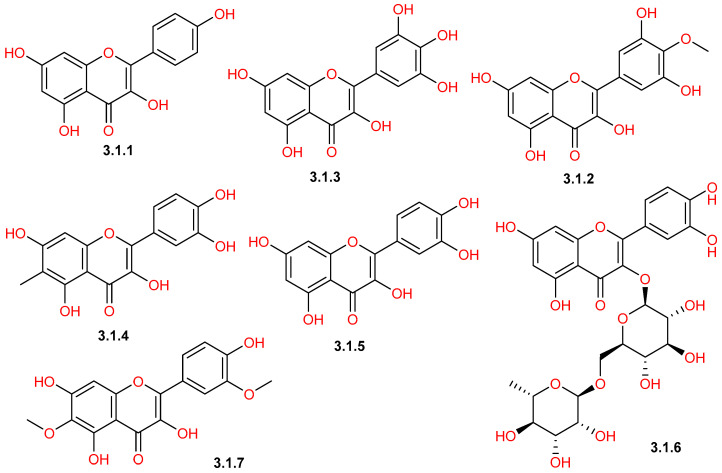 Figure 8
Figure 8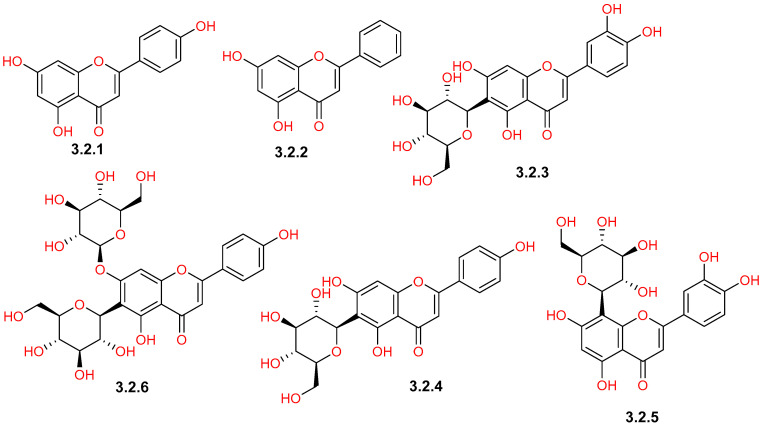 Figure 9
Figure 9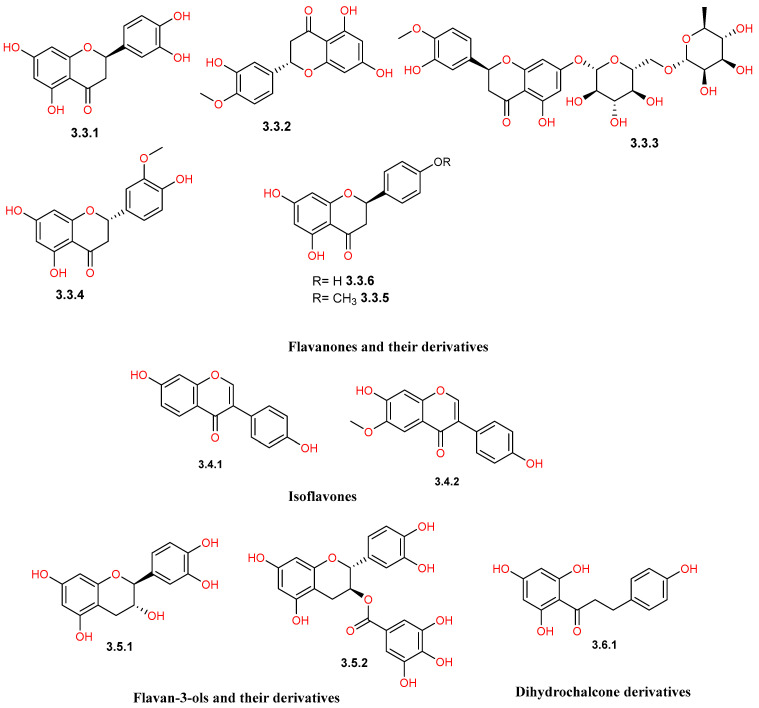 Figure 10
Figure 10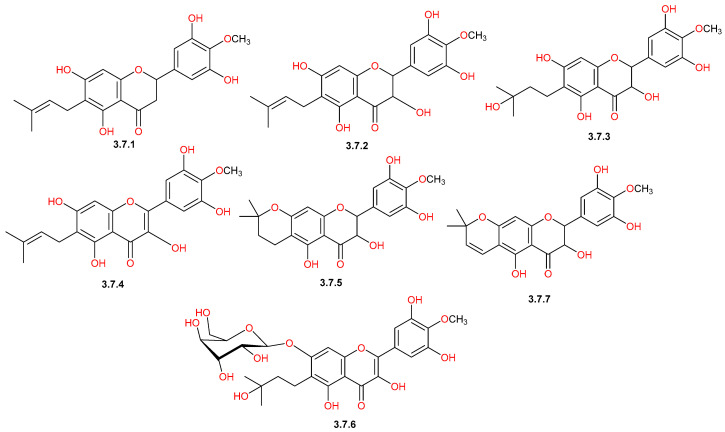 Figure 11
Figure 11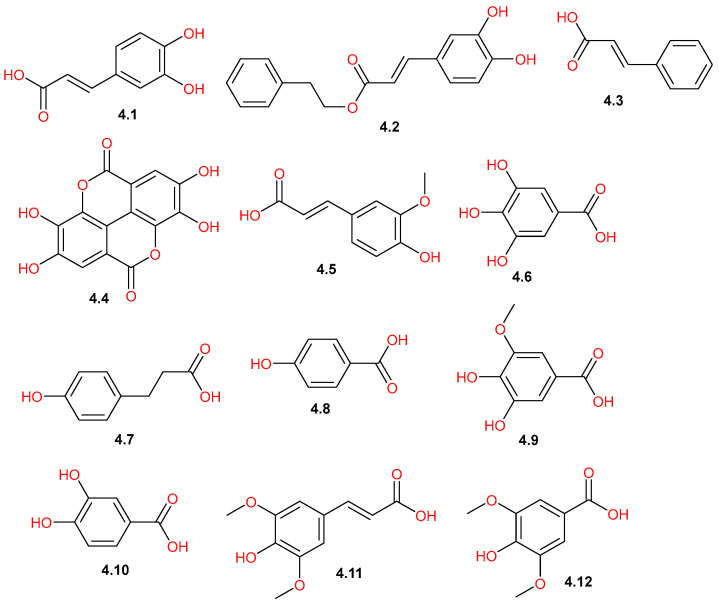 Figure 12
Figure 12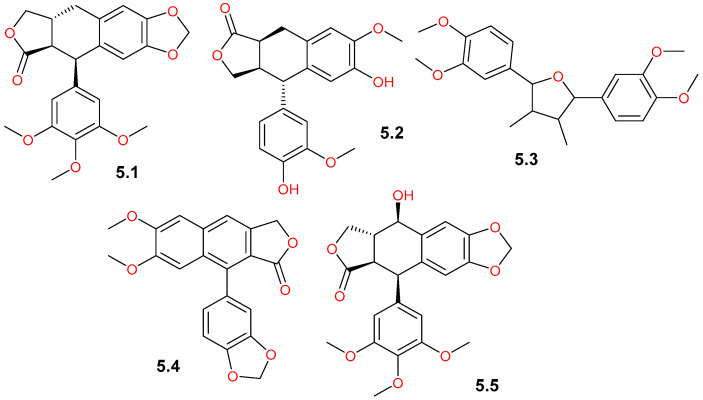 Figure 13
Figure 13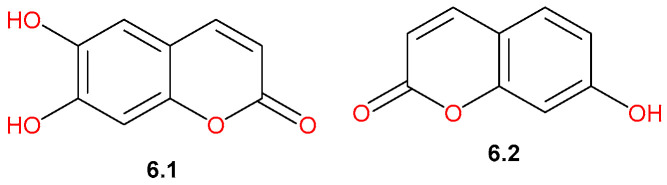 Figure 14
Figure 14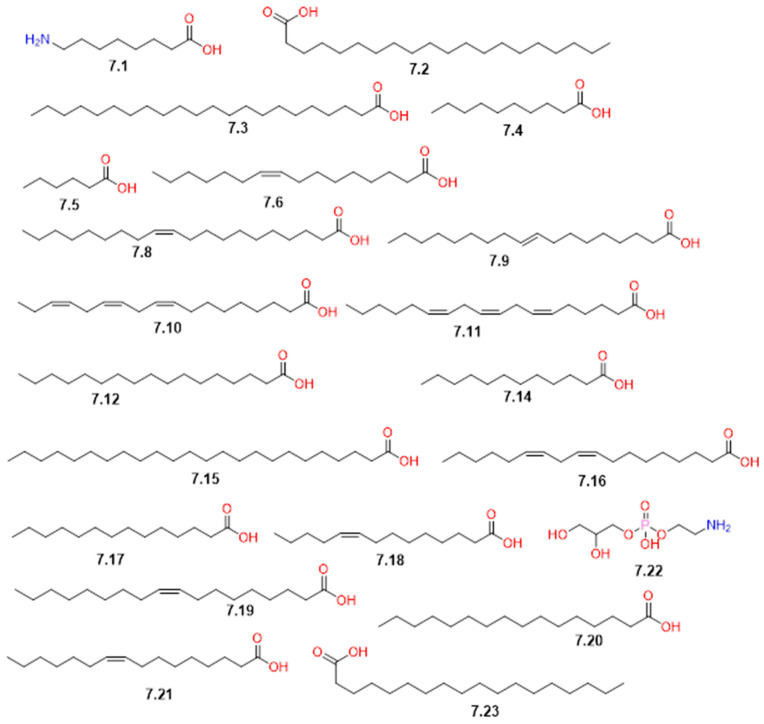 Figure 15
Figure 15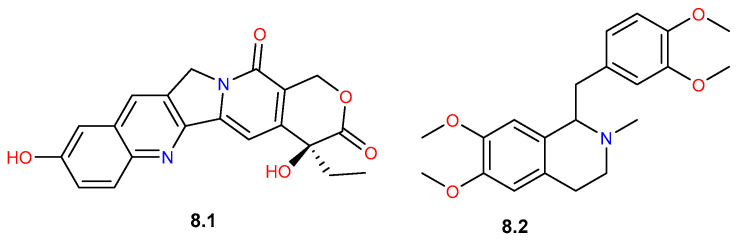 Figure 16
Figure 16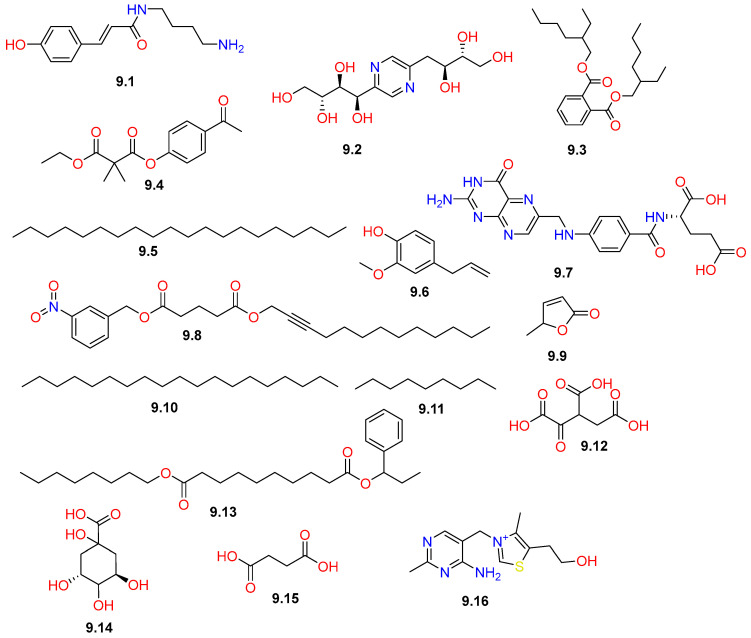 Figure 17
Figure 17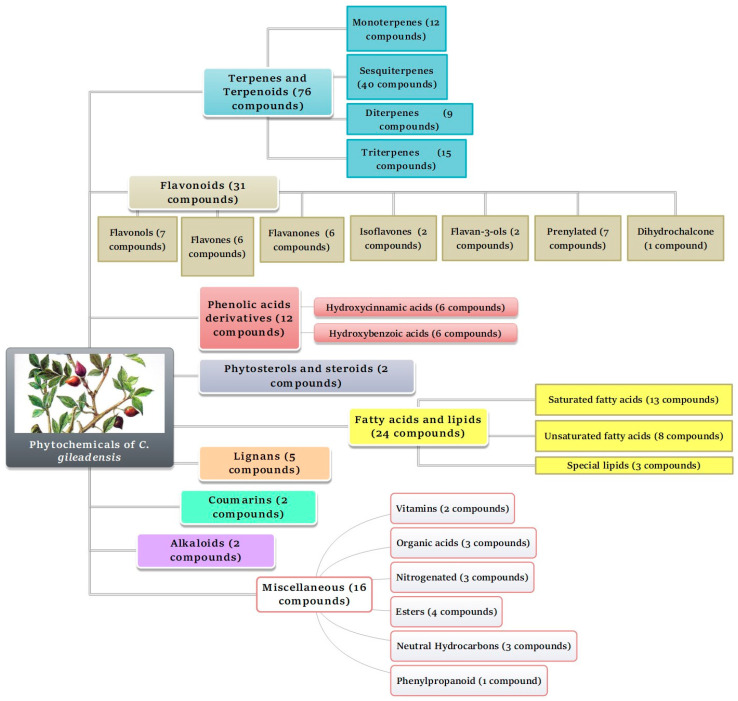 Figure 18
Figure 18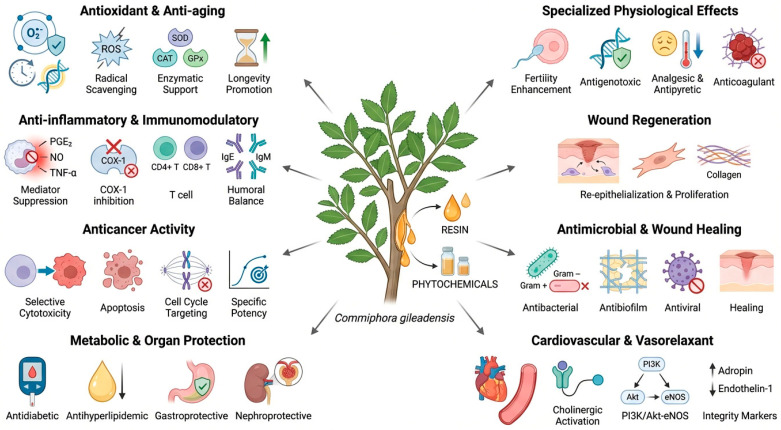 Figure 19
Figure 19Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPharmacological Effects of Medicinal Plants · Medicinal Plants and Neuroprotection · Saffron Plant Research Studies
1. Introduction
Plants provide invaluable resources to humanity. They hold a valuable treasure of secondary metabolites that have helped save lives since ancient times. Over the years, numerous applications of natural products have been identified and developed for human benefit [1]. At present, they represent a major and indispensable source for the discovery and development of new therapeutic agents. Thousands of studies have investigated phytoconstituents and their biological effects on human health, particularly in comparison to synthetic compounds, which may be associated with adverse effects alongside their intended pharmacological benefits [2,3].
The genus Commiphora is part of the family Burseraceae. It includes about 150 to 200 species found mainly in tropical and subtropical areas, such as Africa, Arabia, India, and some regions of South America. This genus is characterized by small trees or shrubs with spiny branches. Their barks release aromatic oleogum resins with characteristic smells [4,5,6]. The Commiphora genus has been known since ancient times for its various healing properties, economic uses, and distinct botanical traits [7]. Additionally, the resins are useful for embalming, incense, and perfumery [4,5,8,9]. Since ancient times, plants of this genus have been used to treat various ailments, including inflammation, arthritis, wounds, obesity, diabetes, hyperlipidemia, and infections. These traditional applications have been validated by modern pharmacological research, which demonstrated anti-inflammatory, antioxidant, antimicrobial, hypolipidemic, and anticancer properties, along with the identification of more than 300 secondary metabolites [4,5,6,9].
Significantly, C. gileadensis (L.) C.Ch. is a well-known aromatic and medicinal plant in the Commiphora genus with a long history and several different names documented in the literature. The species name “gileadensis” and the popular common name “Balm of Gilead” refer to the ancient Gilead region east of the Jordan River [10]. Other popular names include “Balm of Makkah,” which is commonly used in Saudi Arabia and the Arabian Peninsula; “Bisham” and “Basin,” which are regional names frequently used in Arabic-speaking areas; “Mecca Balsam,” which is sometimes used in English-language sources [11], and Apharsemon in Hebrew [12]. These names reflect the historical use of the plant in perfumery, medicine, and religious rituals. In addition, C. opobalsamum and Balsamodendron opobalsamum are widely recognized synonyms [13]. In desert folklore, C. gileadensis is known as a “weather predictor” plant. Its growth and blooming cycles are traditionally used to predict rainfall and changes in weather patterns; this phenomenon is thought to be more prominent in this species than in other members of the genus Commiphora [14].
The resin of C. gileadensis was famous in ancient Arabia and the Middle East for its opulent fragrance as well as medicinal and therapeutic properties. It was a major constituent in incense and perfumes, contributing to its economic and cultural importance for thousands of years. Its essential oil and resin were used to prepare aromatic products that were traded and valued by various communities [14]. It contains a complex mixture of phenolic compounds, essential oils, and other phytochemicals, which potentially contribute to a broad spectrum of traditional therapeutic applications, including skin, liver, digestive, inflammatory, infectious, and even cancerous conditions, supported by evidence of antioxidant, antimicrobial, and hepatoprotective activities [9,15]. The primary methods of traditional use include topical application, oral administration, and inhalation [10,11,13,16,17].
Due to overharvesting and increased aridification brought on by climate change, the species is in danger of going extinct due to a sharp decline in population and loss of genetic diversity. This underscores the necessity of conservation strategies like controlled harvesting, controlled grazing, water management, and seed bank preservation [18].
C. gileadensis produces both resinous and non-resinous components, which are chemically characterized by many studies as attempts to understand their composition and bioactivity. The oleogum resin is rich in terpenoids, including cycloartane derivatives, which are key metabolites identified by NMR-based metabolomics, distinguishing it from other Commiphora species [19]. It also contains volatile compounds, such as α-pinene, sabinene, β-pinene, and p-cymene, which contribute to its characteristic intense, terpenic, and lemony aroma, as well as biological activities, including anti-inflammatory and wound healing effects [20]. Non-resinous components include amino acids, organic acids, sugars, phenolic acids, and polyphenols, which are also present in the cell sap and contribute to the pharmacological profile of the plant [19].
This review aims to present a thorough and comparative summary of the biological activities and phytochemical composition of resinous and non-resinous components of C. gileadensis. It aims to highlight the similarities and differences in their bioactive constituents and pharmacological potential by integrating available data.
2. Materials and Methods
2.1. Literature Search Strategy
An updated comprehensive literature search was conducted using electronic databases, including ScienceDirect, PubMed, Scopus, Google Scholar, and Web of Science. The search extended over the period from 1997 to 2025 and included peer-reviewed journal articles, reviews, books, theses, and authoritative reports. The following Boolean search query was applied (with minor syntax adjustments depending on database requirements): (“Commiphora gileadensis” OR “C. gileadensis” OR “Commiphora opobalsamum” OR “C. opobalsamum” OR “Balsamodendron opobalsamum” OR “Balm of Gilead” OR “Mecca Balsam” OR “Bisham”) AND (“phytochemistry” OR “chemical composition” OR “secondary metabolites” OR “essential oil*” OR “volatile oil*” OR “terpen*” OR “sesquiterpen*” OR “monoterpen*” OR “triterpen*” OR “flavonoid*” OR “alkaloid*” OR “resin” OR “oleogum resin” OR “leaves” OR “stems” OR “barks”) AND (“biological activity” OR “pharmacological activity” OR “antioxidant” OR “anti-inflammatory” OR “antimicrobial” OR “antibacterial” OR “anticancer” OR “cytotoxic*” OR “antidiabetic” OR “wound healing” OR “toxicity” OR “pharmacokinetic*”). Searches were performed using the default settings of each database. Although no formal field restriction was applied, emphasis during screening was placed on studies in which the key terms appeared in the Title, Abstract, or Keywords to enhance specificity. Chemical Abstracts Service (CAS) registry numbers were obtained through searches conducted in the Reaxys database (Elsevier), using compound names and structural information to ensure accurate matching [21].
2.2. Inclusion and Exclusion Criteria
Only English-language publications that provided phytochemical analyses and/or biological evaluations (in vitro or in vivo) of C. gileadensis were included. Articles that addressed other Commiphora species without comparative data on C. gileadensis, lacked methodological rigor, or were duplicate reports were excluded.
2.3. Data Extraction and Synthesis
Relevant data were manually extracted, tabulated, and categorized based on plant part studied (resin, leaves, bark, stems), major phytochemical constituents (e.g., terpenes, flavonoids, alkaloids), biological activities (e.g., antioxidant, antimicrobial, anti-inflammatory, cytotoxic), extraction and analytical methods used, and traditional and ethnopharmacological uses. Comparative tables were designed to highlight similarities and distinctions among plant parts. Emphasis was placed on studies employing validated experimental models and analytical tools such as GC-MS, LC-MS, HPLC, and NMR. Tentatively identified compounds of likely synthetic origin were excluded.
3. Botanical and Taxonomical Characteristics
Burseraceae is a family of flowering plants belonging to the order Sapindales and composed of about 18 genera comprising 700 species of resinous trees and shrubs [22]. Several of its species are economically important for their fragrant resins.
Commiphora Jacq is the most species-rich genus in the family Burseraceae, comprising approximately 190 species, 21 of which are listed in the ‘Red List of Threatened Species’ of The International Union for Conservation of Nature (IUCN) [23]. They are small deciduous trees or shrubs with short, often thorny branches and papery bark. The leaves are usually compound, and the plants can be identified by their resin canals and distinctive seed morphology. Morphological and anatomical features, such as the presence or absence of spines and axial parenchyma, are used to differentiate species within the genus [4,17,24].
C. gileadensis (L.) C.Chr. is a small perennial tree or shrub, native to the southern and western mountains of the Arabian Peninsula. It belongs to the Kingdom Plantae, Phylum Angiosperms, Class Eudicots, Order Sapindales, and Family Burseraceae, and is classified within the Genus Commiphora [25]. It is a dioecious plant bearing scaly, dark-grey branches and small pubescent compound leaves with 3–5 leaflets (Figure 1). The flowers range from cream to red colored. The fruits are edible, red single-seeded drupes with black seeds and with white dividing lines creating the appearance of being four-lobed fruits [14]. When the bark is incised, it exudes a glossy dark brown to black fragrant oleogum resin, a key characteristic feature for the plant. Its best geological conditions are the eroded black basalt and granite rocks, with an elevation range of about 100 to 750 m above sea level, and it was not recorded on coastal plains with deep soil [17,26].
4. Phytochemical Profile of C. gileadensis
C. gileadensis has been the subject of extensive phytochemical research, which has consistently demonstrated that the plant is remarkably rich in secondary metabolites, particularly flavonoids and terpenes, to which a wide spectrum of pharmacological activities is potentially attributed. The phytochemical profile of the plant (Table 1) was represented by 10 chemically distinct groups, dominated by terpenoids, especially mono-, sesqui-, di-, and triterpenes, which were found in a wide array of plant material, including aerial parts, leaves, stems, fruits, bark, and particularly the resin/balsam.
4.1. Terpenes and Terpenoids
4.1.1. Monoterpenes
Monoterpenes are a class of natural compounds derived from two linked isoprene units, giving them a basic structure of ten carbon atoms (C_10_H_16_) [50,51,52]. Monoterpenoids (denoted by the suffix “-oid”) are monoterpenes with additional functional groups such as alcohol and ketone. Both groups are widely found in the essential oils of aromatic plants, where they often form the majority of volatile components [50,51,53]. C. gileadensis is rich in monoterpenoids, which constitute the dominant components of essential oils in specific chemotypes and are primarily responsible for defining its characteristic aroma profile (Table 1 and Table 2). Compounds 1.1.1–1.12 (Figure 2) represent the fundamental monoterpene profile of C. gileadensis in which monoterpenes hydrocarbon drive volatility and aroma intensity, while oxygenated monoterpenoids enhance chemical stability and biological relevance [54]. Monoterpene hydrocarbons, including β-myrcene (1.1.4), sabinene (1.1.8), α-pinene (1.1.5), β-pinene (1.1.6), β-phellandrene (1.1.7), γ-terpinene (1.1.9), and β-thujene (1.1.12), are the primary contributors to the fresh, resinous, and citrus-peppery aroma of the oil [54]. For instance, β-myrcene predominated in EO from Medina (Badr) samples, reaching 17.44%, accompanied by β-phellandrene (9.59%), indicating a monoterpene-hydrocarbon-rich chemotype [11]. In contrast, EO from plants cultivated in the Middle East but native to Saudi Arabia show strong domination of sabinene (22.7%) and α-pinene (14.4%) in fresh aerial parts, while resin and exudates are interestingly dominated by sabinene (43.8–46.4%) and α-pinene (24.0–25.8%), defining the characteristic balsamic aroma [20,54]. Aromatic monoterpenes such as p-cymene (1.1.1) serve as background components that stabilize the warm aromatic character of the oil and frequently co-occur with resin and aerial-part profiles [54]. Its oxygenated derivative, p-cymen-8-ol (1.1.2), was mainly reported in extracts and essential oils of aerial parts, enriching the oxygenated fraction [12,32]. Oxygenated monoterpenes, particularly terpinen-4-ol (1.1.10), are key EO components, reaching 8.5–9.8% in EOs from fresh aerial parts and flowering tops of plants from Makkah and 18.7% in oils from Almog, Palestine [20,33]. Additional contributors of this group, including limonene (1.1.3) and γ-terpinene (1.1.9), provide citrus-fresh nuances and act as biosynthetic or oxidative precursors to oxygenated derivatives [32,54].
4.1.2. Sesquiterpenes
Sesquiterpenes is a diverse class of natural compounds made up of three isoprene units, resulting in a 15-carbon backbone. They are widely distributed in plants, marine organisms, and microbes, and are considered the most structurally varied group among terpenoids. The basic structure of sesquiterpenes can be acyclic, monocyclic, bicyclic, tricyclic, or even multicyclic, and they often include derivatives such as alcohols, ketones, lactones, and epoxides (sesquiterpenoids) [55,56]. Available data suggests that the sesquiterpene group is the most abundant group of compounds reported from C. gileadensis, especially in the aerial parts and resin, with only a few reported from the oil (Table 1). Among these compounds, 4α,10α-dihydroxy-1α,5αH-guaia-6-ene (1.2.18) presented a discrepancy in its reported stereochemical configuration. Although Shen, et al. [35] described it as a known compound, examination of their cited reference revealed a different structure, namely guaia-6(7)-en-4α,10β-diol [35], possessing a different stereochemical configuration. Moreover, this latter compound was identified based on structural similarity to a previously reported compound, guaianediol [57]. Therefore, further investigations are required to confirm the stereochemical configuration of compound 1.2.18.
The sesquiterpene profile of C. gileadensis exhibited a significant structural diversity, and its chemical composition is highly impacted by the plant part used, the preparation technique used, and the geographical origin. Members of the cadinane, germacrane, and eudesmane classes are among the principal reported compounds by many investigations (Figure 3 and Figure 4).
The distribution of sesquiterpene hydrocarbons varied considerably across different chemotypes based on variations in sources, in addition to plant parts and extraction methods. In Ein Gedi, Palestine, the leaves and fruits extracted with MTBE were dominated by β-caryophyllene (1.2.11) (20.12%) and germacrene D (1.2.23) (19.62%) [34]. However, in samples from Medina (Badr) region, the analysis of a 70% ethanolic extract from fresh aerial parts revealed a profile particularly rich in copaene (1.2.13) (11.48%), α-muurolene (1.2.30) (9.01%), and β-selinene (1.2.39) (5.02%) [11]. By contrast, samples from Makkah processed via hydrodistillation of fresh aerial parts and flowering tops show a prevalence of α-calacorene (1.2.8) (9.4%), cadalene (1.2.3) (5.4%), and δ-cadinene (1.2.5) (4.8–5.0%) [33]. Furthermore, samples from the Breiman region utilizing successive solvent extraction of shoots and aerial parts identified γ-himachalene (1.2.27) (21.43%) as a major constituent, highlighting the structural diversity of sesquiterpenes present within the species across different Saudi Arabian localities [32].
Another key class of the C. gileadensis profile is oxygenated sesquiterpenoid derivatives (Table 1 and Table 2; Figure 3 and Figure 4). In Saudi Arabian samples, viridiflorol (1.2.40) and spathulenol (1.2.38) are commonly reported as major components; in the Breiman region, viridiflorol reached 9.63%, while in Makkah and Khulais, it reached 4.9–5.41% [32,33]. Furthermore, dominant sesquiterpenoid markers in Makkah and Khulais samples have been found to be α-cadinol (1.2.7) (10.1%) and β-eudesmol (1.2.21) (11.9%) [32,33]. The chemical complexity of this secondary metabolite class was highlighted by the report of two unique isolated sesquiterpenoids, myrrhanolide D (1.2.33) and myrrhasin A (1.2.34) (Figure 4), from resin samples imported from India and extracted by refluxing with ethyl acetate [35].
4.1.3. Diterpenes
Diterpenes are isoprene-derived compounds consisting of four isoprene units (C_20_). They are classified according to rings in their chemical structures into acyclic, mono-, di-, tri-, tetra-, and macrocyclic, in addition to other miscellaneous structures.
The diterpenoids reported in C. gileadensis represent a chemically diverse group, with a significant existence of verticillane-type and resin acid derivatives (Table 1 and Figure 5). These compounds are distributed across various plant materials, including stems, leaves, bark, and resinous exudates.
Verticillanes are a predominant class of diterpenoids identified in the stems of the plant. (13S,14S)-ent-13,14-epoxyverticillol (1.3.2) and (9S,10S)-ent-9,10-epoxyverticillol (1.3.3) are two epoxy-verticillane derivatives isolated along with the alcohol derivative, ent-verticillol (1.3.8), from the stem extract of C. gileadensis [31,39]. Additionally, (1S,3E,7E,11R)-(+)-verticilla-3,7,12(18)-triene (1.3.7) was remarkable for its presence in both organic extracts and the essential oil of the stems [31,36,39].
The resin and balsam of the plant served as a primary source for diterpene acids or resin acids. Dehydroabietic acid (1.3.1), an abietane-type diterpenoid, was only identified in the resin of C. gileadensis, indicating the role of secretory tissues as major sites for diterpenoid acid accumulation. Similarly, sandaracopimaric acid (1.3.5), a pimarane-type diterpenoid acid, was isolated from resin or balsam, supporting the accumulation of diterpenoid acids in resinous matrices [38].
The plant leaves extracted with integrated ultrasonic-microwave-assisted methods showed the presence of the diterpenoid, sordarin (1.3.6). This extract displayed a capacity to synthesize silver nanoparticles with enhanced antibacterial properties [40].
4.1.4. Triterpenes
Triterpenes are a diverse group of natural compounds with a 30-carbon backbone that are composed of six isoprene units and include steroids, bile acids, and saponins [58]. The majority of triterpenoids are tetracyclic (6-6-6-5; such as lanostanes, cycloartanes, dammaranes, euphanes, and tirucallanes) and pentacyclic (6-6-6-6-5 or 6-6-6-6-6; such as friedelanes, lupanes, oleananes, ursanes, hopanes, and taraxasteranes) types, although acyclic, monocyclic, bicyclic, tricyclic, and hexacyclic triterpenoids have been reported from natural sources, with pentacyclic triterpenoids representing the largest occurrence in nature [59].
The triterpenoid contents reported in C. gileadensis included both tetra- and pentacyclic-type derivatives, which can be categorized into the cycloartane, friedelane, oleanane, and ursane classes (Table 1). They were found to be distributed in the resin, stems, and aerial parts of the plant. Resinous exudates were rich in acetylated, hydroxylated, and epoxy derivatives of the tetracyclic cycloartane-type triterpenoids, as displayed in Figure 6. The acetoxycycloartane derivatives included 1α-acetoxycycloartan-24-ene-2α,3β-diol (1.4.1) and 3β-acetoxycycloartan-24-ene-1α,2α-diol (1.4.2), along with multiple hydroxylated derivatives, including cycloartan-23E-ene-1α,2α,3β,25-tetrol (1.4.5), cycloartan-24-ene-1α,2α,3α-triol (1.4.6), cycloartan-24-ene-1α,2α,3β-triol (1.4.7), and cycloartan-24-ene-1α,3β-diol (1.4.8) [41]. Two epoxycycloartane derivatives, including 24R,25-epoxycycloartane-1α,2α,3β-triol (1.4.9) and its 24S-epimer (1.4.10), as well as the valerate ester derivative of 1.4.6, namely 3β-isovaleroyloxycycloartan-24-ene-1α,2α-diol (1.4.11), were also reported in the resin [41]. In contrast, the stems and aerial parts showed the presence of pentacyclic triterpenoids with friedelane, oleanane, and ursane skeletons, such as canophyllal (1.4.3) and friedelin (1.4.12), oleanonic acid (1.4.13) and oleanonic aldehyde (1.4.14), and the ursane-type urs-12-en-3-one-28-al (1.4.15) [39,42]. Remarkably, the stems showed the presence of the friedelan-type triterpenoid, commigileadin A (1.4.4) [39,43].
4.2. Phytosterols
Phytosterols are triterpene-derived metabolites, but undergo structural rearrangement and side-chain modification, resulting mostly in the production of C28 (e.g., campesterol) and C29 (e.g., β-sitosterol, stigmasterol) sterols. They are characterized by the presence of a sterane (steroid) ring, a β-hydroxyl group at C-3, and unique alkyl side chains at the C-24 position and serve as structural components of plant cell membranes [60]. Several studies reported the existence of phytosterols in the aerial parts, leaves, and resin of C. gileadensis, including guggulsterone (2.1) and stigmasterol (2.2), Figure 7 [35,43,44].
4.3. Flavonoids
Flavonoids are a major subclass of polyphenols, distinguished by a C6-C3-C6 structural framework, consisting of two aromatic rings linked by a three-carbon bridge [61,62]. In the plant kingdom, these secondary metabolites are essential for pigmentation, UV protection, and defense against environmental stressors [63].
Reported phytochemical studies on C. gileadensis revealed a rich flavonoid profile of C. gileadensis, demonstrating a wide spectrum of structural diversity (Table 1 and Figure 8, Figure 9, Figure 10 and Figure 11). Flavonols represented one of the major classes (Figure 8), comprising seven reported compounds with quercetin (3.1.5) as the most commonly detected flavonol in different plant materials, including aerial parts, leaves, and bark [25,32,40,42,43,45]. This class also included kaempferol (3.1.1) and mearnsetin (3.1.2), which are mainly extracted from aerial parts and bark [42,43,45,46]. Within the flavones class (Figure 9), apigenin (3.2.1) was isolated from both aerial parts and bark [11,45], whereas saponarin (3.2.6) was only detected in leaf extracts [40]. The flavanone group (Figure 10) was represented by compounds, such as naringenin (3.3.6), which was detected in both aerial parts and bark [11,43,45], and hesperidin (3.3.3), detected in the aerial parts [11]. Furthermore, the plant is characterized by the presence of a unique prenylated flavonoid group (Figure 11), such as comophorin A (3.7.1) and comophorin B (3.7.2), which are specifically reported in the stem bark [45,46,47]. Finally, other minor classes contributed to this flavonoid profile, including isoflavones such as daidzein (3.4.1) and dihydrochalcones such as phloretin (3.6.1), both of which were identified in bark extracts, as displayed in Figure 10 [45].
4.4. Phenolic Acids
Phenolic acids are a significant subclass of plant polyphenols characterized by a phenol moiety and a resonance-stabilized structure, which enables free radical scavenging through hydrogen atom donation [61,63]. These compounds are generally subdivided into two main subgroups, including hydroxybenzoic acids and hydroxycinnamic acids, with the latter subgroup comprising caffeic and ferulic acids, which are commonly found in edible plants [62].
Reported phytochemical investigations of C. gileadensis identified a diverse group of phenolic acids (Figure 12) found particularly within the stem bark [45]. The hydroxycinnamic acid subgroup was represented by compounds such as caffeic acid (4.1), cinnamic acid (4.3), ferulic acid (4.5), hydro-p-coumaric acid (4.7), and sinapic acid (4.11), which were detected in the bark extract [45], while caffeic acid phenethyl ester (4.2) was identified in the aerial parts [11]. On the other hand, the hydroxybenzoic acid subgroup comprised gallic acid (4.6), which appeared to be abundant in the aerial parts and the bark [11,45]. Additionally, several benzoic derivatives were detected in the bark, including 4-hydroxybenzoic acid (4.8), 3-O-methyl gallic acid (4.9), and protocatechuic acid (4.10) [45]. Notably, syringic acid (4.12) was isolated from aerial part extract [42], while ellagic acid (4.4) was detected in the bark extract [45].
4.5. Lignans
Lignans are a large group of polyphenols that function as structural building blocks of plant cell walls. These compounds are typically dimers, or occasionally trimers and tetramers, of phenylpropane units (C6-C3), known as monolignols, formed through oxidative coupling reactions [64].
By reviewing the available literature, lignans were found to be identified in the leaves of C. gileadensis. Several lignan derivatives (Table 1 and Figure 13) were tentatively identified by UPLC-qTOF-MS in the leaf extracts, including anthricin (5.1), β-conidendrin (5.2), (-)-galbelgin (5.3), and the medicinally important lignan, podophyllotoxin (5.5) [40]. Furthermore, a biotechnological study utilizing tissue culture has tentatively identified justicidin B (5.4) in the leaves, callus, and cell suspension cultures of C. gileadensis by LC-MS/MS analysis, suggesting that the phenylpropanoid pathway remains active in undifferentiated cell systems [23].
4.6. Coumarins
Coumarins are a class of naturally occurring benzopyrone compounds composed of a benzene ring fused to a pyrone ring. These natural products are aromatic and fragrant; their aroma often resembles woodruff, fresh hay, or vanilla. Beyond their organoleptic characteristics, coumarins demonstrate a wide spectrum of biological activities, which are dependent on their chemical substitutions and structural configurations [65].
Reported phytochemical studies targeting analysis of the bark extract of C. gileadensis using the LC-MS/MS technique have led to the tentative identification of two coumarin derivatives (Figure 14), including 6,7-dihydroxycoumarin (6.1) and umbelliferone (6.2) [45].
4.7. Fatty Acids and Lipids
Lipids are oily, fatty, or waxy compounds soluble in organic solvents, comprising four primary subgroups: fats and oils (triacylglycerols), phospholipids, waxes, and steroids [66]. Fatty acids serve as the fundamental building blocks for these lipids and are characterized as hydrophobic organic molecules with unique aliphatic chains, either saturated or unsaturated, terminating in a carboxylic acid moiety, commonly 12 to 18 carbons in length [67]. The chain length, branching, and degree of saturation determine the physical properties of lipids and their biological role [66,67].
In C. gileadensis, this class is represented by a variety of derivatives identified in various plant parts (Table 1 and Figure 15). This class can be categorized into four distinct groups with various lengths, substitutions, and degrees of saturation: saturated fatty acids, unsaturated fatty acids, fatty acid derivatives, and special lipids. Saturated fatty acids constituted the most abundant class detected in the resin, bark, and stem. Among these, palmitic acid (7.20), caprylic acid (7.6), and stearic acid (7.23) were found in high concentrations within the resinous balsam at 15.50%, 12.60%, and 10.73%, respectively [48]. While palmitic acid (7.20) is distributed across the stem and resin [37,48], stearic acid (7.23) and arachidic acid (7.2) were found in both the bark and resin [45,48]. This group also included behenic (7.3), capric (7.4), caproic (7.5), heptadecanoic (7.12), lauric (7.14), lignoceric (7.15), and myristic (7.17) acids, which were reported mainly in the resinous balsam [48].
On the other hand, the unsaturated fatty acids, oleic acid (7.19) was the most abundant member of this group, comprising 33.36% of the balsam oil [48]. This class also comprised monounsaturated acids, such as palmitoleic (7.21) detected in the bark and resin [45,48], alongside myristoleic (7.18), elaidic (7.9), and cis-11-eicosenoic (7.8) acids in the resin [48]. Polyunsaturated fatty acids, including linoleic (7.16), γ-linolenic (7.11), and α-linolenic (7.10) acids, have also been reported in the resin [48]. Regarding fatty acid derivatives, the amino-fatty acid 8-aminocaprylic acid (7.1) was exclusively identified in leaf extracts [40]. Finally, special lipids were tentatively identified through LC-MS analysis in the aerial parts of C. gileadensis, including leaves and branches. They included three compounds: ceramide (7.7), hexosylceramide (7.13), and phosphatidylethanolamine (7.22) [49].
4.8. Alkaloids
Alkaloids class is an unusual family in the Commiphora genus. However, two alkaloids have been tentatively determined during LC-MS/MS-based chemical profiling of the plant and its tissue culture (Figure 16) [23]. 10-Hydroxycamptothecin (8.1) was detected only in the methanol extract of the leaves, whereas laudanosine (8.2) was detected in cell suspension culture, as well as in the seeds and leaves of the wild plant [23].
4.9. Miscellaneous Compounds
The miscellaneous group of compounds (9.1–9.16) displayed in Figure 17 comprises a diverse array of bioactive metabolites identified in various plant materials of C. gileadensis, including the bark, leaves, stems, and aerial parts (Table 1).
They represented several chemical classes, including vitamins, organic acids, nitrogenated compounds, and long-chain hydrocarbons (Figure 17). Particularly, the bark showed the presence of nitrogen-containing derivatives, including a phenolamide, coumaroylputrescine (9.1), and a pyrazine derivative, 2,6-deoxyfructosazine (9.2), as well as carboxylic acids, including oxydisuccinic acid (9.12), quinic acid (9.14), and succinic acid (9.15) [45]. Investigations of the leaves revealed the presence of several miscellaneous esters, including dimethylmalonic acid, 4-acetylphenyl ethyl ester (9.4), glutaric acid, tridec-2-yn-1-yl 3-nitrobenzyl ester (9.8), and sebacic acid, octyl 1-phenylpropyl ester (9.13) [40]. The aerial parts were noted for their di-(2-ethylhexyl)-phthalate (9.3) content, in addition to aliphatic hydrocarbons, including eicosane (9.5), and nonadecane (9.9) [32], as well as nonane (9.11). It is worth noting that the presence of the phthalate derivative (9.3) in the aerial part extract most likely arises from contamination of the extraction solvents rather than being an endogenous plant constituent, as phthalate esters are well-documented contaminants introduced during solvent extraction and analytical procedures [68]. Additionally, eugenol (9.6) was identified in both the aerial parts and stems [33]. Finally, the stem was found to be a source of essential nutrients, such as folic acid (9.7), 5-methyl-2(5)-furanone (9.9), and vitamin B1 (9.16) [37].
4.10. Phytochemistry of Essential Oil
The aromatic and bioactive nature of the plant is highly attributed to the abundant terpenoid content found in the essential oil. Research on the essential oil (EO) of C. gileadensis revealed distinct chemotypic divergence, with its composition significantly based on geographical origin, the plant organ processed, and the specific extraction or analytical techniques used [11,20,32,33]. Table 2 summarizes these variation factors and compares the major chemical components identified by different studies.
The comparative data presented in Table 2 highlighted marked variability in the EO composition of C. gileadensis, which can be directly attributed to differences in geographical origin, plant organ, and extraction or preparation methods. EOs obtained from the Makkah region, Saudi Arabia, were consistently dominated by sesquiterpenes; for example, hydrodistilled oils from fresh aerial parts contained α-calacorene (9.4%), terpinen-4-ol (8.5%), and δ-cadinene (5.0%), while oils from stored aerial material showed the presence of more stable oxygenated sesquiterpenes, such as α-cadinol (10.1%), spathulenol (5.8%), and viridiflorol (4.9%) [33]. In contrast, samples collected from the Badr area, Medina, showed an obviously different chemotype, with steam-distilled oils rich in monoterpene hydrocarbons, particularly β-myrcene (17.44%) and β-phellandrene (9.59%), alongside nonane (10.88%) and the diterpene verticiol (10.56%) [11]. These findings support the influence of geographical variation and local environmental conditions on terpene biosynthesis.
Organ-based variation was also evident. Fresh flowering tops from Makkah were rich in terpinen-4-ol (9.8%), whereas fresh aerial parts from the same region were dominated by α-calacorene (9.4%) [33]. EOs extracted from leaves and fruits cultivated in Ein Gedi (Palestine) exhibited a balanced profile of sabinene (21.11%), β-caryophyllene (20.12%), and germacrene D (19.62%), reflecting mixed mono- and sesquiterpene biosynthesis [34]. The resin and exudate represented a distinct chemical niche. It was mainly made up of monoterpene hydrocarbons. Direct injection analysis showed sabinene at 43.8% and α-pinene at 24.0%. HS-SPME results highlighted sabinene at 46.4% and α-pinene at 25.8%. It also detected other highly volatile components, including α-thujene at 4.3%.
The extraction technique exerts a crucial effect on the chemical profile of the produced EO. Steam or hydrodistillation methods preferentially recover volatile monoterpenes, while solvent-based methods selectively enrich heavier sesquiterpene hydrocarbons. This was illustrated in Medina samples, where steam-distilled oil was characterized by 36.9% monoterpene hydrocarbons dominated by β-myrcene, while a 70% ethanolic extract of the same material was rich in sesquiterpenes, including copaene (1.2.13) (11.48%), α-muurolene (1.2.30) (9.01%), and β-selinene (1.2.39) (5.02%) [11].
Taken together, the pronounced chemical variability of EOs from C. gileadensis emphasizes the need for a standardized source and methodology that will allow a valid comparison of chemical composition and biological activity.
The phytochemical classes reviewed in this study highlight the remarkable chemical diversity of C. gileadensis (Figure 18).
5. Ethnobotanical and Traditional Uses of C. gileadensis
The title plant, C. gileadensis has a long history of use in traditional medicine, particularly in ancient and Arabic medicine (Table 3). It was recognized by names such as Arabian balsam, Bisham, becham, or balessan medicine [23,44,69]. Historically, the sap or resin was the most highly valued material, which was utilized topically for thousands of years as a potent antiseptic and wound-healing agent, particularly in skin conditions such as burns, wounds, and skin infections [23,44,49,69]. Beyond topical applications, the resin was traditionally indicated for headaches, paralysis, hearing disorders, stroke, respiratory ailments, fractures, gastrointestinal ailments, arthritis, and weight reduction, as well as an antidote for scorpion stings and snake bites. It was even applied to the eyes to treat conditions such as cataracts and blurred vision [11,49,69]. The plant bark, especially the inner bark, was frequently employed as an antiseptic for injuries and, when ground into juice, as an anti-allergic treatment for skin conditions, such as burns, eczema, or inflammation [33,37,70]. Moreover, the bark aqueous extract was used as an anti-hypertensive [71]. Traditionally, local populations in Palestine and other Arab regions have utilized the decoction of the flowers and leaves to manage pain, constipation, and to promote urine output or to expel renal calculi [13,71,72,73,74]. Crushed leaves were used in the treatment of eye tumors [17]. Medicinal preparations from seeds and wood were utilized for chest, kidney, and stomach complaints, and to relieve conditions, including rheumatism, scurvy, and jaundice [69,72]. In traditional gynecology, particularly in Yemen, various plant organs have been used to mitigate labor pain, as a contraceptive, and to treat cervical infections [37,69,73]. In Syria, the oil imported from Egypt was particularly indicated for cold, ear problems, excess phlegm (catarrh), and to massage arthritic joints [69]. Other traditional uses of the oil included treatment of epilepsy, tetanus, and gonorrhea [40,69]. Additionally, the twigs were utilized as a natural toothbrush (miswak) to maintain oral hygiene and were traditionally boiled to extract oil for treating traumatic injuries [33,37,69]. In certain regions, such as Oman, extracts were even documented as a historical treatment for rabies and as a cleansing bath for newborns [37].
6. Pharmacological Activities Reported for C. gileadensis
A wide pharmacological potential was reported for C. gileadensis, which was supported by its rich and varied secondary metabolite profile, Table 1 (such as terpenoids and polyphenols), validating its historical and current medicinal applications. Numerous studies were conducted using various in vitro, in vivo, or ex vivo models to investigate multiple bioactivities of various plant parts and unorganized products, including the resin, essential oil, polysaccharide, or crude extracts of C. gileadensis prepared by diverse extraction techniques, as well as drying conditions, and derived from different localities. Additionally, the traditional medicinal uses of the plant have been scientifically validated through numerous reported studies. Some reported biological activities for C. gileadensis are summarized in Table 4.
6.1. Antioxidant Activity
An imbalance between the generation of free radicals and the ability of cells to neutralize or eradicate them leads eventually to oxidative stress. It contributes to the development of many chronic and degenerative conditions, such as cancers, Alzheimer’s disease, and cardiovascular problems. Oxidative stress may result from a number of predisposing factors, such as smoking, UV radiation, and environmental irritants [93]. Antioxidants are molecules that have the capacity to neutralize free radicals, preventing their interaction with vital macromolecules, such as lipids, proteins, and DNA. Therefore, they reduce processes that can contribute to chronic and degenerative conditions through inhibiting lipid peroxidation, protein oxidation, and oxidative DNA damage [94].
Plant polyphenols and terpenoids are antioxidant molecules that exert their activity through several mechanisms, including scavenging and neutralization of free radicals, chelation of metals that catalyze the generation of free radicals (such as iron, copper, zinc, cadmium), and stimulating the secretion of antioxidant enzymes, such as superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx). Furthermore, they impede the activity of enzymes, which produce reactive oxygen species (ROS), such as NADPH oxidase and xanthine oxidase, and restore the activity of other functioning antioxidants, such as vitamin C and vitamin E [95,96,97].
The antioxidant potential of various plant parts of C. gileadensis has been extensively studied, displaying significant activities in both in vitro and in vivo models. In the in vitro models, the extracts investigated demonstrate potent radical scavenging activity using the DPPH, ABTS, and H_2_O_2_ assays. Reported studies comparing the antioxidant potential of the extracts of plant parts indicated that stem peel extracts were more potent radical scavengers than leaf extracts, with reported EC_50_ values of 1.06 µg/mL for the stem extract compared to 3.39 µg/mL for leaf extract by the DPPH method [80]. Extraction procedure significantly impacted these results; for instance, ultrasonic-assisted extraction (USE) of leaves yielded a higher total phenolic content (TPC) than the hydrodistillation method (118.71 vs. 101.47 mg GAE/g DM) and also exhibited greater DPPH radical scavenging [78]. Furthermore, the steam-distilled EO showed stronger radical scavenging activity (IC_50_ 22.2 µg/mL) compared to 70% ethanolic extracts (IC_50_ 56.5 µg/mL); however, the ethanolic extract was notably effective in the β-carotene bleaching (BCB) assay (IC_50_ 75.8 µg/mL) [11].
The in vivo models using diabetic rats demonstrated that C. gileadensis extracts effectively mitigated oxidative stress by strengthening the natural body defense. The aqueous extracts from twigs and leaves were shown to induce the activities of CAT, SOD, and GST, while reducing levels of malondialdehyde (MDA), a marker of lipid peroxidation [79]. Similarly, the butanol fraction of stem bark at a dose of 100 mg/kg has been shown to increase reduced glutathione (GSH) by 82.51% and SOD by 49.03%, while reducing MDA by 33.41% compared to the diabetic control group [45]. The methanol extract of the stem also exhibited high ferric reducing antioxidant power (FRAP) (1.95 mM FE/mg), which was comparable to the reference drug, ascorbic acid [81].
The reported antioxidant potential was largely attributed to the high concentration of terpenoids, polyphenols, and flavonoids. Monoterpenes, such as α-pinene (1.1.5), β-pinene (1.1.6), sabinene (1.1.8), γ-terpinene (1.1.9), and terpinen-4-ol (1.1.10), were suggested as major contributors to the free radical scavenging activity [11,78]. Similarly, sesquiterpenes, including β-caryophyllene (1.2.11), β-copaene (1.2.13), and germacrene D (1.2.23), were related to the protective antioxidant [34,78]. Whereas phenolics and flavonoids, such as quercetin (3.1.5), gallic acid (4.6), caffeic acid (4.1), and rutin (3.1.6), were suggested to play key roles in scavenging free radicals [11].
6.2. Anti-Inflammatory Activity
The reported anti-inflammatory activity of C. gileadensis was demonstrated utilizing various experimental in vitro and in vivo models, showing significant potential in both acute and chronic inflammation (Table 4). These activities are characterized by the multi-targeted suppression of diverse biochemical mediators, immune cells modulation, and inhibition of enzymes associated with inflammatory responses. Additionally, specific preparations of C. gileadensis demonstrated significant effectiveness in suppressing acute and chronic inflammation at non-toxic dosage levels (Table 4).
A methanolic extract of the aerial parts (at a dose of 500 mg/kg) has been shown to significantly reduce the accumulation of Prostaglandin E2 (PGE2) (40.8%), Nitric Oxide (NO) (55.47%), and Tumour Necrosis Factor-alpha (TNF-α) (19.06%) at the site of inflammation [83]. Furthermore, the 70% ethanolic extract of the aerial parts exhibited COX-1 inhibitory activity (450 µg/mL) and prevented protein denaturation (110.5 µg/mL), a key marker for anti-arthritic potential [11]. The ethanol extracts of the aerial parts also significantly reduced carrageenan-induced paw edema and suppressed granuloma formation in rat models, indicating significant efficacy in both acute and chronic inflammatory conditions [76,82]. The aerial parts extract was demonstrated to synergistically potentiate the effects of conventional NSAIDs (non-steroidal anti-inflammatory drugs), such as diclofenac [82].
Recent studies highlighted the ability of C. gileadensis to regulate the immune system by altering T-lymphocyte cells. This effect was primarily evaluated using the sap, methanol, and acetone extracts of the aerial parts. The investigated extracts significantly decreased total pro-inflammatory lymphocytes and peripheral cluster of differentiation (CD) subsets, including CD3^+^, CD4^+^, and CD8^+^ [86,87]. Conversely, the extract of the aerial parts induced a significant increase in T-regulatory (Treg) cells (CD4^+^ CD25^+^ and CD8^+^ CD25^+^), which have the advantage of maintaining immune tolerance and protecting against chronic inflammatory damage [86].
The anti-inflammatory activities are mainly attributed to the rich terpenoids, steroids, and phenolic content of the plant (Table 1). Of these, key identified constituents were suggested as major contributors to the anti-inflammatory action of the plant, including the steroidal guggulsterone (2.1), which was isolated from the leaves, acting as a potent anti-inflammatory agent by antagonizing the bile acid receptor and suppressing the NF-κB pathway, thereby downregulating pro-inflammatory cytokines and chemokines [44,98]. Sesquiterpenes, such as β-caryophyllene (1.2.11), β-copaene (1.2.13), and germacrene D (1.2.23), were suggested to have anti-inflammatory actions [34,78]. The diterpeneoids (resin acids), including dehydroabietic acid (1.3.1) and sandaracopimaric acid (1.3.5), isolated from the resin, exhibited vasorelaxant and anti-inflammatory-related actions on pulmonary arteries via the PI3K/Akt-eNOS signaling pathway [38]. Finally, phenolic compounds, including caffeic acid, gallic acid (4.6), phenethyl ester (4.2), and flavonoids (Table 1) were suggested as potential contributors to the observed COX-1 inhibitory activity and protein denaturation suppression of the investigated aerial parts extract [11].
6.3. Anticancer Activity
The ability of extracts or certain isolated compounds of C. gileadensis to target malignant cells has been extensively investigated (Table 4). They demonstrated broad selective cytotoxic effects against various cancer types (Table 4).
The stem ethanol extract and EO were shown to have strong cytotoxicity against mouse lymphoma (BS-24-1) and MoFir (Epstein–Barr virus (EBV)-transformed human B lymphocytes) [34]. The extracts showed no inhibitory effect on normal human skin fibroblasts, indicating selective cytotoxicity [13,34]. The mechanism of cell death was shown to be associated with caspase-3 activation and DNA fragmentation [34]. Furthermore, the cytotoxicity of the sap was found to be cell cycle-dependent, targeting transformed cells in the S or G_2_/M phases of division, leaving cells in the pre-replicative G_1_ phase unaffected [13].
Diverse extracts and products derived from C. gileadensis exhibited varying degrees of potency against different cancer types. For instance, polysaccharides extracted from the stems showed potent antiproliferative activity (IC_50_ 13.15 µg/mL) against the colorectal cancer lines, SW480 and SW620, through induction of cell death via apoptosis. However, the bark extract demonstrated moderate activity (IC_50_ 24.5 µg/mL) against lung (A549) and cervical (HELA) carcinomas, while callus and cell suspension cultures derived from the plant showed a more specific activity against A549 lung carcinoma [23,85]. Furthermore, prenylated flavonoids isolated from the stem bark showed high inhibitory activity against MCF-7 (breast) and HepG2 (liver) cancer lines, with comophoroside A (3.7.6) showing the greatest activity (IC_50_ 8 µg/mL) [46]. The leaf extract was also reported to significantly decrease HepG2 viability at concentrations of 100 µg/mL [78].
The anticancer potential of the plant was attributed to a diverse group of phytochemicals, particularly terpenoids, flavonoids, and lignans (Table 1). In the EO, β-caryophyllene (1.2.11) served as a primary bioactive constituent, which constituted about 20% of the EO composition. It was shown to act as a selective inducer of caspase-3-dependent apoptosis in mouse lymphoma and human B-lymphocyte cell lines without affecting normal human skin [13,34]. Furthermore, comprehensive metabolite profiling of the plant suggested a group of identified lignans and alkaloids with established antineoplastic properties as contributors to cytotoxic properties, including 10-hydroxycamptothecin (8.1), justicidin B (5.4), and anthricin (deoxypodophyllotoxin) (5.1), detected in leaf, bark, and callus extracts [23,85]. Moreover, the cytotoxic potential was further supported by the flavonoid content, particularly flavonols such as quercetin (3.1.5) and myricetin (3.1.3), as well as prenylated flavonoids isolated from the stem bark [23,46]. Notably, the prenylated flavonoid comophoroside A (3.7.6) demonstrated potent inhibitory action against MCF-7 and HepG2 cell lines [46].
6.4. Antidiabetic Activity
Research on C. gileadensis revealed hypoglycemic potential, insulin level restoration, and immune system modulation to preserve pancreatic integrity. Numerous experimental models have confirmed these effects (Table 4). The leaves, twigs, and stem bark extracts demonstrated a potent hypoglycemic effect. In streptozotocin (STZ)-induced diabetic models, the butanol fraction of the stem bark significantly reduced blood glucose and α-amylase levels and increased insulin levels [45]. Similarly, the aqueous extracts of twigs and leaves were shown to significantly decrease fasting blood sugar and glycated hemoglobin (HbA1c) in alloxan-induced diabetic rats, with the twigs extract exhibiting higher potency than the leaf extract [79]. Evaluations of the antidiabetic properties of the sap and acetone extracts of the aerial parts in STZ-induced diabetic mice revealed that the sap-treated group achieved normal blood glucose levels after only 6 days of treatment, while the acetone group required 15 days [87].
The antidiabetic properties of the plant were highly attributed to its antioxidant capacity and ability to modulate inflammatory responses associated with diabetes. It enhances endogenous antioxidant defenses, such as SOD and GSH, while reducing lipid peroxidation markers, thereby protecting pancreatic β-cells from oxidative damage [45,79]. They also included inhibition of α-amylase, reducing the breakdown of carbohydrates into glucose [45]. Histologically, it was reported that the group of diabetic rats treated with C. gileadensis twig aqueous extract restored normal pancreatic tissues compared to the pancreatic tissues in the untreated group, which showed vacuolation of Langerhans’ islets [79]. Immunological parameters such as IgA, IgG, and IgM were also associated with diabetes. Diabetic patients may have altered levels of IgA, IgG, and IgM, which can affect their immune response, making them more susceptible to infections. Twigs aqueous extract was found to be effective in restoring the immunoglobulin levels [79]. Sap and aerial parts extracts were shown to reduce pro-inflammatory CD4^+^ and CD8^+^ T-lymphocyte subsets in diabetic models. Conversely, they increased the population of Treg cells, CD4^+^ CD25^+^ and CD8^+^ CD25^+^, which help maintain immune tolerance and may prevent the progressive autoimmune destruction of pancreatic β-cells [86,87].
Phytochemical studies identified several potentially bioactive metabolites that may be responsible for these antidiabetic activities. These included 2,6-deoxyfructosazine (9.2) and saponarin (3.2.6), which were identified in the leaf extract and are known to regulate blood sugar levels [40]. Other potential bioactive compounds included phenolics and flavonoids, quercetin (3.1.5), gallic acid (4.6), caffeic acid (4.1), and naringenin (3.3.6), which were identified in the bioactive fractions and are linked to α-amylase inhibition and antioxidant protection [45]. In addition to the previously mentioned antioxidant compounds, which are believed to play a role in mitigating oxidative stress, these compounds may also attenuate the induced progression of diabetes [34,78].
6.5. Wound Healing Activity
The wound healing process is a physiological response initiated after skin injury, involving successive and overlapping functions, including haemostasis (stopping bleeding), inflammation, cell proliferation and maturation, and remodeling, mediated through several cytokines, chemical mediators, and secretions from different types of cells [99]. Studies of the wound healing properties of C. gileadensis (Table 4) showed that it accelerated tissue regeneration, exhibited potent antimicrobial action at the injury site, and exerted significant anti-inflammatory modulation [36,49]. Topical application of the aerial parts (leaves, branches, fresh stems, and EO) extract significantly enhanced the rate of wound reduction and shortened the epithelialization period in both infected (S. aureus) and non-infected excision models [36,49]. Histopathological evaluations demonstrated that the extracts reduced inflammatory cell infiltration, promoted earlier collagen fiber deposition, and stimulated rapid re-epithelization and granulation tissue formation [36,49]. These effects were attributed to a high ceramide content of (69% of methanolic extract), which was able to restore skin barrier function. Additionally, the wound healing potential was supported by the terpenoid compounds such as β-caryophyllene (1.2.11), β-pinene (1.1.6), and α-pinene (1.1.5), which possess recognized antimicrobial and re-epithelization-inducing properties [36,49]. Furthermore, the sap was shown to block the lectin-dependent adhesion of bacterial pathogens, such as Pseudomonas aeruginosa, preventing biofilm-related complications that generally delay healing [36,69]. Traditional application in Hadhramout supported its wound healing properties, as ~90% of local users reported pronounced improvement of burn wounds treated with the plant [30].
6.6. Cardio-Protective and Vasorelaxant Activities
Diabetes raises the oxidative state, which results in several consequences, including cardiovascular effects. A high level of serum triglyceride and LDL cholesterol or low level of HDL cholesterol is linked with fatty depositions within the walls of arteries, with increased risk of heart attack and stroke.
Evaluations of the cardio-protective and vasorelaxant activities of C. gileadensis (Table 4) revealed the presence of multiple mechanisms, involving the activation of muscarinic receptors, modulation of vascular signaling pathways, and the induction of cardiovascular integrity markers [38,71,87]. In a study, the aerial part (branches) extract produced immediate, dose-related hypotensive and bradycardiac effects in rats through the activation of muscarinic cholinergic receptors, a response that was effectively inhibited by atropine sulfate [71]. At a molecular level, using a bio-guided assay, the diterpene resin acids, dehydroabietic acid (1.3.1) and sandaracopimaric acid (1.3.5), induced concentration-dependent relaxation of the phenylephrine (PE)-contracted pulmonary artery [38]. In particular, 1.3.1 was shown to enhance nitric oxide (NO) production via the PI3K/Akt-eNOS signaling pathway [38]. In diabetic models, treatment with the sap, methanol, or acetone extracts of the leaves and branches significantly improved cardiovascular health by inducing positive integrity markers such as adropin and NO and reducing malfunction markers including endothelin-1 (ET-1), vascular endothelial growth factor (VEGF), and cardiac enzymes like creatine kinase-MB (CK-MB) and lactate dehydrogenase (LD) [86]. These cardioprotective effects are further supported by a reduction in pro-inflammatory T-cells and an increase in Treg cells, which help prevent atheroma formation [86]. Additionally, the reported antioxidant properties of the plant further support these properties through mitigating oxidative stress-induced cardiovascular damage. For instance, the aerial parts extract inhibited xanthine oxidase (XO) and showed potent antioxidant activity, suggesting potential in preventing oxidative ischemic injury [11,86].
6.7. Antihyperlipidemic Activity
Hyperlipidemia is a pathological condition characterized by elevated levels of serum lipids due to genetic or lifestyle factors, representing a major risk factor for cardiovascular diseases. Diabetes further exaggerates this risk by disrupting lipid metabolism, resulting in increased total cholesterol (TC), triglycerides (TG), and low-density lipoprotein (LDL), and decreased high-density lipoprotein (HDL, good cholesterol) levels [100]. Recent evidence indicates that oxidative stress may contribute to the pathophysiology of hyperlipidemia [101].
Studies on the antihyperlipidemic potential of C. gileadensis showed its significant capacity to normalize lipid profiles in metabolic disorder models through reducing TC, TG, and LDL and augmenting HDL levels [45,79,86]. In hypercholesterolemic diabetic rats, aqueous extracts of the twigs and leaves effectively lowered TC, TG, and LDL, with the twigs extract demonstrating a notably higher therapeutic efficiency in restoring these parameters to near-normal levels [79]. This efficacy was further supported by the butanol fraction of the stem bark, which was shown to reduce cholesterol levels by enhancing the catabolism of LDL-C and its subsequent elimination as bile acids [45]. Guggulsterone (2.1), a key bioactive sterol in the leaves, was shown to act as an antagonist of the farnesoid X receptor (FXR), the primary bile acid receptor, thereby modulating systemic cholesterol metabolism and downregulating the bile salt export pump [44,102]. Furthermore, the sap and the aerial parts extract demonstrated significant hypolipidemic effects in Type 1 diabetic mice, with the sap demonstrating superior efficacy [86].
6.8. Gastroprotective and Anti-Ulcer Activities
C. gileadensis was shown to have gastroprotective and anti-ulcer properties through a dual mechanism, involving inhibition of gastric secretions and reinforcing the mucosal defensive barrier [74]. The aerial parts extract, administered at doses of 250 and 500 mg/kg, showed significant dose-dependent protection against gastric lesions induced by necrotizing agents (80% ethanol, 0.2 M NaOH, and 25% NaCl), hypothermic restraint stress, and indomethacin [74]. Histopathological evaluations demonstrated that pretreatment with the extract preserved the cytoarchitecture of gastric mucosa, preventing hemorrhagic necrosis and inflammatory cell infiltration. The extract was found to replenish depleted stomach wall mucus and maintain nonprotein sulfhydryl (NP-SH) concentrations, which potentially enhanced prostaglandin synthesis [74]. These effects are primarily attributed to bioactive constituents, such as flavonoids, saponins, and terpenoids (Table 1), through the prevention of free radicals and lipid peroxidation resulting from gastric mucosal damage [74,103,104]. Traditional usage surveys in Hadhramaut supported these gastroprotective properties, with local populations reporting the effective use of the plant for treating stomach ulcers and improving digestion [30,74].
6.9. Hepatoprotective Activity
Oxidative stress from toxins, infections, or diet disrupts hepatic homeostasis by damaging DNA, lipids, and proteins, initiating inflammatory pathways that exacerbate liver injury [105]. Hepatoprotective agents function by minimizing these harmful effects or restoring physiological mechanisms disturbed by hepatotoxins [76].
Promising results were reported by several studies, showing the ability of C. gileadensis to mitigate chemical-induced injury, strengthen antioxidant defense, and restore hepatic architecture [72,73,79]. The aerial parts and bark extracts showed significant protection against CCl_4_ and diethylnitrosamine (DEN)-induced-liver damage, as demonstrated by a marked reduction in serum AST, ALT, ALP, and bilirubin levels [72,73]. The reported hepatoprotective mechanisms involved free radical scavenging, reduction in MDA levels, and restoring endogenous antioxidants, including nonprotein sulfhydryls (NP-SH), GSH, and SOD [45,73,79]. Histopathological evaluations showed that the plant prevented confluent necrosis, steatosis, and inflammatory cell infiltration, maintaining the structural integrity of hepatocytes [72,73,79]. These therapeutic benefits are attributed to the rich phytochemical content of C. gileadensis, flavonoids, and volatile terpenes, which exhibit antioxidant activities promoting hepatocellular recovery [11,31,40].
6.10. Fertility-Enhancing Activity
In a study by Alhazmi (2025), C. gileadensis was shown to ameliorate infertility and erectile dysfunction in STZ-induced diabetic male mice [75]. The oral administration of the sap, methanol, and acetone extracts significantly reversed reproductive decline by significantly restoring testicular weight and increasing testosterone levels in diabetic and control male mice up to 70-fold and 30-fold, respectively, leaving FSH and LH unchanged, indicating a hypothalamic pituitary independent effect. The extracts also elevated the sperm count in addition to motility and percentage of normal sperm morphology. The sap remarkably showed a pronounced increase in testosterone level and sperm count. The plant was shown to exert these effects through the induction of positive erectile markers, such as NO and adropin, and suppression of the vasoconstrictor (negative erectile marker), endothelin [75]. It also reduced CD8^+^ and CD4^+^, which may decrease testes inflammation and induce its recovery in diabetic mice. Furthermore, the extracts restored testicular architecture and enhanced nitric oxide synthase (NOS) immunoreactivity in the testes. These therapeutic outcomes are attributed to the high steroid content and the presence of antioxidant and anti-inflammatory terpenes, which mitigated oxidative damage by reducing lipid peroxidation (LPO) and replenishing GSH, GSH-Px, and SOD levels in testicular tissue [75]. The steroidal derivative, guggulsterone (2.1), was reported to have multiple interactions, including an antagonizing effect on FXR and modulating the activity of other steroid receptors, such as estrogen receptor alpha, progesterone receptor, and pregnane X receptor (PXR) [106,107]. These interactions with key receptors involved in metabolic, reproductive, and steroidal hormone modulations can potentially influence fertility processes.
6.11. Antigenotoxic Activity
Genotoxicity refers to the capacity of physical, chemical, or biological agents to damage DNA, causing permanent and transmissible genetic alterations such as gene mutations and chromosomal abnormalities, which may lead to cancer, heritable defects, neuromuscular and neurodegenerative diseases, immune deficiencies, cardiovascular diseases, metabolic syndrome, aging, and infertility. Anti-genotoxic agents are substances capable of protecting DNA from damage or lessening existing harm [108,109].
Reported data (Table 4) showed that C. gileadensis could protect against chemical-induced damage, such as CCL_4_-induced genotoxicity and metabolic stress [32,45]. The dichloromethane extract reduced the proportion of chromosomal abnormalities in bone marrow cells and DNA fragmentation in hepatocytes [32]. This protective effect extended to the reproductive system, where the extract significantly lowered the frequency of sperm shape anomalies and chromosomal irregularities in spermatocytes [32]. In STZ-induced diabetic rats using the Comet assay, the butanol fraction of stem bark extract (100 mg/kg) reduced tailed DNA, associated with diabetic complications, from 13.00% to 11.00% and restored healthy untailed DNA to 89.00. Reactive oxygen species (ROS) affect DNA metabolism, leading to increased oxidative stress and inflammatory cascade activation that cause organ abnormalities. Therefore, antioxidants would protect the DNA structure [45]. Polyphenolic compounds were reported to have protective effects against DNA damage by reducing ROS and modulating enzymes responsible for the bioactivation of genotoxic agents. Free hydroxyl groups on the B ring (catechol moiety) and C-3 position of the C ring are important structural features for their antigenotoxic activity [108,110].
6.12. Immunomodulatory Activity
Immune reaction plays a critical role in inflammatory and atherosclerotic heart diseases. The immunomodulatory activity of C. gileadensis was mainly attributed to its ability to modulate cellular and humoral immune responses, as demonstrated in models of metabolic stress [79,86,87]. The sap and extracts of aerial parts modulated T-cell populations in diabetic mice, reducing the total lymphocytes and CD3^+^, CD4^+^, and CD8^+^ T-cell subsets, while increasing the Treg CD4^+^ CD25^+^ and CD8^+^ CD25^+^ cell subsets, which help regulate inflammatory cascades [86,87]. Furthermore, the aqueous extracts of the leaves and twigs were shown to restore humoral balance in diabetic rats by normalizing elevated immunoglobulins IgA, IgE, IgG, and IgM levels [79].
Plant polyphenols are recognized for their immunoregulatory effects, achieved by modulating immune cell functions and suppressing proinflammatory cytokine production and immune-related gene expression [111].
6.13. Antiaging Activity
Aging is a progressive decline in the organism’s ability to withstand stress, resulting in a gradual loss of physiological functions. It is driven largely by the accumulation of molecular damage in DNA, proteins, and lipids. Because oxidative reactions rise as intracellular ROS-scavenging capacity declines, antioxidants are thought to slow aging and may contribute to lifespan extension [78]. The anti-aging activity of C. gileadensis (Table 4) was evidenced by its capacity to extend the replicative lifespan of cellular models and mitigate oxidative stress [78]. The leaf extract (30 µg/mL), obtained by both ultrasonic-assisted and hydrodistillation methods, was shown to significantly increase the average lifespan of the K6001 yeast strain from 7.55 to 9.15 generations [78]. These longevity-promoting effects were attributed to the high terpenoid and phenolic contents of the plant, which function as powerful antioxidants that stabilize free radicals, protecting cells from oxidative damage [78]. The terpenic EO constituents of C. gileadensis, including α-pinene (1.1.5) and β-caryophyllene (1.2.11), are known for their antioxidant and anti-inflammatory activities, which may explain their potential geroprotective effects [11,78,112]. In particular, β-caryophyllene (1.2.11) was shown to extend lifespan in Caenorhabditis elegans by over 22%, primarily through reducing oxidative stress and modulating key stress-response genes including SIR-2.1, SKN-1, and DAF-16 [112]. Furthermore, the presence of flavonoids such as quercetin (3.1.5), which was shown to ameliorate aging processes, provided a complementary mechanism for cellular protection [113].
6.14. Analgesic and Antipyretic Activities
The extract of C. gileadensis showed promising analgesic and antipyretic properties (Table 4). In rodent (rats and mice) models, the aerial parts of the plant demonstrated potent dose-dependent peripheral and central analgesic effects. It significantly reduced acetic acid-induced writhing and increased latency in tail-flick and hot-plate tests [76,82]. At a 500 mg/kg dose, it showed superior potency compared to diclofenac, with 100% inhibition of writhing and 93.57% reduction in late-phase formalin-induced paw licking [82]. It also showed an ability to decrease yeast-induced hyperthermia in a dose-and-time dependent manner, indicating antipyretic action [76]. Moreover, a combination of the aerial parts extract (125 mg/kg) with a sub-therapeutic dose of the NSAID, diclofenac (12.5 mg/kg), resulted in a synergistic potentiation of the analgesic effect [82].
These therapeutic properties were mainly attributed to the triterpenes and flavonoids content of C. gileadensis [114,115], in particular, the sesquiterpene β-caryophyllene (1.2.11), which was recognized for its analgesic and anti-inflammatory potential [34].
6.15. Diuretic, Kidney Protective, and Antihyperuricemic Activities
The plant was traditionally utilized to treat urinary retention and promote the expulsion of renal calculi by increasing urine flow, in addition to its nephroprotective and antihyperurecemic effects, supported by pharmacological investigations, indicating potential diuretic and kidney protective activities (Table 4) [11,71,74]. The aerial parts extract was shown to significantly increase urine volume in a dose-dependent manner (22% to 31% increase) without significantly altering the excretion of Na^+^, K^+,^ and Ca^2+^ ions. This effect was potentially attributed to the inhibition of antidiuretic hormone (ADH) or blocking its receptors [76]. Beyond this diuretic action, nephroprotective effects were observed in metabolic stress in vivo models. Treatment with sap and aerial parts extracts reduced elevated serum urea, creatinine, and uric acid levels in diabetic and hypercholesterolemic models, as well as their ability to ameliorate histopathological damage, such as glomerular atrophy and tubular vacuolation, restoring near-normal renal architecture [76,79]. Since blood urea and serum creatinine are considered early biomarkers of diabetic nephropathy [116], the observed reductions in these parameters in diabetic mice suggest that C. gileadensis may exert renal protective effects and could slow the progression of diabetic nephropathy.
High levels of uric acid are closely linked to oxidative stress and the generation of ROS, a condition in which xanthine oxidase (XO) acts to convert xanthine to uric acid. Thus, reduced oxidative stress can improve urate handling and decrease tissue damage [11]. Although the aerial parts extract demonstrated weak in vitro inhibition of XO (IC_50_ = 251.2 µg/mL) compared to allopurinol (IC_50_ = 0.41 µg/mL); however, the involvement of multiple mechanisms, including antioxidant (DPPH and BCB assays), XO and COX-1 inhibitory activity, and inhibition of protein denaturation, could synergistically contribute to its nephroprotective effects [11]. These multiple mechanisms may provide scientific evidence for its use in treating gouty arthritis [17].
The nephroprotective activities are mostly attributed to a variety of phenolic and flavonoid compounds (Table 1), which act as powerful antioxidants, such as caffeic acid phenethyl ester (4.2), hesperetin (3.3.2), hesperidin (3.3.3), chrysin (3.2.2), gallic acid (4.6), rutin (3.1.6), and caffeic acid (4.1) [11]. They are also potent inhibitors of enzymes, such as cyclooxygenase (COX) and lipoxygenase (LOX), that control inflammation, a symptom characterizing gout and gouty arthritis [11].
6.16. Anticoagulant Activity
The sap, as well as the methanol and acetone extracts of branches and leaves of C. gileadensis, exhibited anticoagulative potential by significantly prolonging prothrombin time (PT), activated partial thromboplastin time (aPTT), and INR, which was more effective than heparin or aspirin. The observed effect was attributed to high levels of glycosaminoglycans [88].
Oxidative stress is considered an important factor causing thrombosis, due to the overproduction of reactive oxygen species (ROS) that can significantly impair the function of vascular endothelial cells, platelets, and red blood cells, leading to a cascade of events accelerating the formation of thrombi. Polyphenols and terpenes were reported to interfere with ROS-mediated platelet activation in thrombosis [117,118,119]. Accordingly, the anticoagulant activity of C. gileadensis may also be explained by the polyphenols and terpenes contents, which represent its main composition.
6.17. Antibacterial and Antibiofilm Activities
Numerous studies documented the in vitro and in vivo antibacterial and antibiofilm activities of different extract types, as well as EO of C. gileadensis, against a wide range of Gram-positive (Gram +ve) and Gram-negative (Gram −ve) bacteria, including multidrug-resistant pathogens, such as methicillin-resistant Staphylococcus aureus (MRSA), P. aeruginosa, and Klebsiella pneumoniae (Table 4) [23,33,39,49,70,80,81,85,89,90].
The bark and leaves extracts exhibited significant antibacterial activity against a wide spectrum of pathogens, including S. aureus, P. aeruginosa, and K. pneumoniae [23,85]. Notably, the antibacterial activity against S. aureus was shown to be superior to standard antibiotics, such as ampicillin [85]. Interestingly, bioactivity-guided phytochemical investigations of fresh stems identified a group of ent-verticillane-type diterpenoids, in particular, (9S,10S)-ent-9,10-epoxyverticillol (1.3.3), as the main antimicrobial agents against K. pneumoniae, potentially due to their ability to penetrate bacterial cell walls [39]. Furthermore, the EO demonstrated high antibacterial activity against Bacillus subtilis and S. aureus, which was attributed to potential bioactive compounds, such as β-pinene (1.1.6), β-caryophyllene (1.2.11), and terpinen-4-ol (1.1.10) [23,33,120]. Additionally, extracts derived from cell suspension cultures showed selective antibacterial action against Staphylococcus epidermidis [23]. Maqlam and Bin Kardous [70] developed two topical formulations: a cream and a gel, containing 10% of the bark extract. The gel formulation exhibited greater antibacterial activity against S. aureus, a common skin pathogen (15.7 mm inhibition zone diameter, IZD), compared to both the plant extract cream (IZD = 13.3 mm) and the standard cetrimide cream (IZD = 13.3 mm). This increased efficacy was attributed to the lower viscosity of the gel vehicle, which facilitated the release and diffusion of phenolic compounds into the surrounding medium compared to the cream base [70]. Moreover, a synergistic effect was reported between C. gileadensis extracts and some antibiotics, especially amoxicillin, polymixin B, and tetracycline [81].
Microorganisms use biofilm formation as a survival strategy to stick to surfaces and develop into organized communities covered in an extracellular polymeric matrix that they produce on their own. Compared to planktonic cells, this matrix confers significantly greater resistance to antibiotics and host immune defenses, protecting bacteria and encouraging chronic and persistent infections [121]. C. gileadensis demonstrated strong antibiofilm activities through interfering with bacterial cell adhesion and significantly reducing exopolysaccharide (EPS) content, which is a critical factor in biofilm stability [81]. The shoot extracts were shown to decrease EPS levels in K. pneumoniae by 39% and inhibit biofilm formation in multidrug-resistant strains of Acinetobacter baumannii [81]. Furthermore, the aqueous extract of branches (1 mg/mL) significantly killed more oral anaerobic biofilm-forming multispecies bacteria (unspecified), obtained from biofilms of subgingival and supragingival plaque from human donors, than 2% chlorhexidine [90]. Although the experiment utilized unknown bacterial isolates from plaque, the literature review of the thesis mentioned various bacterial strains typically associated with such infections, including Enterococcus faecalis, Porphyromonas gingivalis, Fusobacterium nucleatum, and Streptococcus mutans [90]. The major essential oil component, terpinene-4-ol (1.1.10), was identified as a potent antibiofilm agent against S. aureus, which was capable of preventing biofilm formation at sub-inhibitory concentrations and disrupting established biofilms, potentially through targeting penicillin-binding protein 2a [120].
6.18. Antiviral Activity
The leaf extract of C. gileadensis demonstrated potent and selective antiviral and virucidal activity against enveloped viruses, including herpes simplex virus type 2 (HSV-2) and respiratory syncytial virus type B (RSV-B). However, it was inactive against non-enveloped viruses, such as coxsackievirus B type 3 (CVB-3) and adenovirus type 5 (ADV-5) (Table 4) [44]. The suggested mechanism involved direct interaction between bioactive compounds in the extracts with receptor proteins on the viral envelope, inhibiting the virus’s ability to bind to host cells, eliminating its ability to cause infection. This activity was primarily attributed to the sterol guggulsterone (2.1), which was isolated from the leaf extract by TLC using a bio-guided method, and identified through HPLC-diode array (PDA) combined with electrospray ionization mass spectrometry (ESI-MS) [44]. Although the leaf extract exhibited selective antiviral properties, research evaluating the bark extract of C. gileadensis showed that it was ineffective as antiviral against a group of viruses, including hepatitis A virus (HAV-0), coxsackievirus, HSV-1, and HSV-2 [91].
Molecular docking and dynamics simulation studies were performed on guggulsterone against the dengue virus, targeting four proteins, including NS5 RNA-dependent RNA polymerase, dengue methyltransferase, NS3 protease-helicase, and dengue virus type 2 envelope glycoprotein. The virtual findings suggested that guggulsterone may be considered as a potent inhibitor of the dengue virus proteins and may be a significant inhibitor of the major envelope protein E of the dengue virus [102]. Notably, (-)-galbelgin (5.3), a lignan isolated from Piper kadsura (Choisy) Ohwi and detected in leaves of C. gileadensis was also reported as anti-human hepatitis B virus [122].
6.19. Antifungal Activity
Candidiasis has become the most common opportunistic invasive mycosis in the world due to its acute progression, difficulty in diagnosis, and high mortality rates, especially in immunocompromised individuals [85]. Although Candida albicans is commonly found in secondary endodontic infections at rates of up to 18%, the emergence of non-albicans species that show intrinsic resistance to traditional antifungal agents is increasingly impeding the success of treatment [85,90]. C. gileadensis has historically been used as an antiseptic to treat infected wounds. Several studies documented that extracts from different parts of this plant demonstrated broad-spectrum antifungal activity against various pathogenic yeasts and molds, including C. albicans, C. glabrata, C. krusei, Cryptococcus neoformans, and Aspergillus niger (Table 4) [33,37,85,89]. The leaf extracts effectively inhibited the growth of C. albicans, while the bark extracts remarkably demonstrated effectiveness against multidrug-resistant Candida species that were less sensitive to standard antifungals, such as itraconazole and voriconazole [23,85].
Phytochemical profiling of leaf extracts tentatively identified sordarin (1.3.6), an antifungal tetracyclic diterpenoid bearing a norbornene system, that was reported to inhibit fungal protein synthesis by targeting eukaryotic translation elongation factor 2 [40,123,124]. High levels of condensed tannins (proanthocyanidins) in the stem peels were believed to play a protective role against fungal pathogens by serving as a chemical defense mechanism [80]. It is worth noting that no specific studies were found describing the detection of proanthocyanidins in C. gileadensis. However, the occurrence of proanthocyanidins was reported in the decoction of C. leptophloeos bark [125,126]. Additionally, the EO contributed to the antifungal properties of C. gileadensis by selectively inhibiting C. glabrata and C. krusei [33]. Abbas, et al. [42] reported that syringic acid (4.12) isolated from the aerial parts of C. gileadensis exhibited moderate anticandidal activity, along with moderate antimalarial and antimycobacterial activities.
The pharmacological activities of C. gileadensi are summarized in Figure 19.
7. Pharmacokinetic and Toxicological Profiles of C. gileadensis
Although C. gileadensis was shown to have broad-spectrum biological activities and a rich phytochemical profile, making it a promising medicinal plant, little is known about its pharmacokinetic and toxicological properties. Even though a long history of traditional use suggests a favorable safety profile, there is a significant gap in the current literature, by focusing on the phytochemical composition and biological activities, with little presentation of thorough ADME and toxicological studies.
Reviewed publications either conducted sub-acute toxicological studies on various C. gileadensis extracts and administration doses and routes, with the oral route being the most popular, or preliminary acute toxicity tests with median lethal dose (LD_50_) determination in some cases. A summary of the reviewed toxicological studies is provided in Table 5.
C. gileadensis was generally categorized as practically non-toxic, and the reported acute oral lethal doses (LD_50_) were determined to be >2000 mg/kg in rats and >5000 mg/kg in mice [85,127]. Although single doses are well-tolerated, high-range doses (2500–5000 mg/kg) were associated with reversible sedation and a reduction in motor activity, possibly due to CNS-depressant effects of the high volatile oil content [85,127]. In a study evaluating the sub-acute toxicological effects (14 days at 1000 mg/kg/day), the extract did not adversely affect renal or hepatic function markers; however, significant dose-dependent reductions in hematological parameters (RBC, HCT, HGB, and WBC) suggested potential bone marrow inhibition on prolonged administration [127].
The ADME and toxicity profiles of the major bioactive sterol, guggulsterone (2.1), were predicted through in silico assessments [102]. Generally, the compound was concluded to be non-toxic, exhibiting no predicted hepatotoxic, immunotoxic, carcinogenic, cytotoxic, or mutagenic effects. It demonstrated high gastrointestinal absorption and blood–brain barrier (BBB) permeability. It was not a substrate for P-glycoprotein and exhibited a bioavailability of >40% in rats with a half-life of approximately 10 h [44,102]. It was predicted to inhibit CYP2C19 and CYP2C9 enzymes, with no inhibitory activity toward CYP1A2, CYP2D6, or CYP3A4 [102].
Another bioactive constituent, β-caryophyllene (1.2.11), was shown to selectively induce apoptosis in tumor cells without affecting normal human fibroblasts, indicating a good therapeutic window for medicinal applications [13,34].
8. Conclusions
C. gileadensis, historically known as the “Balm of Gilead,” possesses a remarkably diverse metabolic profile comprising over 170 secondary compounds, including terpenoids, flavonoids, and phenolic acids, among others, validating its traditional use, particularly as an effective antioxidant, anti-inflammatory, antimicrobial, and anticancer agent. However, because most current phytochemical evidence relies on tentative identifications from hyphenated techniques, such as GC-MS and LC-MS, future research must prioritize the phytochemical isolation and full structural elucidation of pure bioactive constituents to enable robust structure-activity relationship (SAR) analyses. Furthermore, scientific efforts should shift toward identifying specific molecular targets through detailed mechanistic studies and addressing critical gaps in the pharmacokinetic behavior (ADME) and long-term toxicological profile of the plant through thorough in vivo investigations. Therefore, the successful integration of C. gileadensis into modern evidence-based medicine requires a transition to human clinical trials for establishing safety and efficacy protocols. In addition, the expanded use of biotechnological methods, such as plant tissue culture, is essential to ensure a sustainable and standardized supply of bioactive metabolites, while safeguarding the species from ecological threats associated with overharvesting and climate change.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Pawase P.A. Goswami C. Shams R. Pandey V.K. Tripathi A. Rustagi S. GD. A conceptual review on classification, extraction, bioactive potential and role of phytochemicals in human health Future Foods 2024910031310.1016/j.fufo.2024.100313 · doi ↗
- 2Kumar A. PN. Kumar M. Jose A. Tomer V. Oz E. Proestos C. Zeng M. Elobeid T. KS. Major Phytochemicals: Recent Advances in Health Benefits and Extraction Method Molecules 20232888710.3390/molecules 2802088736677944 PMC 9862941 · doi ↗ · pubmed ↗
- 3Monjotin N. Amiot M.J. Fleurentin J. Morel J.M. Raynal S. Clinical Evidence of the Benefits of Phytonutrients in Human Healthcare Nutrients 202214171210.3390/nu 1409171235565680 PMC 9102588 · doi ↗ · pubmed ↗
- 4Yang Y. Sun X. Peng C. Wei J. Yang X. The Genus Commiphora: An Overview of Its Traditional Uses, Phytochemistry, Pharmacology, and Quality Control Pharmaceuticals 202417152410.3390/ph 1711152439598434 PMC 11597752 · doi ↗ · pubmed ↗
- 5Batiha G.E. Wasef L. Teibo J.O. Shaheen H.M. Zakariya A.M. Akinfe O.A. Teibo T.K.A. Al-Kuraishy H.M. Al-Garbee A.I. Alexiou A. Commiphora myrrh: A phytochemical and pharmacological update Naunyn Schmiedebergs Arch. Pharmacol.202339640542010.1007/s 00210-022-02325-036399185 PMC 9672555 · doi ↗ · pubmed ↗
- 6Dekebo A. Juniedi S. Hu X. Jung C. Ethnobotany, Chemistry, and Biological Activities of Some Commiphora Species Resins Gums, Resins and Latexes of Plant Origin: Chemistry, Biological Activities and Uses Murthy H.N. Springer International Publishing Cham, Switzerland 2021127
- 7Sallon S. Solowey E. Gostel M.R. Egli M. Flematti G.R. Bohman B. Schaeffer P. Adam P. Weeks A. Characterization and analysis of a Commiphora species germinated from an ancient seed suggests a possible connection to a species mentioned in the Bible Commun. Biol.20247110910.1038/s 42003-024-06721-539256474 PMC 11387840 · doi ↗ · pubmed ↗
- 8Garang Z. Feng Q. Luo R. La M. Zhang J. Wu L. Wang Z. Zeweng Y. Jiangyong S. Commiphora mukul (Hook. ex Stocks) Engl.: Historical records, application rules, phytochemistry, pharmacology, clinical research, and adverse reaction J. Ethnopharmacol.202331711671710.1016/j.jep.2023.11671737301302 · doi ↗ · pubmed ↗