Tibetan Fritillaria cirrhosa D. Don Extract Ameliorates DSS-Induced Ulcerative Colitis by Repairing Damage to the Intestinal Mucosal Barrier and Regulating the Gut Microbiota
Zhengyang Hao, Xiangjun Chen, Qing Peng, Ruipeng Wu, Huan Zhang, Ping Yin, Xuanfu Yu, Shaokang Wang

TL;DR
Tibetan Fritillaria cirrhosa extract helps treat ulcerative colitis in mice by repairing the gut lining and balancing gut bacteria.
Contribution
The study shows FCD extract's novel therapeutic potential for UC through mucosal repair and microbiota modulation.
Findings
FCD reduced colon shortening and disease activity in DSS-induced colitis mice.
FCD enhanced intestinal barrier proteins like ZO-1 and occludin and increased IL-10 levels.
FCD promoted gut microbiota diversity and reduced pro-inflammatory cytokines.
Abstract
Background/Objectives: Ulcerative Colitis (UC) is a chronic inflammatory disease of the colon that profoundly impacts human health. Conventional pharmacological treatments are associated with serious adverse reactions and toxic side effects. Consequently, the development of natural plant-derived biological agents for UC treatment is an urgent imperative. Methods: Utilising a Dextran Sulfate Sodium (DSS)-induced ulcerative colitis mouse model, with mice receiving low, medium, and high doses of water extract of Tibetan Fritillaria cirrhosa D. Don extract (FCD), alongside a group receiving 5-aminosalicylic acid. The Disease Activity Index (DAI) was calculated, colon length was measured, histological scores were assessed, and histopathological alterations were evaluated. Inflammatory factor were determined by ELISA; mRNA and protein expression in colonic tissue was analysed by RT-qPCR and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
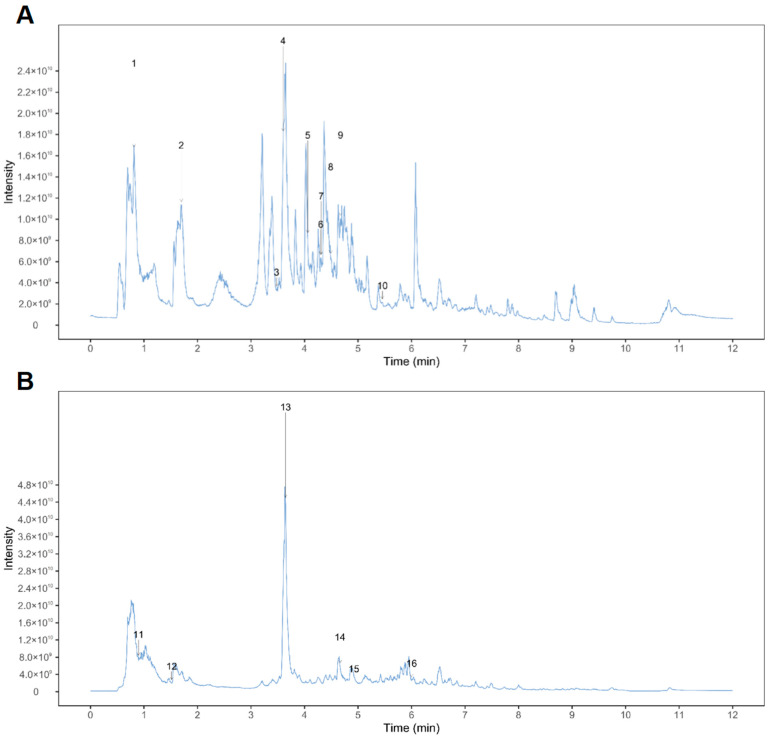 Figure 1
Figure 1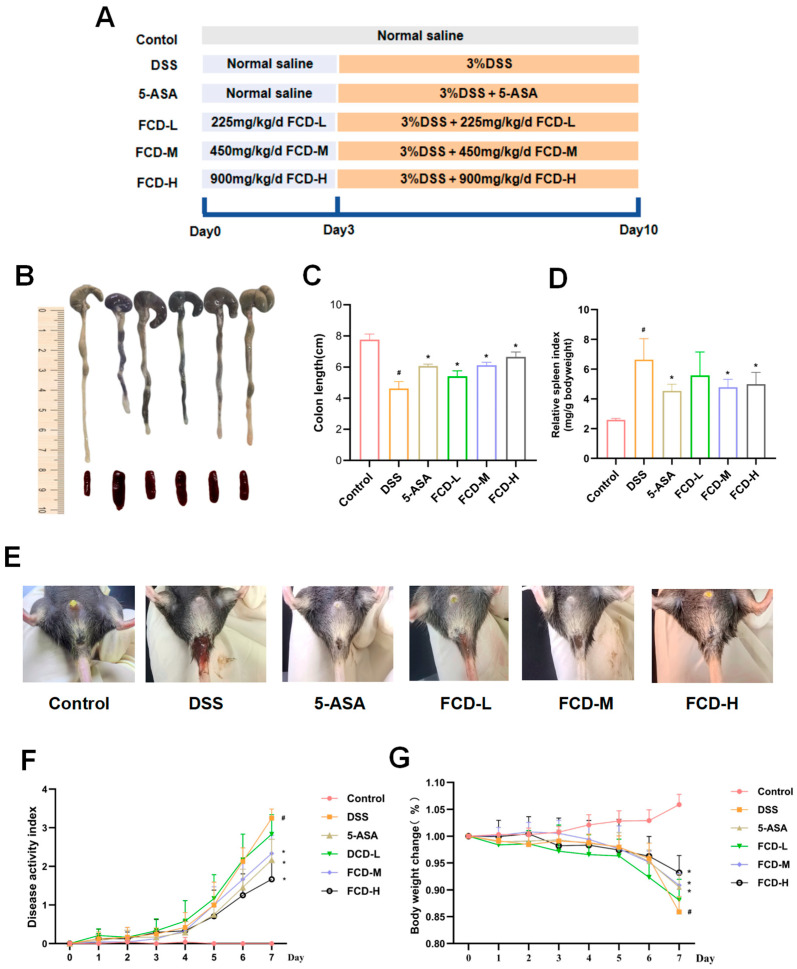 Figure 2
Figure 2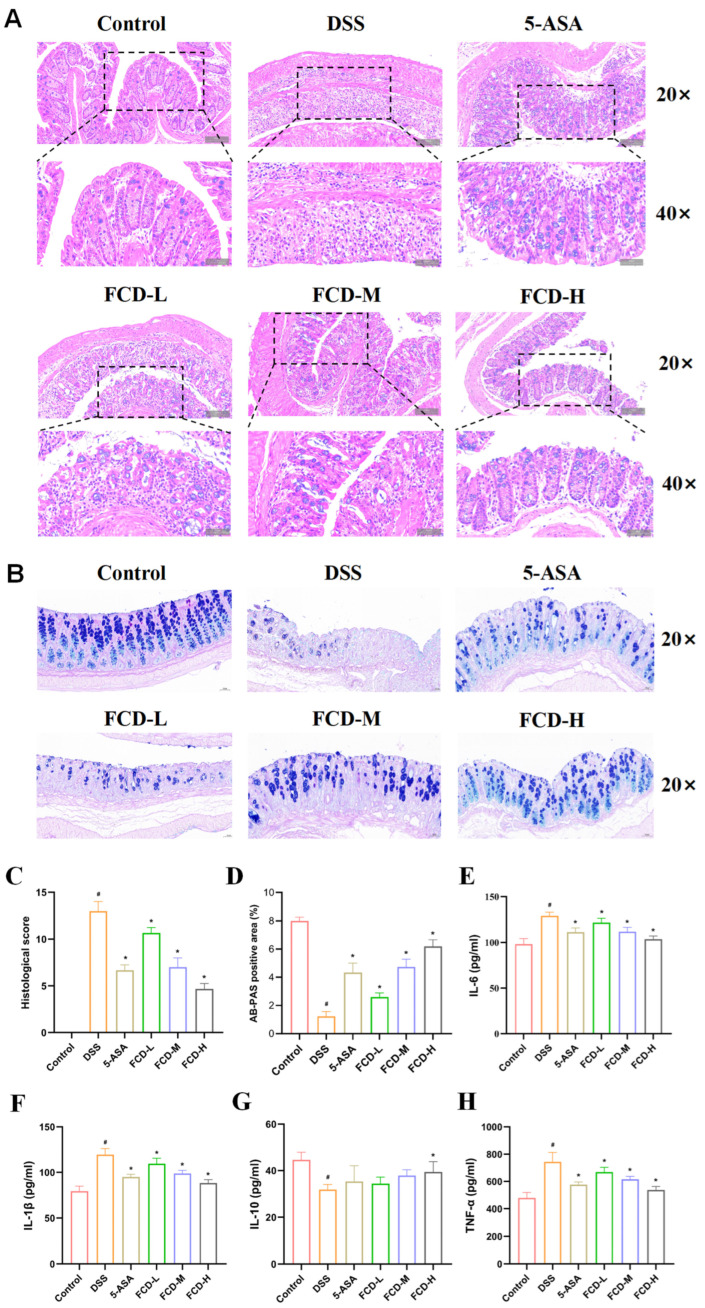 Figure 3
Figure 3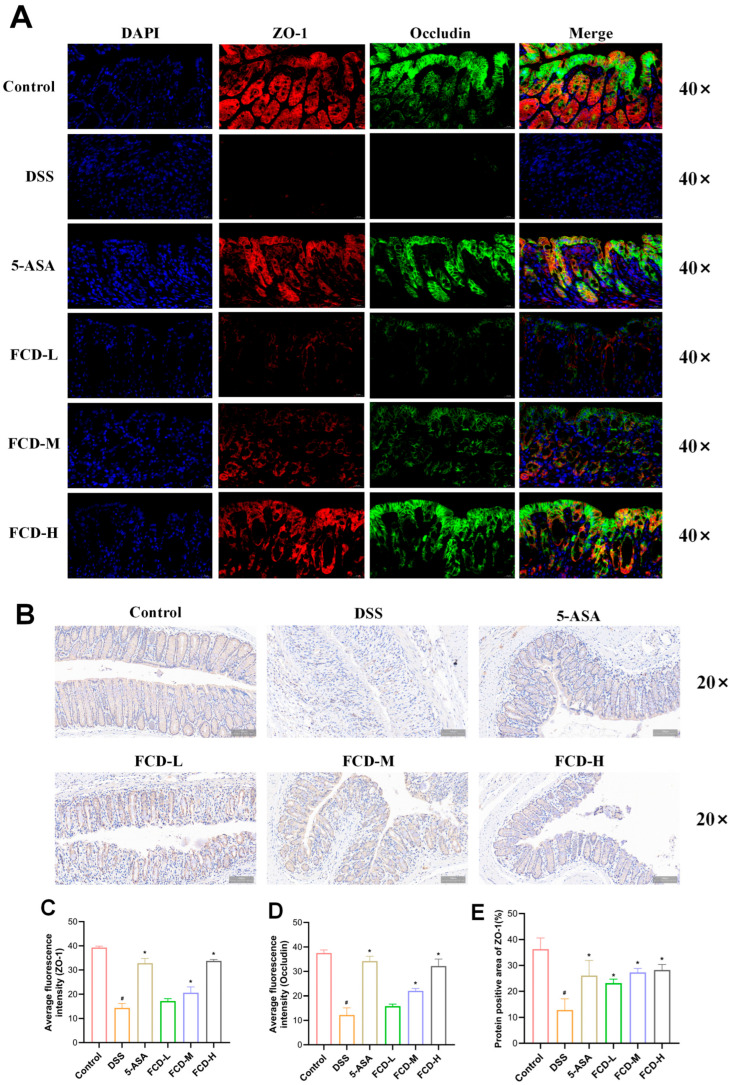 Figure 4
Figure 4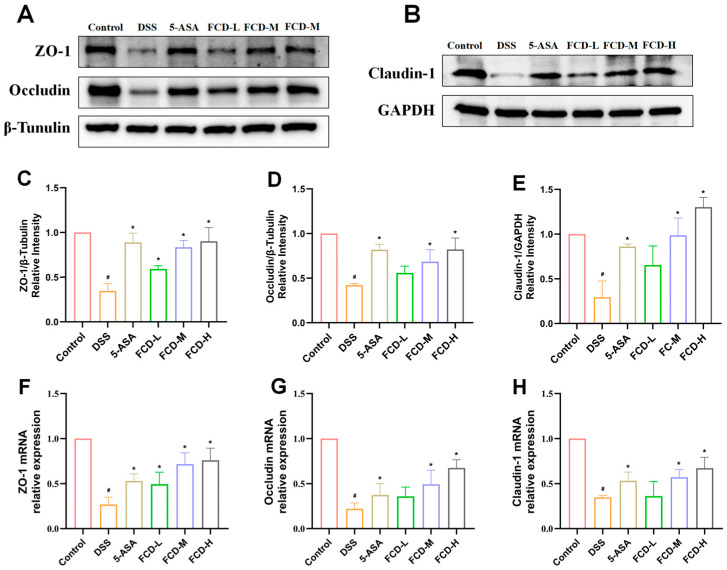 Figure 5
Figure 5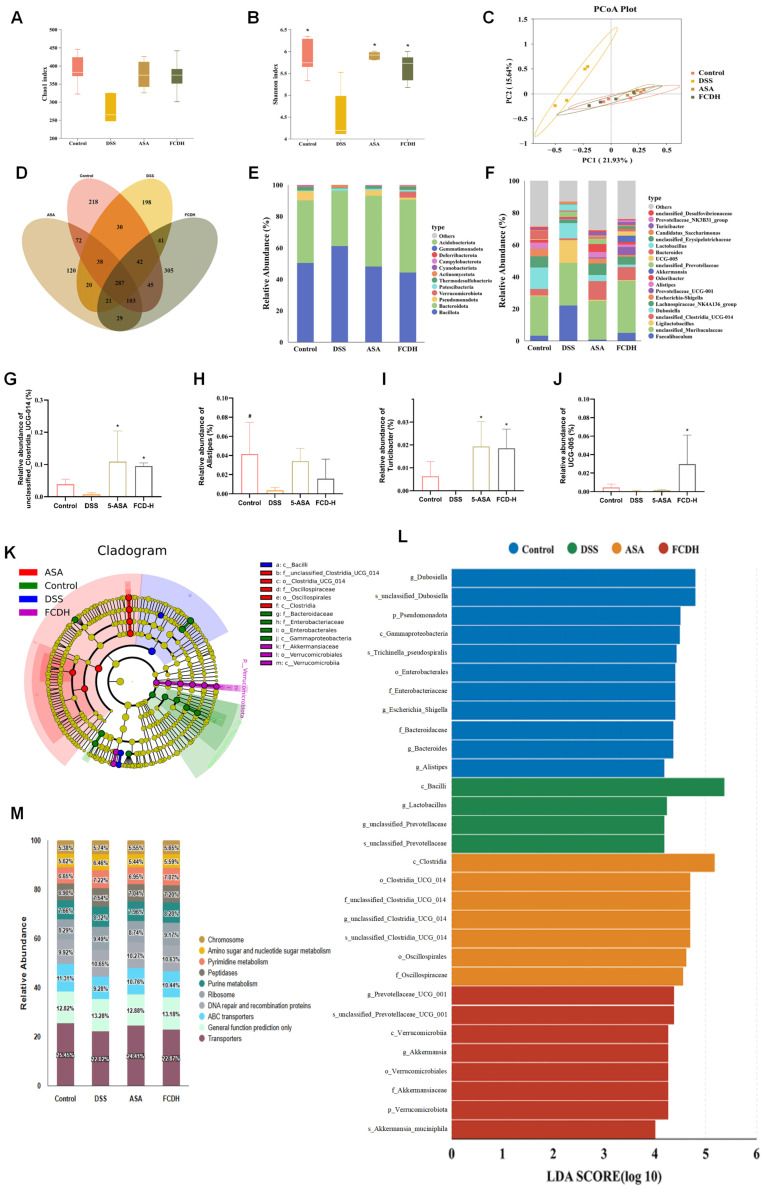 Figure 6
Figure 6- —Special Project of the Central Government in Guidance of Local Science and Technology Development of the Xizang Autonomous Region, China
- —Science and Technology Projects of Xizang Autonomous Region, China
- —Central Government Guidance for Local Project, China
- —Natural Science Foundation of Xizang Autonomous Region, China
- —Internal Scientific Research Project of Xizang Minzu University, China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInflammatory Bowel Disease · Phytochemical Studies and Bioactivities · Gut microbiota and health
1. Introduction
Ulcerative Colitis (UC) is classified as a chronic inflammatory bowel disease (IBD) that primarily affects the rectum, colonic mucosa, and submucosal layer [1]. The symptoms primarily manifest as weight loss, diarrhoea with blood in the stool, and immune dysregulation [2,3]. UC is characterised by recurrent episodes, a protracted course, and a complex, multifactorial pathogenesis, with its precise mechanisms remaining unclear [4,5]. The incidence of the disease is rising in newly industrialised nations while remaining high in Western developed countries. In 2023, the global prevalence of ulcerative colitis is estimated to reach a maximum of 5 million cases, with its incidence continuing to rise on a worldwide basis [6,7]. Several factors have been posited as potential triggers for UC onset, including disruption of the gut microbiota, metabolic disorders, damage to the intestinal barrier, and alterations in intestinal immune regulation [8]. Current treatment modalities for UC principally comprise drug therapy, surgical interventions, and the use of biological inhibitors [9,10]. However, conventional drug treatments have shown limited efficacy, and prolonged use may lead to adverse effects. The primary function of these medications is to reduce inflammation.
The intestinal barrier comprises epithelial cells, a mucus layer, and tight junction proteins. Enhancing intestinal barrier function may exert a protective effect against UC [11]. Impaired intestinal barrier function is commonly associated with various gastrointestinal disorders. Down-regulation of tight junction (TJ) protein expression in intestinal epithelial cells has been demonstrated to increase intestinal permeability, allowing harmful substances such as bacteria and toxins to enter the bloodstream. Experimental studies have shown that UC patients who achieve mucosal healing exhibit superior long-term clinical remission outcomes compared with those who fail to attain mucosal healing [12]. Moreover, the gut microbiota plays a pivotal role in the development of inflammatory bowel diseases [13,14,15,16,17]. Chinese yam (Dioscorea) polysaccharide, Tetrastigma hemsleyanum polysaccharide, Hypecoum leptocarpum and multiple natural products can mediate anti-inflammatory and mucosal repair effects by regulating microbial composition and activity [18,19,20,21]. Consequently, preserving the intestinal barrier and regulating the gut microbiota are recognised as pivotal objectives in the prevention and treatment of UC. At present, 5-aminosalicylic acid (5-ASA) is used in the management of UC. This synthetic compound is a member of the nonsteroidal anti-inflammatory drug (NSAID) class, primarily acting upon the intestinal mucosa to inhibit prostaglandin synthesis, scavenge oxygen free radicals, and reduce intestinal permeability. However, its use may cause adverse effects, including abdominal discomfort, nausea, weight loss, diarrhoea, and headaches [5,22,23]. Therefore, identifying safe and effective treatment options has become a priority for current research.
The genus Fritillaria is one of the most extensive genera within the family Liliaceae [24]. It is native to several Asian countries, including China, Korea, and Japan. FCD is a plateau-endemic medicinal plant belonging to the Fritillaria genus in the lily family [25]. It plays a distinctive role in treating digestive-system ailments in Tibetan medical tradition. Primarily cultivated on the Qinghai–Tibet Plateau at altitudes of 3000 and 4500 m and serves as a key medicinal herb for addressing gastrointestinal heat syndromes. Tibetan medical theory holds that the bitter and cooling properties of the herb clear intense heat from the stomach and intestines, detoxify the body, and promote wound healing. Recent studies have indicated that FCD exerts a mitigating effect on the development of non-alcoholic fatty liver fibrosis in murine models [26]. However, in-depth research into the pharmacological mechanisms of FCD’s effects on the gastrointestinal tract remains largely unexplored.
This study aimed to investigate the efficacy of FCD extract in treating Dextran Sulfate Sodium (DSS)-induced UC in mouse. This pioneering work represents the first demonstration that FCD effectively mitigates colonic tissue damage and inflammatory responses in a DSS-induced UC model by regulating intestinal barrier integrity and maintaining gut microbiota homeostasis. These findings provide novel insights for developing platform-based gastrointestinal protective therapeutics.
2. Materials and Methods
2.1. Reagents and Materials
FCD was procured from the Tibet Snow Region Pharmacy Chain Co., Ltd. (Lhasa, China). DSS and 5-ASA were obtained from SAITONG (Tianjin, China). The enzyme-linked immunosorbent assay (ELISA) kits for IL-6, IL-10, IL-1β, and TNF-α were procured from Enzyme Immunoassay (Shanghai, China). The protein quantification kit for determining protein concentration was purchased from Servicebio (Wuhan, China). The following antibodies were purchased from Affinity: Zonula occludens-1 (ZO-1), claudin-1, occludin, Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) and β-Tubulin.
2.2. Preparation of FCD Extract
A quantity of 100 g of FCD should be added to 1 L of ultrapure water, and the mixture then subjected to the process of decoction for a period of three hours. The decoction should then be collected, after which a further 500 mL of ultrapure water should be added and the mixture should be left to decoct for a period of 1.5 h. The decoctions should then be collected and combined. The decoction should then be concentrated by rotary evaporation, after which the process of drying should be undertaken using a vacuum freeze-dryer (Qingdao Yonghe Chuangxin Electronic Technology Co., Ltd., Qingdao, China). Prior to use, the lyophilized FCD extract powder must be stored at −80 °C.
2.3. UPLC-MS/MS Analysis of FCD
For the analysis of FCD, This project utilises an ultra-high performance liquid chromatography system (Vanquish, Thermo Fisher Scientific, Waltham, MA, USA) for the analysis of FCD. The chromatographic separation of the target compounds was achieved using a Pheomenex Kinetex C18 column (2.1 mm × 100 mm, 2.6 µm, Torrance, CA, USA). Liquid Chromatography (LC) mobile phase A was aqueous, containing 0.01% acetic acid, while phase B comprised a mixture of isopropanol and acetonitrile at a volume ratio of 1:1. The temperature of the sample tray is indicated as follows: 4 °C. Sample volume is 2 μL.
2.4. Animals
The experiment was conducted using male C57BL/6 mice, which were obtained from the China Dashuo Animal Experiment Centre and were of a clean-grade and healthy variety. The subject is male, aged between 6 and 8 weeks, with a weight range of 20–22 g. The animal use licence number is as follows: SCXK(Chuan)2025-0030. The mice were housed at the Experimental Animal Centre of Tibet University for Nationalities under the following conditions: maintain a temperature of (25 ± 2) °C and an air humidity of (50 ± 2)%, with a 12 h light-dark cycle, and provide ample food and water.
2.5. Experimental Design
Forty-eight mice were randomly divided into six groups, with eight mice in each group, a control group, a DSS group, a positive drug group (5-ASA), a low-dose FCD group (FCD-L), a medium-dose FCD group (FCD-M), and a high-dose FCD group (FCD-H). During the experimental period, in the preceding three days, the Control, DSS, and 5-ASA groups were administered normal water via gavage. The FCD-L, FCD-M and FCD-H groups were administered FCD extract concentrations of 225 mg/kg, 450 mg/kg and 900 mg/kg, respectively, via gavage. From the fourth day onwards, with the exception of the control group, the remaining groups were provided with drinking water that had been supplemented with 3% sodium dextran sulphate, with the objective of inducing UC. Concurrently, the 5-ASA group received 100 mg/kg 5-aminosalicylic acid via gavage, while the FCD-L, FCD-M, and FCD-H groups continued gavage at the aforementioned concentrations. On the tenth day, the mice were euthanised by cervical dislocation [27]. Collect serum, colon, liver, spleen, and caecal contents.
2.6. Disease Activity Index Analysis and Histopathology
A daily assessment of the DAI is to be conducted following the administration of the DSS treatment [28,29]. Subsequent to the conclusion of the experiment, photographs were taken of the anus, colon, and spleen, and measurements were taken of the length of the colon from the caecum to the anus and of the spleen. For the purpose of histopathological assessment, fix colon and liver tissue sections with 4% paraformaldehyde. Paraffin-embedded colonic specimens were sectioned for haematoxylin and eosin (H&E) staining and Alcian Blue—Periodic Acid Schiff (AB-PAS) staining. The 3D digital slide scanner model P250 FLASH (Dangier, Jinan, China) was utilised in this study.
2.7. Alcian Blue—Periodic Acid Schiff Stain Staining
Following deparaffinization and rehydration, colonic tissue sections from mice were subjected to Alcian blue staining (for acidic mucins) and the periodic acid-Schiff reaction (for neutral mucins). The sections were then counterstained with hematoxylin to visualise nuclei, followed by dehydration, clearing, and mounting. This staining protocol was employed to distinguish goblet cells and characterise mucin types.
2.8. Cytokine Assay
The levels of cytokines IL-1β, IL-6, IL-10, and TNF-α in serum samples were measured using an enzyme-linked immunosorbent assay (ELISA) kit, with the experimental methodology adhering to the kit instructions.
2.9. Immunofluorescence and Immunohistochemistry
For the purpose of immunofluorescence analysis, colon sections were deparaffinised and rehydrated prior to being subjected to an overnight incubation at 4 °C with antibodies ZO-1 (1:100) and occludin (1:200). The sections were then subjected to incubation with CY3H- and FITC-labelled secondary antibodies, with nuclei counterstained using DAPI. Image acquisition was performed using a 3D digital slide scanner, and the mean fluorescence intensity per image was determined using ImageJ software (V1.52a) analysis system.
For the purpose of immunohistochemical analysis, colon tissue sections underwent a process of dewaxing and rehydration, employing a combination of xylene and graded ethanol solutions. Antigen retrieval was subsequently performed by heating the sections in boiling sodium citrate buffer within an induction furnace. The primary antibody, ZO-1, was applied at a concentration of 1 mg/mL for a period of 12 h at a temperature of 4 °C. Subsequently, horseradish peroxidase-labelled secondary antibody was applied, followed by 3,3′-Diaminobenzidine (DAB) staining, haematoxylin counterstaining, counterstaining with blue in running water, dehydration with graded ethanol solutions and xylene, and coverslipping. A semi-quantitative analysis of the target protein was performed using ImageJ software, with protein expression intensity represented by the area occupied by positive protein.
2.10. Western Blot Analysis
The extraction of total colonic proteins was then undertaken using Radioimmunoprecipitation Assay Lysis Buffer. The centrifuge should be operated at 12,000 rpm for a period of 15 min. Proteins were subsequently extracted, and their concentrations were determined using a Bicinchoninic Acid Assay (BCA) protein assay kit based on the biuret reaction. The primary antibodies employed in this study were: GAPDH, β-Tubulin, ZO-1, occludin, and claudin-1. Subsequently, secondary antibodies were incubated at room temperature for 1 h, and protein bands were captured using an automated chemiluminescent imaging system. The relative expression of target proteins was then normalised against corresponding GAPDH and β-Tubulin internal controls.
2.11. Reverse Transcription Quantitative Polymerase Chain Reaction
Total RNA was extracted from colon tissue using the M5 Universal RNA Mini Kit (Mei5bio, Beijing, China), with RNA concentration and purity determined using an NanoDrop Microvolume Spectrophotometers (Thermo Scientific, Waltham, MA, USA). Utilising GAPDH as the housekeeping gene, the relative expression levels of target genes were calculated by means of the 2^−ΔΔCt^ method. The primer sequences for RT-qPCR are listed in Table 1. The RT-qPCR primer sequences are listed in Table 1. Primers were purchased from Shanghai Sanang Biotechnology Co., Ltd. (Shanghai, China).
2.12. 16SrRNA Sequencing
Intestinal content samples were dispatched to Novogene Co., Ltd. (Beijing, China). The 16S rRNA gene sequencing was conducted in Beijing, China. The workflow encompassed a series of meticulous procedures, including DNA extraction, PCR amplification, magnetic bead purification and recovery, quantitative fluorescence analysis, library preparation, and high-throughput sequencing. The primers 515F (5′-GTGCCAGCMGCCGCGGTAA-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′) were used to target the V3–V4 hypervariable region of the bacterial 16S rRNA gene. After quantifying the purified PCR products using Qubit and qPCR, sequencing was performed on either the NovaSeq 6000 (Illumina Inc., San Diego, CA, USA), including principal coordinate analysis (PCoA), non-metric multidimensional scaling (NMDS), α diversity analysis, and bacterial abundance difference analysis were all performed using Novigen’s cloud platform (www.novogene.com, accessed on 6 October 2025).
2.13. Statistical Analysis
Data analysis and bar chart creation were performed using GraphPad Prism 10.1.2 software. Statistical differences were determined using one-way analysis of variance (ANOVA) combined with Tukey’s post hoc test, with significance levels set at p < 0.05.
3. Results
3.1. Identification of FCD Extract
The major components of FCD extract were determined by Mass spectrometry/mass spectrometry chromatography (Figure 1A,B). The components of the identified compounds are shown in Table 2.
3.2. FCD Alleviates Symptoms of DSS-Induced Colitis in Mice
We first evaluated the effects of FCD on UC mice (Figure 1). The results showed that compared with the control group, mice in the DSS group exhibited a significant decrease in body weight. When compared with the DSS group, all 5-ASA and FCD groups demonstrated an increase in body weight. Furthermore, the FCD-L, FCD-M, and FCD-H groups demonstrated a trend towards increased body weight (Figure 2G). Following DSS-induced colitis, DAI scores significantly increased, indicating heightened inflammatory severity in mice. However, administration of 5-ASA and FCD solution markedly reduced DAI scores (Figure 2F). Furthermore, the colon length in the DSS group was significantly shorter than that in the control group. and spleen weight ratio was significantly elevated, these symptoms were markedly alleviated in the 5-ASA and FCD treatment groups (Figure 2B–D). Observation of the mice’s anuses revealed that those in the DSS group exhibited more severe haematochezia, a symptom that improved following administration of 5-ASA and FCD (Figure 2E).
3.3. FCD Ameliorates Histopathological Alterations in DSS-Induced Mouse Colitis
In H&E-stained colonic sections, the control group exhibited intact colonic architecture with no histological alterations, whereas sections from the DSS group demonstrated inflammatory cell infiltration, ulceration, crypt and marked epithelial damage (Figure 3A). Furthermore, histopathological scores in mice from the 5-ASA group and all FCD groups were lower than those in the DSS group, with a concentration-dependent reduction observed across all FCD groups (Figure 3C). Histological evaluation revealed that 5-ASA, FCD-M, and FCD-H significantly enhanced intestinal architecture and mitigated DSS-induced colonic inflammation. Liver sections revealed no significant differences across all groups (Supplementary Figure S1). The findings suggest that FCD is efficacious in ameliorating UC in murine models without causing hepatic damage.
AB-PAS-stained colonic tissue revealed a reduction in goblet cell numbers within the DSS group compared to the control group, demonstrating that intestinal mucosal damage following DSS treatment leads to diminished goblet cell populations. Both the 5-ASA and FCD groups exhibited mitigation of DSS-induced goblet cell. These findings confirm that FCD exerts a certain ameliorative effect on DSS-induced colitis in mice (Figure 2B,D).
3.4. Regulation of DSS-Induced Inflammatory Cytokine Levels in Mice by FCD
In order to gain further insight into the inflammatory status of FCD-treated DSS mice, serum levels of inflammatory cytokines were measured. In comparison with the control group, the mean levels of IL-6, IL-1β, and TNF-α were significantly elevated in the DSS group. In comparison with the DSS group, both 5-ASA and FCD groups demonstrated significant decreases. Concurrently, the mean level of IL-10 was found to be significantly reduced in the DSS group in comparison with the control group. Compared with the DSS group results, only the FCD-H group showed significant growth (Figure 3E–G).
3.5. FCD Ameliorates DSS-Induced Intestinal Barrier Damage in Mice
Immunofluorescence results revealed a significant decrease in the number of ZO-1 and occludin-positive cells in the DSS group compared with the control group. In comparison with the DSS group, the 5-ASA, FCD-M, and FCD-H groups demonstrated a substantial upregulation of ZO-1 and occludin-positive cells (Figure 4A,C,D). Immunohistochemical analysis revealed that the ZO-1 protein expression levels in the DSS group were lower than those in the Control group. In comparison with the DSS group, ZO-1 expression levels increased in all 5-ASA and FCD groups, with the most pronounced increase observed in the FCD-H group. Furthermore, FCD groups exhibited a dose-dependent increase in expression (Figure 4B,E). Western blot analysis revealed that the expression levels of ZO-1, occludin, and claudin-1 were reduced in the DSS group. Conversely, there was a significant elevation in the expression levels of tight junction proteins in the 5-ASA, FCD-M, and FCD-H groups (Figure 5A–E). RT-qPCR results demonstrated that tight junction protein expression was significantly reduced in the colon of the model group, whereas it was markedly upregulated in the 5-ASA, FCD-M, and FCD-H groups, with increased transcriptional levels (Figure 5F–H). In conclusion, these findings suggest that FCD has a protective effect against DSS-induced intestinal barrier damage.
3.6. FCD Modulates DSS-Induced Gut Microbiota Dysbiosis
Intestinal microbiota, essential for maintaining gut homeostasis, plays a pathogenic role in the initiation and progression of intestinal inflammation when dysbiosis occurs, Therefore, we used 16S rRNA gene sequencing to analyse the contents of the colon. The OTU counts for the four groups (Control, DSS, 5-ASA and FCD-H) were 218, 198, 120 and 305, respectively (Figure 6D). Evaluating intestinal flora α-diversity using the Chao1 and Shannon indices revealed that DSS-induced α-diversity declined, whereas FCD treatment reversed this trend (Figure 6A,B). PCoA and NMDS analyses further revealed differences in the overall microbial community structure among the Control, DSS, 5-ASA, and FCD-H groups (Figure 6C).
At the phylum level, Bacillota increased and Bacteroidota decreased in the DSS group compared to the control group. However, FCD and 5-ASA treatment reversed this trend, as demonstrated by reduced Bacillota and increased Bacteroidota. FCD significantly improved dysbiosis in unclassified_Clostridia_UCG-014, UCG-005, Lactobacillus and Turicibacter (Figure 6E–J). LEFSE analysis (LDA > 3.0) was used to investigate the presence of biomarkers in the Control, DSS, 5-ASA and FCD-H groups. At the genus level, significant differences in biomarkers were observed in the control group for the following genera: Dubosiella, Alistipes, Bacteroides, Escherichia_Shigella and Dubosiella. In the DSS group, significant differences in biomarkers were observed for Lactobacillus and unclassified_Prevotellaceae. Within the 5-ASA group, unclassified_Clostridia_UCG_014 showed significant differences as a biomarker. In the FCD-H group, Prevotellaceae_UCG-001 and Akkermansia were identified as biomarkers. In conclusion, FCD was effective in alleviating the imbalance in intestinal flora associated with DSS (Figure 6K,L).
The relative abundance distribution of functional genes in the Control, DSS, ASA, and FCDH groups is as follows: Transporters were among the most abundant functional categories across all groups (accounting for 22.02–25.45%), while the abundance of metabolism-related functions (e.g., purine metabolism, pyrimidine metabolism) exhibited gradient changes among different treatment groups. The abundance of “only general functional predictions” in the DSS group (13.28%) was significantly higher than that in the control group (12.82%), while the abundance of ABC transporters in the FCDH group (10.44%) was slightly lower than that in the ASA group (10.76%) (Figure 6M). These intergroup differences in functional gene abundance indicate that different treatments exert specific regulatory effects on the functional gene composition of the samples.
4. Discussion
UC is a chronic, recurrent inflammatory disease characterised by common biological phenotypes, including diarrhoea, bloody stools, and weight loss, which significantly impacts patients’ quality of life. FCD is a traditional Xizang herbal medicine with potential immunomodulatory and anti-inflammatory effects. Nevertheless, its effects on UC remain unclear. The present study aims to investigate the protective effects of FCD and its underlying mechanisms in a DSS-induced UC mouse model.
In this study, UPLC–MS/MS was first employed for compositional analysis, ultimately identifying Sipeimine, Peiminine, Peimine, and Peimisine as the primary representative components likely playing a central role [30,31,32].This study established a UC mouse model using 3% DSS. The DSS group exhibited a range of adverse outcomes, including weight loss, shortened colons, increased spleen-to-body weight ratios, reduced food intake, and bloody stools. Detailed microscopic analysis of the tissue samples revealed the presence of mucosal erosion, accompanied by the destruction of both the submucosal and serosal layers. Conversely, the 5-ASA and FCD groups exhibited a marked reduction in DAI scores, indicating superior clinical and histological outcomes in comparison to the model group, accompanied by a significant amelioration of inflammatory symptoms. The mucus secreted by goblet cells forms a protective layer over the surface of gastrointestinal epithelial cells, thereby effectively isolating pathogenic microorganisms from invasion. Inflammation has been demonstrated to damage these goblet cells, resulting in reduced mucin secretion [33]. Using of AB-PAS staining facilitates the visualisation of acidic mucins in colon tissue, which manifest as blue, while neutral mucins are characterised by a purple appearance. The findings demonstrated a substantial decrease in mucin levels within the colon of the model group, while both the 5-ASA group and the FCD group exhibited a significant increase in mucin content. The results of this study indicate that FCD substantially alleviates DSS-induced UC-associated symptoms.
The oral administration of DSS has been demonstrated to induce severe damage to the colonic mucosa, characterised by increased intestinal permeability, mucosal thickening, and extensive inflammatory cell infiltration, ultimately leading to severe intestinal inflammation [34]. Research indicates that multiple pro-inflammatory cytokines (e.g., IL-1β, IL-6, IL-10, and TNF-α) play a pivotal role in the inflammatory response of UC and contribute to the onset and progression of the disease [35,36]. In the DSS-induced colitis model, elevated levels of inflammatory markers IL-1β, IL-6, IL-10, and TNF-α can be observed. The excessive secretion of these substances has been demonstrated to disrupt immune homeostasis, thereby exacerbating intestinal inflammation and mucosal ulceration. ELISA results indicate that, in comparison with the control group, the DSS group demonstrated increased inflammatory cell infiltration and elevated inflammatory cytokines, whereas both the 5-ASA group and the FCD group exhibited notable improvements.
In the absence of underlying pathologies, the gastrointestinal tract maintains an intact barrier function that prevents the invasion of pathogens, regulates the immune system, and maintains normal bodily functions [37,38]. The intestinal barrier primarily consists of the mucus barrier and the mechanical barrier [39,40]. The superficial layer of the mucosa is composed of epithelial tissue, forming a crucial physiological boundary between the internal environment and the external world. The function of these epithelial cells is to separate the contents of the cavity from the external environment, and it has been determined that tight junction proteins are key components in this process of epithelial recognition [41,42]. Disruption of the epithelial TJ barrier in the gut allows the entry of harmful molecules into the lumen, resulting in the disruption of the mucosal immune system and subsequent inflammation [43]. In the typical instance of UC, there is a downregulation of the three tight junction proteins ZO-1, Occludin and Claudin-1, which leads to a disruption of the intestinal barrier function [44,45,46,47]. Therefore, the present study sought to assess colonic intestinal integrity by examining the distribution and expression of tight junction proteins in the colon. The experimental findings demonstrated that the administration of FCD significantly ameliorated the impairment of the intestinal barrier induced by UC. Patients suffering from UC frequently exhibit signs of gut microbiota dysbiosis, which is primarily characterised by a decrease in microbial diversity [48]. Extensive research indicates that DSS induces changes in the microbial diversity and community composition of mice. A substantial body of research has demonstrated that the administration of faecal microbiota transplantation (FMT) from healthy human donors into DSS-induced UC mouse models can effectively alleviate the associated symptoms in these animals [49]. Consequently, the restoration of gut microbiota homeostasis has become a key focus of current UC research [50,51,52]. This study used 16S rRNA sequencing analysis to reveal that FCD can reverse DSS-induced gut dysbiosis and enhance the diversity of the gut microbiota. It can also modulate the abundance of specific bacterial genera. An increase in Bacteroidetes and a decrease in Bacillota were observed, consistent with previous reports [53]. Additionally, the abundances of unclassified_Clostridia_UCG-014, UCG-005 and Turicibacter increased significantly within the FCD-H group.
In this experiment, the relative abundance of the purine metabolism pathway in the gut microbiota of mice in the Control group was 7.68%. while in DSS-induced UC model mice, this pathway’s abundance significantly increased to 9.32%. This aligns with previous findings that “the gut microbiota functional pathways in UC model mice involve purine metabolism”, suggesting that abnormal activation of the purine metabolism pathway is one characteristic of UC gut microbiota dysregulation and may contribute to the inflammatory pathological process. Following administration of ASA and FCDH, pathway abundance decreased to 7.96% and 8.20%, respectively, approaching baseline levels in the Control group. This aligns with the regulatory trend reported in the literature that “Luteolin treatment for UC reduces gut microbiota-associated purine metabolic activity” [54]. This suggests that ASA and FCDH may target the purine metabolism pathway by inhibiting its excessive activation, reshaping the gut microbiota metabolic functional network, and reducing inflammation-related metabolites, thereby alleviating UC-associated intestinal inflammatory damage. This provides metabolic pathway-level evidence for subsequent analysis of their specific action targets. In this experiment, the relative abundance of ABC transporters in DSS-induced UC model mice (9.28%) was significantly lower than that in the blank control group (11.31%). consistent with literature reports indicating a trend toward downregulation of certain ABC transporter subtypes in the colon of patients with active UC. This confirms that abnormal transporter abundance represents a common pathological alteration associated with UC. Following administration of ASA and FCDH, the relative abundance of these transporters recovered to 10.76% and 10.44%, respectively, approaching control levels. echoing literature findings that UC therapeutics can modulate ABC transporter expression. Combined with reports indicating that altered expression of this transporter is a functional contributor to UC pathogenesis rather than merely a concomitant feature of epithelial injury, these results provide experimental support for ASA and FCDH intervention in UC through regulating ABC transporter abundance. They also substantiate the potential therapeutic value of this transporter as a target for UC treatment [55].
5. Conclusions
In conclusion, FCD not only protects the integrity of the intestinal barrier by enhancing the distribution and expression of tight junction proteins, but also ameliorates DSS-induced UC symptoms by improving dysbiosis in the gut microbiota of mice. This study provides a theoretical foundation and basis for the further drug development and clinical application of FCD, while also opening new avenues for the prevention and treatment of ulcerative colitis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Voelker R. What Is Ulcerative Colitis?JAMA 202433171610.1001/jama.2023.2381438306113 · doi ↗ · pubmed ↗
- 2Danne C. Skerniskyte J. Marteyn B. Sokol H. Neutrophils: From IBD to the Gut Microbiota Nat. Rev. Gastroenterol. Hepatol.20242118419710.1038/s 41575-023-00871-338110547 · doi ↗ · pubmed ↗
- 3Kang N. Fan Z. Yang L. Shen J. Shen Y. Fang Z. Li B. Yang B. Wang J. Camel Milk Protein Ameliorates Ulcerative Colitis by Modulating Gut Microbiota and Amino Acid Metabolism Nutrients 20251778010.3390/nu 1705078040077650 PMC 11902107 · doi ↗ · pubmed ↗
- 4Zhang Y. Ji W. Qin H. Chen Z. Zhou Y. Zhou Z. Wang J. Wang K. Astragalus Polysaccharides Alleviate DSS-Induced Ulcerative Colitis in Mice by Restoring SCFA Production and Regulating Th 17/Treg Cell Homeostasis in a Microbiota-Dependent Manner Carbohydr. Polym.202534912282910.1016/j.carbpol.2024.12282939643403 · doi ↗ · pubmed ↗
- 5Garfias Noguez C. Ramírez Damián M. Ortiz Moreno A. Márquez Flores Y.K. Alamilla Beltrán L. Márquez Lemus M. Bermúdez Humarán L.G. Sánchez Pardo M.E. Microencapsulation and Probiotic Characterization of Lactiplantibacillus Plantarum LM-20: Therapeutic Application in a Murine Model of Ulcerative Colitis Nutrients 20251774910.3390/nu 1705074940077619 PMC 11901509 · doi ↗ · pubmed ↗
- 6Ng S.C. Shi H.Y. Hamidi N. Underwood F.E. Tang W. Benchimol E.I. Panaccione R. Ghosh S. Wu J.C.Y. Chan F.K.L. Worldwide Incidence and Prevalence of Inflammatory Bowel Disease in the 21st Century: A Systematic Review of Population-Based Studies Lancet 20173902769277810.1016/S 0140-6736(17)32448-029050646 · doi ↗ · pubmed ↗
- 7Le Berre C. Honap S. Peyrin-Biroulet L. Ulcerative Colitis Lancet 202340257158410.1016/S 0140-6736(23)00966-237573077 · doi ↗ · pubmed ↗
- 8Wei F.-H. Xie W.-Y. Zhao P.-S. Ji Z.-H. Gao F. Chen C.-Z. Zhang Z. Gao W. Yuan B. Crataegus Pinnatifida Polysaccharide Alleviates DSS-Induced Colitis in Mice by Regulating the Intestinal Microbiota and Enhancing Arginine Biosynthesis Phytomedicine 202514215679410.1016/j.phymed.2025.15679440315641 · doi ↗ · pubmed ↗