Generation of Novel Natural Products by Disrupting Azaphilone Synthesis in Penicillum sclerotiorum E23Y-1A
Wenjun Chang, Yanhua Yang, Ruijun Duan, Heye Qin, Shiwen Chen, Yanbo Zeng

TL;DR
Scientists disrupted a gene in a marine fungus to create new natural products, including two novel compounds with potential medicinal uses.
Contribution
The study demonstrates that deleting the A00667 gene redirects fungal metabolism to produce novel natural products.
Findings
Deleting A00667 led to the production of two new meroterpenoids, sclerotilins A and B.
Compound 6 showed moderate antimicrobial activity against Staphylococcus aureus.
Compound 3 exhibited moderate cytotoxicity against human cancer cell lines.
Abstract
Marine-derived filamentous fungi are a rich source of structurally diverse and biologically active natural products. However, many biosynthetic gene clusters (BGCs) in fungi remain silent under standard conditions. In this study, we employed a metabolic shunting strategy to disrupt azaphilone biosynthesis in the marine-derived fungus Penicillium sclerotiorum E23Y-1A by deleting the pathway-specific regulator gene A00667. HPLC analysis revealed the emergence of new metabolite peaks in the mutant strain Δ667 compared to the wild type. Subsequent purification yielded seven compounds: the mutant produced two novel meroterpenoids sclerotilins A and B (1 and 2) along with the known steroids ergosta-5,7,22-trien-3β-ol (3) and cerevisterol (4), while the wild type yielded the known steroid (22E)-5α,8α-epidioxyergosta-6,22-dien-3β-ol (5) and two azaphilones geumsanol G (6) and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
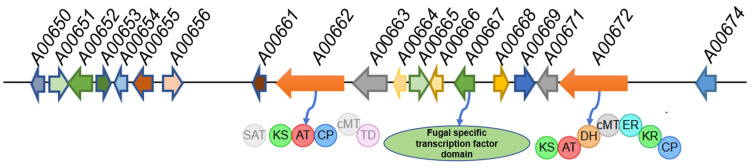 Figure 1
Figure 1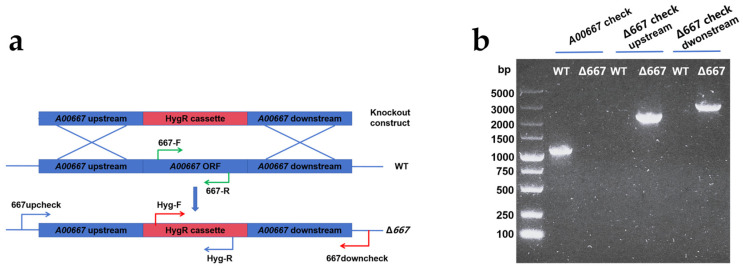 Figure 2
Figure 2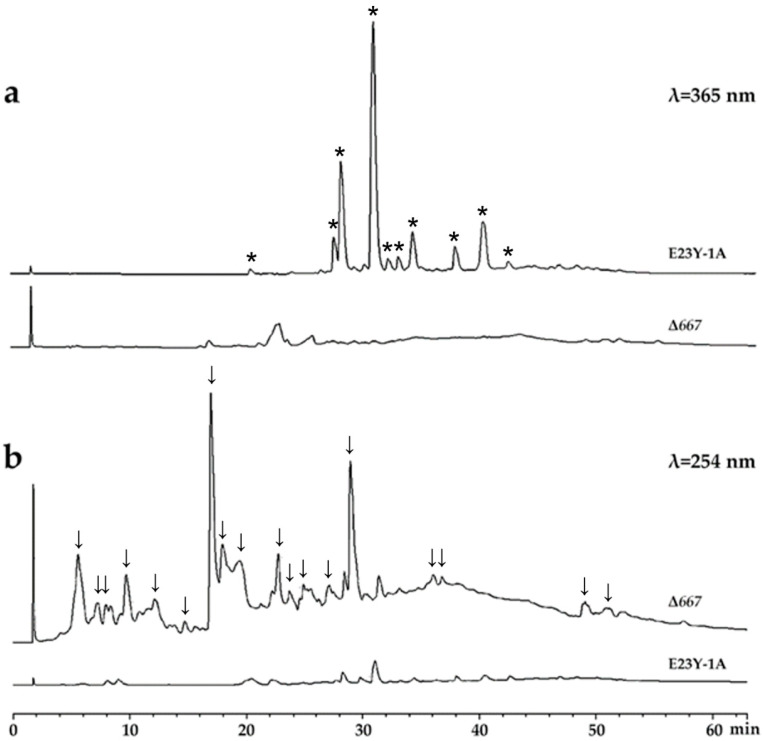 Figure 3
Figure 3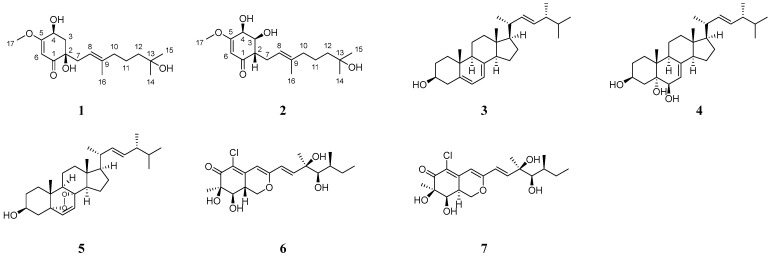 Figure 4
Figure 4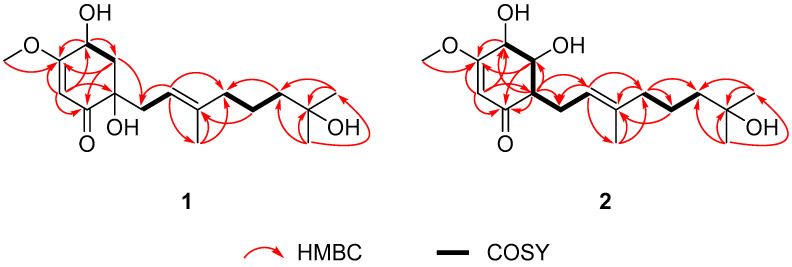 Figure 5
Figure 5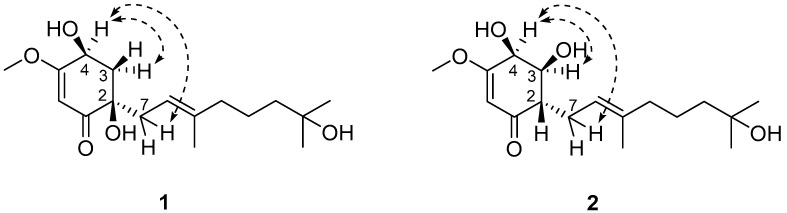 Figure 6
Figure 6- —Hainan Provincial Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSteroid Chemistry and Biochemistry · Fungal Biology and Applications · Microbial Natural Products and Biosynthesis
1. Introduction
The marine environment is an invaluable source of novel natural products characterized by their structural diversity, significant bioactivity, and chemical novelty [1,2,3]. It has been reported that various filamentous fungi of marine origin, such as Aspergillus, Penicillium, and Fusarium, are capable of synthesizing a range of bioactive compounds that are pharmaceutically promising [4,5,6,7].
One major challenge in natural product research is the ability to discover novel, biologically active compounds. Advances in genome sequencing and biosynthetic gene cluster (BGC) prediction have revealed that fungi harbor millions of cryptic BGCs, underscoring their immense potential to produce novel natural products [8]. Diverse strategies including One Strain Many Compounds (OSMAC), cocultivation, and genetic, epigenetic, and metabolic engineering techniques are now being used to unlock their potential for producing novel natural products [9,10,11,12,13]. Metabolic shunting can be controlled through enzymatic activity, which allows researchers to engineer microorganisms by introducing or modifying genes for the production of pharmaceuticals or other valuable compounds [13,14].
Penicillium sclerotiorum E23Y-1A is a filamentous fungus isolated from the marine sponge Holoxea sp. [15]. In our previous studies, we found that azaphilones constitute the primary metabolic composition of P. sclerotiorum, leading to the identification of dozens of novel azaphilone compounds [15,16]. To explore its biosynthetic potential beyond azaphilone production, in this work, we implemented a metabolic shunting strategy by deleting a key regulatory gene involved in the azaphilone biosynthetic pathway. This intervention successfully suppressed azaphilone biosynthesis and enabled the discovery of new metabolites. We achieved the characterization of novel natural products while also confirming gene A00667 as a pathway-specific regulator essential for azaphilone biosynthesis.
2. Results
2.1. The P. sclerotiorum Genome and Azaphilone BGC
The genome of P. sclerotiorum strain E23Y-1A was assembled into 181 contigs, totaling 35.9 Mbp with a GC content of 48.48% and 10,868 genes predicted. Genome mining of P. sclerotiorum with antiSMASH (version 8.0.4) revealed 70 biosynthetic gene clusters (BGCs), dominated by 29 nonribosomal peptide synthetases (NRPSs), 20 polyketide synthases (PKSs, 8 of which were hybrids), and 12 terpene synthases. BLAST (version 2.17.0) analysis revealed that a PKS/terpene hybrid cluster (cluster 12.4), located on scaffold 12 of the P. sclerotiorum genome, exhibits homology to several known azaphilone biosynthetic gene clusters responsible for the biosynthesis of azasperpyranones A–F, asperfuranone, and chaetoviridin E [17,18,19,20]. The 12.4 cluster harbors two core biosynthetic genes encoding a highly reducing polyketide synthase (HRPKS, GenBank PX533551) and a non-reducing polyketide synthase (NRPKS, GenBank PX533544, Figure 1, Table S1). The genetic architecture comprising both HRPKS and NRPKS is a hallmark of many characterized azaphilone biosynthetic pathways, such as those for asperfuranone, chaetoviridin and, azanigerone [18,19,21]. The NRPKS protein A00662, which comprises starter unit acyl-CoA transacylase (SAT), β-ketoacyl synthase, acyltransferase, acyl carrier protein, and a terminal methyltransferase domain, exhibits 93% identity to Psc_Aza_B and 66% identity to CazM (Figure 1). These two homologs are, respectively, involved in the biosynthesis of halogenated azaphilones in the termite-symbiotic fungus P. sclerotiorum SNB-CN111 and in the chaetoviridin biosynthetic pathway [19,22]. Meanwhile, the HRPKS A00672 was identified to feature a seven-domain organization typical of HRPKSs, comprising β-ketoacyl synthase, acyltransferase, dehydratase, methyltransferase, enoylreductase, keto-reductase, and acyl carrier protein (Figure 1). A BLAST search revealed that its homologs include Psc_Aza_A (94% identity), a key azaphilone biosynthetic protein in P. sclerotiorum SNB-CN111 [22], and ATEG_07659 (67.0% identity), a protein within the azasperpyranone A biosynthetic cluster [17]. In addition to the core PKS genes, the 12.4 BGC also encodes other key enzymes for azaphilone biosynthesis, such as an FAD-dependent monooxygenase (A00668, GenBank PX533549) and a halogenase (A00669, GenBank PX533550), which identifies it as the putative azaphilone-producing cluster in P. sclerotiorum E23Y-1A (Figure 1, Table S1). The 12.4 cluster harbors two fungal-specific transcription factors, A00652 and A00667 (GenBank PX533548), which contain a TFMHR domain predominantly found in Aspergillus and Penicillium species (Figure 1, Table S1). BLAST analysis revealed that both proteins share similarity with Zn(II)2_Cys_6-type transcription factors, suggesting their regulatory role in the azaphilone biosynthetic pathway of P. sclerotiorum. Consequently, we targeted A00667 for gene deletion in this study to assess its necessity for azaphilone production in P. sclerotiorum.
2.2. Generation and Confirmation of Mutant Strain Δ667
The Δ667 mutants were generated by replacing the A00667 gene in wild-type P. sclerotiorum with a hygromycin resistance (HygR) cassette via homologous recombination, selected on hygromycin-containing plates, and confirmed by PCR (Figure 2). No specific 1087 bp band corresponding to the wild-type A00667 gene was amplified in the Δ667 strain, indicating successful gene disruption (Figure 2b). Furthermore, to verify the correct integration at the target locus, the upstream and downstream junctions were amplified using primer pairs 667upcheck/Hyg-R and Hyg-F/667downcheck, respectively (Figure 2a). This yielded the expected 2254 bp and 3035 bp fragments from the Δ667 genomic DNA (Figure 2b). Collectively, these results confirm that the A00667 gene was successfully replaced at the correct chromosomal locus.
2.3. Effect of Gene A00667 Disruption on Metabolite Production
To investigate the metabolic changes resulting from A00667 disruption, we performed HPLC analysis on crude extracts from both the wild-type strain E23Y-1A and the mutant strain Δ667. The results showed that all dominant peaks present in the wild-type strain E23Y-1A were absent in the mutant strain Δ667 when detected at 365 nm (Figure 3a). Building on our prior work and the metabolic characterization performed in this study, we confirmed that these peaks correspond predominantly to azaphilones in E23Y-1A [15,16,23], indicating that the knockout of gene A00667 severely disrupted the biosynthesis of azaphilone compounds. In contrast, when detected at 254 nm, a set of new peaks eluting between 0 and 30 min emerged in the HPLC profile of Δ667 compared to that of E23Y-1A, suggesting the production of novel compounds in the mutant strain (Figure 3b). These results clearly demonstrate that A00667 is involved in the biosynthesis regulation of azaphilone compounds and that its disruption led to the production of several novel metabolites.
2.4. Structural Elucidation of New Compounds
To compare the chemical composition between the wild-type strain E23Y-1A and the mutant strain Δ667, crude extracts from both strains were purified by silica gel chromatography and semi-preparative HPLC. This purification afforded seven compounds (Figure 4), comprising two novel meroterpenoids sclerotilins A and B (1 and 2) [24,25] and two known steroids—ergosta-5,7,22-trien-3β-ol (3) [26,27] and cerevisterol (4) [28,29]—from the mutant strain 667, as well as three known compounds from the wild-type strain E23Y-1A: the steroid (22E)-5α,8α-epidioxyergosta-6,22-dien-3β-ol (5) [30,31], along with two azaphilones, geumsanol G (6) [32,33] and 5-chloro-3-[(1E,3R,4R,5S)-3,4-dihydroxy-3,5-dimethyl-1-hepten-1-yl]-1,7,8,8a-tetrahydro-7,8-dihydroxy-7-methyl-(7R,8R,8aS)-6H-2-benzopyran-6-one (7) [34,35]. The structures of the known compounds were confirmed by comparing their 1D/2D NMR spectroscopic data with those reported in the literature.
Sclerotilin A (1) was obtained as a brown oil. Its molecular formula was determined to be C_17_H_28_O_5_, indicating four degrees of unsaturation, based on HRESIMS data of the sodium adduct ion at m/z 355.1829 (calcd. 355.1829 for C_17_H_28_O_5_Na, [M + Na]^+^). The ^1^H NMR spectrum of compound 1 showed signals attributable to two olefinic protons (δH 5.21, 5.36), four methyls (δH 1.21, 1.22, 1.59, 3.81), four methylenes (Table 1), and one methine (δH 4.51). The ^13^C NMR spectrum of compound 1 (Table 1) revealed the presence of 17 carbon signals, comprising 4 methyl carbons (δC 56.8, 29.4, 29.3 and 16.5), 5 methylene carbons (δC 43.2, 40.7, 40.1, 36.9 and 22.4), 3 methine carbons (δC 117.7, 98.7 and 66.2), and 5 quaternary carbons (δC 200.4, 176.3, 139.4, 75.4 and 71.2). The ^1^H-^1^H COSY spectrum revealed three isolated spin-systems: H-3/H-4, H-7/H-8, and H-10/H-11/H-12 (Figure 5). Subsequently, based on the key HMBC correlations from H-6 to C-1, C-2, C-4, C-5, H-3 to C-1, C-5, and H-4 to C-3, C-5, the presence of a cyclohexanone moiety was revealed. Moreover, the key HMBC correlations from H-16 to C-8, C-9, C-10 and from H-14 to C-12, C-13, C-15 revealed the existence of a farnesyl moiety. This farnesyl side chain was located on the cyclohexanone at C-2 based on the key HMBC correlations from H-3 to C-1 and C-7. The structural elucidation revealed that compound 1 is closely related to the known natural product cyclohexenoneterpene D, isolated from the mangrove-associated fungus Penicillium sp. N-5 [25]. They possess an identical core skeleton and differ only in the positional arrangement of certain substituents. Compound 1 bears a hydroxyl group at C-13 of the side chain rather than at C-15, supported by HMBC correlations observed from H-14 to C-13 and C-15. Its cyclohexenone ring contains a methoxy substituent at C-5, as evidenced by HMBC correlations from H-17 to C-5 and from H-3 to C-5. Furthermore, two hydroxyl groups are positioned at C-2 and C-4, based on HMBC correlations from H-6 to both C-2 and C-4. Thus, the planar structure of 1 was established as shown in Figure 4. The ROESY correlations of H-4 to H-7 and H-3a δH 2.58 (dd, J = 12.8, 5.5 Hz) suggest that H-4 and H-3a δH 2.58 (dd, J = 12.8, 5.5 Hz) are in α-orientation (Figure 6). Consequently, OH-2, H-3b δH 1.99 (dd, J = 12.8, 9.9 Hz), and OH-4 are in β-orientation. Thus, the relative configuration of compound 1 is defined as 2R**, 4S** (Figure 6).
Sclerotilin B (2) was obtained as a brown oil. The molecular formula of 2 was determined as C_17_H_28_O_5_ by HRESIMS at m/z 355.1839 [M + Na]^+^ (calcd. for C_17_H_28_O_5_Na, 355.1829), indicating four degrees of unsaturation. Analysis of the ^1^H and ^13^C NMR spectral of 2 revealed structural similarity with 1. The main difference is that compound 2 lacks one oxygenated quaternary carbon and one methylene group present in 1, with these being replaced by two methine carbons, indicating that the position of hydroxyl substitution has been changed (Figure 4). Further evidence was provided by ^1^H-^1^H COSY correlations of H-4/H-3/H-2/H-7/H-8 and key HMBC correlations from H-6 to C-2, H-2 to C-4, C-8, and H-3 to C-1, C-5, which collectively confirm the positioning of the two hydroxyl groups on the cyclohexenone ring at C-3 and C-4, respectively (Figure 5). Thus, the analyses of 1D and 2D NMR spectra cumulatively established the structure of compound 2, as depicted in Figure 4. The ROESY correlations of H-4 to H-7 and H-3 suggest that H-4 and H-3 are in α-orientation. Consequently, H-2, OH-3 and OH-4 are in β-orientation. Thus, the relative configuration of compound 2 is defined as 2S**, 3S**, 4*S** (Figure 6).
2.5. Antimicrobial and Cytotoxic Activities
The antimicrobial activity of the isolated compounds was evaluated against five clinically relevant bacterial strains, including Escherichia coli ATCC 25922, Pseudomonas aeruginosa ATCC 27853, Staphylococcus aureus ATCC 6538, methicillin-resistant Staphylococcus aureus ATCC 43300, and Candida albicans ATCC 10231. The results indicate that compounds 1, 2, 3, 4, and 7 showed no inhibitory activity against any of the tested pathogens. As detailed in Table 2, compound 5 exhibited weak activity solely against E. coli ATCC 25922, with an MIC of 150 µg/mL. In contrast, compound 6 displayed moderate activity against S. aureus ATCC 6538 (MIC = 17.36 µg/mL), along with weak inhibition against E. coli ATCC 25922 and P. aeruginosa ATCC 27853, with higher MIC values of 125 µg/mL and 62.5 µg/mL, respectively.
The cytotoxicity of compounds 1–7 was evaluated against five human cancer cell lines: K562 (myeloid leukemia), BEL-7402 (liver cancer), SGC-7901 (gastric cancer), A549 (non-small-cell lung cancer), and HeLa (cervical cancer). As shown in Table 3, only compound 3 exhibited moderate cytotoxic activity across all five cell lines.
Ergosterols have been extensively studied for their anticancer properties. These compounds can exert direct antitumor effects both in vitro and in vivo by mechanisms such as promoting oxidative stress, upregulating the Foxo3/Bim/Fas apoptotic axis, and antagonizing sex hormone receptors, showing activity against laryngeal, breast, prostate, bladder, and melanoma cancers [36,37,38]. In line with these findings, compound 3 (ergosta-5,7,22-trien-3β-ol) has been reported to display antiproliferative activity against K562 cells at 20.0 μM, as observed by loss of cell surface roundness and luster under microscopic examination [39]. Furthermore, ergosta-5,7,22-trien-3β-ol has demonstrated selective anticolorectal-cancer activity through down-regulation of Bcl-2, up-regulation of Bax and caspase-3, induction of mitochondrial depolarization and ROS accumulation, G2/M phase arrest in HCT116 cells, and molecular docking into the hydrophobic cleft of Bcl-2 to promote apoptosis [40]. Additionally, compound 3 exhibited moderate, dose-dependent growth inhibition against rhabdomyosarcoma (A673), colorectal carcinoma (HCT116), and breast cancer (MCF-7) cells, although it induced detectable apoptosis only at the highest tested concentration of 30 μM [41].
3. Materials and Methods
3.1. Fungal Identification, Genome Sequencing and BGC Prediction
The strain P. sclerotiorum E23Y-1A was originally isolated from the marine sponge Holoxea sp. (Quanfu Island, Sansha, China) in March 2019 and was preserved in our laboratory [16]. Wild-type of P. sclerotiorum E23Y-1A was cultured on potato dextrose agar (PDA) for 5 days. The spores were scraped from the plate and collected for DNA extraction using the SDS method [42]. The whole genome was sequenced using Illumina NovaSeq PE150 at the Beijing Novogene Bioinformatics Technology Co., Ltd. (Beijing, China). The BGCs were analyzed by the antiSMASH [43].
3.2. Preparation of A00667 Knockout Construct
The 1102-bp upstream and 1648-bp downstream sequences of A00667 were amplified with primer pairs 667up-F/667up-R and 667down-F/667down-R, respectively, and cloned into the EcoRI/ClaI and XhoI/KpnI sites of vector pCT74 to construct a fusion plasmid where the hygromycin resistance cassette (Hyg-R) was flanked by the A00667 homologous arms. The complete A00667 knockout construct (upstream-Hyg-R-downstream) was PCR-amplified from the pCT74 plasmid template with primers 667delete-F and 667delete-R. This amplified linear fragment was then used for fungal transformation.
3.3. Generation of A00667 Deletion Mutant
Protoplasts of P. sclerotiorum were prepared using a two-step enzymatic digestion protocol. Fresh spores were first inoculated into 50 mL of LMM medium and incubated at 28 °C with shaking at 200 rpm for 11 h. The resulting mycelia were harvested by filtration and subjected to the first digestion in an 8 M KCl solution containing 15 mg/mL Yatalase and 10 mg/mL Lysing enzymes at 32 °C for 3 h. After collection by centrifugation at 5000 rpm, the mycelia underwent a second digestion in a fresh 8 M KCl solution containing 15 mg/mL Lywallzyme and 10 mg/mL Driselase under the same conditions (32 °C, 3 h).
The digested mixture was filtered to collect the protoplasts, which were then pelleted by centrifugation at 800× g for 10 min at 4 °C. The pellet was washed twice with STC buffer (1.2 M sorbitol, 10 mM CaCl_2_, and 10 mM Tris-HCl, pH 7.5) and finally resuspended in 200 μL of the same buffer. The A00667 knockout construct was subsequently introduced into the protoplasts using a PEG/CaCl_2_-mediated transformation method as described by Fan et al. [44].
Putative transformants were selected on PDA plates supplemented with 100 μg/mL hygromycin and subjected to PCR confirmation. Correct in-locus recombination was verified using primer pairs 667upcheck/Hyg-R and Hyg-F/667downcheck, while the complete deletion of the A00667 gene was confirmed using the internal primer pair 667-F/667-R. The verified A00667 deletion mutants were designated Δ667. Primers used in this study are listed in Table S2.
3.4. Fungal Fermentation and Metabolite Isolation
For fermentation, both the wild-type and mutant strain Δ667 were subjected to a 40-day static fermentation at room temperature in eighty 1000 mL flasks, each containing 80 g rice and 120 mL water (autoclaved at 121 °C for 20 min). After fermentation, the cultures were extracted exhaustively with ethyl acetate (EtOAc).
The crude extract was subjected to silica gel column chromatography (CC, 200 mesh) using a gradient elution of petroleum ether/EtOAc (from 9:1 to 1:9, v/v), followed by CH_2_Cl_2_/MeOH (9:1 v/v), yielding twenty fractions (E1–E20) from the wild type E23Y-1A and eighteen fractions (C1–C18) from the mutant strain Δ667. Compound 5 (4.0 mg) was obtained from fraction E4 using silica gel CC eluted with petroleum ether/EtOAc (8:1, v/v). Fraction E17 was separated by silica gel CC eluted with petroleum ether/EtOAc (5:1, v/v) to yield 7 (5.0 mg) and then eluted with CH_2_Cl_2_/MeOH (40:1, v/v), followed with semi-preparative HPLC (MeOH/H_2_O, 70:30, v/v) to afford 6 (60.0 mg, tR = 7.1 min).
Fraction C2 was treated with 100% methanol and centrifugated at 12,000 rpm to give a chloroform-soluble precipitate, compound 3 (3.0 mg). Fraction C10 was purified by gel silica CC with a stepwise CH_2_Cl_2_/MeOH elution (80:1 to 20:1, v/v) to give 4 (1.5 mg) and nine subfractions (C10A–C10I). Subfraction C10E was purified by silica gel CC (CH_2_Cl_2_/MeOH, 40:1, v/v) and subsequent semi-preparative HPLC using (MeOH/H_2_O, 70:30, v/v) to afford compound 2 (1.6 mg, tR = 6.9 min). Similarly, subfraction C10G was purified by silica gel CC (CH_2_Cl_2_/MeOH, 80:1, v/v) followed by semi-preparative HPLC (MeOH/H_2_O, 70:30, v/v) to give 1 (2.9 mg, tR = 7.7 min).
Compound 1: brown oil; [α : −13.0 (c 0.1, MeOH); UV (MeOH) λmax (logε) 259 (2.71) nm; HRESIMS m/z: 355.1829 [M + Na]^+^ (calcd. for 355.1829, C_17_H_28_O_5_Na); ^1^H NMR (600 MHz) and ^13^C NMR (125 MHz) data in CDCl_3_; see Table 1.
Compound 2: brown oil; [α : +31.0 (c 0.1, MeOH); UV (MeOH) λmax (logε) 252 (2.58) nm; HRESIMS m/z: 355.1839 [M + Na]^+^ (calcd. for 355.1829, C_17_H_28_O_5_Na); ^1^H NMR (600 MHz) and ^13^C NMR (125 MHz) data in CDCl_3_; see Table 1.
3.5. HPLC Analysis on Crude Extracts of the Wild Strain E23Y-1A and the Mutant Strain Δ667
Crude extracts of both E23Y-1A and Δ667 were prepared at a concentration of 1 mg/mL for HPLC analysis using a gradient elution starting with MeOH/H_2_O (5:95, v/v) for 5 min, followed by a linear increase to 100% MeOH over 45 min, and then an isocratic hold at 100% MeOH for 10 min. the flow rate was 0.5 mL/min with UV detection at 365 nm and 254 nm.
3.6. General Experimental Procedures
^1^H and ^13^C NMR data were acquired using Bruker Avance III 400 and DRX 600 spectrometers (Bruker Biospin AG, Ettlingen, Germany). All chemical shifts are referenced to tetramethylsilane (TMS) and reported in parts per million (ppm), while coupling constants are presented in Hertz (Hz). HRESIMS analyses were performed on an Agilent G6520 Q-TOF mass spectrometer (Agilent, Santa Clara, CA, USA). Column chromatography was performed using silica gel (200–300 and 300–400 mesh; Qingdao Haiyang Chemical Group Co., Ltd., Qingdao, China) and Sephadex LH-20 (Merck, Darmstadt, Germany). Optical rotations were measured with a Perkin–Elmer 241MC polarimeter (PerkinElmer, Fremont, CA, USA). ECD and UV data were acquired using a Jasco J-810 spectropolarimeter (JASCO, Tokyo, Japan). Infrared (IR) spectra were recorded on a Nicolet 380 FT-IR spectrometer (Thermo, Waltham, MA, USA) using samples prepared as KBr pellets. HPLC separation was conducted using an Agilent 1260 series system employing a DAD G1315D detector and an Eclipse XDB-C18 column (5 μm, 9.4 × 250 mm) (Agilent, Santa Clara, CA, USA).
3.7. Antimicrobial Activity Assay
The minimum inhibitory concentrations (MICs) of the isolated compounds against Escherichia coli ATCC 25922, Pseudomonas aeruginosa ATCC 27853, Staphylococcus aureus ATCC 6538, methicillin-resistant Staphylococcus aureus ATCC 43300, and Candida albicans ATCC 10231 were determined using a 96-well microtiter plate broth microdilution assay. Each test compound was dissolved in dimethyl sulfoxide (DMSO), and 5 µL of the resulting solution was transferred into nutrient broth to prepare a series of two-fold serial dilutions. Subsequently, 100 µL of each compound dilution was combined with 100 µL of a bacterial suspension which was adjusted to approximately 1.0 × 10^5^ CFU/mL in each well. Levofloxacin or vancomycin were employed as positive controls, while DMSO served as the negative control. Following incubation at 25 °C for 24 h, the MIC was determined visually as the lowest concentration at which no turbidity was observed. All experiments were conducted in triplicate to ensure reproducibility.
3.8. Cytotoxic Detection
All compounds were evaluated for cytotoxic activity against five human tumor cell lines: K562 (myeloid leukemia), BEL-7402 (liver cancer), SGC-7901 (gastric cancer), A549 (non-small-cell lung cancer), and HeLa (cervical cancer). These cell lines were obtained from the Cell Bank of the Type Culture Collection at the Shanghai Institute of Cell Biology, Chinese Academy of Sciences. Cytotoxicity was assessed using a modified MTT assay as described by Zeng et al. [16]. Absorbance measurements were performed at 570 nm using a Multiskan FC photometric microplate reader (Thermo Fisher Scientific). Cisplatin served as the positive control.
4. Conclusions
In this study, we successfully disrupted the azaphilone biosynthetic pathway in P. sclerotiorum E23Y-1A by deleting the regulatory gene A00667, leading to the suppression of azaphilone production and the emergence of new metabolites. From the mutant strain, two novel meroterpenoids sclerotilins A and B (1 and 2) along with two known steroids were isolated and structurally characterized. The antimicrobial screening showed that most compounds were inactive; however, compound 5 exhibited weak activity against E. coli ATCC 25922, and compound 6 showed moderate activity against S. aureus ATCC 6538 along with weak effects against E. coli and P. aeruginosa. In cytotoxicity testing, only compound 3 exhibited moderate activity against all five tested human cancer cell lines—K562, BEL-7402, SGC-7901, and A549. The research confirms A00667 as a key regulator of azaphilone biosynthesis and demonstrates that metabolic shunting through genetic manipulation can effectively redirect fungal metabolism to yield novel natural products.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lu W.Y. Li H.J. Li Q.Y. Wu Y.C. Application of marine natural products in drug research Bioorg. Med. Chem.20213511605810.1016/j.bmc.2021.11605833588288 · doi ↗ · pubmed ↗
- 2Altmann K.H. Drugs from the oceans: Marine natural products as leads for drug discovery Chimia 20177164610.2533/chimia.2017.64629070409 · doi ↗ · pubmed ↗
- 3Chu L. Huang J. Muhammad M. Deng Z. Gao J. Genome mining as a biotechnological tool for the discovery of novel marine natural products Crit. Rev. Biotechnol.20204057158910.1080/07388551.2020.175105632308042 · doi ↗ · pubmed ↗
- 4Carroll A.R. Copp B.R. Grkovic T. Keyzers R.A. Prinsep M.R. Marine natural products Nat. Prod. Rep.20244116220710.1039/D 3NP 00061 C 38285012 · doi ↗ · pubmed ↗
- 5Lv F. Zeng Y. Novel bioactive natural products from marine-derived Penicillium fungi: A review (2021–2023)Mar. Drugs 20242219110.3390/md 2205019138786582 PMC 11122844 · doi ↗ · pubmed ↗
- 6Sun L. Wang H. Yan M. Sai C. Zhang Z. Research advances of bioactive sesquiterpenoids isolated from marine-derived Aspergillus sp.Molecules 202227737610.3390/molecules 2721737636364202 PMC 9659078 · doi ↗ · pubmed ↗
- 7Gao Y. Wang J. Meesakul P. Zhou J. Liu J. Liu S. Cao S. Cytotoxic compounds from marine fungi: Sources, structures, and bioactivity Mar. Drugs 2024227010.3390/md 2202007038393041 PMC 10890532 · doi ↗ · pubmed ↗
- 8Zhang X. Wang T.T. Xu Q.L. Xiong Y. Zhang L. Han H. Ge H.M. Genome mining and comparative biosynthesis of meroterpenoids from two phylogenetically distinct fungi Angew. Chem.20181308316832010.1002/ange.20180431729797385 · doi ↗ · pubmed ↗