Targeted Inactivation of an α/β Hydrolase Gene Enables Discovery of Heterodimeric Nenestatins
Wenzheng Wei, Xiaodong Jiang, Yiguang Zhu, Wenjun Zhang, Chunfang Yang, Qingbo Zhang, Changsheng Zhang

TL;DR
Scientists discovered new chemical compounds called NENs by inactivating a specific gene in a type of bacteria.
Contribution
Targeted gene inactivation revealed new NEN analogs and highlighted the role of Nes5 in their biosynthesis.
Findings
Inactivating the nes5 gene produced five new NEN analogs (NENs E–I) and a known compound.
NEN A and NEN B were absent in the mutant, showing Nes5 is essential for their production.
Structures of the new analogs were determined using spectroscopy and computational methods.
Abstract
Nenestatins (NENs) belong to benzo[b]fluorene-containing atypical angucyclines, a structurally diverse class of microbial natural products. Bioinformatic analysis of the NEN biosynthetic gene cluster (nes BGC) from the deep-sea sediment-derived Micromonospora echinospora SCSIO 04089 implicated Nes5 as an α/β hydrolase. The targeted inactivation of the nes5 gene led to the accumulation of five new analogs, NENs E–I (1–5), together with the known monomer homo-dehydrorabelomycin E (6). Their structures were elucidated by comprehensive spectroscopic analysis and electronic circular dichroism calculations. Notably, both NEN A and NEN B were absent in the Δnes5 mutant, indicating that Nes5 is essential for their biosynthesis; however, the exact function of Nes5 requires further exploration.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
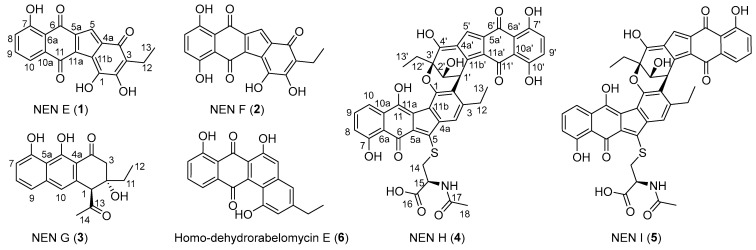 Figure 1
Figure 1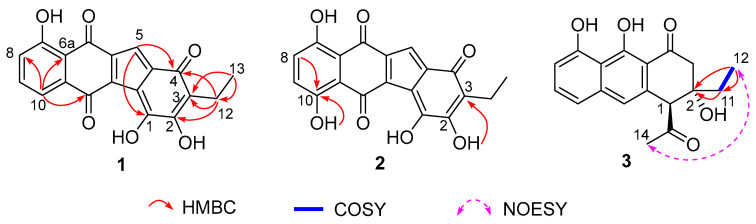 Figure 2
Figure 2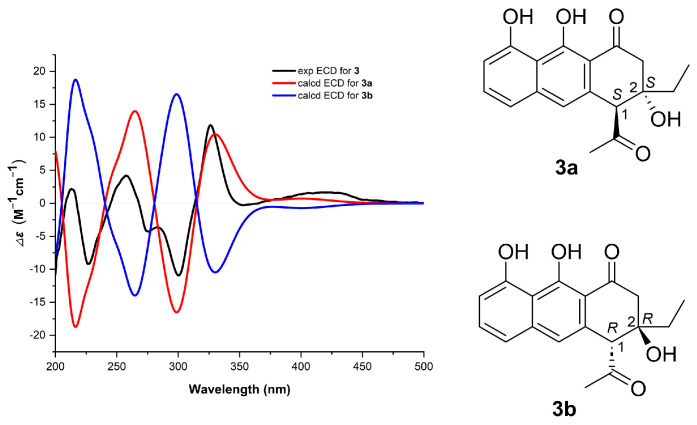 Figure 3
Figure 3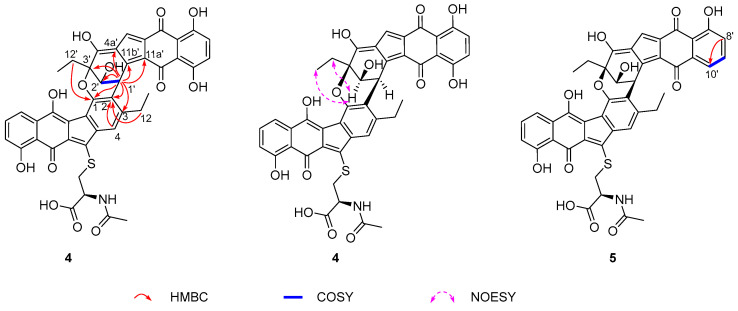 Figure 4
Figure 4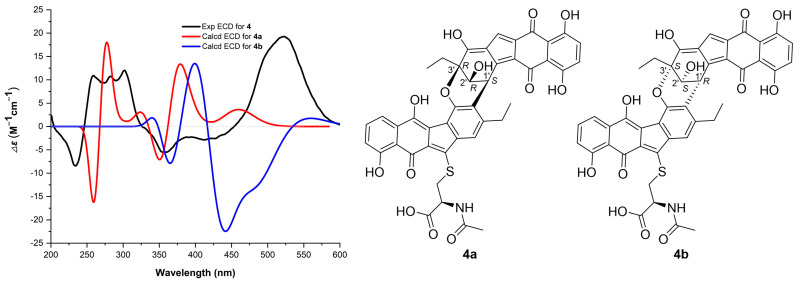 Figure 5
Figure 5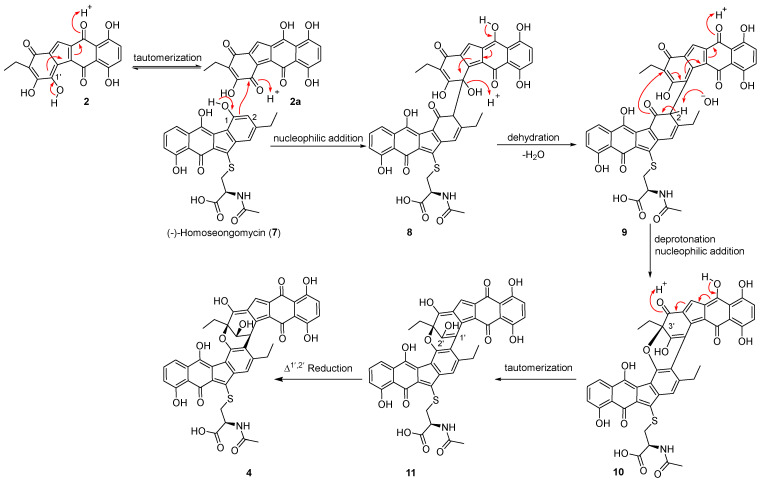 Figure 6
Figure 6- —National Natural Science Foundation of China
- —Key Science and Technology Plan Projects in Nansha District
- —National Key Research and Development Program of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Natural Products and Biosynthesis · Peptidase Inhibition and Analysis · Marine Sponges and Natural Products
1. Introduction
Benzo[b]fluorene-containing atypical angucyclines are a structurally diverse family of microbial natural products with significant biological activities [1,2]. Their biosynthesis typically originates from type II polyketide synthases (PKSs) [2,3,4]. The resulting tetracyclic precursors undergo a B-ring cleavage and subsequent rearrangement to form the characteristic benzofluorene core, which is further diversified by tailoring modifications such as oxidation, glycosylation, and dimerization [2,3,4,5]. Among all post-tailoring modifications, dimerization is especially critical for enhancing biological activity. For instance, lomaiviticin A exhibits nanomolar to picomolar cytotoxicity against a range of human cancer cell lines, with potency two orders of magnitude greater than that of its monomers [6,7,8]. Similarly, the asymmetric heterodimer difluostatin A shows superior antibacterial activity compared to its constituent monomers [9]. These cases highlight that dimerization is critical for the biological activities of this class of compounds.
Nenestatins (NENs) are a family of benzo[b]fluorene-containing atypical angucyclines isolated from the deep-sea sediment-derived actinomycete Micromonospora echinospora SCSIO 04089, with NEN B featuring an asymmetric dimeric architecture [10,11]. Although a previous study suggested that the NmrA-family protein Nes18 might be responsible for the C-2–C-1′ dimerization to generate NEN B, this enzymatic activity has yet to be biochemically confirmed in vitro [11]. Recently, in our ongoing effort to investigate the biosynthetic pathway of NENs, an α/β hydrolase encoding gene, nes5, was disrupted, which led to the accumulation of a series of novel NEN congeners (Figure S1). Herein, we report the targeted inactivation of nes5 in M. echinospora SCSIO 04089 and the isolation, structure elucidation, and bioactivity evaluation of five new nenestatin analogs, NENs E–I (1–5), together with the known analog homo-dehydrorabelomycin E (6) [12], from the Δnes5 mutant (Figure 1). Notably, NENs H and I (4 and 5) represent two novel dimers.
2. Results and Discussion
The nes5 gene was inactivated in M. echinospora SCSIO 04089 to generate the Δnes5 mutant. Fermentation of this mutant in a medium supplemented with macroporous resin HP20 yielded six nenestatin analogs (1–6), including three new monomers, NENs E–G (1–3), two new dimers, NENs H and I (4 and 5), and the known compound homo-dehydrorabelomycin E (6) [12] (Figure 1).
NEN E (1) was obtained as a red amorphous powder. Its molecular formula was established as C_19_H_12_O_6_ by HRMS (m/z: 335.0567 [M − H]^−^, calcd for C_19_H_11_O_6_, 335.0561), indicating fourteen degrees of unsaturation. Analysis of the NMR data of 1 (Table 1 and Figure S2) revealed signals for an ABC three-spin system (δC/δH 134.6/7.53, 121.5/7.05, and 118.0/7.50), a sp^2^-hybridized methine (δC/δH 115.0/6.88), a quartet methylene (δC/δH 15.8/2.32), a triplet methyl group (δC/δH 13.1/0.97), and thirteen sp^2^-hybridized non-protonated carbons. Comparative NMR analysis showed that 1 is highly similar to nenestatin C [11]. The signals for an AB-spin system (δC/δH 121.7/7.22, C-1; δC/δ_H_ 137.7/5.94, C-2) and a non-protonated carbon (δC 76.3, C-3) were present in nenestatin C, while absent in 1. Instead, three sp^2^-hybridized methines (δC 175.1, 156.1, 120.2), two of which are oxygenated, were observed in 1. The HMBC correlations from H_3_-13 to C-12 (δC 15.8)/C-3 (δC 120.2), from H-5 to C-4, and from H_2_-12 to C-2 (δC 156.1)/C-3/C-4 (δC 183.9) suggested the presence of a double bond between C-2 and C-3 (Figure 2). Additionally, the carbon at δC 175.1 was assigned as C-1 by the ^4^J long-distance HMBC correlation from H-5 to C-1. Furthermore, the 10-OH in nenestatin C was absent in 1. The assignment of 1 was further supported by HMBC correlations from H-10 to C-6a/C-8/C-11 (Figure 2).
NEN F (2) was isolated as a red amorphous powder. The molecular formula of 2 was established as C_19_H_12_O_7_ through HRMS (m/z: 351.0515 [M − H]^−^, calcd for C_19_H_11_O_7_, 351.0510), indicating the incorporation of an additional oxygen atom compared to 1. Analysis of the NMR data of 2 (Table 1 and Figure S3) revealed that the structures of 2 and 1 were highly similar. Different from the olefinic methine CH-10 (δC/δH 118.0/7.50) present in 1, an oxygenated non-protonated C-10 (δC 155.6) was found in 2. The assignment was confirmed by the HMBC correlations from 10-OH and H-8 to C-10 (Figure 2). Additionally, the exchangeable proton at δH 9.59 was assigned to the 2-OH group based on its HMBC correlation to C-3 (δC 120.1) (Figure 2). Therefore, 2 was determined as a C-10 hydroxylated derivative of 1.
NEN G (3) was isolated as a brown amorphous powder. Its molecular formula was established to be C_18_H_18_O_5_ through HRMS (m/z: 313.1081 [M − H]^−^, calcd for C_18_H_17_O_5_, 313.1081). The NMR data of 3 (Table 1 and Figure S4) were highly similar to those of huanglongmycin H [13]. The only difference was that the methyl group in huanglongmycin H was replaced with an ethyl group (δC/δH 32.1/1.51; 1.64, CH_2_-11; 6.80/0.97, CH_3_-12) in 3. The assignment was supported by the COSY correlate between H_2_-11/H_3_-12, together with HMBC correlations from H_3_-12 to C-2/C-11 and from H_2_-11 to C-2 (Figure 2). The NOESY correlation between H_3_-14 and H_3_-12 indicated a syn orientation of the acetyl group at C-1 and the ethyl group at C-2 (Figure 2). Finally, the absolute configuration of 3 was assigned to be 1S,2S, given that the experimental electronic circular dichroism (ECD) spectrum of 3 matched well with that calculated for the model compound 3a (1S,2S) (Figure 3 and Figure S5). The assignment was further corroborated by the closely matched specific rotation values of 3 = −21.0°) and huanglongmycin H ( = −21.4°).
NEN H (4) was isolated as a purple amorphous powder. Its molecular formula was established as C_43_H_33_NO_13_S through HRMS (m/z: 802.1607 [M − H]^−^, calcd for C_43_H_32_NO_13_S, 802.1600), corresponding to 28 degrees of unsaturation. Analysis of the ^1^H, ^13^C and 2D NMR data of 4 revealed the presence of two structural units, A and B (Table 2 and Figure S6). The NMR data for the unit A closely resembled those of (-)-homoseongomycin [14]. The key difference was at the C-2 position: while (-)-homoseongomycin has a sp^2^-hybridized methine (δC/δH 114.3/6.63, CH-3), the unit A exhibited a non-protonated carbon at δC 126.5 (C-2). The assignment was supported by HMBC correlations from H_2_-12 and H-4 to C-2 in 4 (Figure 4). The unit B displayed ^1^H and ^13^C NMR data consistent with those of that in nenestatin D [11]. The obvious difference was the presence of a sp^3^-hybridized CH-1′ (δC/δH 36.1/5.12) in the unit B of 4, instead of the sp^2^-hybridized and non-protonated C-1′ (δC 141.3) in nenestatin D [11]. The assignment was supported by the COSY correlation between H-1′ and H-2′, and the HMBC correlations from H-1′ to C-2′/C-3′/C-4a′/C-11a′/C-11b′. Moreover, HMBC correlations from H-1′ to C-1, C-2, and C-3 indicated a connection between CH-1′ and C-2 (Figure 4). The proposed structure accounted for only 27 degrees of unsaturation, one fewer than that determined by HRMS data. Further analysis of the HMBC spectrum revealed a long-range HMBC correlation from CH_2_-12′ to C-1, suggesting an ether linkage between C-1 and C-3′. This additional ring satisfied the final degree of unsaturation, thereby completing the determination of the planar structure of 4. The relative configuration of H-2′ and CH_2_-12′ was assigned as syn by NOESY correlations between H-2′/H_2_-12′ and H-2′/H_3_-13′ (Figure 4). Therefore, four possible absolute configurations could be assigned to 4, namely, 1′S,2′R,3′R (4a), 1′R,2′S,3′S (4b), 1′S,2′S,3′S (4c), and 1′R,2′R,3′R (4d). Finally, the absolute configuration of 4 was assigned to be 1′S,2′R,3′R, given that the experimental ECD spectrum of 4 matched well with that calculated for the model compound 4a but was different from that computed for 4b, 4c, and 4d (Figure 5, Figures S7 and S8).
NEN I (5) was isolated as a purple amorphous powder. Its molecular formula was established to be C_43_H_33_NO_12_S through HRMS (m/z: 786.1658 [M − H]^−^, calcd for C_43_H_32_NO_12_S, 786.1651), indicating one oxygen atom fewer than that of 4. A detailed comparison of the NMR data of 5 and 4 revealed their high structural similarity (Table 3 and Figure S9). A key difference was observed at the C-10′ position. Compound 4 features a sp^2^-hybridized and oxygenated carbon at C-10′ (δC 155.8), while compound 5 exhibited signals for a sp^2^-hybridized methine group at this position (δC/δH 117.4/7.54, CH-10′). The assignment was confirmed by the COSY correlation between H-9′ (δH 7.47) and H-10′ (δH 7.54), and HMBC correlations from H-8′ to C-10′. The absolute configuration of 5 was assigned as 1′S,2′R,3′R based on the identical experimental ECD spectra of 5 and 4, and their comparable specific rotation values: = −251.0° for 5 and = −262.7° for 4 (Figure S7). Finally, 5 was determined as a C-10′ dehydroxylation derivative of 4.
Compound 6 was determined to be homo-dehydrorabelomycin E [12], based on comparing its NMR data with those previously reported (Figure S10).
Bioinformatic analysis identified Nes5 as a homologue (62.7% identity) of the deacylase FlsH (Figure S11) [8]. In the fluostatin biosynthetic pathway, FlsH is characterized to catalyze the hydrolysis of acyl fluostatins to prevent their spontaneous conversion into toxic quinone methides, which act as key transient intermediates that non-enzymatically drive the formation of fluostatin dimers [8]. We therefore hypothesize that disruption of the nes5 gene would result in the accumulation of acyl nenestatins. However, no acyl nenestatins were detected in the Δnes5 mutant, implying that Nes5 does not possess a deacylase function similar to that of FlsH. Given that the abolished production of NENs A and B (Figure S1 for their structures) in the Δnes5 mutant, Nes5 should be essential for their biosynthesis. However, the exact function of Nes5 in the biosynthesis of NENs A and B requires further exploration.
Notably, five new nenestatin congeners, including two novel dimers, 4 and 5, were isolated. This suggests that Nes5 may catalyze or regulate an as-yet-unidentified key step, and its absence leads to precursor diversion and the subsequent formation of 4 and 5. Finally, we propose a plausible nonenzymatic way for the dimerization of compounds 4 and 5 (Figure 6) [11]. The formation of 4 is likely initiated by deprotonation of the 1-OH group in (-)-homoseongomycin (7) to generate a phenolate anion, which undergoes resonance and nucleophilic addition to 2a, the keto-enol tautomer of compound 2, forming the C-2–C-1′ bond in the intermediate 8. Next, 8 undergoes tautomerization followed by dehydration to yield intermediate 9. Deprotonation at CH-2 of 9 induces a nucleophilic attack by O-1 on C-3′ to form the key C-1–C-3′ ether bond in 10. Finally, a tautomerization step and subsequent enzymatic or nonenzymatic reduction in the Δ^1′,2′^ double bond in the intermediate 11 affords compound 4. Alternatively, a coupling reaction between 7 and 1 could lead to the formation of compound 5. The proposed formation of 4 and 5 resembles the nonenzymatic C−C coupling to produce several natural product artifacts [8,15]. However, the proposed pathway requires further in vitro chemical experiments to confirm.
The biosynthetic machinery of compound 3 is likely encoded by the nes BGC. The nes chain length factor (CLF) probably exhibits loose control in the chain length, and thus allows the formation of both decaketide and nonaketide precursors [16,17,18]. Therefore, we propose that a shunt nonaketide intermediate is processed via a pathway analogous to that of 4-acetylchrysophanol to yield compound 3, as previously observed in the heterologous expression of the fluostatin BGC (Figure S12) [19].
The compounds 1–6 were evaluated for antibacterial activity against six bacterial strains (Staphylococcus aureus ATCC 29213, Enterococcus faecalis ATCC 29212, Acinetobacter baumannii, Klebsiella pneumoniae ATCC 13883, Mycobacterium smegmatis MC^2^ 155, and Micrococcus luteus) via broth microdilution [20]. None of the compounds showed inhibitory activity at a concentration of 64 µg·mL^−1^. Separately, the cytotoxicity of compounds 1–5 was assessed against four human cancer cell lines (SF-268, HepG-2, MCF-7, A549) using the SRB method [21]. None of the compounds exhibited cytotoxicity against these cell lines at concentrations up to 100 µM. All tested compounds were inactive in both the antibacterial and cytotoxicity assays under the conditions employed, indicating that their structural features do not confer activity against the bacterial strains or cell lines tested. Future studies could focus on structural diversification to enhance bioactivity or on screening against additional biological targets.
3. Materials and Methods
3.1. General Experimental Procedures
Optical rotations were measured with an MCP 500 polarimeter (Anton, Graz, Austria). UV spectra were recorded on a UV-2600 spectrophotometer (Shimadzu, Kyoto, Japan). Circular dichroism (CD) spectra were recorded on a Chirascan CD spectropolarimeter (Applied Photophysics Ltd., Surrey, UK). IR spectra were measured on an IR Affinity-1 FT-IR spectrometer (Shimadzu, Kyoto, Japan). ^1^H, ^13^C, and 2D NMR spectra were recorded on a Bruker AVANCE III HD 700 MHz NMR spectrometer (Bruker Biospin GmbH & Co. KG., Rheinstetten, Baden-Württemberg, Germany), with TMS as an internal standard. HRESIMS data were measured using a MaXis 4G UHR-TOFMS spectrometer (Bruker Daltonics GmbH & Co. KG., Bremen, Germany). Materials for column chromatography (CC) were silica gel (100–200 mesh; 300–400 mesh; Jiangyou Silica Gel Development Co., Ltd., Yantai, Shandong, China), Sephadex LH-20 (40–70 μm; Amersham Pharmacia Biotech AB, Uppsala, Sweden), and YMC*GEL ODS-A (12 nm S-50 μm; YMC Company Ltd., Kyoto, Japan). Thin layer chromatography (TLC, 0.1–0.2 or 0.3–0.4 mm) was conducted with precoated glass plates (silica gel GF254, 10–40 nm, Jiangyou Silica Gel Development Co., Ltd., Yantai, Shandong, China), Medium pressure liquid chromatography (MPLC) was performed with automatic flash chromatography (Cheetahtmmp 200, Bonna-Agela Technologies Co., Ltd., Tianjin, China) with a monitoring wavelength of 220 nm and a collecting wavelength of 254 nm. Semipreparative HPLC was performed on a Hitachi HPLC station (Hitachi-L2130, Hitachi, Tokyo, Japan) with a diode array detector (Hitachi L-2455, Hitachi, Tokyo, Japan) using a ODS column (Kinetex C18, 250 mm × 10.0 mm, 5 μm; Phenomenex, Torrance, California, USA).
3.2. Construction of M. echinospora SCSIO 04089/Δnes5 Mutant
To generate a nes5 in-frame deletion mutant, two homologous DNA fragments flanking the target gene were amplified from M. echinospora SCSIO 04089 genomic DNA using primer pairs Nes5-UF/Nes5-UR and Nes5-DF/Nes5-DR (Table S1). The PCR products were purified and ligated into the pre-digested thermal-sensitive Streptomyces–Escherichia coli shuttle vector pKC1139, resulting in the construction of the gene knockout plasmid pKC1139/Δnes5. Following sequence confirmation, the deletion plasmid was introduced into E. coli ET12567/pUZ8002 by transformation and subsequently transferred into M. echinospora SCSIO 04089 via conjugation. Exconjugants were cultured at 28 °C for two successive generations to facilitate double-crossover recombination and then shifted to 37 °C to cure the plasmid. Gene knockout was verified by PCR using the flanking primers Nes5-TF and Nes5-TR (Table S1).
3.3. Fermentation, Extraction, and Isolation
The mutant strain M. echinospora SCSIO 04089*/Δnes5* was cultured on an ATCC172 agar plate (soluble starch 20.0 g L^−1^, glucose 10.0 g L^−1^, yeast extract 5.0 g L^−1^, Aobox casein 5.0 g L^−1^, CaCO_3_ 19.0 g L^−1^, artificial sea salt 10.0 g L^−1^, pH 7.0) at 28 °C for 7 days. A piece of mycelia was then inoculated into a 250 mL Erlenmeyer flask containing 50 mL of A1 medium (soluble starch 10.0 g L^−1^, yeast extract 4.0 g L^−1^, bacterial peptone 2.0 g L^−1^, artificial sea salt 10.0 g L^−1^, pH 7.0) and incubated at 28 °C with shaking at 200 rpm for 3 days to prepare seed cultures. The seed cultures were subsequently transferred into 1000 mL Erlenmeyer flask containing 200 mL of the N4 medium (soluble starch 15.0 g L^−1^, fish peptone 8.0 g L^−1^, bacterial peptone 5.0 g L^−1^, glycerol 7.5 g L^−1^, CaCO_3_ 2.0 g L^−1^, KBr 0.2 g L^−1^, artificial sea salt 30.0 g L^−1^, prewashed HP20 resin 5%, v/v, pH 7.0). A total of 21 L of culture was prepared and incubated under the same conditions (28 °C, 200 rpm) for 7 days. During fermentation, the secondary metabolites were adsorbed by the HP20 resin, which changed color from white to black. The colored HP20 resin was collected by filtration and washed several times with 4 L of CH_3_CN. The combined solvents were concentrated under vacuum to yield 3.5 g of crude extract. The extract was fractionated by MPLC with an ODS column. Elution was performed using a linear gradient under the following program: solvent system (solvent A, H_2_O containing 5% CH_3_OH; solvent B, CH_3_OH); 0% B to 60% B (0–40 min), 60% B to 85% B (40–80 min), 85% B to 100% B (80–100 min), 100% B (100–120 min); flow rate at 20 mL·min^−1^ to yield seven fractions (Fr.1–Fr.7). Further purification of Fr.1–Fr.7 was carried out by semi-preparative HPLC (Kinetex C18, 250 mm × 10.0 mm, 5 μm; Phenomenex, Torrance, California, USA) using a linear gradient under the following program: solvent system (solvent A, H_2_O; solvent B, CH_3_CN); 10% to 80% B (0–25 min), 100% B (25.1–30 min), 100% B to 10% B (30–30.1 min), 10%B (30.1–35 min); flow rate at 2.5 mL min^−1^. This process resulted in the isolation of compounds 1 (4.6 mg, tR = 21.0 min) and 2 (2.1 mg, tR = 20.3 min) from Fr.2, compound 3 (7.1 mg, tR = 23.0 min) from Fr.3, compounds 4 (2.6 mg, tR = 24.7 min) and 5 (5.4 mg, tR = 27.2 min) from Fr.4, and compound 6 (6.6 mg, tR = 28.8 min) from Fr.7.
Compound 1: red amorphous powder. UV (CH_3_OH) λmax (log ε) 209 (2.74), 278 (2.01), 424 (0.74), 488 (0.60) nm; IR (film) νmax 3750, 3387, 1560, 1456, 1313, 1213, 1004 cm^−1^; ^1^H and ^13^C NMR data, Table 1; HRESIMS m/z: 335.0567 [M − H]^−^ (calcd for C_19_H_11_O_6_, 335.0561).
Compound 2: red amorphous powder. UV (CH_3_OH) λmax (log ε) 205 (2.16), 277 (1.17) nm; IR (film) νmax 3749, 3444, 1682, 1541, 1206, 1026 cm^−1^; ^1^H and ^13^C NMR data, Table 1; HRESIMS m/z: 351.0515 [M − H]^−^ (calcd for C_19_H_11_O_7_, 351.0510).
Compound 3: brown amorphous powder. = −21.0° (c 0.03, CH_3_OH); UV (CH_3_OH) λmax (log ε) 223 (2.93), 268 (2.38), 407 (0.73) nm; IR (film) νmax 3749, 1690, 1541, 1204, 1024 cm^−1^; ^1^H and ^13^C NMR data, Table 1; HRESIMS m/z: 313.1081 [M − H]^−^ (calcd for C_18_H_17_O_5_, 313.1081).
Compound 4: purple amorphous powder. = −262.7° (c 0.03, CH_3_OH); UV (CH_3_OH) λmax (log ε) 207 (2.36), 284 (1.24), 327 (0.90), 508 (0.60) nm; IR (film) νmax 3408, 1681, 1204, 1024, 710 cm^−1^; ^1^H and ^13^C NMR data, Table 2; HRESIMS m/z: 802.1607 [M − H]^−^ (calcd for C_43_H_32_NO_13_S, 802.1600).
Compound 5: purple amorphous powder. = −251.0° (c 0.03, CH_3_OH); UV (CH_3_OH) λ_max_ (log ε) 207 (2.26), 284 (1.18), 330 (0.88), 498 (0.50) nm; IR (film) νmax 3397, 1672, 1204, 1024, 709 cm^−1^; ^1^H and ^13^C NMR data, Table 3; HRESIMS m/z: 786.1658 [M − H]^−^ (calcd for C_43_H_32_NO_12_S, 786.1651).
Compound 6: brown amorphous powder. UV (CH_3_OH) λ_max_ (log ε) 208 (2.62), 232 (2.30), 451 (0.44) nm; IR (film) νmax 3741, 1601, 1456, 1250, 746 cm^−1^; HRESIMS m/z: 333.0076 [M − H]^−^ (calcd for C_20_H_13_O_5_, 333.0768).
3.4. TDDFT-ECD Calculations
All quantum chemical calculations were performed using the Gaussian 09 (Revision D.01) software package [22]. Conformational searches were conducted using the Molecular Merck Force Field (MMFF) as implemented in Spartan’14 V1.1.4 software (Wavefunction Inc., Irvine, CA, USA). For compound 3, all conformers with a population greater than 1% were re-optimized at the B3LYP/6-311G (d,p) level of theory, while for compound 4, the CAM-B3LYP/6-311G(d,p) level was employed. These geometry optimizations utilized the IEFPCM solvation model to simulate a CH_3_OH environment. Subsequently, time-dependent density functional theory (TDDFT) calculations at the corresponding theory levels (i.e., B3LYP/6-311G (d,p)/IEFPCM(CH_3_OH) for 3 and CAM-B3LYP/6-311G(d,p)/IEFPCM(CH_3_OH) for 4) were conducted to obtain the ECD spectra for the stable conformers. Finally, the overall theoretical ECD spectrum for each compound was generated by Boltzmann averaging the individual conformer spectra using SpecDis 1.71 [23].
3.5. Antibacterial Assays
The antibacterial activities of the compounds 1–6 were evaluated against six indicator strains: Staphylococcus aureus ATCC 29213, Enterococcus faecalis ATCC 29212, Acinetobacter baumannii, Klebsiella pneumoniae ATCC 13883, Mycobacterium smegmatis MC^2^-155, and Micrococcus luteus, using the broth microdilution method [20]. Indicator strains were grown on a rotary shaker at 37 °C for 12 h. The cultures were diluted with sterilized medium to an optical density (OD_600_) of 0.04–0.06 and then further diluted 1000-fold before being dispensed into 96-well microtiter plates. Each compound was tested in triplicate over a dilution series ranging from 64 to 0.25 µg mL^−1^. After 16 h of incubation, the minimum concentrations that completely inhibited visible growth of the tested strains were determined from two independent experiments.
3.6. Cytotoxic Activity Assays
The cytotoxicities of compounds 1–5 were evaluated against SF-268 (human glioma cell line), HepG-2 (human liver carcinoma cell line), MCF-7 (human breast adenocarcinoma cell line), and A549 (human lung adenocarcinoma cell line) (The cell lines were purchased from the National Collection of Authenticated Cell Cultures in Shanghai, China) by the SRB method [21]. The cells were cultivated in RPMI 1640 medium [24]. Cells (180 µL) with a density of 3 × 10^4^ cells mL were seeded onto 96-well plates and incubated for 24 h at 37 °C, 5% CO_2_. Subsequently, 20 µL of different concentrations of 1–5, ranging from 0 to 100 µM in DMSO, was added to each plate well. An equal volume of DMSO was used as a negative control. After further incubation for 72 h, the cell monolayers were fixed with 50% (w/v) trichloroacetic acid (50 µL) and then stained for 30 min with 0.4% (w/v) SRB dissolved in 1% acetic acid. The unbound dye was removed by repeatedly washing with 1% acetic acid. The protein-bound dye was dissolved in a 10 mM Tris-base solution (200 µL) for the determination of the OD_570_ value using a microplate reader. The cytotoxic compound cisplatin was used as a positive control. All data were obtained in triplicate and are presented as means ± S.D. IC_50_ values were calculated with the SigmaPlot 14.0 software using the non-linear curve-fitting method.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kharel M.K. Pahari P. Shepherd M.D. Tibrewal N. Nybo S.E. Shaaban K.A. Rohr J. Angucyclines: Biosynthesis, mode-of-action, new natural products, and synthesis Nat. Prod. Rep.20122926432510.1039/C 1NP 00068 C 22186970 PMC 11412254 · doi ↗ · pubmed ↗
- 2Zhang Z. Pan H.X. Tang G.L. New insights into bacterial type II polyketide biosynthesis F 1000 Research 2017617210.12688/f 1000 research.10466.128299197 PMC 5321127 · doi ↗ · pubmed ↗
- 3Fan K. Zhang Q. The functional differentiation of the post-PKS tailoring oxygenases contributed to the chemical diversities of atypical angucyclines Synth. Syst. Biotechnol.2018327528210.1016/j.synbio.2018.11.00130533539 PMC 6260466 · doi ↗ · pubmed ↗
- 4Mikhaylov A.A. Ikonnikova V.A. Solyev P.N. Disclosing biosynthetic connections and functions of atypical angucyclinones with a fragmented C-ring Nat. Prod. Rep.2021381506151710.1039/D 0NP 00082 E 33480893 · doi ↗ · pubmed ↗
- 5Ma L. Zhang Q. Yang C. Zhu Y. Zhang L. Wang L. Liu Z. Zhang G. Zhang C. 1.06—Assembly Line and Post-PKS Modifications in the Biosynthesis of Marine Polyketide Natural Products Comprehensive Natural Products III Liu H.-W. Begley T.P. Elsevier Oxford, UK 2020139197
- 6Colis L.C. Woo C.M. Hegan D.C. Li Z.W. Glazer P.M. Herzon S.B. The cytotoxicity of (-)-lomaiviticin A arises from induction of double-strand breaks in DNA Nat. Chem.2014650451010.1038/nchem.194424848236 PMC 4090708 · doi ↗ · pubmed ↗
- 7Herzon S.B. The Mechanism of Action of (-)-Lomaiviticin A Acc. Chem. Res.2017502577258810.1021/acs.accounts.7b 0034728956437 PMC 5874803 · doi ↗ · pubmed ↗
- 8Huang C. Yang C. Zhang W. Zhang L. De B.C. Zhu Y. Jiang X. Fang C. Zhang Q. Yuan C.-S. Molecular basis of dimer formation during the biosynthesis of benzofluorene-containing atypical angucyclines Nat. Commun.20189208810.1038/s 41467-018-04487-z 29802272 PMC 5970136 · doi ↗ · pubmed ↗