Characteristics of a Dinophysis cf acuminata Population from a Tidewater Glacier Lagoon in a Temperate Latitude: Applications to Dinophysis Studies
Patricio A. Díaz, María García-Portela, Gonzalo Álvarez, Francisco Rodríguez, Iván Pérez-Santos, Daniel Varela, Michael Araya, Camila Schwerter, Ángela M. Baldrich, Barbara Cantarero, Beatriz Reguera

TL;DR
This paper studies a unique population of Dinophysis acuminata in a tidewater glacier lagoon in Chile, highlighting its role in shellfish poisoning and its adaptation to a rare ecosystem.
Contribution
The study presents a novel PTX2-containing Dinophysis cf acuminata population in a unique tidewater glacier lagoon environment.
Findings
Dinophysis acuminata was the only species observed in the lagoon's cold, brackish waters.
Cell densities varied seasonally, reaching up to 2800 cells per liter in summer.
Genetic analysis confirmed the species' relation to European and North American strains and identified Mesodinium rubrum as the plastid source.
Abstract
Dinophysis acuminata, the main agent of diarrhetic shellfish poisoning (DSP) worldwide, shows a high variability in morphology and toxin content between strains from contrasting habitats. Most frequent uncertainties in morphological discrimination are within the “D. acuminata complex”, but confusion with other species (e.g., D. norvegica, D. fortii) also occurs. Here we describe a unique PTX2-containing population of Dinophysis cf acuminata observed during opportunistic samplings in San Rafael Lagoon (Chilean Patagonia), the only tidewater glacier lagoon remaining in the glacier with the world’s lowest latitude. Dinophysis acuminata was the only Dinophysis species observed during three seasonal surveys in the well-mixed cold (4–7° C) and brackish (salinity 14–15) waters of the lagoon. Cell densities ranged from 500 cells L−1 (winter) to 2800 cells L−1 (summer). Partial sequences of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
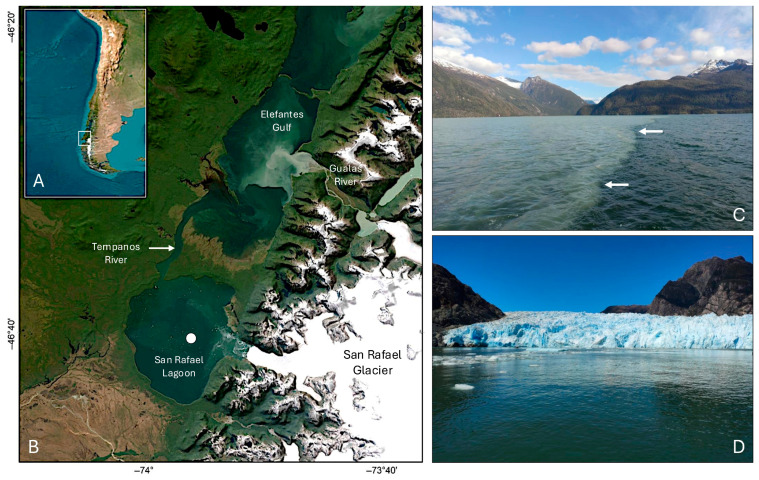 Figure 1
Figure 1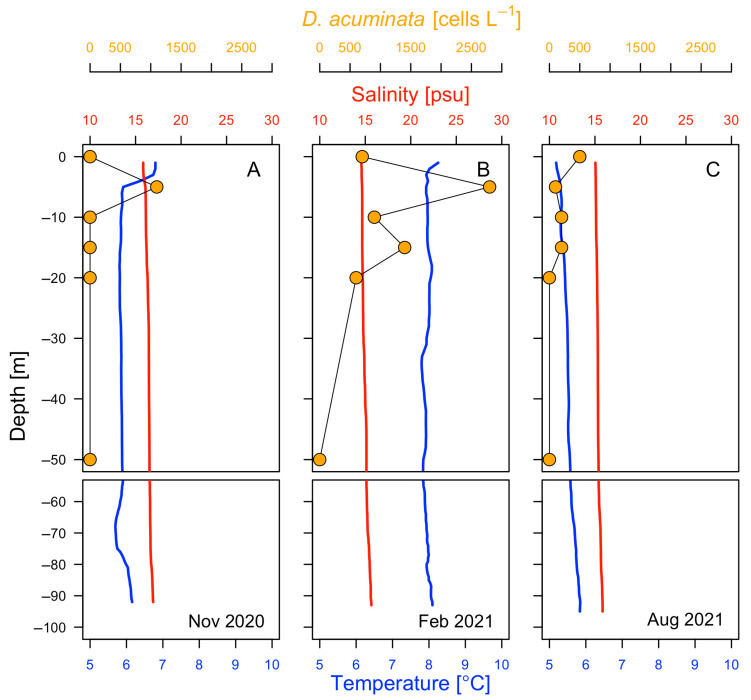 Figure 2
Figure 2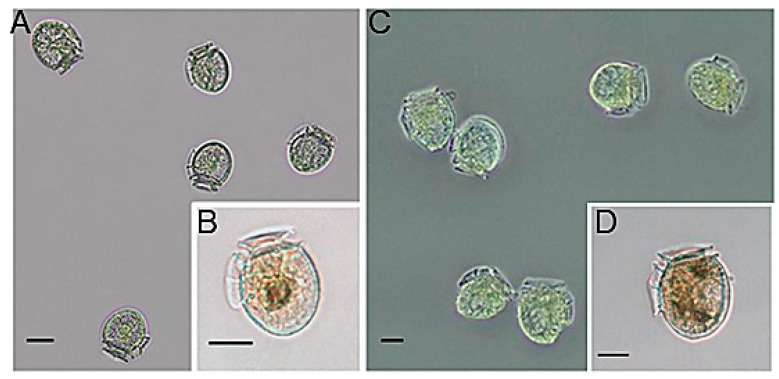 Figure 3
Figure 3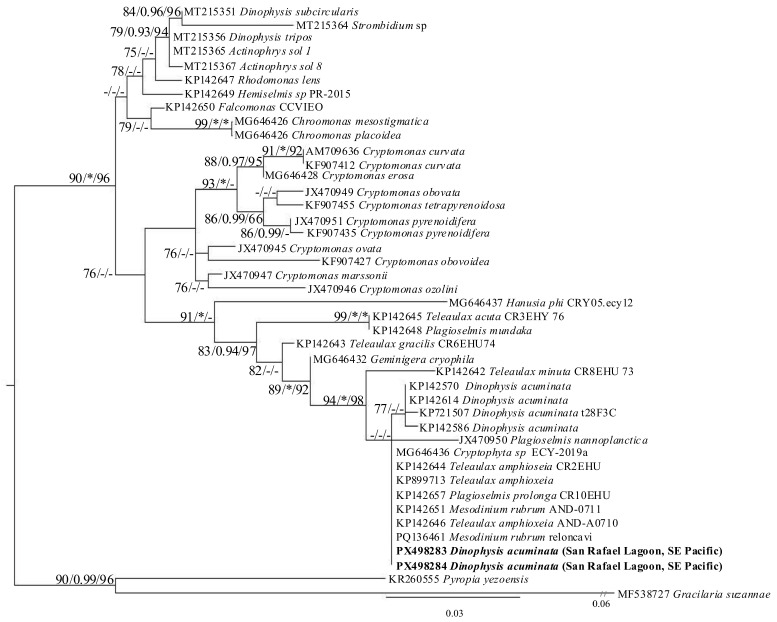 Figure 4
Figure 4 Figure 5
Figure 5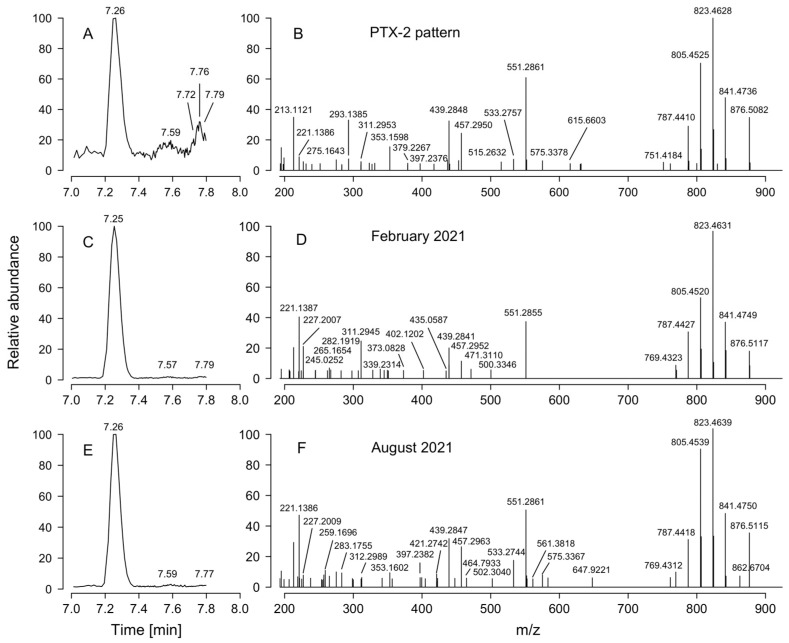 Figure 6
Figure 6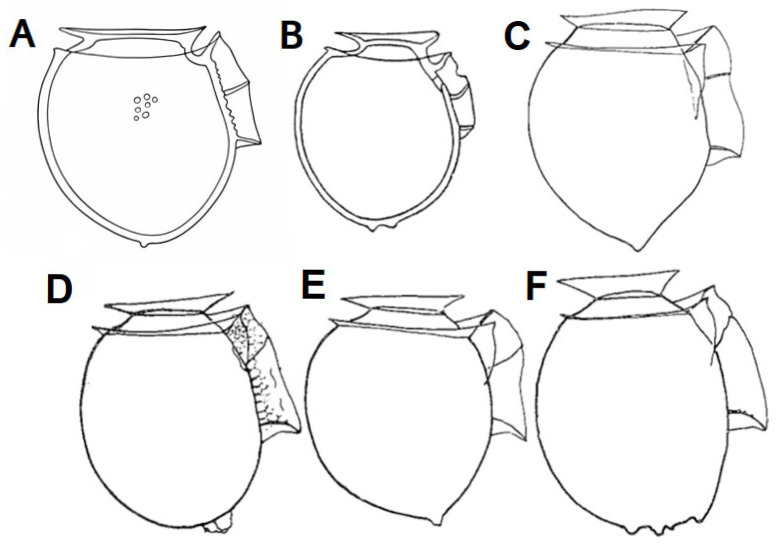 Figure 7
Figure 7| Tolerance | Cell Maximum TS | Location | Geographic | References | ||
|---|---|---|---|---|---|---|
| Temperature (°C) | Salinity | Temperature (°C) | Salinity | |||
|
| ||||||
| 12.5–22.0 | >33.5 | 13.0–16.0 | 31–34 | NW Spain | 43° N | [ |
| 13.6–19.7 | 32.0–35.4 | 17.0–18.0 | 32.0–33.0 | 42°15′ N; 8°50′ W | [ | |
| 15.0–18.0 | 32.0–34.4 | 17.0–18.0 | 32.0–33.0 | 42°23′ N; 8°45′ W | [ | |
| 16.0–18.0 | <35.5 | 17.0 | 35.1 | ~39°30′–42°30′ N; 09° W | [ | |
| 13.0–18.0 | 22.0–32.0 | 23.0–26.0 | W Sweden (Fjords) | 58°13′ N; 11°33′ E | [ | |
| 15.0–19.0 | 29.0–34.0 | 19.0 | 30.0–33.0 | Netherland (N Sea) | ~52° N; ~6° E | [ |
|
| ||||||
| 7.0–8.0 | 6.7 | NE Sweden | ~58° N; ~18° E | [ | ||
| 5.5–21.0 | 5.5–6.9 | ~18.0 | 5.5–6.9 | NE/NW Finland | ~59° N; ~22° E | [ |
| 5.5–21.0 | 5.5–6.9 | 12.0 | 5.5–6.9 | |||
|
| ||||||
| 11.0–22.0 | 29.0–38.0 | 11.5–12.5 | 36.0–38.0 | E Aegean Sea | ~40° N; ~22° E | [ |
|
| ||||||
| 7.2–17.1 | 25.3–34.9 | 10.4 | 34.4 | Loch Ewe Scotland | 57°50′ N; 5°36′ W | [ |
| – |
| |||||
| 10.0–29.4 | 22.7–34.0 | 18.0 | 28.0 | SE Sea Japan | ~35° N; ~135° E | [ |
| 17.3 ± 3.9 | 32.7 ± 0.85 | N Japan | ~39° N; ~141° E | [ | ||
| 13.0–26.9 | 27.6–34.1 | 16.0–18.0 | 32.0–33.0 | SW Japan | 33° N; 135° E | [ |
|
| ||||||
| 11.1–26.6 | 17.7–25.2 | 11.1–26.6 | 23.8 | E USA, New York | 40°53′ N; 73° W | [ |
| 8.7–26.7 | 8.2–26.7 | 13.0–24.0 | 40°53′ N; 73° W | [ | ||
|
| ||||||
| 17.0–19.0 | 23.0–29.0 | ~18.0 | ~25.0 | NW Brazil | 26°47′ S; 48°37′ W | [ |
| 5.7–18.4 | 31.2–34.2 | ~8.0–13.0 | SW Argentine | ~38–56° S; 57–69° W | [ | |
| 22.0–26.0 | 25.0–32.3 | 26.0 | 30.2 | SW Uruguay | ~30–35° S; ~53–58° W | [ |
|
| ||||||
| 10.5–19.6 | 4.7–30.73 | 11.19–12.52 | 19.75–22.59 | 43°47′ S; 72°56′ W | [ | |
| 10.2–16.9 | 17.4–32.1 | 16.1–16.5 | 17.4–20.0 | N Chilean fjords | 44°36′ S; 72°48′ W | [ |
| 10.8–18.8 | 8.4–32.0 | 12.6 | 31.3 | 41°36′ S; 72°24′ W | [ | |
| 5.2–8.2 | 15.0–16.0 | 8.2 | 15.0 | San Rafael Lagoon | 46°40′ S; 73°55′ W | This work |
| 5.8–10.4 | 23.8–32.5 | 8.0 | 30.6 | S Chilean fjords | 54°49′ S; 70°12′ W | [ |
|
| ||||||
| 13.8–17.0 | 33.8–34.3 | 14.6–14.8 | 34.2–34.3 | NE New Zealand | ~41°20′ S; 174°07′ E | [ |
| Toxin | Abbreviation | Molecular | Diagnostic | Calculated | CE |
|---|---|---|---|---|---|
| Okadaic acid | OA | C44H68O13 | [M − H]− | 803.4587 | 55 |
| Dinophysistoxin-1 | DTX-1 | C45H70O13 | [M − H]− | 817.4744 | 55 |
| Dinophysistoxin-2 | DTX-2 | C44H68O13 | [M − H]− | 803.4587 | 55 |
| Yessotoxin | YTX | C55H82O21S2 | [M − 2H]2− | 570.2322 | 34 |
| homo-Yessotoxin | homoYTX | C56H84O21S2 | [M − 2H]2− | 577.2401 | 34 |
| Azaspiracid-1 | AZA-1 | C47H71NO12 | [M + H]+ | 842.5049 | 34 |
| Azaspiracid-2 | AZA-2 | C48H73NO12 | [M + H]+ | 856.5206 | 34 |
| Azaspiracid-3 | AZA-3 | C46H69NO12 | [M + H]+ | 828.4893 | 40 |
| Gymnodimine | GYM | C32H45NO7 | [M + H]+ | 508.3421 | 34 |
| Pinnatoxin-G | PnTX | C42H63NO7 | [M + H]+ | 694.4677 | 50 |
| 13-desMe-SPX C | SPX-1 | C42H61NO7 | [M + H]+ | 692.4521 | 34 |
| Pectenotoxin-2 | PTX2 | C47H70O14 | [M + H2O]+ | 876.5104 | 30 |
- —Environmental control of Dinophysis bloom development and their contribution to lipophilic toxin events in the Chilean Patagonia
- —FONDECYT
- —COPAS COASTAL
- —CIEP
- —FONDECYT
- —ANID-FONDECYT
- —Galician Networks of Excellence
- —Spanish Ministry MICIU
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine Toxins and Detection Methods · Marine Invertebrate Physiology and Ecology · Aquatic Ecosystems and Phytoplankton Dynamics
1. Introduction
Among a variety of harmful algal blooms (HABs) and impacts, blooms of toxin-producing microalgae (toxic HABs) are the most damaging to public health (human poisonings) as well as to the shellfish (harvesting closures), fish (mass mortalities) and aquaculture industries. Some microalgae produce potent toxins which even at low cell concentrations are accumulated and transformed by filter feeders and transferred through food webs [1,2]. This “particulate” transmission of toxins was formerly considered the only way harm was caused during paralytic (PSP) and diarrhetic shellfish poisoning (DSP) events. Evidence has accumulated in the last 20 years of extracellular excretion or leakage of toxins into the environment [3,4]. In laboratory experiments, the extracellular fraction may comprise 65–90% of the total toxin produced by Dinophysis [5,6]. In other words, measuring toxins in particulate form underestimates the real amount of toxins produced and the potential to cause harm to marine organisms [7,8].
MacKenzie et al. [9] pioneered the use of passive samplers or “SPATT” (solid-phase adsorption toxin-tracker) resins which adsorb the “dissolved toxins” fraction, dissolved meaning here the fraction of toxins which are not retained after filtering seawater through 0.22 µm pore-sized filters. These toxins are actively released or simply leak into the environment and when found free outside the cells (detectable using the appropriate resin sampler) can affect a much larger range of organisms, from protists to invertebrates [8]. They can also adhere to organic debris and sediment and be detected in plankton concentrates when the toxic cells are no longer present; they may also become available to bottom-dwelling detritivorous invertebrates and browsers [4,10].
Accumulation of toxins in bivalve mollusks (cultures and shellfish beds) exposed to endemic blooms of Dinophysis represents the main threat to sustainable exploitation of shellfish resources in Europe [11] and northeastern Japan [12,13] and is an emergent hazard in the US [14]. Diarrhetic shellfish toxins affect consumers of shellfish collected without sanitary control and cause lengthy harvesting bans whenever toxins in shellfish flesh exceed safe limits enforced by regulations [15].
The genus Dinophysis Ehrenberg (1841) comprises over a hundred species worldwide [16], of which 10 have been associated with shellfish poisoning [15]. Toxic species of Dinophysis produce one or two groups of lipophilic toxins: (i) okadaic acid and derivatives (the diarrhetic shellfish toxins, DSP) and (ii) pectenotoxins [15]. Pectenotoxins (PTXs) were traditionally included with okadaates in the DSP complex because they are both produced by Dinophysis and are coextracted with lipophilic solvents used for standard toxin tests [17]. Okadaic acid and derivatives (Dinophysistoxins) are protein-phosphatase inhibitors and the only ones with diarrhetic effects [18,19]. Pectenotoxins may kill mice when administered via intraperitoneal injection and cause hepatotoxicity but are harmless by oral intake [20,21]. Based on recent toxicological studies, the World Health Organization consider that PTXs do not pose a risk to human health, and this group of toxins is no longer under regulation. Nevertheless, pectenotoxins are actin-polymerase inhibitors which affect cellular motility [22], and there is a growing number of studies in vitro about the noxious effects of PTXs in early larval stages of bivalves and fish [23,24]. Concerning subacute effects on human health, okadaic acid and derivatives are considered to be tumour promoters [25], while PTX2 has been found to have a selective apoptotic effect on carcinogenic cells [26,27]. Thus, the same organisms (Dinophysis species) produce both tumour promoters and suppressors.
Dinophysis species, in particular those belonging to the “Dinophysis acuminata complex”, are endemic in many coastal areas with relevant shellfish resources [15]. Monitoring Dinophysis spp. may be difficult for various reasons. First, they develop low biomass blooms. Concentrations of a few hundred cells per litre have been associated with toxicity in shellfish, and cell numbers above 10^3^ cell L^−1^ are considered a bloom [17,28], but they can be present most of the year below the detection levels (40–100 cells L^−1^) of traditional monitoring programmes. Second, they show a high intraspecific morphological and toxinological variability, i.e., differences in size and shape and in toxin profile and content can be higher between strains of the same species than between different species from the same location. Lastly, there is low inter-species variability in the nuclear (LSU, SSU and ITS regions) and mitochondrial gene (cox1 and cob) sequences normally used to discriminate between morphologically close species and applied in the design of molecular probes for their identification [29,30,31,32]. Considering that the morphological characteristics commonly used to diagnose Dinophysis species are the shape, relative size, and ornamentation of the large hypothecal plates [33,34,35], species identification can be cumbersome when morphologically similar species, each one with their array of morphotypes, co-occur in the same sample, and molecular probes do not help much to solve the uncertainties.
Key issues to predict toxic blooms of Dinophysis species and the subsequent contamination of shellfish are related to the identification of their nutritional sources (ciliate prey) and life history strategies (overwintering cells and origin of the inoculum or “population source”) [36].
Dinophysis species are “plastidic specialist non-constitutive mixotrophs” (pSNCM), i.e., their plastids are not constitutive, but stolen (kleptoplastids) from very selected prey (specialists) [37]. A three-species food chain—D. acuminata fed the ciliate Mesodinium rubrum, the later fed cryptophyte microflagellates—was established in laboratory cultures two decades ago [38] and later used with other toxic Dinophysis (D. acuta, D. caudata, D. fortii, D. infundibulus, D. norvegica, D. sacculus and D. tripos) [39,40]. However, there are records of field populations of Dinophysis with kleptoplastids outside the Teleaulax/Geminigera clade which suggest the possibility of alternative prey [41,42,43]. Given the mixotrophic nature of Dinophysis species, it is crucial to add its confirmed ciliate prey (Mesodinium rubrum + major complex or any other suspected plastid-bearing ciliate) to the growth equation for a sound bloom prediction model. But so far, few Dinophysis-focused surveys have included analyses of the accompanying potential prey (plastid bearing microciliates) nor of the plastid sequences of both, predator and suspected prey, to confirm their common cryptophyte origin [44,45,46,47,48].
In addition, unlike other HAB dinoflagellate species (e.g., Alexandrium and Protoceratium), Dinophysis species do not seem to rely on benthic stages (sexual cysts) for seeding. Their complex polymorphic life cycle includes sexual processes and formation of planozygotes. These mobile diploid planozygotes can divide to produce vegetative cells, short-cutting the sexual-cyst stage [49]. Thus, the whole life cycle comprises free-swimming motile forms (holoplanktonic life cycle) which occur in the water column; the next season inoculum will be formed by free mobile forms which overwintered in some quiescent state, dispersed in the water column or in some unknown retention area acting as a “pelagic seed bank”. This term was coined by Smayda [50] to describe the accumulation of dinoflagellates in upwelling fronts like “pelagic seeds” able to act as inocula for future populations when advected to the coast during downwelling.
This paper describes a singular population of Dinophysis acuminata found in San Rafael Lagoon, a remote relatively unexplored tide-water glacier lagoon of pristine waters and spectacular nature (Figure 1). San Rafael Lagoon, site of a reduced colony of leopard seals (Hydrurga leptonyx) and other protected fauna, is part of the San Rafael Glacier National Park [51], designated a World Biosphere Reserve by UNESCO in 1979. This national park, located in the southernmost sector of Northwestern Chilean Patagonia [52], i.e., in a temperate latitude, is site of the Northern Patagonian Ice Field (3500 km^2^), the third largest in the southern hemisphere, after Antarctica and the Southern Patagonian ice fields. These features make San Rafael Park and its water bodies a highly sensitive observatory for the effect of climate change in glacier dynamics [53,54]. Nevertheless, there have been only a few attempts to study the plankton communities (mainly zooplankton) of San Rafael Lagoon [55], an extreme environment formed by the retreat of its homonymous glacier. There is only one survey in the early 1990s, during the international Raleigh Expedition, describing a low diversity net-phytoplankton community comprised by a few diatoms species [56].
Dinophysis specimens described here are from opportunistic sampling carried out during seasonal oceanographic surveys to study the hydrography and water mass circulation of the lagoon and adjacent water bodies. In the framework of a One Health approach to the study of toxic algal blooms, the one-way effect of toxins in humans is not the only harm to be considered. The remoteness of San Rafael Lagoon, with neither aquaculture nor exploited shellfish beds, the absence of anthropogenic pollution, stable environmental conditions, and the occurrence during different season surveys (including winter) of only one species of Dinophysis, Dinophysis cf acuminata, make it a natural mesocosm for ecophysiology and population dynamic studies of a cosmopolitan Dinophysis species. It can be taken as a model case study to explore the effect of bioactive (particulate or extracellular) compounds of Dinophysis on a low diversity local fauna permanently exposed to a single lipophilic toxin producer under stable environmental conditions.
Map showing: (A) Chile. Box delimits the Northern Patagonia, Aysén province; (B) San Rafael Glacier National Park comprising the Northern Patagonian Ice Field, San Rafael Lagoon, and location of the sampling station, Témpanos “River” channel and the “Elephants Gulf”; photographs of the (C) Frontogenesis in Elefantes Gulf (white arrows mark the salinity front and (D) San Rafael Glacier Lagoon.
2. Results
2.1. Hydrographic Conditions and Dinophysis Distribution
Vertical distribution of physical properties was measured with CTD casts in spring (November 2020), summer (February 2021) and winter (August 2021). There were light differences in temperature (T) and salinity (S) between surface and bottom (>90 m) waters in summer (T: 8.26 °C and S: 14.56 at the surface; 8.09 °C and 15.67 at 93 m) and winter (T: 5.18 °C and S: 15.5 at surface; 5.84 °C, 15.84 at 95 m). In spring, there was a mild thermal gradient between 0 m (6.8 °C) and 5 m (6.0 °C), and a uniform value from 5 m (6.0 °C) to 92 m (6.16 °C). In winter, there was a very mild thermal inversion from >70 m depth to the bottom (Figure 2). Thus, the lagoon offers a cold (5.2 to 8.3 °C) and brackish (14.6–16.9) but very stable environment with narrow ranges of variability.
Dinophysis cf acuminata was the only species of Dinophysis present in samples collected during the three seasonal surveys. Cells occurrence in winter (500 ± 445 cells L^−1^) at the surface was well above the detection limits in conventional monitoring programmes using microscopic counts following Utermöhl [57] (40–100 cells L^−1^). Moderate bloom levels occurred in spring (2800 ± 980 cells L^−1^) and summer (1000 ± 630 cells L^−1^). Vertical distribution of Dinophysis cells varied seasonally; cell maxima were found at the surface in winter and in a sharp maximum at 5 m in spring and summer. This 5 m depth coincided with the base of a weak (maybe diurnal) thermocline observed in the CTD profiles (Figure 2). Low vertical resolution sampling (bottles every 5 m) precluded the possibility to explore the vertical microstructure and determine if Dinophyis cells were actively aggregated in a thin layer. It is not certain that the observed peak in Dinophysis cells coincided with the maximal intensity of a hypothetical thin layer.
Vertical profiles of temperature (°C), salinity (CTD casts) and cell density of Dinophysis acuminata (cells L−1) during (A) Spring, (B) Summer and (C) Winter surveys in San Rafael Lagoon (Aysén, Chile).
2.2. Morphological Observations
The micrographs shown here are from summer (February) samples, the only season with a relevant occurrence of the small forms. Dinophysis specimens observed in San Rafael Lagoon exhibited a quite regular and constant shape (contour of the large hypothecal plates) and two distinct size-classes or morphotypes during the three seasonal (spring, summer and winter) samplings (Figure 3). The large forms were subovate and slightly asymmetric, and the large hypothecal plates were well ornamented and had a coarse appearance which resembled that of the cold temperate species Dinophysis norvegica. Cell measurements were L (length) = 43.53 ± 2.32 µm and D (dorso-ventral depth) = 39.03 ± 2.32 µm, n = 44; the L:D ratio was 1.12 ± 0.05. The smaller forms were more oval and symmetric, and measurements were L = 27.68 ± 3.41 µm and D = 25.90 ± 3.65 µm wide, n = 9; the L:D ratio was 1.07 ± 0.08 (Figure 3).
Light micrographs of Lugol’s preserved (A,B) small and (C,D) vegetative Dinophysis cf acuminata cells from San Rafael Glacier Lagoon in late February 2021. Scale bar = 20 μm in all frames.
2.3. General Alignment of 23S rDNA Sequences and Phylogenetic Analyses
The alignment of the plastid-derived sequences of 23S rDNA comprised 43 sequences with a total aligned length of 381 bp (target sequences in this study, 2_23S_San Rafael_Dinophysis PX498283 and 6_23S_San Rafael_Dinophysis PX498284 were 312 and 306 bp long, respectively). The dataset included 55 parsimony-informative sites, 49 singleton sites, and 277 constant sites. ModelFinder selected the TPM3u + G4 model as the best-fit substitution model according to the Bayesian Information Criterion (BIC). The maximum log-likelihood value obtained for the final tree was −1533.570.
The resulting maximum-likelihood (ML) phylogeny recovered the main cryptophyte lineages, including Teleaulax, Plagioselmis, Geminigera, Cryptomonas, Chroomonas, Rhodomonas, and Hemiselmis. As these sequences are plastid-derived, the resulting phylogeny reflects the cryptophyte origin of the kleptoplastids retained by Dinophysis cells rather than the phylogenetic position of the Dinophysis host itself. The two plastid-derived sequences obtained from Dinophysis (SH-aLRT/aBayes/UFBoot = 91.1/1/86) clustered within the Teleaulax–Plagioselmis-Geminigera assemblage (e.g., MG646432, KP142648, KP142644, KP142657, KP142646, KP899713). The two sequences obtained in this study were grouped in a subclade (94.1/1.00/98) together with T. amphioxeia and Plagioselmis sequences. Both Dinophysis-derived 23S rDNA sequences showed very high similarity to these cryptophyte sequences: 2_23S_San_Rafael_Dinophysis was identical (100%) to Teleaulax amphioxeia (KP142644 and KP899713) and Plagioselmis prolonga (KP142657), while 6_23S_San_Rafael_Dinophysis shared 98.7% identity with the same sequences. These results confirm that the plastids in Dinophysis cells from San Rafael belong to a Teleaulax/Plagioselmis-type cryptophyte lineage (Figure 4).
Maximum likelihood phylogenetic tree inferred from 23S rDNA sequences, including sequences of Dinophysis species plastids. Pyropia yezoensis and Gracilaria suzannae were used as outgroups. The 23S sequences of Dinophysis cells obtained in this study are in bold. The scale bar represents inferred evolutionary distance in changes/site. Node support represents Shimodaira–Hasegawa approximate likelihood ratio test (SH-aLRT), approximate Bayesian (aBayes) and ultrafast bootstrap (UFBoot) values. Only the values ≥ 70%/≥0.90/≥90% (SH-aLRT/aBayes/UFBoot, respectively) are shown. Asterisks represent maximum statistical support (100%/1/100%).
2.4. General Alignment of ITS rDNA Sequences and Phylogenetic Analyses
A total of 71 ITS rDNA sequences were included in the final alignment, which had a length of 646 bp. ModelFinder selected TIM2 + F + G4 as the best-fitting nucleotide substitution model according to the BIC. The ML tree inferred under this model had a log-likelihood of –3560.7830. The seven ITS sequences obtained in this study were highly similar and clustered within the Dinophysis acuminata complex clade.
Individual untrimmed sequence lengths ranged from 451 to 560 bp (PX494338 D_ac_8: 512 bp; PX494339 D_ac_9: 451 bp; PX494340 D_ac_R6: 534 bp; PX494341 D_ac_R8: 513 bp; PX494342 D_ac_S1: 514 bp; PX494343 D_ac_S2: 560 bp: PX494344 D_ac_S3: 548 bp), based on nucleotide positions excluding gaps. Pairwise nucleotide identities among the seven sequences ranged from 99.56% to 100%, with at most 2 nt differences between any pair of sequences. ML patristic distances were likewise very small (from 3 × 10^−6^ to 0.00563 substitutions per site), confirming extremely low ITS divergence within the group.
The ITS rDNA phylogeny (Figure 5) showed the characteristic pattern reported for the genus Dinophysis, in which several species form closely related lineages and deeper relationships remain only partially resolved due to the low genetic divergence and extensive rDNA overlap within the group. The D. acuminata complex appears as a genetically heterogeneous assemblage that includes sequences identified as D. acuminata and D. norvegica, with the latter forming a well-supported internal subclade (AJ506982, OM939691, EU927486, OM939684, AJ506985, MK860873).
Notably, additional sequences identified as D. ovum (GU452504, AM931581) and D. sacculus (MT365103, AJ012007, PP270373) also fall within this complex, reflecting the lack of clear ITS boundaries among members of the D. acuminata–D. sacculus–D. ovum–D. norvegica assemblage.
Sequence AY040569, deposited as D. acuta but placed within the D. acuminata complex clade, displayed 99.4–99.8% identity to the seven D. acuminata sequences obtained in this study (only 1–3 nucleotide differences). In contrast, its identity with the other two sequences of D. acuta clustered separately from D. acuminata complex (KF871424 and MT365112) was markedly lower (92.1–93.4%). These results strongly indicate that AY040569 was misidentified and likely corresponds to a D. acuminata specimen rather than to D. acuta.
In the ITS rDNA tree, the node with 99/1/99 (SH-aLRT/aBayes/UFBoot) support corresponds to the D. norvegica clade. Our sequence PX494341 (identified as D. acuminata) does not fall within this group but appears as its immediate sister lineage. This topology reflects the very short genetic distances between D. acuminata and D. norvegica (Figure 5).
In addition, pairwise comparisons showed that PX494341 shares 96.3–98.6% identity with the D. norvegica sequences but a higher identity of 99.6% with both OM939692 and PX494342, which were identified as D. acuminata. PX494341 belonged to DNA extracted from five cells corresponding to a small-sized morphotype (27.68 ± 3.41 µm length × 25.90 ± 3.65 µm width), whereas the remaining isolates corresponded to a larger morphotype (43.53 ± 2.32 µm length × 39.03 ± 2.32 µm width). Despite these morphological differences, D. acuminata PX494341 grouped with the larger morphotype sequences and exhibited an almost identical nucleotide composition.
Maximum likelihood phylogenetic tree inferred from ITS rDNA, including sequences of Dinophysis species. Three sequences within genus Prorocentrum (P. donghaiense, P. mexicanum and P. rathymum) and two sequences within genus Phalacroma (P. rotundatum and P. rapa) were used as outgroups. The ITS sequences of Dinophysis cells obtained in this study are in bold. The scale bar represents inferred evolutionary distance in changes/site. Node support represents Shimodaira–Hasegawa approximate likelihood ratio test (SH-aLRT), approximate Bayesian (aBayes) and ultrafast bootstrap (UFBoot) values. Only the values ≥ 70%/≥0.90/≥90% (SH-aLRT/aBayes/UFBoot, respectively) are shown. Asterisks represent maximum statistical support (100%/1/100%).
2.5. Particulate Toxin Analysis
LC-MS analyses of lipophilic toxins in extracts from plankton net-tows collected in summer (February) and winter (August) 2021 revealed the only presence of pectenotoxins (PTX2). A chromatographic peak was detected at 7.26 min with a parent mass [M-H] + 876.5082 m/z which corresponded to PTX2; its presence was confirmed by its characteristic MS/MS fragment at m/z 841.4736, 823.4628, 805.4525 and 787.4401 m/z (Figure 6).
In February 2021, total PTX-2 was 50.2 ng NT^−1^, whereas in August 2021 the total content of the same toxin was slightly lower (35.9 ng NT^−1^). Assuming a fluent (without deflections and/or clogging) passage of seawater through the vertical haul, these values per net-tow would be equivalent to 80 pg PTX2 L^−1^ and 57 pg of particulate PTX2 L^−1^ in February and August, respectively.
LC-MS chromatograms with their retention times (left) and mass spectra (right) of pectenotoxin-2 detected in: (A,B) PTX-2 standard pattern; (C,D) net-tow from February 2021 and (E,F) net-tow from August 2021.
3. Discussion
Dinophysis acuminata is the most widely distributed species of the genus Dinophysis and the most frequently cited agent of diarrhetic shellfish poisoning. This work describes a population of D. cf acuminata which is unique for various reasons: first, its uniform morphology (cell contour) in samples collected in three seasonal surveys (winter, summer and autumn); second, its persistence in moderate cell numbers even during the winter months when the species is under detection limits in most of its usual growing areas. Lastly, it is the only species of Dinophysis which seems to be adapted to the location under study. This singular population is reported from a unique place, Laguna San Rafael, a tidewater glacier lagoon with boreal-like waters but located in a temperate latitude and with very stable conditions.
3.1. Biogeographic Considerations
Dinophysis acuminata is considered a euryhaline and eurythermal species with a capacity to grow under a wide range of salinity and temperature values [40]. This general statement often results from a simple association of its occurrence with physical conditions (temperature, salinity, light) which do not always correspond to those in the depth range where the species grows actively (intrinsic growth rate) or where the population has been entrained and transported (from shelf waters to the coast or vice versa). Furthermore, intraspecific differences in adaptations between strains from different latitudes or systems can be as large as between two different species (Table 1).
In a niche-based study of the phytoplankton community in Ría de Vigo, Velasco-Senovilla et al. [79] found that D. acuminata is very tolerant and at the same time exhibits high marginality. In other words, it is able to endure a wide range of adverse conditions, including absence of prey up to two months [80], but population growth occurs within a very narrow environmental window [47,81,82,83]. The diverse set of conditions where species of the D. acuminata complex bloom (Table 1) suggests there is a variety of strains adapted to endure very different ranges of temperature and salinity. But population increases occur within narrow ranges related to different growth seasons and in response to site-specific triggers. The Baltic Sea shows the lowest temperature and salinity limits and the highest latitude [63,64], but there are reports from unmonitored higher latitude areas visited during ad hoc surveys and scientific expeditions (see “Dana” in Section 3.3). The lower limit for growth (~5 °C) agrees with the temperature identified as the lower limit for D. acuminata division in laboratory cultures [84].
3.2. Considerations on Morphological Variability in Dinophysis cf acuminata
D. acuminata exhibits highly variable morphology. Size and shape of its large hypothecal plates and repartition of the left sulcal lists between daughter cells are affected by the stage in the life cycle (small cells produced by depauperating division) and by the species’ vegetative division by desmoschisis as well as the nutritional status of the cell (a range of sizes from well-fed vacuolate cells to thin starving specimens) [85]. There is also an intraspecific morphological variability which has been explained on the basis of biogeographic differences, but there is controversy on whether these differences are inherited traits or phenotypic responses to environmental conditions which can be stimulated in the laboratory with acclimation experiments [31]. In any case, cosmopolitan species with broad niches present morphological differences between strains growing in different latitudes or hydrodynamic systems. This issue has been well illustrated in the case of species of the genus Tripos and the variability (size and shape) of their apical and lateral horns [86].
Among a dozen well-known species of Dinophysis, the magnitude of their morphological variability as a source of uncertainty in taxonomic identification has been biased by their toxic potential and the location of the impacted aquaculture sites. The” Dinophysis acuminata complex” includes various morphotypes of D. acuminata, D. sacculus and D. ovum, which may have overlapping distributions and are major contributors to DSP events in temperate latitudes. These three taxa cannot be distinguished with conventional rDNA probes applied for species discrimination. Nevertheless, the same or higher uncertainties can be met with other “Dinophysis complexes” which include more than one recognized species of Dinophysis (e.g., “D. caudata/D. tripos” and “D. acuminata/D. norvegica” complexes) and have not drawn much attention.
3.3. Taxonomical Assignation and Toxin Profile of Dinophysis from San Rafael Lagoon
Dinophysis specimens collected in San Rafael during three different seasons showed a very regular shape. The morphotypes described here, with their coarse and rotund appearance, are reminiscent of some morphotypes described by Paulsen in his section of “Icelandic Dinoflagellates. On Northern Dinophysis and Phalacroma species” collected in Icelandic fjords (>68° N) during the Dana expedition [87].
In a vertical net-haul from Patreks Fjord, Paulsen probably found a multispecific bloom of D. norvegica, D. acuminata and a few more Dinophysis species, each one presenting an array of cell cycle- and life cycle-related forms. The listed taxa included Dinophysis artica; D. islandica; “Acuminoide Dinophysis” (all these with antapical protuberances as in D. acuminata, D. borealis, D. lachmanni), and D. skagii, and common descriptive terms for D. norvegica included “Atlantic robust forms”, “pointed/obtusa/angulata forms” as well as currently used forms (debilor, crassior). But different taxa, drawings and proposed names were not equally adopted by the most influential taxonomists. D. borealis was included in a group of Norwegian Dinophysis species examined (pore theca morphology) by Balech [88], who found them in samples from cold temperate to subarctic latitudes (between Oslofjord and Tromsø) (Figure 7). Balech [88] considered there were no reasons to keep D. borealis as a separate species from D. acuminata Claparède and Lachmann and considered them to be synonyms. Thus, D. borealis corresponds to broad D. acuminata morphotypes (L: 39–53, L:D ratio 1.07–1.18) with a quite rounded antapical end, with or without antapical nodes or protuberances. Later he renamed the broad morphotypes, illustrated with the same Norwegian specimen outlines, as D. acuminata var acuminata [89]. The small size-class observed in summer samples from San Rafael Lagoon is compatible with a small cell formation process during depauperating division of the vegetative cells [90]. These small cells behave as male gametes and form anisogamous mating-gamete pairs with larger cells [49]. Production of small cells seems to be a density-dependent process [79] and is maximal at the peak of annual population growth [91]. Recently, in situ imaging technology (Imaging FlowCytobot) has provided evidence of a mass production of small cells in populations of D. acuminata aggregated in thin layers preceding mating and formation of planozygotes (2n) with two trailing flagella [92]. This sexual division is segregated in time from the vegetative division at dawn and confirmed early observations by MacKenzie [93] in field populations of D. acuta in Marlborough Sound, New Zealand.
(A) Dinophysis islandica (modified from Figure 11S in [87]); (B) Dinophysis borealis (modified from Figure 14U in [87]); (C) Dinophysis norvegica (modified from Figure 2G in [88]); (D–F) Dinophysis acuminata (modified from Figures 9–11 in [89]).
D. acuminata morphotypes from San Rafael Lagoon are quite different from the more oval and smooth D. ovum, a member of the D. acuminata complex predominant in high density (>5 × 10^4^ cells L^−1^) blooms reported in southern Brazil [94], Uruguay [74] and off Buenos Aires [95], all these subject to the influence of the plume from La Plata River. They also differ from D. fortii, with its characteristic straight ventral margin, misidentified as a member of the D. acuminata complex in samples from southeastern Brazil [96]. Lastly, they are clearly distinct from previous observations of D. acuminata forms in field samples along the Chilean coasts, including the southernmost Chilean fords in the Magallanes province (Díaz et al. [97] and Table 1), as well as in field and cultured specimens from the Patagonian Pacific coast (off Chiloé Island and outer fjord waters) shown in Paredes-Mella et al. [98].
Phylogenetic analyses of the ITS region clustered the sequences of San Rafael specimens, both small and large morphotypes, with those of D. acuminata from North America, western Japan, Australia and Europe and with other morphologically close “species” included in the D. acuminata complex (i.e., D. ovum and D. sacculus). Therefore, the morphological variations observed between the small and large morphotypes and between other forms included in the D. acuminata complex do not seem to correspond to genetic differentiation, suggesting they all belong to the same genetic lineage within the D. acuminata complex.
Several studies have drawn attention to the lack of an appropriate molecular tool to discriminate between morphologically close species of Dinophysis. Sequences of genes used as genetic markers in eukaryotes (i.e., nuclear LSU and SSU rDNA, ITS, and mitochondrial cox1 and cob genes) show little variability between different species of Dinophysis. Problems to identify species grouped in the “D. acuminata complex continue 35 years after the first mention [99]. But, this is not the only “complex” within the genus Dinophysis. Phylogenetic analysis of the ITS rDNA tree here showed D. norvegica grouped in a subclade very close to D. acuminata as its immediate sister lineage. A very short genetic distance between D. acuminata and D. norvegica agrees with observations from Edvardsen et al. [100] who concluded that species delineation between D. acuminata and D. norvegica using morphometrics and genetics was not always apparent and suggested that hybrids existed in Norwegian waters. The coarse texture of the hypothecal plates and broad proportions of D. acuminata specimens in the present study make some forms with slightly tapered antapical end look similar to small cells of D. norvegica (Figure 7). In fact, some D. acuminata images shown in Stern et al. [43] (e.g., Figure 2 Dph2, Dph7, Dph12) seem to be misidentifications corresponding to medium-small sized cells of D. norvegica (see D. norvegica forma crassior in Hansen [101]) and to forms labelled as D. norvegica from the Kattegat and published in Larsen and Moestrup [34] as Figure 7D,E. In Loch Ewe (57° 50′ N), a fjord in Northwest Scotland, UK, data available online of cell counts from the official HAB monitoring shows that morphologically close D. acuminata and D. norvegica are occasionally enumerated together as D. acuminata/D. norvegica complex https://data.marine.gov.scot/dataset/scottish-coastal-observatory-loch-ewe-site (accessed on 28 January 2026).
Raho et al. [30] proposed sequencing mitochondrial genes as a better marker for Dinophysis species. These also failed to resolve the “acuminata complex”, but in an extensive phylogeny tree inferred from mitochondrial cox1 gene sequences (507 bp) of Dinophysis prepared by Park et al. [31], D. norvegica and D. acuta formed individual clades and the other Dinophysis species were separated into two heterogeneous clades: the D. caudata group, which includes the large pedunculated Dinophysis species, and the “D. acuminata complex” group.
We are not in a position to make a sound taxonomic assignment of the Dinophysis morphotypes. With the available genetic information and cell contour diagnosis, we can confirm that it is a member of the D. acuminata complex and close to the broad forms classified as D. acuminata var acuminata in Balech [88].
Toxin analyses of the net-tows in which D. acuminata was the only potentially toxic species showed the presence of PTX2 as the only toxin in the extracts. No okadaates were detected. This simple profile coincides with the predominant PTX2 profile found in most D. acuminata populations along the Chilean coast [97] and in several cultivated strains from southern Chile [98]. But D. acuminata blooms associated with PTX2 and DTX1 have been occasionally reported in Southern Chile [97,102].
The results of toxin concentration per unit of volume, i.e., PTX2 in ng per net-tow (ng PTXs NT^−1^) have to be taken as semiquantitative information. Quantification of toxins in phytoplankton net-tows and extrapolation to toxin per cell can be biased by multiple factors which were not considered in this work [4]. But they can be compared with results obtained by other experts using the same plankton collection systems and applying the same analytical protocols.
During three cruises in the Argentine Sea (∼38–56° S) in years when D. acuminata occurred using the same sampling and analytical procedures [73], maximum values of toxin per net-tow were 624 ng PTX2 NT^−1^ (autumn 2012), 997 ng PTX-2 NT^−1^ (spring 2013) and 44 ng PTX-2 NT^−1^ (late summer 2013). But, the plankton nets used in those cruises had a diameter (60 cm) three times larger than ours (20 cm), so the volume of filtered seawater was 9 times higher and the toxin per unit of volume has to be divided by the same factor. Mutatus mutandi, the observed “normalized” concentrations of particulate PTX2 in plankton net tows in the Argentine Sea (2012–2013) were 69 (autumn), 111 (spring) and 5 (summer) ng PTX-2 NT^−1^ cf, 50.2 (summer) and 35.9 (winter) ng PTX-2 NT^−1^ in Laguna San Rafael in 2021.
The above estimates suggest moderate levels of PTX2 in San Rafael Lagoon. Unfortunately, the concentration of extracellular toxins was not measured. Nevertheless, a chronic exposure to moderate-low levels of toxic cells (particulate) and their exudates (extracellular toxins) may result in equally noxious effects in sensitive organisms and a relevant accumulation in filter-feeders [103].
3.4. Confirmation of Mesodinium rubrum + Major Complex as the Ciliate Prey for D. acuminata in San Rafael Lagoon
Dinophysis species are plastidic specialist non-constitutive mixotrophs (pSNCM), i.e., they need to steal plastids from a very specific prey to perform photosynthesis. Identification of the prey is a key issue in the Dinophysis population growth equation. Phylogenetic analysis of the (plastid-derived) 23S rDNA sequences confirmed their alignment with the Teleaulax/Geminigera clade. Plagioselmis is now deleted from the clade’s nomination after confirmation that it is the haploid (n) stage in the life cycle of Teleaulax amphioxeia [104]. This is the first time that plastids of wild populations of D. acuminata from Chile have been sequenced. In an earlier study, Díaz et al. [41] found a different kind of plastid in D. acuta, D. caudata, D. tripos and D. subcircularis from Chilean Patagonia. Plastid sequences in picked cells from the four species aligned with those in cryptophytes from clade V (Rhinomonas, Rhodomonas and Storeatula). That study showed a predominance of red cryptophyte plastids different from the Teleaulax/Plagioselmis/Geminigera clade in field populations of phototrophic Dinophysis spp., but D. acuminata was not included in the occurring species. Previous studies with Dinophysis species from northwestern Scotland [43] had reported the first case of phototrophic Dinophysis harbouring plastids related to the Rhodomonas/Storeatula clade.
3.5. Singularities of San Rafael Lagoon and Its Dinophysis Populations: Applications for Research and Monitoring
Monitoring and early warning of lipophilic toxin events face several issues derived from the characteristics of the toxin-producing species: (i) detection of low-density populations which may occur for months below detection levels; (ii) difficulties to discriminate between co-occurring Dinophysis species with similar morphology but different toxic potential; and (iii) identification of the inoculum source, which is not easy with holoplanktonic species which do not rely on resting cysts as a seeding strategy [36,85]. None of these issues seem to pose a problem in the unique environment of San Rafael Lagoon. Populations of D. acuminata were present well above the detection limits with microscopic cell counts using the Utermöhl [57] technique, even during the winter survey. There was a single homogeneous population of D. acuminata which did not have to be discriminated from co-occurring morphologically close species of Dinophysis. Lastly, the inoculum population was available all the time, and there was a 2–3 doublings difference between the (winter) minimum and the (summer) maximal cell densities observed.
Park et al. [31] illustrated the morphological variation in cultures of 54 strains of D. acuminata complex isolated from Korean coastal water. All the original isolates had a Dinophysis cf ovum appearance, but culture samples taken during the exponential growth phase showed a continuum of shapes from D. acuminata to D. ovum morphotypes which yielded identical sequences of the mitochondrial cox1 gene. Unfortunately, we have no information on how many days each strain was incubated before the images were taken, and all strains (regardless their isolation site) were grown with identical conditions (20 °C, salinity 30, 14:10 light:dark cycle with cool-white, fluorescent lamps providing a light intensity of 140 μmol photons m^−2^ s^−1^).
Cultured microalgal cells sometimes show important morphological differences (contours become smoother and loose sharpness) compared with wild specimens. These differences increase with days of incubation and are not subject to some field constraints (e.g., turbulence or laminar flows, shear currents, etc.) with important effects on shape but infrequently measured or related to morphological variability. But with no doubt, and until a higher resolution molecular tool is achieved, the results of Park et al. [31] support the view that the “Dinophysis acuminata complex” may be the result of phenotypic variants of D. acuminata determined by intrinsic factors, such as life cycle, cell cycle stages, nutritional status [85] and extrinsic factors, such as changing environmental conditions and the geographical location [31]. The stable environmental conditions in San Rafael Lagoon and the regular shapes observed in Dinophysis cells there give support to the hypothetical relationship between environmental stability and shape (hypothecal plate contours). The closest morphotypes in appearance to the Dinophysis from San Rafael Lagoon are the broad sub-Artic forms with coarse hypothecal plates described by Paulsen [87] and Balech [89] in areas with similar low temperatures but saltier waters. In contrast, the illustrated morphotypes from the cold and much fresher Baltic waters [64] agree with the slender forms with a delicate appearance which Balech classified as D. acuminata var. lachmannii. A possible explanation is that shape and texture in Dinophysis are mainly determined by the turbulence regime and that salinity and temperature may be proxies for other hydrodynamics features. Temperature may have an indirect effect on size: cells of the same species from colder areas are usually larger, and a relationship between temperature and division rates (which affects size) has been shown in culture experiments with Dinophysis species [84].
The D. acuminata population from San Rafael Lagoon contained only pectenotoxins (PTX2). Extracellular PTX2 has been detected with “solid phase adsorption toxin-tracker (SPATT) resins as far south as 62° S in summer, off the South Shetlands (Antartica) (1.5 °C and S = 34) associated with scattered cells of D. norvegica and D. acuminata [105]. The recent deregulation of pectenotoxins by the European Union Commission [106] has been a relief for shellfish growers exporting bivalves to Europe provided they use current standard LC-MS equipment for their toxin testing. Nevertheless, regions with shellfish resources which continue using mouse bioassay for toxin tests may suffer prolonged false positives and economic losses if their local strains have a simple PTX-predominant toxin profile. Notwithstanding the new scenario concerning Dinophysis toxins, growing evidence of the harmful effects of PTXs on early larval stages of marine organisms in vitro needs to be supported by observations in the field.
San Rafael National Park is protected from human activities affecting environmental health, and there is no shellfish exploitation in the lagoon, but a unique wild fauna. A thorough inventory of the local fauna, in particular the recruitment processes in larval stages of fish and invertebrates, would shed light on species sensitivity or tolerance of environmental levels of PTX. In summary, Laguna San Rafael, the only tide-water glacier lagoon remaining in the Northern Patagonian Ice Field, could be used as a privileged natural mesocosm and as a baseline to study the effect of chronic exposure to low levels ofPTXs in marine organisms. Progress could be achieved too on extreme habitat conditions affecting biodiversity and limiting (or selecting) determined species of interest, such as Dinophysis, their accompanying species, as well as their competitors.
For all the above reasons, we conclude that what makes San Rafael Lagoon a unique environment is the combination of an Austral temperature regime with a warm-temperate latitude light regime. Dinophysis populations and their ciliate prey will have suitable light intensity and day length (photoperiod) for growth in late winter–early spring (time for the spring bloom in southwestern Europe) when there is 24 h darkness in polar latitudes. The eddy circulation reported by Soto et al. [107] may act as a retention area and explain the maintenance of moderate overwintering cell densities of Dinophysis. The extreme conditions of the lagoon, at the extreme latitude of existing glaciers, will act as a filter of stress-tolerant organisms and promote low diversity communities. It may also explain the absence of D. norvegica, co-occuring with D. acuminata in any other cold-temperate, boreal/austral system mentioned in Table 1.
3.6. Future Research Directions
The study of Davenport in the early 1990s [56] is the only information available on the phytoplankton communities in the lagoon. Sampling in that study was carried out with horizontal hauls of plankton nets of 40 µm mesh size in a single survey in summer. But the author pointed to the very low diversity (a bloom of Coscinodiscus) and the homogeneous composition of the phytoplankton communities throughout a 24 h cycle.
There are many questions related to Dinophysis population dynamics and early warning systems (EWS) which can be elegantly tested in Laguna San Rafael, a natural observatory extremely sensitive to climate change effects on calving glaciers.
There are ongoing research projects on the lagoon circulation and connectivity with adjacent water bodies, as well as on climate change-driven changes in the calving glacier’s limits. The challenge for HAB experts is to elucidate the hypothetical role of San Rafael Lagoon as a Dinophysis retention area for overwintering populations, or as a Dinophysis incubator and pelagic seed bank ready to export seed populations to the neighbouring fjords and channels. Alternatively, the lagoon may be an isolated body of water with connections limited to a thin surface layer. This last possibility would justify the morphology of a Dinophysis strain distinct from other populations in fjords and inner seas in Southern Chile.
4. Materials and Methods
4.1. Study Area
The Chilean Patagonian coast (southeastern Pacific Ocean), bordered to the east by the Andes Mountain Range, is one of the most extensive (from 41.5° S to 56.7° S) and complex fjord and channel systems in the world. Northern Chilean Patagonia extends from Puerto Montt (41.5° S) in the Los Lagos Province to the Taitao Peninsula (46.5° S) in Aysén [108]. The Northern Patagonian Icefield (NPI) caps the Andes Mountain range throughout the Northern Patagonia limits, and San Rafael Glacier is in its southernmost sector (Figure 1). San Rafael Glacier, one of 39 calving glaciers in the NPI, discharges into San Rafael Lagoon [52,53,109]. San Rafael Lagoon (46°40′ S; 74°56′ W) is the only tidewater glacier lagoon remaining in the Northern Patagonian Icefield and the lowest latitude tidewater glacier in the world. This fan-shaped (15 km long, 10 km wide) body of water has a maximal depth of 250 m in the area close to the glacier and temperatures ranging from 5.5 to 6.5 °C [110]. The lagoon is connected to the Pacific Ocean through “Río Témpanos” (=“Ice Floe River”, not really a river but a channel about 15 m deep) and Golfo Elefantes (Elephants Gulf), an elongated fjord.
Soto et al. [107] developed a high-resolution hydrodynamic model which described an anticyclonic eddy in the central part of the lagoon. The sampling station in this work, in the middle point of this eddy, was chosen as representative of the main central area of San Rafael Lagoon which may act as a retention area for phytoplankton populations.
Dinophysis specimens described here are from opportunistic samples collected with oceanographic bottles and vertical net-tows in Laguna San Rafael during cruises aimed to study the hydrodynamics and connectivity of the lagoon with adjacent water bodies. (Figure 1).
4.2. Sampling Overview
Between November 2020 and August 2021, three oceanographic cruises (15 November 2020; 25 February and 5 August 2021) were carried out in a southern section (46°20′–46°40′ S) of NW Patagonia on board R.V. Dr. Jürgen Winter. On each cruise, opportunistic sampling was carried out in one station (Figure 1, white dot on San Rafael Lagoon) to collect bottle samples for phytoplankton observations.
During each visit to the station, vertical profiles of temperature, salinity and in vivo chlorophyll a fluorescence were obtained with an AML (AML Oceanographic, Victoria, BC, Canada) CTD profiler model Metrec-XL equipped with a Turner Designs CYCLOPS-7 (Turner Designs, San Jose, CA, USA) submersible fluorometer (excitation 460 nm, emission, 620–715 nm).
Samples for quantitative analyses of micro-phytoplankton were collected with Niskin bottles at six discrete depths from surface to 50 m depth (0, 5, 10, 15, 20 and 50 m). Subsamples of 100 mL were collected in opaque glass jars and immediately fixed with acidic Lugol’s solution [111]. Samples for semiquantitative analysis of toxins in the plankton (toxin content per litre) were collected with vertical net-tows (20 μm mesh Nitex, 20 cm Ø,) through the upper 20 m of the water column; the filtered volume per vertical tow was about 628 L [73]. Samples were reduced to a final volume of 50 mL (adding filtered seawater if needed) and filtered through Whatman GF/F fiberglass filters (25 mm Ø, 0.7 μm pore size) (Whatman, Maidstone, UK). The filters and filtered material were placed in a cryotube, mixed with 1 mL analysis grade methanol and stored in the laboratory at −20 °C until analysis.
4.3. Phytoplankton Analyses
For quantitative analyses of phytoplankton, 10 mL of the Lugol’s fixed bottle samples were left to sediment overnight and analyzed under an inverted Olympus CKX41 microscope (Olympus, Tokyo, Japan) using the method described in Utermöhl [57]. Large but less abundant taxa, such as Dinophysis, were counted at a magnification of ×100, so that the detection limit was 100 cells L^−1^. Confidence limits of the cell counts were calculated according to Andersen and Throndsen [112].
4.4. Dinophysis Morphometry and Imaging
Observations were conducted on acidic Lugol’s fixed Dinophysis cells at ×20 magnification using a Nikon Eclipse TS2 inverted microscope (Nikon Corporation, Tokyo, Japan), equipped with an industrial-grade digital microscopy camera (model X7CAM4K16MPA) with 4K full-HD resolution and autofocus capabilities, enabling high-quality image acquisition under bright-field illumination. Maximum cell length (L) and dorso-ventral depth (D) of the large hypothecal plates were measured following the criteria of Balech [88]. Measurements were performed on digital images acquired using ToupTek XCamView software (version V1.8_20240530) and subsequently processed and analyzed with the free software ImageJ (version 1.54i) [113]. A total of 44 normal-sized cells and 9 small forms of Dinophysis were measured.
4.5. Single Cell Isolation, PCR Amplification and Sequencing
Cells of Dinophysis from the acidic Lugol’s preserved samples collected in Laguna San Rafael were individually picked with a capillary pipette and washed three times in drops of distilled water on a glass slide. Samples were photographed and measured with a Nikon Eclipse TS2 equipped with a 4K digital camera and transferred to 200 µL tubes. Then, a cold shock with liquid nitrogen was applied and samples were kept at −80 °C before immediate PCR analyses.
After a series of unsuccessful assays on single cells, tubes containing between 4 and 33 specimens showed positive results. ITS (ITS1-5.8S-ITS2) and partial plastid 23S rRNA gene were amplified using the pairs of primers ITSF01/PERK-ITS-AS (5′-TCCCTGCCCTTTGTACACAC-3′/5′-GCTTACTTATATGCTTAAATTCAG-3′, [114]) and p23Sr_f1/23Sr_r1 (5′-GGACAGAAAGACCCTATGAA-3′/5′-TCAGCCTGTTATCCCTAGAG-3′, [115]). The amplification reaction mixtures (20 mL) were performed using Horse-Power™ Taq DNA Polymerase MasterMix (2x) (Canvax, Valladolid, Spain). DNA was amplified in an Eppendorf Mastercycler EP5345 (Eppendorf AG, New York, NY, USA). PCR conditions were as follows: initial denaturation for 5 min at 94 °C, followed by 35 cycles of 35s at 94C, 35 s at 54 °C (ITSF01/PERK-ITS-AS), 55 °C (p23Sr_f1/23Sr_r1), and 1 min at 72 °C, followed by a final extension for 7 min at 72 °C. An 8 µL aliquot of each PCR reaction was checked by agarose gel electrophoresis (1% TAE, 50 V) and GelRed™ nucleic acid gel staining (Biotium, Hayward, CA, USA). PCR products were purified with ExoSAP-IT™ (USB Corporation, Cleveland, OH, USA). Sequencing reactions were performed using the Big Dye Terminator v3.1 reaction cycle sequencing kit and migrated in a SeqStudio genetic analyser (both at Applied Biosystems, Foster City, CA, USA) at the CACTI sequencing facilities (Universidade de Vigo). The ITS and plastid 23S rDNA sequences obtained in this study were deposited in GenBank (ITS Acc. Numbers are Dinophysis_ac_8 PX494338, Dinophysis_ac_9 PX494339, Dinophysis_ac_R6 PX494340, Dinophysis_ac_R8 PX494341, Dinophysis_ac_S1 PX494342, Dinophysis_ac_S2 PX494343 and Dinophysis_ac_S3 PX494344 and plastid 23S Acc. Numbers are PX498283 and PX498284 for 2_23S_San_Rafael_Dinophysis and 2_23S_San_Rafael_Dinophysis).
4.6. Alignment and Phylogenetic Analyses
Plastid 23S and internal transcribed spacer (ITS) rDNA sequences were analyzed separately. Each group of sequences was inspected, aligned and trimmed using Geneious Prime^®^ 2024.0.7 (Biomatters Ltd., London, UK). Each alignment was performed using MAFFT v7 [116] with the E-INS-i algorithm, and pairwise identity (%) matrix was calculated with Geneious Prime^®^ 2024.0.7 (Biomatters Ltd., London, UK). Patristic distances (measured in substitutions per site) were calculated using the software PATRISTIC v1.0 [117] and represented the sum of branch lengths between pairs of taxa in the phylogenetic tree. This distance was used to quantify evolutionary divergence among sequences.
Phylogenetic analyses were conducted in IQ-TREE v3.0.1 [118]. The best-fitting nucleotide substitution model was determined automatically using ModelFinder [119] according to the Bayesian Information Criterion (BIC), resulting in selection of the TPM3v + G4 model in the case of the 23S alignment and TIM2 + F + G4 for the ITS. The phylogenetic trees were represented using the ML method. Node support was assessed using three complementary methods: Shimodaira–Hasegawa approximate likelihood ratio test (SH-aLRT; 1000 replicates) [120], approximate Bayesian (aBayes) support [121] and ultrafast bootstrap (UFBoot; 1000 replicates) [122]. The final consensus maximum-likelihood tree was visualized in FigTree v1.4.4 [123], displaying all three support values at each node (SH-aLRT/aBayes/UFBoot). Following standard practice, nodes with SH-aLRT ≥ 80%, aBayes ≥ 0.95 and UFBoot ≥ 95%, were considered to have strong statistical support, whereas values between SH-aLRT 70–79%, aBayes 0.90–0.94 or UFBoot 90–94% indicated moderate support. Nodes below these thresholds were regarded as weakly supported.
4.7. Toxin Analysis
4.7.1. Sample Extraction
The filters and filtered net-towed material kept with methanol in cryotubes were thawed and concentrated by centrifugation (10,000 g; 20 min) to obtain a final volume of 1 mL. Each sample was then mixed with 1 mL of methanol (100%) and sonicated with a Branson Sonic Power 250 disruptor cellular sonifier (Thermo Fisher Scientific, San Jose, CA, USA) to extract lipophilic toxins. The extract obtained was clarified by centrifugation (10,000 g; 20 min), and the supernatant was then filtered through a 0.20 μm Clarinert nylon syringe filter (13 mm Ø) (MZ-Analysentechnik GmbH, Mainz, Germany). To analyze free OA, DTX1, PTX2, PTX2sa, YTX, and other lipophilic toxins, an aliquot of the sample (0.5 mL) was placed in an amber vial and kept at −20 °C until analysis. To analyze total OA, samples were subjected to alkaline hydrolysis following the standard procedure of the EU Reference Laboratory for Marine Biotoxins [124].
4.7.2. Toxin Detection and Quantification
The presence of lipophilic toxins in the extracts was checked by UHPLC-MS/MS using a Dionex Ultimate 3000 chromatographic system (UHPLC) (Thermo Fisher Scientific, San Jose, CA, USA) coupled to a Thermo Q Exactive Focus by means of a HESI-II electrospray interface, following the method described by Regueiro et al. [125], but modified in order to use a shorter column and to allow enough time for the elution of the toxins. Chromatographic separation was performed using a column Gemini NX-C18 (50 mm × 2 mm; 3 μm) from Phenomenex (Torrance, CA, USA). The flow rate was set to 0.35 mL min^−1^, and the injection volume was 10 μL. The mobile phase was used in gradient mode as follows: 85% of eluent A (100% water containing 6.7 mM NH_4_OH) and 15% of eluent B (90% acetonitrile:10% water with 6.7 mM NH_4_OH) was held for 1 min, followed by a linear increase to 80% B for 2.85 min and then an increase to 85% B for 0.15 min, and 90% B for 0.75 min and 100% B for 3.25 min. Finally, the gradient was returned to the initial conditions over 2 min and the column was re-equilibrated for 1 min. The mass spectrometer was operated using positive and negative acquisition experiments with the following settings: spray voltage positive 3500 V, negative 3000 V, sheath gas pressure 30, auxiliary gas flow 4, capillary temperature 350 °C, automatic gain control (AGC) of the C-Trap 5 × 10^4^, and mass resolution of 70,000 FWHM.
For quantification and confirmation, data acquisition was performed using selected-ion-monitoring (SIM) with data-dependent MS/MS (ddMS^2^). All analyses were performed using a mass inclusion list, including the precursor ion masses, expected retention time window, and collision energy (CE) for each toxin (Table 2). The toxin concentrations of OA, DTX-1, PTX-2, PTX-2sa, and YTX in the extracts were quantified by comparing the areas or peaks obtained in the chromatograms with those of certified reference materials obtained from NCR, Canada. The detection limits of the analyses (LOD) were 0.32 ± 0.03 ng mL^−1^ for OA; 0.24 ± 0.02 ng mL^−1^ for DTX1; and 2.61 ± 0.25 ng mL^−1^ for PTX2. Limits of quantification (LOQ) were 0.50 ± 0.03 ng mL^−1^ for OA; 0.50 ± 0.03 for DTX1; and 4.33 ± 0.25 for PTX2.
The total amount of toxins per sample extract corresponded to vertical net-tows and was expressed as ng of toxin per net-tow (ng NT^−1^) as in Fabro et al. [73]. The net-tow estimate was divided by the volume of filtered seawater during the vertical tow, and the toxins were expressed as pg toxin L^−1^. Considering the fact that the seawater passing through the net may suffer deflections, and also that the mesh can get clogged and thus may not be able to be filter efficiently, data obtained here must be considered as semiquantitative.
5. Conclusions
Opportunistic samplings revealed the occurrence of Dinophysis cf acuminata in a tidewater glacier lagoon formed by the climate change-driven retreat of San Rafael Glacier, in San Rafael Lagoon National Park and Reserve of the Biosphere.
The broad and coarse morphotypes of Dinophysis found in San Rafael Lagoon, with a toxin profile of pectenotoxins (PTX2) only, are similar to the description of Paulsen’s [87] broad and semi-circular (L:D = 1.18) forms first reported in Boreal waters (northwestern Iceland) and considered by Balech [89] to be synonyms of D. acuminata var. acuminata.
Partial sequences of ITS rDNA aligned D. acuminata from San Rafael Lagoon with another member of the D. acuminata complex from North America, Europe and Japan. Partial sequences of their plastid derived 23S rDNA, the first plastid sequence obtained from field specimens of D. acuminata in Chile, confirmed ciliates of the Mesodinium rubrum + major complex as their prey and plastid source.
The detection of moderate (>500 cells L^−1^) cell densities of Dinophysis cf acuminata in San Rafael Lagoon as well as the regular shape of the cells in the three seasonal (spring, summer and winter) surveys appear associated with extreme but stable conditions of a lagoon with an Austral temperature regime and a warm-temperate latitude light regime. These features added to the occurrence of one single species of Dinophysis with a simple toxin profile makes the lagoon a natural observatory for ecophysiology, ecotoxicology and population dynamic studies with a focus on the Dinophysis acuminata complex in a highly sensitive environment to climate change.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Van Egmond H.P. Aune T. Lassus P. Speijers G.J.A. Waldock M. Paralytic and diarrhoeic shellfish poisons: Occurrence in Europe, toxicity, analysis and regulation J. Nat. Toxins 199324182
- 2Granéli E. Codd G.A. Dale B. Lipiatou E. Maestrini S.Y. Rosenthal H. EUROHAB Science Initiative: Harmful Algal Blooms in European Marine and Brackish Waters Office for Official Publications of the European Communities Luxembourg 199997
- 3Fux E. Bire R. Hess P. Comparative accumulation and composition of lipophilic marine biotoxins in passive samplers and in mussels (M. edulis) on the West Coast of Ireland Harmful Algae 2009852353710.1016/j.hal.2008.10.007 · doi ↗
- 4Pizarro G. Paz B. Gonzáles-Gil S. Franco J.M. Reguera B. Seasonal variability of lipophilic toxins during a Dinophysis acuta bloom in Western Iberia: Differences between picked cells and plankton concentrates Harmful Algae 2009892693710.1016/j.hal.2009.05.004 · doi ↗
- 5Nielsen L.T. Krock B. Hansen P.J. Production and excretion of okadaic acid, pectenotoxin-2 and a novel dinophysistoxin from the DSP-causing marine dinoflagellate Dinophysis acuta—Effects of light, food availability and growth phase Harmful Algae 201323344510.1016/j.hal.2012.12.004 · doi ↗
- 6Nagai S. Suzuki T. Nishikawa T. Kamiyama T. Differences in the production and excretionkinetics of okadaic acid, dinophysistoxin-1, and pectenotoxin-2 between cultures of Dinophysis acuminata and Dinophysis fortii isolated from western Japan J. Phycol.2011471326133710.1111/j.1529-8817.2011.01076.x 27020356 · doi ↗ · pubmed ↗
- 7Tillmann U. John U. Toxic effects of Alexandrium spp. on heterotrophic dinoflagellates: An allelochemical defence mechanism independent of PSP-toxin content Mar. Ecol. Prog. Ser.2002230475810.3354/meps 230047 · doi ↗
- 8Long M. Krock B. Castrec J. Tillmann U. Unknown extracellular and bioactive metabolites of the genus Alexandrium: A review of overlooked toxins Toxins 20211390510.3390/toxins 1312090534941742 PMC 8703713 · doi ↗ · pubmed ↗