Histone Deacetylase 19 Controls Powdery Mildew Susceptibility by Attenuating Biosynthesis of Cuticular Wax and Salicylic Acid
Mengdi Zhang, Wenrui Zhao, Pengfei Zhi, Haoyu Li, Cheng Chang

TL;DR
A wheat enzyme called TaHDA19 makes the plant more vulnerable to powdery mildew by reducing protective waxes and a key chemical signal.
Contribution
TaHDA19 is newly identified as a regulator of wheat susceptibility to powdery mildew through epigenetic control of wax and salicylic acid.
Findings
Knockdown of TaHDA19 increases cuticular wax and salicylic acid, reducing powdery mildew susceptibility.
TaHDA19 suppresses TaECR and TaSARD1 transcription via histone deacetylation.
Cuticular wax and salicylic acid act as chemical cues affecting wheat's resistance to Blumeria graminis.
Abstract
Phytopathogenic Ascomycetes Blumeria graminis f. sp. tritici (Bgt) causes wheat powdery mildew disease and impacts global wheat production. Decoding the molecular wheat-Bgt interaction could facilitate the wheat disease resistance breeding. In this study, we elucidated that wheat histone deacetylase 19 (TaHDA19) regulates susceptibility to Bgt pathogen by suppressing biosynthesis of cuticular wax and salicylic acid (SA). Knockdown of wheat TaHDA19 gene expression led to in enhanced cuticular wax and SA accumulation, potentiated Bgt conidia germination and appressoria formation, attenuated formation of Bgt haustoria and microcolonies. Histone deacetylase TaHDA19 is enriched at the TaECR and TaSARD1 promoter regions to facilitate histone deacetylation, and thus suppressing TaECR and TaSARD1 transcription. In addition, we identified cuticular wax and SA regulated by TaHDA19 as chemical…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
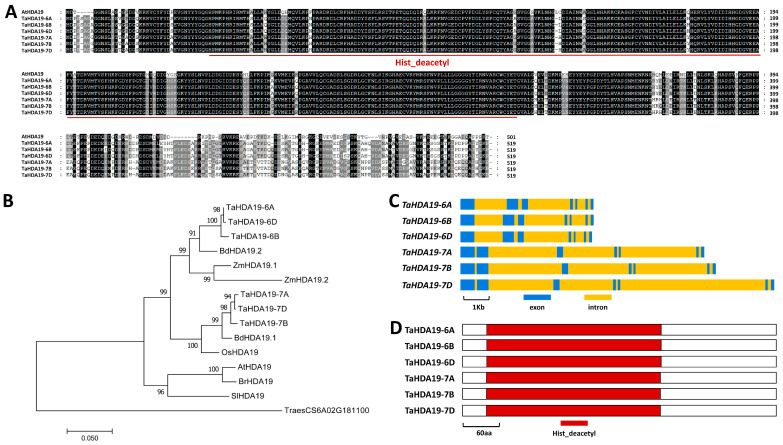 Figure 1
Figure 1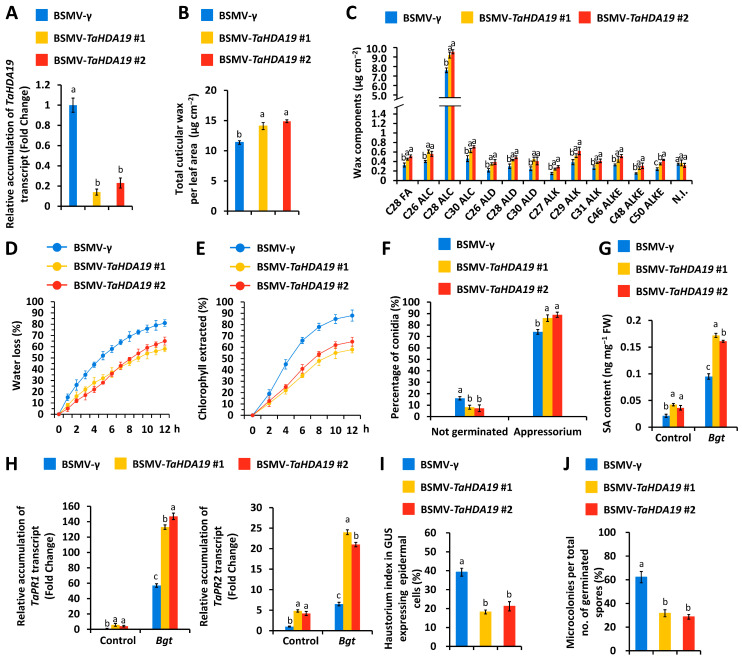 Figure 2
Figure 2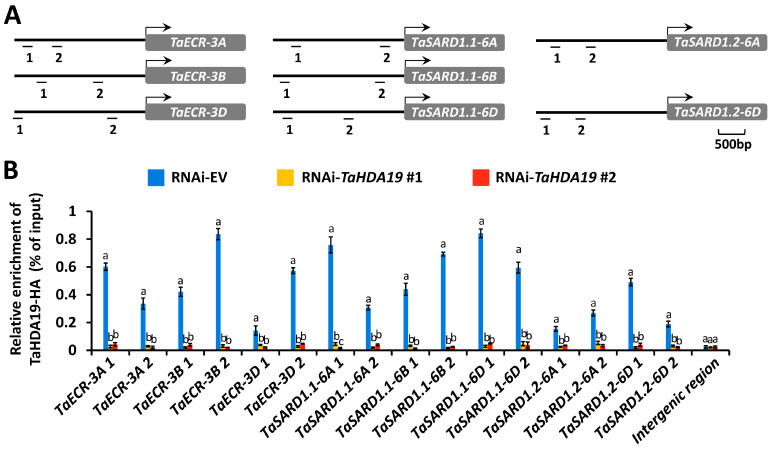 Figure 3
Figure 3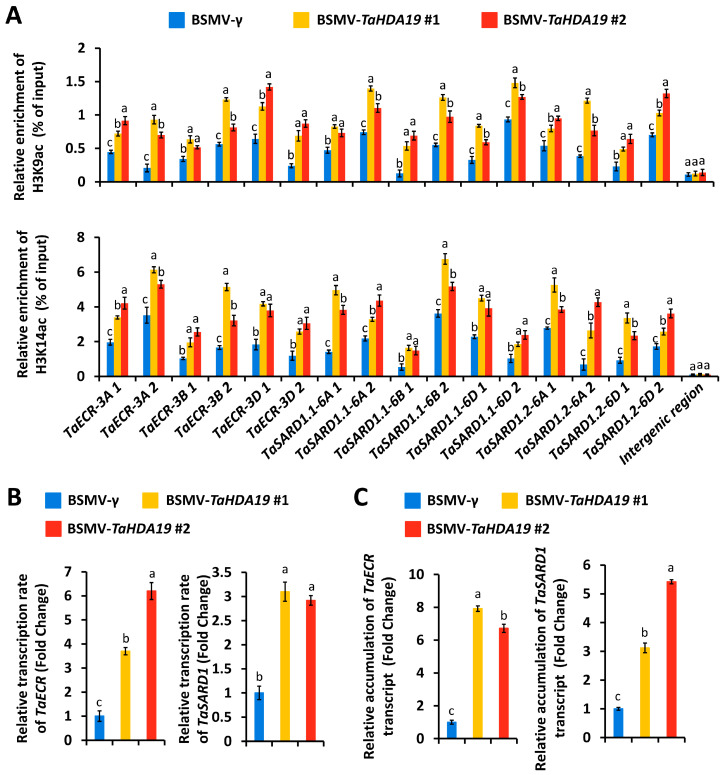 Figure 4
Figure 4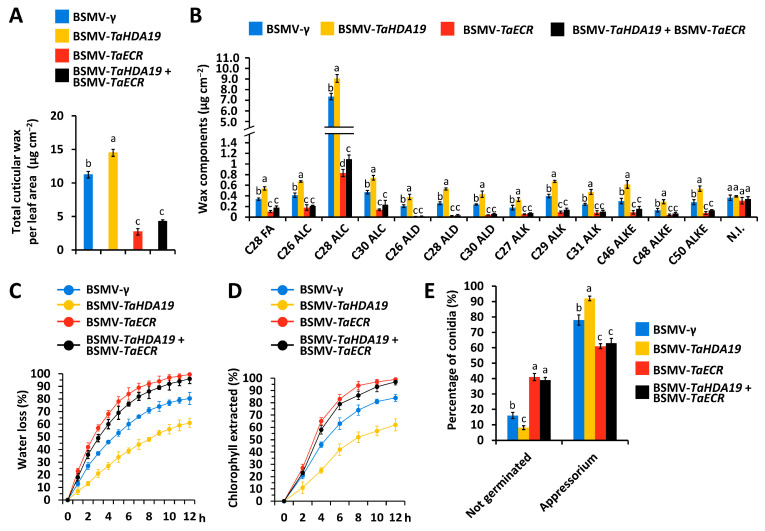 Figure 5
Figure 5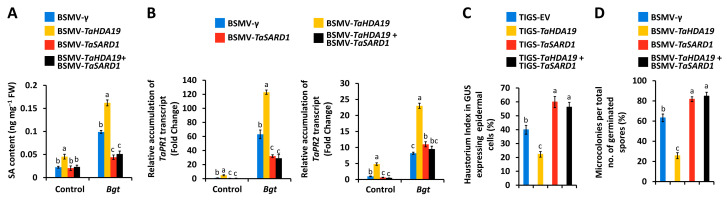 Figure 6
Figure 6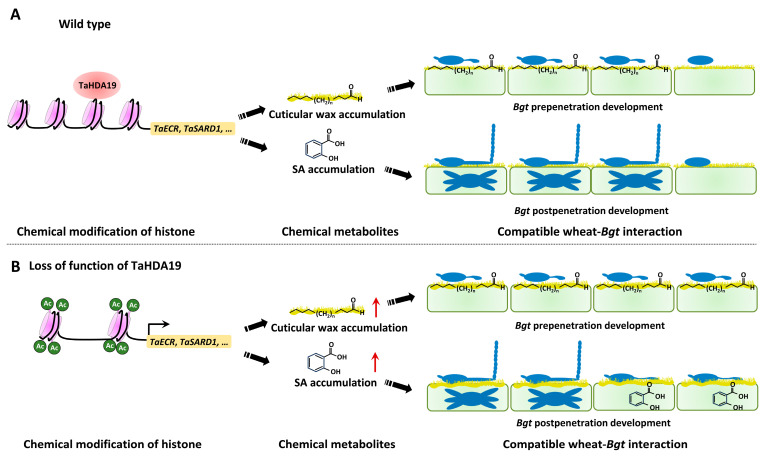 Figure 7
Figure 7- —Shandong Provincial Natural Science Foundation
- —Qingdao Science and Technology Bureau Fund
- —Qingdao University Fund
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Surface Properties and Treatments · Plant-Microbe Interactions and Immunity · Polysaccharides and Plant Cell Walls
1. Introduction
Phytopathogenic Ascomycetes Blumeria graminis f. sp. tritici (Bgt) in the family Erysiphaceae infects bread wheat (Triticum aestivum L.), one of the most widely cultivated staple crop plants, to cause the devastating wheat powdery mildew disease [1]. The air-borne conidia of Bgt could infect wheat leaf blade, leaf sheath, stem, and spike to interfere with plant physiological processes and affect plant developmental events [2]. It was estimated that Bgt infection could cause above 5% loss of global wheat production [3]. The grain end-use quality of bread wheat is also dampened by the powdery mildew disease [1,2]. Bgt conidia germinates on the epidermal surface of wheat aerial organs like leaf blades and stems, undergoes the development of feeding structure, haustorium, in wheat epidermal cells, and culminates into the microcolony formation [1,2]. Decoding the molecular wheat-Bgt interaction is essential for the wheat breeding against powdery mildew disease.
As an important histone posttranslational modification (PTM), acetylation occurring at the histone N-terminal tail could neutralize the histone positive charge, weaken the histone-DNA association and facilitate gene transcription. Through removing the histone acetylation mark, histone deacetylases (HDAC) could suppress gene transcription [4,5,6]. Some histone deacetylases have been characterized in Arabidopsis thaliana to regulate plant development and response to environ-mental challenges. For instance, Arabidopsis histone deacetylase AtHDA9 represses the flowering activator gene AGAMOUS-LIKE 19 (AtAGL19) in response to vernalization to regulate flowering time [7]. In addition, Arabidopsis histone deacetylase AtHDA9 function in concert with the SANT-domain containing protein POWERDRESS (PWR) and the transcription factor ABA-INSENSITIVE-4 (ABI4) to regulate drought stress response [8]. However, the function of histone deacetylases in cereal crops like the important staple crop bread wheat remains to be disclosed.
In this study, we explored the regulation of wheat histone deacetylase TaHDA19 on wheat powdery mildew susceptibility, and revealed that TaHDA19 mediates histone deacetylation at TaECR and TaSARD1 promoters to suppresses cuticular wax and salicylic acid (SA) biosynthesis and thus finetune wheat susceptibility to Bgt pathogen. This study reported for the first time the important role of histone deacetylase TaHDA19 in the regulation of wheat powdery mildew susceptibility and provide important information for the wheat disease resistance breeding.
2. Materials and Methods
2.1. Wheat and Bgt Pathogen Maintenance
Seedlings of wheat cultivar Yannong 999 were cultivated in climate chambers under 16 h, 20 °C light/8 h, 18 °C dark cycle. Bgt isolate E09 was maintained on the seedlings of Yannong 999.
2.2. Gene Transcription Analysis
RT-qPCR and nuclear run-on assays were performed to analyze the transcript accumulation and transcription rate of genes as described [9,10,11]. TaHDA19 gene is analyzed using primers 5′TTCAAGAGCGACCTCCTGA3′/5′AGATTCTCTCTTAACTCT3′.
2.3. Gene Silencing Assay
BSMV-VIGS and TIGS assays were performed to silence genes as described [9,10,11,12]. For the BSMV-VIGS, two independent silencing fragments targeting two conserved regions of TaHDA19 gene were amplified using primers 5′AAGGAAGTTTAACGGAGCGGAGGAAGGAGA3′/5′AACCACCACCACCGTGTCTGCTACTTCTACGAC3′ and 5′AAGGAAGTTTAAAATAATCCCCAAACTTGTG3′/5′AACCACCACCACCGTGTCTTCGACGGCCTCTAC3′. For the TIGS, fragments of TaHDA19 gene were amplified using primers 5′GGGGACAAGTTTGTACAAAAAAGCAGGCTTCCGGAGCGGAGGAAGGAGA3′/5′GGGGACCACTTTGTACAAGAAAGCTGGGTCGTCTGCTACTTCTACGAC3′.
2.4. Wheat-Bgt Interaction Analysis
Bgt conidial germination, appressorial formation, haustorial development and microcolony formation were statistically analyzed to judge the wheat-Bgt interaction as described [9,10,11]. Bgt conidial germination and appressorial formation rates were described as a percentage of Bgt conidia with germ tube development or appressorial formation. Bgt microcolony index were described as a percentage of germinated Bgt with microcolony formation. Bgt haustorium index was expressed as a percentage of Bgt-infected wheat cells with haustoria formation.
2.5. Cuticular Wax and Free SA Measurements
Measurement of cuticular wax and free SA in wheat leaves by gas chromatography-mass spectrometry (GC-MS) and high-performance liquid chromatography (HPLC) were conducted as described previously [9,10,11,13].
2.6. Wheat Leaf Cuticle Permeability Analysis
Water loss and chlorophyll leaching assays were conducted to judge the wheat leaf cuticle permeability as described [14,15].
2.7. Chromatin Immunoprecipitation (ChIP) Assay
For analyzing the TaHDA19 distribution at promoter regions of TaECR and TaSARD1 genes, the ChIP assay was conducted using the wheat protoplast cells. We cotransfected the wheat protoplast with TaHDA19-HA and RNA interference (RNAi) constructs, and performed a ChIP assay to characterize the distribution of TaHDA19-HA at TaECR and TaSARD1 promoters, and the coding sequence of TaHDA19-6A gene was employed as a representative TaHDA19 gene for expressing the TaHDA19-HA fusion protein. For analyzing the histone acetylation H3K9ac and H3K14ac at TaECR and TaSARD1 promoters, the ChIP assay was conducted using leaves of BSMV-VIGS wheat plants. We silenced all endogenous TaHDA19 genes using BSMV-VIGS, and performed a ChIP assay to analyze H3K9ac and H3K14ac at TaECR and TaSARD1 promoters. The ChIP assay was conducted as described [9,10,11,13].
2.8. Statistical Analysis
Three technical replicates for each assay were statistically analyzed by one-way ANOVA with Duncan’s post hoc test. bars with different letters are significantly different (p < 0.05), and letters were assigned in alphabetical order. Similar results were obtained from three independent biological replicates for these assays.
3. Results
3.1. Identification of Wheat TaHDA19 Genes
Through searching the wheat genome database using protein sequence of Arabidopsis histone deacetylase AtHDA19, we identified TaHDA19 genes in wheat chromosomes 6A (TraesCS6A02G184100), 6B (TraesCS6B02G212600), 6D (TraesCS6D02G171000), 7A (TraesCS7A02G365600), 7B (TraesCS7B02G261800), and 7D (TraesCS7D02G356800) (Figure 1A). Phylogenetic tree reconstruction validated that these allelic TaHDA19 proteins are wheat close homologs of HDA19 proteins from Arabidopsis thaliana (At), Brassica rapa (Br), Solanum lycopersicum (Sl), Zea mays (Zm), Oryza sativa (Os), Brachypodium distachyon (Bd), and Triticum aestivum (Ta). TaHDA6 (TraesCS6A02G181100) protein sequence was employed as an external branch in the phylogenetic analysis (Figure 1B). As shown in Figure 1C, 7 exons and 6 introns exist in the genome sequence of allelic TaHDA19 genes. Histone deacetylase (Hist_deacetyl) domain was identified from all TaHDA19 proteins (Figure 1D).
3.2. Regulation of TaHDA19 Gene on the Wheat Cuticular Wax and SA Biosynthesis and Powdery Mildew Susceptibility
To analyze the function of TaHDA19 genes in the compatible wheat-Bgt interaction, we conducted the BSMV-VIGS assay to silence all endogenous TaHDA19 genes in the Bgt-susceptible wheat cultivar Yannong 999. As shown in the Figure 2A, accumulation levels of TaHDA19 gene transcript were significantly reduced by BSMV-VIGS targeting two conserved regions of TaHDA19 genes (BSMV-TaHDA19 #1 and BSMV-TaHDA19 #2). GC-MS results indicated that total cuticular wax loads increased from 11.29 μg cm^−2^ in the BSMV-γ control leaves to above 14.08 μg cm^−2^ in the TaHDA19 gene-silenced wheat leaves (Figure 2B). As shown in the Figure 2C, accumulation of major wax constituents like VLC fatty acids (FA), alcohols (ALC), aldehydes (ALD), alkanes (ALK), alkyl esters (ALKE) were increased to different extent in the wheat leaves with silenced TaHDA19 gene. Measurement of wheat leaf water loss rate and chlorophyll leaching levels revealed that cuticle permeability of wheat leaves decreased in the BSMV-TaHDA19 #1 and BSMV-TaHDA19 #2 plants, compared with BSMV-γ control plants (Figure 2D,E). We examined the Bgt prepenetration events, conidia germination and appressoria formation, on the TaHDA19 gene-silenced wheat leaves. As shown in Figure 2F, approximately 8% more Bgt conidia germination and 12% more appressoria formation was observed on the wheat leaves with silenced TaHDA19 gene. These results indicated that TaHDA19 gene negatively regulate wheat cuticular wax biosynthesis and Bgt prepenetration events, conidia germination and appressoria formation.
We measured the levels of plant hormone salicylic acid (SA) in the TaHDA19 gene-silenced wheat leaves. HPLC results showed that accumulation of free SA significantly increased in the wheat leaves with silenced TaHDA19 gene (Figure 2G). As shown in the Figure 2H, accumulations of SA marker genes TaPR1 and TaPR2 transcripts were remarkable increased in the wheat leaves with silenced TaHDA19 gene. We examined the Bgt postpenetration events, haustoria development and microcolony formation, on the TaHDA19 gene-silenced wheat leaves. As shown in Figure 2I,J, approximately 17% less Bgt haustoria development and 23% less microcolony formation was observed on the wheat epidemral cells or leaves with silenced TaHDA19 gene. These results indicated that TaHDA19 gene negatively regulates wheat SA biosynthesis and facilitate Bgt postpenetration events, haustoria development and microcolony formation.
3.3. Distribution of Wheat Histone Deacetylase TaHDA19 Protein at Promoter Regions of TaECR and TaSARD1 Genes
Previous studies revealed that wheat wax biosynthesis gene TaECR and SA biosynthesis activator gene TaSARD1 are regulated by epigenetic events like histone acetylation [9,10,11,13]. We conducted the ChIP-qPCR assay to analyze the putative distribution of histone deacetylase TaHDA19 at promoter regions of TaECR and TaSARD1 genes. We firstly expressed the fusion protein TaHDA19-HA in the wheat protoplast cell. Considering that wheat TaHDA19 isoforms shared high sequence identity, we employed allele TaHDA19-6A as a representative TaHDA19 gene in the expression of the fusion protein TaHDA19-HA. TaSARD1.2 gene was not identified from the wheat chromatin 6B in previous study [16], and TaSARD1.2-6B gene promoter was not included in this study. As shown in Figure 3, TaECR and TaSARD1 promoter fragments were immuno-precipitated with the antibody against the fusion protein TaHDA19-HA, indicating that that wheat TaHDA19 protein occupied at promoter regions of TaECR and TaSARD1 genes.
3.4. Epigenetic Regulation of Wheat Histone Deacetylase TaHDA19 on the Transcription of TaECR and TaSARD1 Genes
We conducted the ChIP-qPCR assay to analyze the putative regulation of histone deacetylase TaHDA19 on the histone acetylation at TaECR and TaSARD1 promoters. The accumulation of histone acetylation marks H3K9ac and H3K14ac at TaECR and TaSARD1 promoters were significantly increased in the TaHDA19 gene-silenced wheat leaves (Figure 4A). These ChIP-qPCR results implicate that histone deacetylase TaHDA19 protein is enriched at TaECR and TaSARD1 promoters to mediate histone deacetylation. We then performed the nuclear run-on assay to examine TaHDA19 regulation on the TaECR and TaSARD1 gene transcription. As shown in Figure 4B, transcription rates of TaECR and TaSARD1 genes were remarkably enhanced in the TaHDA19 gene-silenced wheat leaves. As revealed by the RT-qPCR analysis, accumulation levels of TaECR and TaSARD1 transcript were significantly increased in the TaHDA19 gene-silencing wheat leaves (Figure 4C). These data collectively suggested that histone deacetylase TaHDA19 directly suppresses of TaECR and TaSARD1 gene transcription by promoting histone deacetylation at TaECR and TaSARD1 promoters.
3.5. Functional Characterization of TaHDA19 and TaECR Genes in the Regulation of Wheat Cuticular Wax Biosynthesis and Bgt Prepenetration Susceptibility
We simultaneously silenced TaHDA19 and TaECR genes using BSMV-VIGS and analyzed the cuticular wax accumulation by GC-MS. As shown in Figure 5A, total cuticular wax loads significantly decreased from 11.23 μg cm^−2^ in the BSMV-γ control leaves and 14.41 μg cm^−2^ in the TaHDA19 gene-silenced wheat leaves to 4.33 μg cm^−2^ in the wheat leaves with silenced TaHDA19 and TaECR genes. Major wax constituents were remarkable decreased in the TaHDA19/TaECR genes co-silenced wheat leaves (Figure 5B). As shown in Figure 5C,D, cuticle permeability of wheat leaves obviously increased in the wheat plants with co-silenced TaHDA19/TaECR genes, compared with BSMV-γ control plants. We examined the Bgt prepenetration events, conidia germination and appressoria formation, on the TaHDA19/TaECR genes co-silenced wheat leaves. As shown in Figure 5E, significantly increased ration of Bgt conidia germination and appressoria formation was observed on the wheat leaves with co-silenced TaHDA19/TaECR genes, compared with control plants.
3.6. Functional Characterization of TaHDA19 and TaSARD1 Genes in the Regulation of Wheat SA Biosynthesis and Bgt Postpenetration Susceptibility
We simultaneously silenced TaHDA19 and TaSARD1 genes using BSMV-VIGS and analyzed the SA accumulation by HPLC. As shown in Figure 5A, accumulation of free SA significantly decreased in the wheat leaves with co-silenced TaHDA19 and TaSARD1 genes, compared with control plants or TaHDA19 gene-silenced plants (Figure 6A). Similarly, transcript levels of SA marker genes TaPR1 and TaPR2 were significantly decreased in the wheat leaves with silenced TaHDA19 and TaSARD1 genes, compared with BSMV-γ control leaves and TaHDA19 gene-silenced. We examined the Bgt postpenetration events, haustoria development and microcolony formation, on the TaHDA19/TaSARD1 genes co-silenced wheat epidermal cells or leaves. As shown in Figure 6C,D, significantly increased percentage of Bgt haustoria development and microcolony formation were observed on the wheat epidermal cells or leaves with silenced TaHDA19 and TaSARD1 genes, compared with controls and TaHDA19 gene-silenced wheat epidermal cells or leaves.
4. Discussion
4.1. Wheat TaHDA19 Gene Suppresses Wax Biosynthesis to Dampen Bgt Prepenetraion Development
In this study, we identified allelic TaHDA19 genes from wheat chromosomes 6A, 6B, 6D, 7A, 7B, and 7D as AtHDA19 gene homologs in the agriculturally important crop bread wheat (Triticum aestivum L., AABBDD). Silencing of wheat TaHDA19 gene resulted in enhanced cuticular wax accumulation, potentiated Bgt conidia germination and appressoria formation. Importantly, we demonstrated that histone deacetylase TaHDA19 protein is enriched at TaECR promoters to promote histone deacetylation and suppress TaECR gene transcription. Importantly, silencing of cuticular wax biosynthesis gene TaECR could dampen the cuticular wax accumulation and attenuate conidia germination and appressoria formation of Bgt pathogen on the TaHDA19 gene-silenced wheat plants. These results implied that wheat histone deacetylase TaHDA19 protein epigenetically suppresses wax biosynthesis by promoting histone deacetylation at TaECR promoters, thereby negatively regulating biosynthesis of wheat cuticular wax essential for stimulating Bgt prepenetraion development events, conidia germination and appressoria formation.
Previous studies showed that the wheat Spt-Ada-Gcn5-acetyltransferase (SAGA) complex mediate the histone acetylation at TaECR promoters to activate the TaECR transcription and stimulate cuticular wax biosynthesis [13]. Herein, histone deacetylase TaHDA19 was demonstrated to promote histone deacetylation at TaECR gene promoters and attenuate cuticular wax biosynthesis. Therefore, it is intriguing the examine to the potential antagonistic action of SAGA complex and TaHDA19 in the regulation of TaECR gene transcription and cuticular wax biosynthesis. In the dicot model plant A. thaliana, AtHDA19 protein usually interacts with the transcriptional corepressor TOPLESS (TPL) to form the HDA19-TPL repressor complex in the regulation of plant flower development and hormone response [17,18]. Identifying the wheat TPL proteins might provide more insight into the mechanism underlying regulation cuticular wax biosynthesis. In addition, previous studies revealed that wax components VLC aldehydes could chemically stimulate conidia germination and appressoria formation of B. graminis [19,20]. In this study, VLC aldehydes overaccumulation and enhanced Bgt conidial germination were observed in the wheat leaves with silenced TaHDA19 gene. Therefore, it is proposed that TaHDA19 negatively regulates Bgt conidia germination and appressoria formation probably via suppressing the production of wax cues VLC aldehydes.
4.2. Wheat TaHDA19 Gene Represses SA Biosynthesis to Facilitate Bgt Postpenetraion Development
Herein, we demonstrated that silencing of wheat TaHDA19 gene resulted in enhanced SA accumulation, attenuated Bgt haustoria development and microcolony formation. Notably, we found that histone deacetylase TaHDA19 protein is enriched at TaSARD1 promoters to promote histone deacetylation and epigenetically suppress TaSARD1 gene transcription. Notably, silencing of SA biosynthesis activator gene TaSARD1 could dampen the SA biosynthesis and potentiate Bgt haustoria development and microcolony formation in the TaHDA19 gene-silenced wheat plants. These results suggested that wheat histone deacetylase TaHDA19 protein epigenetically suppresses SA biosynthesis by promoting histone deacetylation at the SA biosynthesis activator gene, thereby negatively regulating SA biosynthesis to facilitate Bgt postpenetraion development events, haustoria development and microcolony formation. Although we have measured the SA accumulation in the control (basal) and Bgt-induced (induced) wheat leaves in the TaHDA19 gene-silenced BSMV-VIGS wheat leaves, constructing TaHDA19 gene-knockout CRISPR mutant and analyzing SA accumulation in the time-course after Bgt-infection could provide more insight into the regulation of TaHDA19 gene on wheat SA biosynthesis.
In A. thaliana, AtHDA19 gene negatively regulate SA biosynthesis to maintain susceptibility to the hemibiotrophic pathogenic bacterium Pseudomonas syringae [21,22]. In this study, we demonstrated that TaHDA19 gene suppresses SA biosynthesis to facilitate Bgt postpenetration susceptibility. These studies suggested that the suppression of HDA19 gene on the SA biosynthesis might be conserved in the dicot Arabidopsis and monocot bread wheat. In addition, SWI/SNF chromatin remodeler TaSWI3B, TaSWI3D and DNA methyltransferase TaMET1 were identified as epigenetic suppressor of TaSARD1 gene transcription and SA biosynthesis [9,10,11]. As summarized by prior reviews, different epigenetic regulators usually function in concert to regulate gene transcription. Characterizing potential interplays of histone deacetylase TaHDA19 protein with SWI/SNF type chromatin remodeler and DNA methyltransferase could provide novel insight into the epigenetic mechanism regulating SA biosynthesis.
As depicted in Figure 7, we proposed a model of the wheat histone deacetylase TaHDA19 regulating the wheat powdery mildew susceptibility. In the wild-type plants (Figure 7A), wheat histone deacetylase TaHDA19 protein epigenetically suppresses biosynthesis of cuticular wax and SA by promoting histone deacetylation at promoters of TaECR and TaSARD1 genes. Cuticular wax provides chemical cues stimulating Bgt prepenetraion development events, conidia germination and appressoria formation, whereas hormone SA initiates plant defense response and restricts Bgt postpenetraion development events, haustoria development and microcolony formation. In the absence of wheat histone deacetylase TaHDA19 (Figure 7B), histone acetylation is potentiated at TaECR and TaSARD1 gene promoters, which is associated with the epigenetic activation of TaECR and TaSARD1 genes and enhanced cuticular wax and SA biosynthesis. As a result, Bgt pre- and postpenetration susceptibility was altered.
Many questions are unaddressed in this study. For instance, causality between TaHDA19-mediated histone deacetylation and target gene transcriptional repression remains to be disclosed. Identifying TaHDA19 protein interacting partners by yeast-two hybrid and characterizing genome-wide targets of TaHDA19 protein by ChIP-seq could shed novel light into the gene transcriptional regulation by TaHDA19. Furthermore, although TaHDA19/TaSARD1 and TaHDA19/TaECR genes co-silencing experiments support the pathway positioning of TaHDA19 gene with TaSARD1 and TaECR genes, pathway linearity and parallel regulation remains to revolved. Moreover, functional characterization of TaHDA19 gene in wheat powdery mildew susceptibility was facilitated by BSMV-VIGS technique. Although two independent silencing fragments targeting different regions of TaHDA19 gene were employed in the BSMV-VIGS assay, this technique has limitations like off-target silencing, transient nature, and effects across homoeologs. Generating stable transgenic TaHDA19 gene-overexpression wheat plants or TaHDA19 gene-knockout mutant might provide more insight into TaHDA19 function and regulation in future research.
Multiple Susceptibility (S) genes that facilitate the wheat susceptibility to host-adapted pathogens have been identified [23,24,25]. Editing of these S genes by genome editing techniques like CRISPR-Cas9 and transcription activator-like effector nucleases (TALENs) could reverse susceptibility and confer disease resistance [26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41]. Although TaHDA19 gene negatively regulates cuticular wax and SA biosynthesis to finetune pre-and post-penetration susceptibility to Bgt and could not be simply classified as a S gene, constructing the TaHDA19-knockout wheat plants by CRISPR-Cas9 and TILLING techniques might attenuate wheat post-penetration susceptibility to Bgt and could contribute to wheat breeding against powdery mildew in future research.
5. Conclusions
Herein, we characterized the function of wheat histone deacetylase gene TaHDA19 in the regulation of compatible wheat-Bgt interaction. Our studies revealed that histone deacetylase TaHDA19 protein epigenetically suppressed biosynthesis of wheat cuticular wax and SA by promoting histone deacetylation at the promoter regions of wax biosynthesis gene TaECR and SA biosynthesis activator gene TaSARD1, thereby differentially affecting Bgt pre- and postpenetration events. These findings elucidated the novel epigenetic regulation of wheat chemical cues essential for the compatible wheat-Bgt interaction and provide valuable information for developing Bgt-resistant wheat varieties in future research.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kusch S. Qian J. Loos A. Kümmel F. Spanu P.D. Panstruga R. Long-term and rapid evolution in powdery mildew fungi Mol. Ecol.202333 e 1690910.1111/mec.1690936862075 · doi ↗ · pubmed ↗
- 2Mapuranga J. Chang J. Yang W. Combating powdery mildew: Advances in molecular interactions between Blumeria graminis f. sp. tritici and wheat Front. Plant Sci.202213110290810.3389/fpls.2022.110290836589137 PMC 9800938 · doi ↗ · pubmed ↗
- 3Savary S. Willocquet L. Pethybridge S.J. Esker P. Mc Roberts N. Nelson A. The global burden of pathogens and pests on major food crops Nat. Ecol. Evol.2019343043910.1038/s 41559-018-0793-y 30718852 · doi ↗ · pubmed ↗
- 4Kumar V. Thakur J.K. Prasad M. Histone acetylation dynamics regulating plant development and stress responses Cell. Mol. Life Sci.2021784467448610.1007/s 00018-021-03794-x 33638653 PMC 11072255 · doi ↗ · pubmed ↗
- 5Chen X. Ding A.B. Zhong X. Functions and mechanisms of plant histone deacetylases Sci. China Life Sci.20206320621610.1007/s 11427-019-1587-x 31879846 · doi ↗ · pubmed ↗
- 6de Rooij P.G.H. Perrella G. Kaiserli E. van Zanten M. The diverse and unanticipated roles of histone deacetylase 9 in coordinating plant development and environmental acclimation J. Exp. Bot.2020716211622510.1093/jxb/eraa 33532687569 PMC 7586748 · doi ↗ · pubmed ↗
- 7Kim W. Latrasse D. Servet C. Zhou D.X. Arabidopsis histone deacetylase HDA 9 regulates flowering time through repression of AGL 19Biochem. Biophys. Res. Commun.201343239439810.1016/j.bbrc.2012.11.10223237803 · doi ↗ · pubmed ↗
- 8Khan I.U. Ali A. Khan H.A. Baek D. Park J. Lim C.J. Zareen S. Jan M. Lee S.Y. Pardo J.M. PWR/HDA 9/ABI 4 complex epigenetically regulates ABA dependent drought stress tolerance in Arabidopsis Front. Plant Sci.20201162310.3389/fpls.2020.0062332528497 PMC 7266079 · doi ↗ · pubmed ↗