Identification and Characterization of Pathogenic Fusarium Species Causing White Mold Disease in Cultivated Morels (Morchella spp.) in China
Luzhen Wang, Qi Zhao, Muqing Bai, Yongwei Wang, Keling Liu, Rujia Liang, Frederick Leo Sossah, Odeshnee Naicker, Chunlan Zhang

TL;DR
This study identifies twelve Fusarium species causing white mold disease in cultivated morels in China, with F. verticillioides being the most common.
Contribution
The first comprehensive identification of twelve Fusarium species responsible for white mold disease in morels.
Findings
Twelve Fusarium species were identified as causing white mold disease in morels, with F. verticillioides being the most prevalent.
All twelve identified Fusarium species were confirmed to be pathogenic to morel ascocarps through pathogenicity assays.
Most isolates were recovered from northern China, highlighting regional differences in pathogen distribution.
Abstract
White mold disease (WMD) is a major constraint to Morchella cultivation in China, leading to significant yield and quality losses. While Fusarium species are recognized plant pathogens, their diversity and role in WMD of morels have been poorly understood. This study aimed to identify and characterize Fusarium species associated with WMD in cultivated morels. Symptomatic ascocarps were collected from 22 cultivation bases across 16 provinces in China. A total of 120 Fusarium isolates were recovered and identified using morphological traits and multi-locus phylogenetic analysis. Twelve Fusarium species were identified, F. acuminatum, F. avenaceum, F. clavum, F. compactum, F. falciforme, F. flocciferum, F. ipomoeae, F. mucidum, F. oxysporum, F. proliferatum, F. subglutinans, and F. verticillioides, with most isolates recovered from northern China. Among them, F. verticillioides was the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
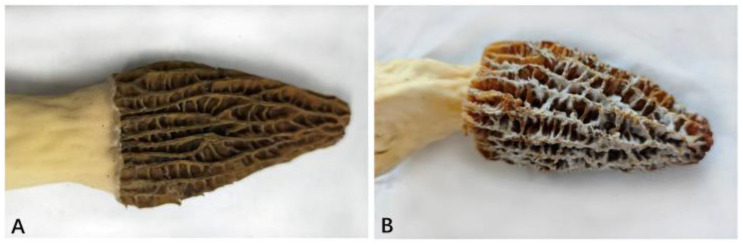 Figure 1
Figure 1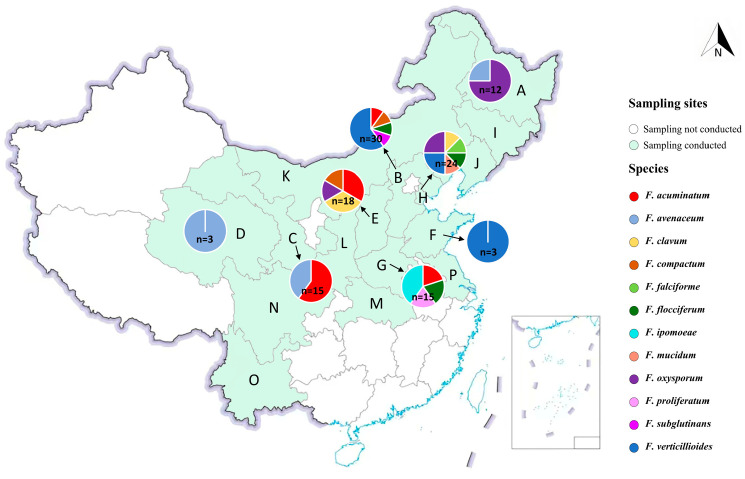 Figure 2
Figure 2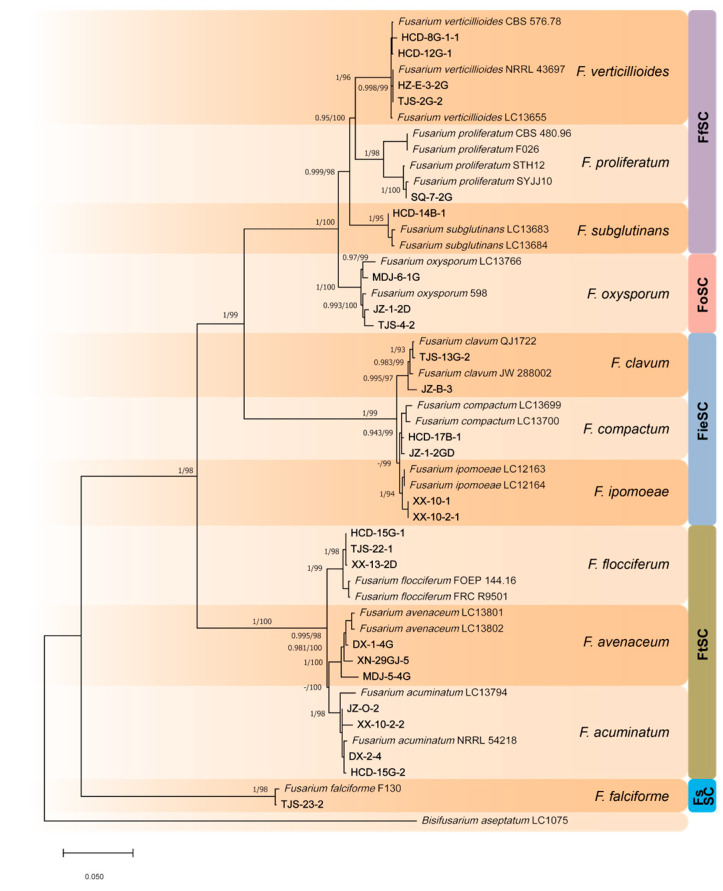 Figure 3
Figure 3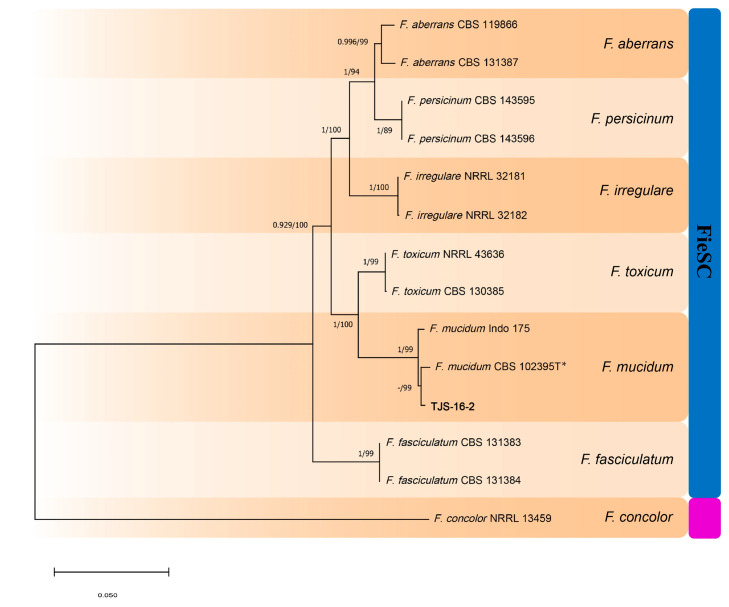 Figure 4
Figure 4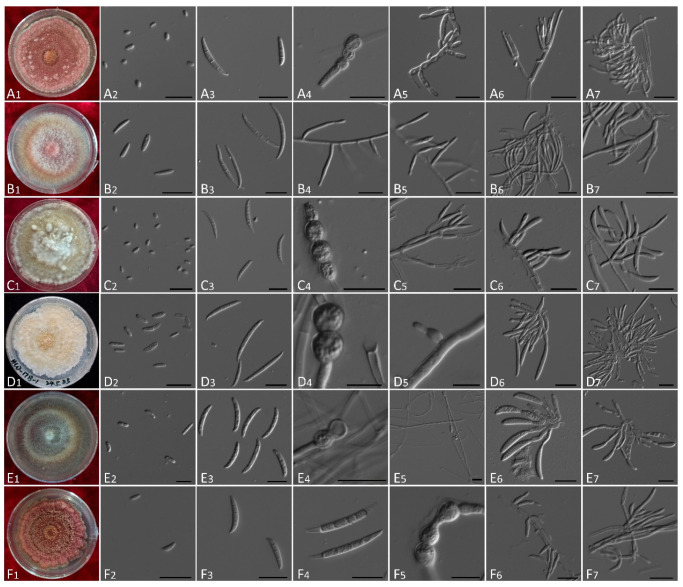 Figure 5
Figure 5 Figure 6
Figure 6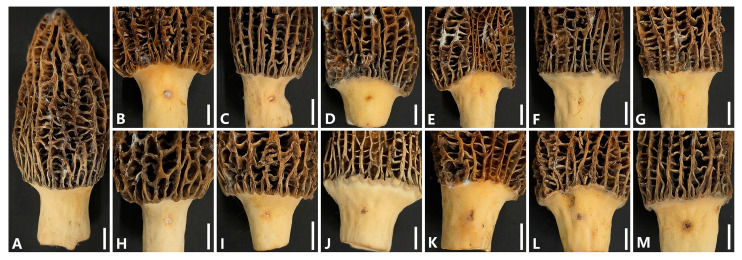 Figure 7
Figure 7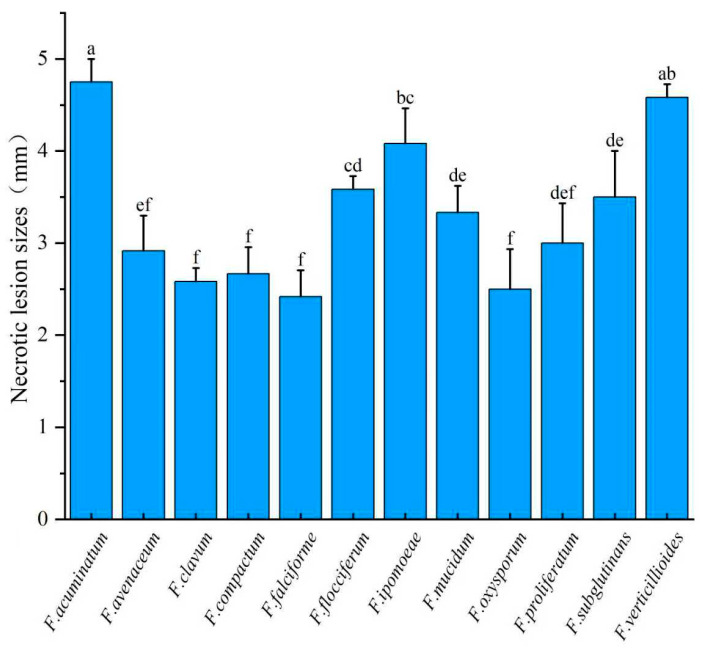 Figure 8
Figure 8- —Jilin Provincial Department of Science and Technology Plan projects
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycotoxins in Agriculture and Food · Mycorrhizal Fungi and Plant Interactions
1. Introduction
Morels (Morchella spp.), members of the order Pezizales, family Morchellaceae, are highly prized edible Ascomycetes renowned for their distinctive flavor, nutritional value, and medicinal properties [1]. These fungi thrive in cool, humid environments and are valued for their bioactive compounds, including amino acids, adenosine, vitamins, and polysaccharides [2,3]. Modern pharmacological studies have indicated antioxidant, anti-inflammatory, immunomodulatory, hepatoprotective, and hypoglycemic activities associated with morel consumption.
Given their high economic potential, the artificial cultivation of Morchella species has attracted growing global interest. A major breakthrough occurred in the 1980s when Ower (1982) successfully domesticated and produced morel fruiting bodies under controlled indoor conditions [4]. In China, field cultivation of morels began in 2012 [5] and has since expanded rapidly, reaching approximately 10,667 ha by 2021. The main cultivated species in China include M. importuna, M. sextelata, and M. septimelata, with M. sextelata accounting for over 85% of the national cultivation area. Morel production currently ranks 25th among the 29 commercially cultivated edible fungi in the country [6].
Despite its rapid expansion, large-scale morel cultivation faces significant challenges, particularly from fungal diseases that reduce both yield and quality. Each year, disease outbreaks affect approximately 25% of the total cultivation area. Several fungal pathogens have been implicated in morel diseases, including Neonectria candida, Paecilomyces penicillatus, and Pseudodiploöspora longispora, which are associated with white mold and pileus rot [7,8,9]. Other reported pathogens include Fusarium incarnatum (stipe rot), Fusarium nematophilum (stalk rot), and various species of Clonostachys, Lecanicillium, and Trichoderma [10,11]. Additionally, cobweb disease in morels has been attributed to Cladobotryum and Hypomyces species [12,13,14].
These diverse pathogens often produce dense, white mycelial growth on morel ascocarps, collectively described as white mold disease (WMD). Under favorable environmental conditions, particularly high humidity and temperature, WMD can spread rapidly, leading to severe production losses [8,15]. Currently, WMD is regarded as the most destructive disease affecting Morchella cultivation. Liu et al. (2023), using high-throughput ITS amplicon sequencing, identified P. longispora and Clonostachys solani as frequently occurring WMD pathogens [16]. However, metabarcoding has limitations, particularly in resolving closely related species. Thus, comprehensive pathogen identification using pure cultures and multi-gene phylogenetic analyses is needed.
To address these limitations and enhance understanding of the fungal pathogens associated with WMD in morels, in this study, we conducted a comprehensive investigation of Fusarium species affecting M. sextelata (Figure 1A). Specifically, the objectives of this study were as follows: (1) To isolate and identify Fusarium species associated with symptomatic morel ascocarps collected from diverse cultivation sites across China, using morphological and multi-locus phylogenetic analyses. (2) To evaluate the pathogenicity of representative isolates under controlled conditions. (3) To assess virulence variability among species to clarify their roles in WMD epidemiology. This study reveals the species diversity and pathogenic potential of Fusarium associated with diseases in morel cultivation. These results provide an important basis for the development of targeted prevention and control strategies for sustainable morel production.
2. Materials and Methods
2.1. Field Survey and Sampling
From 2023 to 2025, a field survey targeting WMD of morels was conducted across 22 cultivation sites in 16 major morel-producing provinces in China: Gansu, Heilongjiang, Hebei, Henan, Hubei, Inner Mongolia, Jiangsu, Jilin, Liaoning, Qinghai, Shaanxi, Shandong, Shanxi, Sichuan, Tianjin, and Yunnan. Sampling sites were selected based on the presence of symptomatic morel ascocarps exhibiting signs of WMD (Figure 1B). In total, 832 symptomatic ascocarps were collected. All samples were placed in sterile plastic bags, transported on ice, and stored at 4 °C prior to laboratory analysis.
2.2. Fungal Isolation and Culture
Fungal isolates were obtained using two complementary methods. For samples exhibiting sporulation, mature sporocarps (e.g., conidiophores, ascomata) were excised and suspended in sterile water to prepare spore suspensions. After serial dilution (10^−1^ to 10^−4^), 150 μL of each suspension was plated in triplicate on water agar (WA) supplemented with 100 μg/mL ampicillin and 100 μg/mL streptomycin to suppress bacterial contamination. After 24–36 h of incubation at 25 °C, germinated spores were individually transferred to potato dextrose agar (PDA) containing the same antibiotic concentrations (PDA-AS). Axenic cultures were established by subculturing single colonies onto fresh PDA.
For samples lacking visible sporulation, a tissue isolation method modified from Zhang et al. (2022) and Xiao et al. (2021) was used [17,18]. Sections (~3 × 3 mm) from the interface of healthy and diseased tissue were surface sterilized in 70% ethanol (1 min), followed by 1% NaClO (1 min), and rinsed three times in sterile distilled water. After drying on sterile filter paper, tissues were placed on half-strength PDA-AS and incubated at 25 °C. Emerging hyphal tips were transferred to PDA to establish pure cultures. All isolates and reference specimens were deposited at Changchun University Fungal Culture Collection (CCUCC), China.
2.3. Morphological Identification
Morphological characterization was conducted based on both macroscopic and microscopic features (Supplementary Table S1). Colony morphology, pigmentation, and growth rate were assessed after 7 days of incubation on PDA at 25 °C in the dark. Colony color was referenced using the Methuen Handbook of Colour [19]. Micromorphological features were examined using carnation leaf agar [20] and PDA, following the methods of Leslie and Summerell (2006) and Lombard et al. (2019) [21,22]. Observed traits included the presence and structure of sporodochia, conidiophores, phialides, aerial and sporodochial conidia, and chlamydospores. Samples were mounted in water and examined under a Carl Zeiss Axioscope 5 microscope (Carl Zeiss Microscopy GmbH, Oberkochen, Germany) equipped with Differential Interference Contrast (DIC) optics. Images were captured using a Carl Zeiss Axiocam 712 mono digital camera (Carl Zeiss Microscopy GmbH, Oberkochen, Germany), and at least 30 measurements were taken per structure using ZEISS ZEN 3.10 software.
2.4. DNA Extraction, PCR Amplification and Sequencing
Genomic DNA was extracted from 7-day-old cultures of each isolate grown on PDA at 25 °C using a plant DNA extraction kit (EE111-02; Beijing TransGen Biotech Co., Ltd., Beijing, China) according to the manufacturer’s instructions. Four genetic loci were targeted for amplification: the internal transcribed spacer (ITS) region using primers ITS4/ITS5 [23], calmodulin (CaM) using CL1/CL2A [24], translation elongation factor 1-alpha (tef1) using EF-1/EF-2 [25,26], and the second largest subunit of RNA polymerase II (rpb2) using fRPB2-5F2/7cR [27,28].
PCR reactions were conducted in a 50 µL volume containing 25 µL of 2× ExTaq PCR Mix (Sangon Biotech, Shanghai, China), 19 µL nuclease-free water, 0.5 µL of each primer (10 µM), and 5 µL of DNA template. Amplification conditions followed published protocols (Table 1). PCR products were verified by 1% agarose gel electrophoresis and sequenced by Sangon Biotechnology (Shanghai, China). Sequences were edited and assembled using PhyloSuite v1.2.3. All newly generated sequences were deposited in GenBank (Table 2).
2.5. Phylogenetic Analysis
Initial identification of all isolates was performed by comparing the ITS sequences against the NCBI GenBank database using BLASTn (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 15 April 2025). For accurate species-level resolution, phylogenetic analyses were conducted on a subset of representative isolates using a multi-locus dataset comprising the internal transcribed spacer (ITS) region, the translation elongation factor 1-alpha (tef1), calmodulin (CaM), and the second largest subunit of RNA polymerase II (rpb2).
Reference sequences from type or ex-type strains of Fusarium species were retrieved from GenBank (Table 3) [29,30,31]. Multiple sequences (ITS, tef1, CaM and rpb2) were aligned using MAFFT within PhyloSuite v1.2.3 with auto strategy, and misalignments were corrected manually where necessary. Then, two independent phylogenetic algorithms, Maximum Likelihood (ML) and Bayesian Inference (BI), were employed for phylogenetic analyses according to the sequences (ITS, tef1, and rpb2) concatenated in PhyloSuite v1.2.3.
Specifically, phylogenetic analyses of TJS-16-2 and related strains were performed using a combined CaM, rpb2, and tef1 dataset. The CaM gene sequence of TJS-16-2 was deposited in GenBank under accession number PQ595068. The CaM gene was selected because it is a core barcode for the identification of Fusarium species and provides high resolution for distinguishing closely related taxa, including strains such as TJS-16-2 that are prone to misidentification.
The Maximum-Likelihood (ML) analyses were carried out using MEGA X v10.2.6, with 1000 bootstrap replicates under the selected model tested using the MEGA X v10.2.6 software’s integrated Model Selection tool [32]. The Bayesian Inference (BI) analyses were performed using PhyloSuite v1.2.3, and modelFinder within PhyloSuite v1.2.3 was used to estimate the best-fitting model for sequences used in this study. For both the ML and BI analyses, the phylogenetic trees were viewed in MEGA X v10.2.6 and FigTree v1.4.4 (http://tree.bio.ed.ac.uk/software/figtree, accessed on 16 September 2025), respectively.
2.6. Pathogenicity Testing
To determine whether the 12 Fusarium species isolated from diseased Morchella ascocarps were all pathogenic, pathogenicity verification was carried out according to Koch’s postulates. One strain from each of the 12 Fusarium species was selected as a test isolate (representative isolates were selected based on geographic origin and colony morphology, Supplementary Table S2). Each isolate was grown on PDA for 14 days at 25 ± 1 °C in darkness to obtain active mycelial growth. Colonies were rinsed with sterile distilled water, suspensions were filtered through cheesecloth and adjusted to 1 × 10^6^ spores/mL using a hemocytometer and a 10 μL droplet of the spore suspension was applied onto the stipe surface. Sterile water served as the control. Each isolate was inoculated onto three independent ascocarps of M. sextelata (biological replicates), which were then transferred to an artificial climate chamber maintained at 18 °C and 90% relative humidity in darkness. Symptom development was monitored daily for 5 days post-inoculation. Lesion average diameter (mm) was measured using a digital caliper at 4 days post-inoculation.
To complete Koch’s postulates, symptomatic tissues were excised, surface-sterilized, and re-cultured on PDA [33]. The identity of re-isolated fungi was confirmed by morphological examination and sequencing of the internal transcribed spacer (ITS) region, following Zhang et al. (2021) [34]. All assays were conducted in triplicate, and the experiment was repeated twice to validate reproducibility.
2.7. Data Analysis
Pathogenicity data were subjected to one-way analysis of variance (ANOVA) using SPSS v27 (IBM Corp., Armonk, NY, USA). Prior to analysis, normality and homogeneity of variance tests were performed to verify the assumptions required for ANOVA. Post hoc comparisons of lesion sizes were conducted using Duncan’s multiple range test at p < 0.05. Because the sample size was equal across groups and the number of treatments was relatively large, Duncan’s test was selected for its efficiency and suitability for the present analysis. Graphical data (e.g., lesion size categorization) were visualized using Origin 2024 (OriginLab Corporation, Northampton, MA, USA).
3. Results
3.1. Symptom Description and Survey Findings
Symptoms observed in the field were consistent across sites and included dense, white, villous mycelial overgrowth, premature wilting, decay of ascocarps, and necrotic lesions (Figure 1B), in contrast to healthy ascocarps (Figure 1A).
Based on field survey results and molecular identification, the presence and diversity of Fusarium species varied by region (Figure 2). Northern and north-central provinces, including Heilongjiang, Shaanxi, and Hebei, exhibited higher species richness and isolation frequencies compared with southern provinces. Isolation frequency was defined as the proportion of Fusarium species isolated from a single sampling location (province/municipality) relative to the total number of Fusarium species identified across all locations.
Other fungal genera isolated from the samples were distinguished from Fusarium based on macroscopic characteristics (e.g., mycelial morphology and colony color), microscopic features (e.g., macroconidia), and phylogenetic analysis using the ITS single-gene marker.
Fusarium species were isolated from morel cultivation bases in Heilongjiang, Hebei, Gansu, Qinghai, Shanxi, Shandong, and Henan provinces, as well as Tianjin Municipality. In contrast, no Fusarium isolates were obtained from Jilin, Liaoning, Shaanxi, Hubei, Sichuan, Yunnan, or Jiangsu provinces, or from the Inner Mongolia Autonomous Region, among the 16 provinces and municipalities surveyed (Figure 2). Pie charts at each sampling site illustrate the relative abundance of each Fusarium species identified. Notably, F. acuminatum and F. verticillioides were widely distributed and frequently isolated from multiple locations.
3.2. Molecular Identification
BLAST analysis of ITS sequences from 120 isolates obtained from symptomatic morel ascocarps revealed that all belonged to the genus Fusarium. To resolve species, the isolates were initially grouped based on colonial morphology and micromorphological characteristics. Representative isolates were then selected from each group by considering their geographic distribution, such that isolates from locations with higher isolation frequencies were proportionally represented. In total, 27 representative isolates were selected for multilocus phylogenetic analyses using ITS, tef1, CaM, and rpb2 sequences. Reference species included in the phylogenetic analyses belonged to the same species complexes and exhibited high phylogenetic relatedness to the unidentified isolates based on tef1 sequence comparisons, thereby improving the robustness of species delimitation.
The concatenated phylogenetic trees (ITS-tef1-rpb2 and CaM-tef1-rpb2; Figure 3 and Figure 4) resolved 12 distinct Fusarium species: F. acuminatum, F. avenaceum, F. clavum, F. compactum, F. falciforme, F. flocciferum, F. ipomoeae, F. mucidum, F. oxysporum, F. proliferatum, F. subglutinans, and F. verticillioides. These species were grouped into five Fusarium species complexes, namely the F. tricinctum species complex (FtSC), F. oxysporum species complex (FoCS), F. fujikuroi species complex (FfSC), F. solani species complex (FsSC) and F. incarnatum-equiseti species complex (FieSC). High statistical support was observed at most nodes, with maximum likelihood (ML) bootstrap values ≥ 70% and Bayesian posterior probabilities (BPP) ≥ 0.90. Fusarium concolor NRRL 13459 and Bisifusarium aseptatum LC1075 were used as outgroups to root the trees.
3.3. Morphological Characteristics
Morphological characteristics of the twelve Fusarium species isolated from symptomatic ascocarps of M. sextelata are summarized in Supplementary Table S1, with representative structures illustrated in Figure 5 and Figure 6. All isolates produced rapidly growing colonies on PDA, exhibiting diverse pigmentation and aerial mycelial textures ranging from sparse and villiform to dense and flocculent. All these characteristics were observed from strains cultured on PDA medium.
Colony features were generally consistent with descriptions of respective species in Leslie and Summerell (2006) [21]. For example, isolates of F. acuminatum and F. flocciferum formed compact, pinkish colonies with rose pigmentation, while F. clavum and F. compactum showed dense, flocculent growth and produced yellow to brown hues. In contrast, F. oxysporum and F. proliferatum colonies were villiform with violet pigmentation. Microscopically, all isolates produced typical fusarioid macroconidia, with the exception of F. verticillioides. Such spores were falcate to fusiform, hyaline, and varied in septation (1–7 septa). In addition, most species (excluding F. ipomoeae) could also produce oval to reniform microconidia from mono- or polyphialides. Notably, F. verticillioides and F. proliferatum produced chains of microconidia but lacked chlamydospores, while chlamydospores were abundant and diagnostic in species such as F. acuminatum, F. mucidum, and F. compactum. Sporodochial structures were observed in most species but lacking in F. flocciferum and F. mucidum.
Diagnostic traits distinguishing among species are detailed in Supplementary Table S1. Morphological observations were consistent with molecular identification and supported delimitation of Fusarium species.
3.4. Pathogenicity Tests
Representative isolates of 12 Fusarium species were selected for pathogenicity assays (Supplementary Table S2). Isolates were considered virulent when obvious brown necrotic lesions developed near the inoculation sites on morel ascocarps shortly after inoculation, resulting in tissue damage. No obvious disease symptoms were observed within 1 to 2 days after inoculation. At 4 days post-inoculation, control ascocarps showed no symptoms (Figure 7A). In contrast, all twelve tested isolates induced distinct brown necrotic lesions with mean diameters ranging from 2.42 ± 0.29 to 4.75 ± 0.25 mm (n = 3) around the inoculation sites on morel ascocarps (Figure 7B–M). This demonstrated that all the identified Fusarium species were virulent to morel ascocarps (Figure 7 and Figure 8). All inoculated isolates were successfully re-isolated from symptomatic tissues, and their identity was confirmed via morphological and molecular methods. No fungi were recovered from the control group. These results fulfilled Koch’s postulates and confirmed the pathogenicity of all tested Fusarium species toward M. sextelata.
4. Discussion
Many members of the family Nectriaceae are known to infect fungal ascocarps, including species in the genera Dialonectria and Neonectria. The genus Fusarium, which also belongs to Nectriaceae, is large and ecologically diverse, encompassing species that function as saprotrophs, endophytes, or pathogens infecting a wide range of edible mushrooms and plant hosts [30,35,36]. Fusarium incarnatum was the first species reported to infect morels, and F. nematophilum was later confirmed as the causal agent of morel white mold disease (WMD). Other Fusarium species are known pathogens of edible mushrooms: F. moniliforme causes wilt disease in Pleurotus ostreatus, F. solani induces wilt disease in Agaricus bisporus and leads to yield losses, and F. chlamydosporum is responsible for white hair disease in Auricularia auricula. In addition, Fusarium species are among the most important pathogens of cereals and other crops, particularly wheat, maize, and soybean, posing major constraints to modern agricultural production [37,38,39,40]. Recent studies have further shown that some Fusarium species possess entomopathogenic potential [41], highlighting the broad pathogenic spectrum of this genus.
With the rapid expansion of Morchella cultivation, fungal diseases caused by Fusarium and other pathogens have become an increasing concern for producers. Fusarium species are widely distributed in nature; however, based on current research, only F. incarnatum and F. nematophilum have been identified as pathogens of morel ascocarps [29,42,43], and the involvement of other Fusarium species in WMD has remained poorly understood. To address this knowledge gap, we isolated 120 Fusarium strains from 832 diseased morel ascocarps and identified twelve Fusarium species associated with WMD in cultivated morels. This study represents the first report demonstrating that these twelve Fusarium species are pathogenic to cultivated morels.
Among the twelve pathogenic species identified, only F. clavum, F. compactum, and F. ipomoeae belong to the F. incarnatum–equiseti species complex (FieSC), together with the previously reported F. incarnatum and F. nematophilum. The remaining nine species were assigned to the F. fujikuroi species complex (FfSC), F. oxysporum species complex (FoSC), F. solani species complex (FsSC), and F. tricinctum species complex (FtSC). Unlike previous studies, which focused primarily on the FieSC, the present study demonstrates that Fusarium species from multiple species complexes beyond the FieSC can also cause WMD in morels, thereby expanding the known etiological diversity of this disease.
Morphological comparisons further supported the distinction between the Fusarium species identified in this study and previously reported pathogens of morels. In the study by Guo et al. [42], colonies of F. incarnatum were described as white to brown with slender, tapering macroconidia, characteristics that are partly shared with F. clavum, F. compactum, and F. ipomoeae. However, F. incarnatum has been reported to produce a distinct moldy odor, whereas no obvious odor was observed for F. clavum, F. compactum, or F. ipomoeae in the present study. In addition, microconidia are absent in F. ipomoeae [30], further differentiating it from F. incarnatum. In a previous study on stipe rot of M. sextelata caused by F. nematophilum, this species exhibited colony characteristics similar to Pseudodiploospora longispora, including white colonies with regular margins and distinct concentric rings [8]. In contrast, the Fusarium isolates obtained in this study produced pale rose, purple, or brown pigments, and none formed concentric rings, confirming that they are distinct from previously reported morel pathogens. Notably, F. proliferatum and F. verticillioides formed characteristic microconidial chains [44,45], and F. mucidum produced short, stout macroconidia. Furthermore, F. avenaceum, F. subglutinans, and F. verticillioides did not produce chlamydospores [46,47]. These morphological traits clearly distinguish the species identified in this study from F. incarnatum and F. nematophilum.
The frequent recovery of F. verticillioides, a well-known pathogen of maize and other cereals [48,49,50], suggests that certain Fusarium species possess strong ecological adaptability, enabling them to colonize diverse hosts, including morels. Although the precise mechanisms underlying WMD in morels remain unclear, and effective control strategies are still lacking, the results of this study highlight the importance of strict disease prevention, early detection, and integrated management in morel cultivation systems. Field investigations indicate that WMD occurrence is closely associated with high temperature and high humidity during the fruiting stage. In addition, large quantities of spawn and nutrient bags containing wheat and other plant-derived materials are commonly used in morel production. These nutrient bags are often prepared by farmers under relatively crude conditions, with inappropriate material selection and insufficient sterilization. Once grains become moldy or deteriorated, they may serve as reservoirs for pathogenic fungi, which can be introduced into cultivation soils and ultimately lead to WMD outbreaks.
To reduce the incidence of WMD, several measures are recommended, including the use of high-quality wheat grains and plant materials with thorough sterilization, maintenance of a clean production environment, regular monitoring of pathogen levels in soil and water sources, and optimization of cultivation systems to achieve effective control of temperature and humidity. When WMD symptoms are observed, priority should be given to collecting ascocarp samples showing typical symptoms at early disease stages to improve the success rate of pathogen isolation and identification.
Overall, this study demonstrates that multiple Fusarium species from diverse species complexes can cause WMD in morels, adding substantial complexity to disease management alongside other known pathogens such as Pseudodiploospora, Clonostachys, and Paecilomyces. The combined use of morphological characterization and multilocus phylogenetic analyses provides a robust framework for accurate pathogen identification and will facilitate the development of effective management strategies for WMD in Morchella cultivation. Future studies will focus on elucidating the biological characteristics of the isolated strains and exploring targeted prevention and control technologies based on these findings.
5. Conclusions
This study revealed a diverse range of Fusarium species associated with WMD in cultivated morels in China and clarified their pathogenic roles. Twelve species across five major Fusarium species complexes were identified, and all of these species exhibited virulence to morel ascocarps. These findings expand our understanding of the pathogens affecting Morchella and highlight the need for effective diagnostics and integrated management to reduce disease impacts in morel cultivation. Future research should explore how environmental and farming practices influence the spread and severity of these pathogens to support sustainable morel production.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Tietel Z. Masaphy S. True morels (Morchella)-nutritional and phytochemical composition, health benefits and flavor: A review Crit. Rev. Food Sci. Nutr.2018581888190110.1080/10408398.2017.128526928350213 · doi ↗ · pubmed ↗
- 2Cai Y. Ma X. Liu W. Review on the nutritional value and health care functions of Morchella Edible Med. Mushrooms 2021292027
- 3Rotzoll N. Dunkel A. Hofmann T. Quantitative studies, taste reconstitution, and omission experiments on the key taste compounds in Morel mushrooms (Morchella deliciosa fr.)J. Agric. Food Chem.2006542705271110.1021/jf 053131 y 16569064 · doi ↗ · pubmed ↗
- 4Ower R. Notes on the development of the morel ascocarp: Morchella esculenta Mycologia 19827414214410.1080/00275514.1982.12021480 · doi ↗
- 5Liu Q. Ma H. Zhang Y. Dong C. Artificial cultivation of true morels: Current state, issues and perspectives Crit. Rev. Biotechnol.20183825927110.1080/07388551.2017.133308228585444 · doi ↗ · pubmed ↗
- 6Ma F. Qiu Y. Zhang C. Xu G. Dai T. Current Situation, Problems and Countermeasures of Morel Industry in Jiangxi Province Edible Fungi China 2022418792+9710.13629/j.cnki.53-1054.2022.11.015 · doi ↗
- 7He X.-L. Peng W.-H. Miao R.-Y. Tang J. Chen Y. Liu L.-X. Wang D. Gan B.-C. White mold on cultivated morels caused by Paecilomyces penicillatus Fems Microbiol. Lett.2017364 fnx 03710.1093/femsle/fnx 03728199636 · doi ↗ · pubmed ↗
- 8Sun J. Yu S. Lu Y. Liu H. Liu X. Proposal of a new family Pseudodiploösporeaceae fam. nov. (Hypocreales) based on phylogeny of Diploöspora longispora and Paecilomyces penicillatus Mycol.-Int. J. Fungal Biol.202314607310.1080/21501203.2022.2143919 PMC 993080736816772 · doi ↗ · pubmed ↗