New Insights from Two Historic Boletellus-Type Specimens in China Based on Next-Generation Sequencing
Bing-Qian Yang, Hong-Kang Shen, Yang Wang, Xin Zhang, Fa-Ming Long, Yi-Lin He, Yan-Chun Li, Gang Wu, Jing Zhou

TL;DR
This study uses DNA sequencing to reclassify two historic mushroom species from China, resolving long-standing taxonomic uncertainties.
Contribution
The study provides new taxonomic classifications for two historic Boletellus-type specimens using genome-skimming and NGS.
Findings
B. serpentipileus is reclassified as Leccinum serpentipileum and no longer belongs to the Boletellus genus.
B. vulgaris is synonymized with A. fusisporus and is now classified under Austroboletus.
The integration of NGS data and morphological analysis clarifies the taxonomic status of these historic specimens.
Abstract
Boletellus is a morphologically distinctive genus within the family Boletaceae, characterized by basidiospores with longitudinally striate ornamentation. Although the species diversity of this genus in China has been well documented in recent years, several historically described species published by early Chinese mycologists have been largely overlooked. To clarify the taxonomic identities of these historic Boletellus species from China, this study applied a genome-skimming approach to perform next-generation sequencing (NGS) on the historical type specimens of B. serpentipileus and B. vulgaris. The integration of NGS data with Sanger sequencing and morphological re-examination enabled a comprehensive taxonomic reassessment, which revealed that B. serpentipileus and B. vulgaris are not members of Boletellus, but belong to Leccinum and Austroboletus, respectively. Accordingly, the new…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
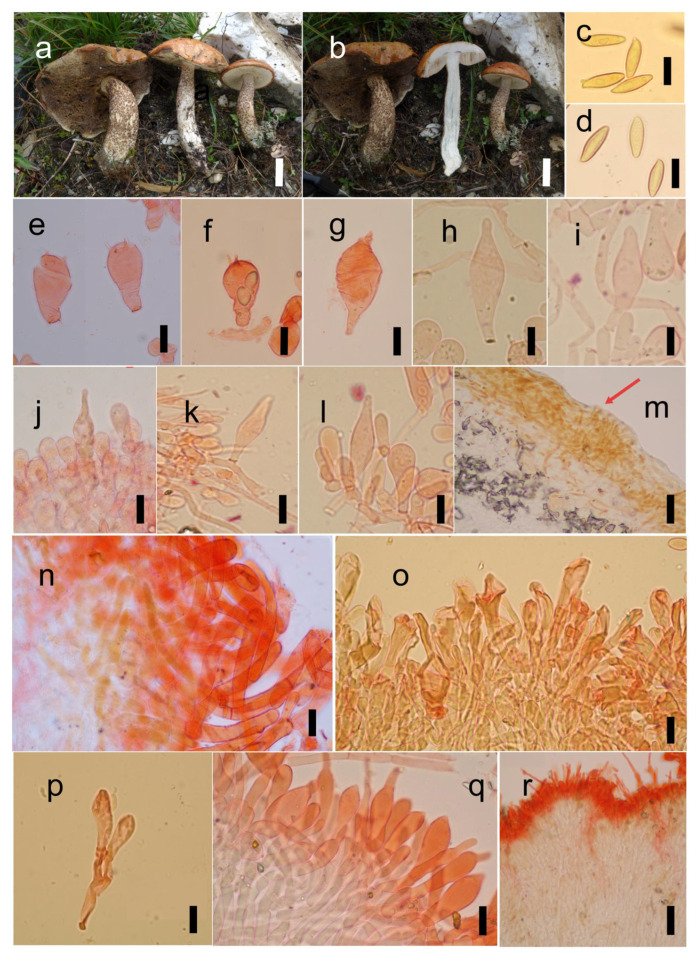 Figure 4
Figure 4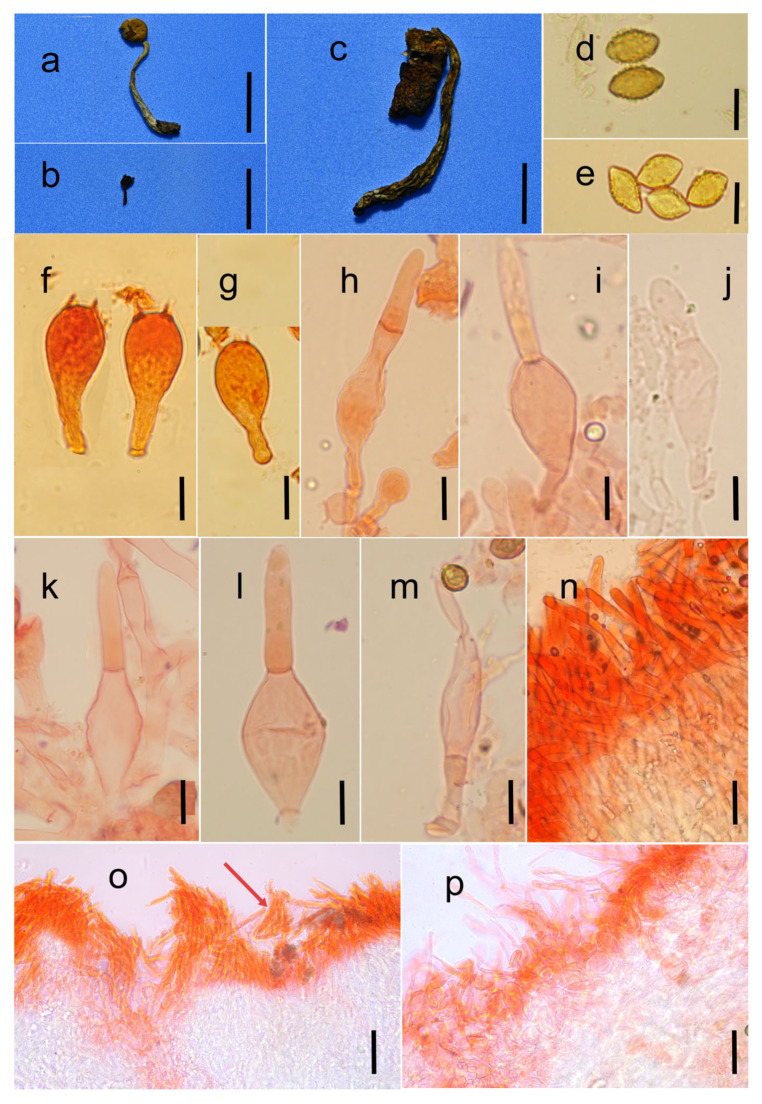 Figure 5
Figure 5- —Biological Resources Programme, Chinese Academy of Sciences
- —National Natural Science Foundation of China
- —Natural Science Foundation of Yunnan Province
- —CAS “Light of West China” Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Slime Mold and Myxomycetes Research · Plant Pathogens and Fungal Diseases
1. Introduction
The genus Boletellus was originally established by Murrill [1], based on collections from the southeastern USA, to accommodate Boletus ananas [2]. The diagnostic features of the genus include the pileus covered with erect, conical, or appressed fibrillose scales; context typically yellow and often bruising blue or sometimes unchanged; and basidiospores bearing striate ornamentation [3,4]. According to the recent research by Xue et al. (2023) [5], there were 17 species of Boletellus recorded in China. However, due to limited study of type specimens of some early described species, the true species diversity within this group remains to be clarified. For example, despite being described decades ago, the two Boletellus species, namely B. serpentipileus [6] and B. vulgaris [7], have not been revisited, leaving their true taxonomic positions unresolved.
Genome skimming is a low-coverage whole-genome sequencing strategy based on next-generation sequencing (NGS) technology, enabling efficient retrieval of phylogenetic marker genes and genomic fragment information [8]. Unlike traditional genome sequencing methods, which require complex sample preparation and involve high-cost procedures, genome skimming can obtain genomic data more rapidly and at a lower cost [9]. This approach holds significant potential to become a mainstream technology in genomic research and has been increasingly applied in recent years to fungal systematics and taxonomic identification studies [10,11]. This potential makes it possible to revisit early described Boletellus species, which were originally defined solely by morphological data.
By integrating NGS data with conventional morphological and molecular evidence, this study aims to (1) clarify the phylogenetic placements of B. serpentipileus and B. vulgaris; (2) evaluate the consistency between their morphological characteristics and molecular phylogenetic relationships; and (3) to propose the potential taxonomic revisions.
2. Materials and Methods
2.1. Morphological Characterization
Voucher specimens in this study were deposited in the Herbarium of Cryptogams at the Kunming Institute of Botany, Chinese Academy of Sciences (KUN-HKAS), and the Fungarium of the Guangdong Institute of Microbiology (GDGM). Macroscopic descriptions were based on field notes and digital images of basidiomata. Microstructural features were examined by light microscopy, whereas basidiospore ornamentation was observed under a scanning electron microscope (SEM) ZEISS Sigma 300 [12]. Tissue sections were prepared freehand, rehydrated in 5% KOH solution (Guangdong Guanghua Sci-Tech Co., Ltd., Guangzhou, Guangdong, China), and stained with 1% Congo red (Tianjin Guangfu Technology Development Co., Ltd., Tianjin, China) when necessary. Photomicrographs were captured using a ZEISS Axio Scope A1 microscope (ZEISS Technology (Suzhou) Co., Ltd., Suzhou, China) equipped with a Motic MG T45 digital camera system. Basidiospore measurements are expressed as “n/m/p”, indicating that n basidiospores were measured from m basidiomata of p collections. Spore dimensions (length/width) are presented as “(a–)b–c(–d)”, where the range b–c contains 90% of the measured values, with the extreme values a (minimum) and d (maximum) shown in parentheses. The length/width ratio of basidiospores is denoted by “Q”, while “Q” (in bold) represents the arithmetic mean ± standard deviation of the basidiospore length/width ratios.
2.2. DNA Extraction and Sequencing Methods
This study employed both Sanger sequencing and next-generation sequencing techniques. Sanger sequencing was preferentially applied to all specimens; however, for those yielding poor sequencing quality, NGS was subsequently employed. In the Sanger sequencing process, total genomic DNA was extracted from dried samples using the Ezup Column Fungal Genomic DNA Purification Kit (Sangon Biotech Co., Ltd., Shanghai, China), from materials dried according to the manufacturer’s instructions. The nuclear rDNA region, comprising the internal transcribed spacers 1 and 2 with the 5.8S rDNA (ITS) and the nuclear ribosomal large subunit (nrLSU), were amplified using the universal primer pairs ITS1-F (or ITS5) + ITS4 [13,14] and LROR + LR5 [15], respectively. For amplification of partial sequences of the RNA polymerase II largest subunit gene (rpb1), the RNA polymerase II second largest subunit gene (rpb2), the second-largest subunit (rpb2), and the translation elongation factor 1-α gene (tef1-α), the specific primers RPB1-B-F + RPB1-B-R (or RPB2-B-F2 + RPB2-B-R) and EF1-B-F2 + EF1-B-R, designed by Wu et al. (2014) [16], were employed. PCR amplification was performed using the following reaction system (25 µL total volume): 12.5 µL of 2× Hieff^®^ PCR Master Mix (Yeasen Biotechnology (Shanghai) Co., Ltd., Shanghai, China), 9.5 µL of ddH_2_O (Sangon Biotech (Shanghai) Co., Ltd., Shanghai, China), 1 µL each of forward and reverse primers (100 µM), and 1 µL of total DNA solution. The reaction conditions were as follows: initial denaturation at 94 °C for 4 min, followed by 35 cycles of denaturation at 94 °C for 60 s, annealing at 48 °C, 52 °C, or 53 °C for 60 s, and extension at 72 °C for 80 s, with a final extension at 72 °C for 8 min. The PCR products were resolved by electrophoresis on 1% agarose gels in 1× TAE buffer. Amplification results were visualized and documented using a gel imaging system. Positive reactions containing the target band were then sequenced on an ABI-3730-XL DNA Analyzer (Applied Biosystems, Foster City, CA, USA).
Next-generation sequencing was applied to historical type specimens (more than 20 years old) of B. serpentipileus (KUN-HKAS31116) and B. vulgaris (GDGM4454) and conducted on the MGI DNBSEQ-T7 platform, generating about 5 Gb of sequencing data per sample. For genomic data analysis, raw reads were quality-controlled using FASTP v1.0.164 (Shenzhen Institutes of Advanced Technology, Chinese Academy of Sciences, Shenzhen, China) [17] and assembled with SPAdes v4.2.065 (Saint Petersburg State University, St. Petersburg, Russia) [18] under default parameters. Detailed procedures, parameter settings, and result interpretations for quality control and assembly, respectively, are available at the following links: https://github.com/OpenGene/fastp and https://github.com/ablab/spades (accessed on 1 March 2025). Subsequently, library comparisons were performed, and the target fragments—ITS, nrLSU, tef1-α, rpb1, and rpb2—were extracted from the assembled genome contigs [19]. The obtained sequences were double-checked using NCBI’s BLAST (available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 20 March 2025) to assess their reliability. Through this approach, a total of 7 sequences were obtained for the aforementioned two species. The specific operational procedures for sequence extraction are as follows:
Build a comparison database
makeblastdb -in ../contigs.fasta -dbtype nucl -parse_seqids -out index
Target fragment extraction analysis
time blastn
-db index
-query 1.fas
-outfmt ‘6 delim= @ sseq’
-num_threads 18 -out 2.fas
1.fas: sequence file for comparison
2.fas: output file for target sequences
2.3. Sequence Alignment and Matrix Assembly
For the assembled genomic data, we used BUSCO v5.5.0 (University of Geneva, Geneva, Switzerland) [20] to assess the assembly quality of the obtained contigs.fasta. Since the specimens belong to the order Boletales, the “boletales_odb10” lineage-specific dataset was utilized for this assessment. In this study, due to the poor sequencing quality of both the type specimen B. serpentipileus (KUN-HKAS31116) and the type specimen B. vulgaris (GDGM4454), the proportion of complete single-copy orthologous genes was relatively low, which was insufficient to support the construction of a phylogenomic tree. Therefore, we instead employed a method involving the selection of gene sequence fragments for tree reconstruction.
For the sequences obtained in this study, we conducted preliminary analysis using the BLASTn program of the GenBank database. Results with sequence coverage exceeding 90% and similarity above 96% were selected, with one to three representative sequences chosen per species. These downloaded sequences were merged with those we newly generated. For the genus Leccinum, phylogenetic analysis was conducted using four gene matrices—nrLSU, tef1-α, rpb1, and rpb2. ITS sequences were excluded due to their excessive variation among Leccinum species [21]. For the genus Austroboletus, five matrices were generated: ITS, nrLSU, tef1-α, rpb1, and rpb2. The following steps on phylogenetic analysis were performed using PhyloSuite v1.2.3 (Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China) [22]: sequence alignment was performed with MAFFT v7.313 (Immunology Frontier Research Center, Osaka University, Osaka, Japan) using the simplest FFT-NS-1 strategy [23]; aligned sequences were trimmed using the “manual trim” mode of trimAl v1.2 (Centre for Genomic Regulation, Barcelona, Spain) (command line parameters: -cons 25.0 -gt 0.9 -st 0.0 -w 1.0) [24]; and the “Concatenate Sequence” tool was used to merge multiple sequences in a specified order to generate a new, complete concatenated sequence.
2.4. Molecular Phylogenetic Analyses
For the combined dataset, molecular phylogenetic analyses were conducted using Maximum Likelihood (ML) and Bayesian Inference (BI) methods. To ensure the biological rationality of the combined analysis, phylogenetic trees were first constructed separately for each gene. Topological congruence was assessed based on the criterion proposed by Nuhn et al. (2013) [25], which considers clades with conflicting topologies supported by bootstrap values >70% as significantly incongruent. No significant topological conflicts were detected among the individual gene trees; therefore, the sequences were concatenated for subsequent joint analyses. Maximum Likelihood analysis was performed using RAxML v8.2.10 (Heidelberg Institute for Theoretical Studies, Heidelberg, Germany) [26] called via the graphical interface raxmlGUI 2.0.0-beta.3 (University of Gothenburg, Gothenburg, Sweden) [27]. The substitution model was set to GTRGAMMAI, with all other parameters kept at their default settings. Bootstrap support values were obtained from 1000 non-parametric bootstrap replicates. All analyses were run on a Windows system utilizing 4 CPU threads. Bayesian Inference Analysis was carried out using PhyloSuite v1.2.3 according to the following workflow: Prior to analysis, PartitionFinder 2 (Australian National University, Canberra, Australia; Macquarie University, Sydney, Australia) [28] was used to select optimal partitioning schemes and substitution models under the corrected Akaike Information Criterion (AICc). The search was conducted using a greedy algorithm with the candidate model set comprising 24 nucleotide substitution models (ranging from JC to GTR + I + G). The model selection results indicated the following best-fit models: for the genus Leccinum matrix, GTR + I + G for nrLSU, SYM + G for tef1-α and rpb1, and K80 + G for rpb2; for the genus Austroboletus matrix, GTR + I + G for ITS and nrLSU, SYM + G for tef1-α and rpb2, and HKY + I + G for rpb1. Bayesian Inference was subsequently executed in MrBayes v3.2.6 (Swedish Museum of Natural History, Stockholm, Sweden; Florida State University, Tallahassee, FL, USA) [29]. The evolutionary model for each partition was specified via the nst and rate parameters to correspond with the optimal models previously identified by PartitionFinder 2. Two independent runs were performed simultaneously, each consisting of one cold chain and three heated chains (four chains in total). The analysis was run for 1.0 × 10^6^ generations, sampling tree topologies and model parameters every 1000 generations. MrBayes utilizes Markov Chain Monte Carlo (MCMC) for Bayesian Phylogenetic Inference. The ideal state of the MCMC run is to reach the target posterior probability distribution. During execution, the “Progress” window displays the Average Standard Deviation of Split Frequencies (ASDSF) in real time. An ASDSF value below 0.01 was considered indicative of convergence. If the run terminated without achieving convergence, the analysis was extended until convergence was reached, after which the final result file (con.tre) was obtained. For this study, both the Leccinum dataset and the Austroboletus dataset were ultimately run for 2.0 × 10^6^ generations.
3. Results
3.1. Molecular Phylogeny
In this study, a total of 29 sequences were newly generated, comprising five ITS, seven nrLSU, six tef1-α, two rpb1, and five rpb2 sequences. Specimens KUN-HKAS152686, KUN-HKAS152693, and KUN-HKAS154141 were sequenced using the Sanger method. Specimens KUN-HKAS31116, GDGM4454, and KUN-HKAS154142 were sequenced using NGS. For specimen KUN-HKAS123627, the ITS and nrLSU sequences were obtained via Sanger sequencing, while the tef1-α, rpb1, and rpb2 sequences were obtained via NGS. All of the sequences have been submitted to GenBank (Table 1). For the genus Leccinum, a concatenated phylogenetic analysis was performed based on the nrLSU, tef1-α, rpb1, and rpb2; the combined dataset comprised 17 species and 2535 nucleotide sites. For the genus Austroboletus, a concatenated phylogenetic analysis was conducted using ITS, nrLSU, tef1-α, rpb1, and rpb2; the combined dataset comprised 22 phylogenetic species and 2616 nucleotide sites. The trees reconstructed using Maximum Likelihood (ML) and Bayesian Inference (BI) methods exhibited similar topologies. Therefore, only the ML trees were presented (Figure 1 and Figure 2).
In the Leccinum analysis, we performed ITS sequence alignment for a specimen (KUN-HKAS154142) collected from France. The alignment showed 100% coverage and greater than 99% similarity to that of an authentic L. aurantiacum specimen (Champ-41) from France, which is the type locality for this species. Therefore, this specimen (KUN-HKAS154142) was identified as L. aurantiacum, and its DNA sequences of other genes were included in the multi-gene (nrLSU + tef1-α + rpb1 + rpb2) phylogenetic analysis. The results revealed that the clade containing the type specimen of B. serpentipileus (KUN-HKAS31116) was clearly positioned outside the genus Boletellus. Instead, it was clustered within Leccinum and formed a monophyletic group distinct from L. aurantiacum (KUN-HKAS154142) (Figure 1). In the Austroboletus analysis, phylogenetic analysis using a multi-gene dataset (ITS + nrLSU + tef1-α + rpb1 + rpb2) placed the type specimen of B. vulgaris (GDGM4454) outside the genus Boletellus and, specifically, within a Austroboletus clade alongside A. fusisporus. Consequently, based on the molecular phylogenetic results and supported by the morphological evidence (as detailed below), we proposed to transfer B. serpentipileus to the genus Leccinum as L. serpentipileum, and to treat B. vulgaris as a synonym of A. fusisporus.
3.2. Morphological Study of Type Specimens
Boletellus serpentipileus was originally published by Zang & Yuan (1999) from Yunnan Province in China, with the original description stating: “Pileus 79 cm latus, convexus demum planoconvexus, siccus, rugulosus, reticulato–venosus, brunneo–flavus, serpentiformis. Contextus 11.6 cm crassus, albus, immutabilis. Hymenium aureum. Tubuli 0.51 cm longi, flavi, adnexi vel sinuato–adnexi. Pori angulares vel irregulares. 1418 per cm. Stipe 79 cm longus. 1.52.2 cm crassus, clavatus, reticulatus, basim versus bulbosus, brunneo–flavus. Mycelio albido. Basidiosporae 2123 × 66.5 μm angusto–ellipsoideae, disstincte longitudinalis rugiformis. Basidia 2530 × 810 μm, clavata. 4–sporigera. Tramae tubi paralleloneurae [6].”
Based on the re-examination of the holotype specimen of B. serpentipileus, which is in suboptimal condition, the identifiable morphological characteristics are as follows: Basidiomata medium-sized to large. Pileus yellowish brown when dry, stipe surface with squamulose ornamentation, central stipe. Basidiospores (Figure 3a–d and Figure 4c) 16–19 × 4–6 µm, smooth, elongated ellipsoid, with slightly thickened walls (approx. 0.5 µm thick). Basidia 13.5–30 × 8–10 μm, clavate, sterigmata up to 3 μm long, pale yellow in 5% KOH. Cheilo- and pleurocystidia not observed. Hymenophoral trama boletoid. Pileipellis an intricate ixotrichoderm. Stipitipellis hymeniform, approximately 60–90 μm thick; caulocystidia 24–45 × 6–13 μm, broadly fusoid–ventricose, thin-walled. Clamp connections absent in all tissues.
Boletellus vulgaris was originally published by Bi et al. (1982) from Guangdong Province in China, with the original description stating: “Pileus 1.6 cm latus, glutinosus, fulvus, margine sordide flavus, dein plano-convexus, villoso-tomentosus, margine porrectus, integris. Contexto albo, immutabili, 3 mm crasso, sapor nullus. Stipes centralis, flavobadius, 5 × 0.3 cm, leviter sinuosus, cylindraceus, solidus, sequalis vel ad basin subincreasatus, tomentosus, cuticula facile delapsus. Tubuli punicei, immutabiles, 3 mm longi, ad stipitem appendiculati, poris angularis, 1 mm diam. Sporae fusiformes, sub microscopio pallide rubropurpureae, verrucosae, 12−17 × 6.6−9 μm. Solitarius ad terram in silva [7].”
Based on the re-examination of the holotype specimen of B. vulgaris, which is in suboptimal condition, the identifiable morphological characteristics are as follows: Basidioma small. Pileus fulvous with a central stipe. Basidiospores (Figure 4e–f and Figure 5d) 13–15(–16) × 8–10 μm, verrucose, fusiform, and inequilateral in lateral view with a slightly depressed suprahilar area; smooth or exhibiting shallow pits at the apex and base, appearing brownish yellow in 5% KOH. Basidia 19.5–33 × 8–16.5 μm, clavate, with sterigmata up to 3.5 μm long and pale yellowish in 5% KOH. Cystidia, pileipellis, and stipitipellis not observed.
3.3. Taxonomy
Leccinum serpentipileum (M. Zang & M.S. Yuan) Bing-Qian Yang & G. Wu, comb. nov. (Figure 3a–d and Figure 4).
**Basionym:**Boletellus serpentipileus M. Zang & M.S. Yuan, Acta bot. Yunn. 21(1): 39 (1999).
MycoBank: MB 862341.
Type: CHINA, Sichuan Province, Ganzi Tibetan Autonomous Prefecture, Jiulong County, Jichou Mountain, 10 September 1996, Ming-Sheng Yuan 2662 (KUN-HKAS 31116).
Diagnosis: Medium to large basidiomata; a pileus surface often orange–yellow and covered with a glutinous tomentum; context white, not discoloring or turning slightly reddish when injured; basidiospores smooth and ellipsoid; and pileipellis an intricate ixotrichoderm.
Description: Basidiomata medium to large. Pileus 6–14 cm in diam., hemispherical; margin slightly incurved, orange–yellow to yellowish brown, covered with yellowish-brown tomentum, viscid when moist; context white, 1–2 cm thick, not discoloring or turning slightly reddish when injured. Hymenophore sinuate to adnate, surface initially pale yellow to white, becoming brown at maturity, staining brown when bruised, tubes about 1.5–2 cm long, white to pale yellow when young, becoming yellowish brown at maturity, turning slightly reddish when bruised; pores angular to irregular, 0.5–1 mm wide. Stipe central, 7–13 cm long, 1.5–3 cm in diam., solid, often curved, subequal, yellowish to brownish or pallid in the upper part, white to pale yellow in the middle part, gradually whitish towards the base, entirely covered with brown verruculae on the surface; context white, becoming slightly bluish-black when injured; basal mycelium whitish.
Basidiospores [42/2/4] (14–)15–18(–19) × 4–5(–6) um [Q = (2.80–)3–4.25(–4.75), Q = 3.41 ± 0.40], angusto-ellipsoideae, inequilateral in lateral view with a slight suprahilar depression, elongate ellipsoid in ventral view, yellowish to brownish in 5% KOH, smooth, slightly thick-walled (ca. 0.5 μm thick). Basidia 22–34 × 8–18 μm, clavate, four-spored, sterigmata up to 4 µm long, hyaline in 5% KOH. Hymenophoral trama boletoid. Pleurocystidia 19–44 × 6–17 μm, broadly fusoid–ventricose to subfusiform, with a rounded rostrum at the apex, hyaline in 5% KOH, thin-walled. Cheilocystidia 29–59 × 5–16 μm, fusoid–ventricose, subfusiform, or narrowly lageniform, occasionally with an elongated rostrum, slenderer than pleurocystidia, concolorous with pleurocystidia, thin-walled. Pileipellis an intricate ixotrichoderm, embedded in a gelatinous matrix, approximately 150 μm thick, composed of interwoven, smooth, thin-walled hyphae 5–10 μm wide; terminal elements 23–64 × 6–15 μm, subcylindrical, light yellowish to brownish. Stipitipellis 70–120 μm thick, hymeniform; caulocystidia 27–70(–80) × 4–16 μm, broadly fusoid–ventricose, with a bluntly acute apex or bearing a long beak, thin-walled. Stipital trama composed of longitudinally arranged hyphae 5–10 μm diam. Clamp connections absent in all tissues of the basidioma.
The microscopic characteristics of L. serpentipileum: ((a,b,d–n,q,r) KUN-HKAS 123627; (c,o,p) KUN-HKAS31116, holotype). (a,b) basidiomata, (c,d) basidiospores, (e,f) basidia, (g–i) pleurocystidia, (j–l) cheilocystidia, (m,n) pileipellis, (p) caulocystidia, and (o,q,r) stipitipellis. Scale bars: (a,b) = 3 cm; (c–l,n–q) = 10 µm; and (m,r) = 40 µm. The red arrow in subpanel m indicates the gelatinous substance.
Habitat and distribution: Solitary or scattered on the ground in subalpine coniferous forests; currently known from northwestern Yunnan and western Sichuan, China.
Additional specimens examined: CHINA, Yunnan Province, Diqing City, Shangri-La County, alt. ca. 3300 m, 1 September 2017, Gang Wu 2474 (KUN-HKAS 123627); Yunnan Province, Shangri-La County, near the Timber Inspection Station of Daxue Mountain, 8 September 2014, Jian-Wei Liu 270 (KUN-HKAS91078).
Austroboletus fusisporus (Kawam. ex Imazeki & Hongo) Wolfe, Biblthca Mycol. 69: 96 (1980) [1979] (Figure 3e–i and Figure 5).
**Basionym:**Porphyrellus fusisporus Kawam. ex Imazeki & Hongo, Acta phytotax. geobot., Kyoto 18(4): 110 (1960).
**Synonym:**Boletellus vulgaris C.S. Bi, in Bi, Loh & Zheng, Acta bot. Yunn. 4(1): 57 (1982).
Diagnosis: Very small to minute basidiomata; pink hymenophore; stipe covered with distinct pale brown reticula; amygdaliform to fusiform basidiospores ornamented with large, regular to irregular subcylindrical structures; and an ixotrichoderm pileipellis.
**Description:**Basidiomata small. Pileus 1.6–3 cm in diam., glutinous, fulvous (tawny), paler towards margin; becoming plano-convex, villose–tomentose; margin extended, forming a membranous veil that embraces the stipe at the young stage, then fragmented and appendiculate at the margin when aged; context white, 0.3–0.5 cm thick, not discoloring when injured. Hymenophore adnate, surface pale pinkish; tubes up to 0.3–0.6 cm long, pink, not discoloring when bruised; pores angular, 1 mm in diam. Stipe central, 3–5 cm long, 0.3–0.6 cm in diam., solid, slightly sinuous, cylindrical, equal or slightly thickened at base, yellowish-brown, covered with pale brown reticula on the surface; basal mycelium white.
Basidiospores [60/3/3] (10.5–)12–15(–17) × (6–)7–10 um [Q = (1.3–)1.5–1.75(–2), Q = 1.6 ± 0.15], amygdaliform to fusiform, inequilateral in lateral view with a slight suprahilar depression; amygdaliform in ventral view, yellowish to brownish yellow in 5% KOH, ornamented with large, regular to irregular, subcylindrical warts (1–1.5 μm high, 0.5–2 μm wide) on the surface, smooth or with shallow pits at the apex and base. Basidia 15–59 × 7–29 μm, clavate, four-spored, sometimes two- or three-spored, sterigmata up to 3.5 μm long, pale yellow in 5% KOH; Hymenophoral trama boletoid. Cheilo- and pleurocystidia consisting of two cells: the upper cells cylindrical to digitate, measuring 17–40 × 3–6.5 μm; the lower cells clavate to broadly clavate, measuring 25–39 × 8–17 μm. These cells contain pale yellowish brown to pale brown pigments, appearing pale brownish yellow to yellowish brown in KOH, and are thin-walled. Pileipellis an intricate ixotrichoderm, about 70–165 μm thick, composed of interwoven, smooth, thin-walled hyphae measuring 4–8 μm in diameter; terminal cells 19–78 × 3.5–8 μm, subcylindrical to subclavate, hyphae transparent yellow in 5% KOH. Caulocystidia scarce, resembling hymenial cystidia, with the upper cells measuring 12–18 × 3–5 μm and the lower cells 15–30 × 8–12 μm. Stipitipellis a hyphoepithelium: the outer layer consists of filamentous hyphae measuring 2–11 μm in width, with clavate to subcylindrical terminal cells sized 15–64 × 5–10 μm, appearing pale yellowish to brownish yellow in KOH. Clamp connections absent in all tissues of the basidioma.
The microscopic characteristics of A. fusisporus: ((a,n–p) KUN-HKAS152686; (b,d) GDGM4454 holotype of B. vulgaris; (c,e–m) KUN-HKAS152693). (a–c) basidiomata, (d,e) basidiospores, (f,g) basidia, (h–j) pleurocystidia, (k–m) cheilocystidia, (n,o) pileipellis, and (p) stipitipellis. Scale bars: (a–c) = 2 cm; (d–n) = 10 um; and (o,p) = 40 um. The red arrow in subpanel o indicates the gelatinous substance.
Habitat and distribution: Scattered on soil in tropical forests dominated by plants of the family Fagaceae; currently known in Japan [45], South Korea [46], and China [4,30].
Specimens examined: CHINA, Guangdong Province, Zhaoqing City, near Qingyun Temple on Dinghu Mountain, 19 August 1980, Zhi-Shu Bi 454 (GDGM 4454, holotype of B. vulgaris); Hunan Province, Changsha City, Lishanchong, 4 September 2023, Wu-Ping Luo, voucher no. SF1442787849763 (KUN-HKAS 152686); Yunnan Province, Lincang City, 14 July 2024, collected by an anonymous enthusiast, voucher no. SF1454082419825 (KUN-HKAS 152693).
4. Discussion
In this study, we re-examined two early described species of Boletellus, B. serpentipileus and B. vulgaris, by integrating NGS data with Sanger sequencing and morphological observation. Our results indicate that B. serpentipileus falls within the genus Leccinum, while B. vulgaris should be treated as a later synonym of A. fusisporus.
Regarding B. serpentipileus, Zang et al. (1999) [6] initially assigned this species to the genus Boletellus based on the morphological observation of distinctly longitudinally striate basidiospores. However, re-examination of the type specimen revealed basidiospores with a smooth, non-striate surface, measuring 16–19 × 4–6 µm, which differ from the original description. This morphological feature is inconsistent with Boletellus, but shows affinity with Leccinum, as the species also possesses a stipe entirely covered with brown squamules and smooth basidiospores. Accordingly, molecular phylogenetic analysis was conducted using DNA data obtained from this type specimen (KUN-HKAS31116) via NGS. The results demonstrate that the corresponding clade is unequivocally positioned outside the genus Boletellus and is nested within Leccinum. Based on combined morphological and molecular phylogenetic analyses, this study proposes the reclassification of this species into the genus Leccinum. Morphologically, Leccinum serpentipileum resembles L. aurantiacum. However, Leccinum aurantiacum has a pileus that is vivid red to red–brown and whitish context turning violaceous gray, gray, or blackish when bruised. Furthermore, it has smaller basidia (15–25.0 × 6.5–10.5 μm) and an intricate trichodermal pileipellis, with larger terminal elements often intracellularly granular-incrusted with red–brown pigment [47].
Early mycologists considered Boletellus to encompass species with basidiospores that were smooth (over 20 μm long), reticulate, verrucose, or longitudinally striate [48,49]. According to it, Bi et al. (1982) [7] initially assigned B. vulgaris to section Allospori of the genus Boletellus due to its fusiform basidiospores with verrucose ornamentation. However, the genus Boletellus was later delimited to encompass solely species exhibiting longitudinally striate ornamentation [50]. This refined definition necessitated a re-examination of the type specimen of B. vulgaris. Our re-observation confirmed that the basidiospore ornamentation is consistent with the description by Bi et al. (1982) [7]. However, the presence of pink tubes and reticulate stipe conflict with the modern delimitation of Boletellus. To clarify its phylogenetic position, NGS was employed in this study to obtain DNA data from this type specimen (GDGM4454). Molecular phylogenetic analysis demonstrated that B. vulgaris is nested within a clade that includes A. fusisporus in the genus Austroboletus. Based on morphological and molecular evidence, this study proposes to treat B. vulgaris as a synonym of A. fusisporus. Morphologically, they share several distinctive characteristics: small to minute basidiomata; a glutinous, fulvous pileus with an extended margin; pink tubes; a densely reticulate stipe surface; and basidiospores of similar size, ornamented with irregular, subcylindrical tubercles of varying sizes; cheilo- and pleurocystidia composed of two cells; and an intricate ixotrichoderm pileipellis.
This study, based on in-depth analyses of historical specimens of the genus Boletellus, validates the feasibility of next-generation sequencing technology in successfully getting the draft genomic data of aged specimens. This approach not only provides technical support for taxonomic study on historic herbarium collections, but also opens new avenues for exploring the potential of historical specimens in the studies on phylogeography and evolutionary history.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Murrill W.A. The Boletaceae of North America-1Mycologia 19091418
- 2Curtis M.A. Contributions to the Mycology of North America Am. J. Sci. Arts 18486349353
- 3Halling R.E. Fechner N. Nuhn M. Osmundson T. Soytong K. Arora D. Binder M. Hibbett D. Evolutionary relationships of Heimioporus and Boletellus (Boletales), with an emphasis on australian taxa including new species and new combinations in Aureoboletus, Hemileccinum and Xerocomus Aust. Syst. Bot.20152812210.1071/SB 14049 · doi ↗
- 4Wu G. Li Y.C. Zhu X.T. Zhao K. Han L.H. Cui Y.Y. Li F. Xu J.P. Yang Z.L. One hundred noteworthy boletes from China Fungal Divers.2016812518810.1007/s 13225-016-0375-8 · doi ↗
- 5Xue R. Zhang X. Xu C. Xie H.J. Wu L.L. Wang Y. Tang L.P. Hao Y.J. Zhao K. Jiang S. The subfamily Xerocomoideae (Boletaceae, Boletales) in China Stud. Mycol.20231069519710.3114/sim.2022.106.0338298571 PMC 10825750 · doi ↗ · pubmed ↗
- 6Zang M. Yuan M.S. Contribution to the knowledge of new basidiomycoteous taxa from China Acta Bot. Yunnanica 1999213944
- 7Bi C.S. Loh T.C. Zheng G.Y. Bi C.S. Loh T.C. Zheng G.Y. Basidiomycetes from Dinghu mountain of China 2. some new species of Boletaceae 1Acta Bot. Yunnanica 198245564
- 8Thakur K. Next generation sequencing—Techniques and its applications J. Pharmacogn. Phytochem.2018713161320