Astaxanthin as a Therapeutic Candidate for Nociceptive and Inflammatory Pain: Mechanisms and Perspectives
Mamoru Takeda, Risako Chida

TL;DR
Astaxanthin, a natural compound, may help relieve pain and inflammation with effects similar to common drugs like ketamine and celecoxib.
Contribution
The paper presents new evidence that astaxanthin can act as a natural analgesic and anti-inflammatory agent.
Findings
Astaxanthin shows anesthetic effects on pain comparable to ketamine.
It has anti-inflammatory properties similar to the drug celecoxib.
Astaxanthin modulates neuronal excitability and inhibits cyclooxygenase-2.
Abstract
Recently, complementary and alternative medicine (CAM) has been actively employed for patients experiencing symptoms unresponsive to Western medical treatments like drug therapy. The natural compounds carotenoids and astaxanthin (AST) have demonstrated various beneficial biological actions for human health in several studies. Given their broad pharmacological activities and reduced toxicity, ASTs possess significant potential as resources for the development of natural analgesic drugs. Given recent studies showing that AST can modulate neuronal excitability, including nociceptive sensory transmission through voltage-gated Ca2+ channels and the n-methyl-D-aspartate (NMDA) glutamate receptor, and inhibit the cyclooxygenase-2 cascade, AST holds promise as a CAM, particularly as a therapeutic agent for nociceptive and pathological pain. Based on the in vivo research findings from our…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
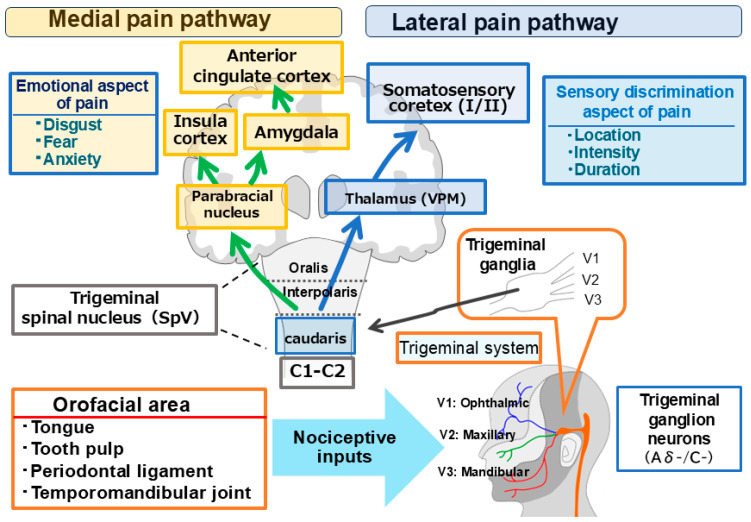 Figure 1
Figure 1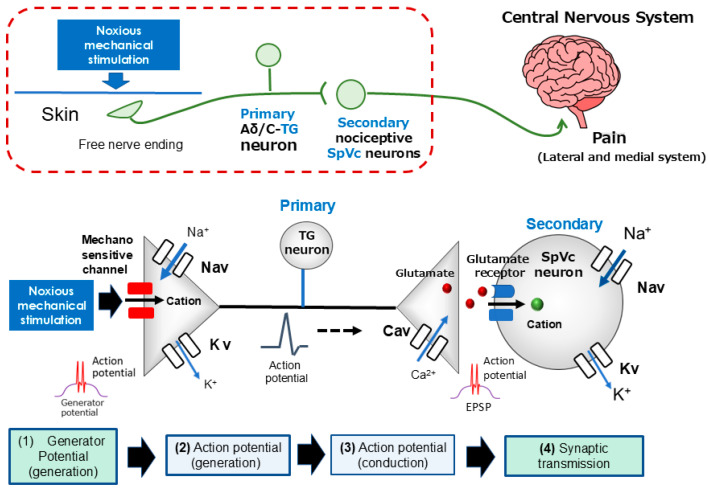 Figure 2
Figure 2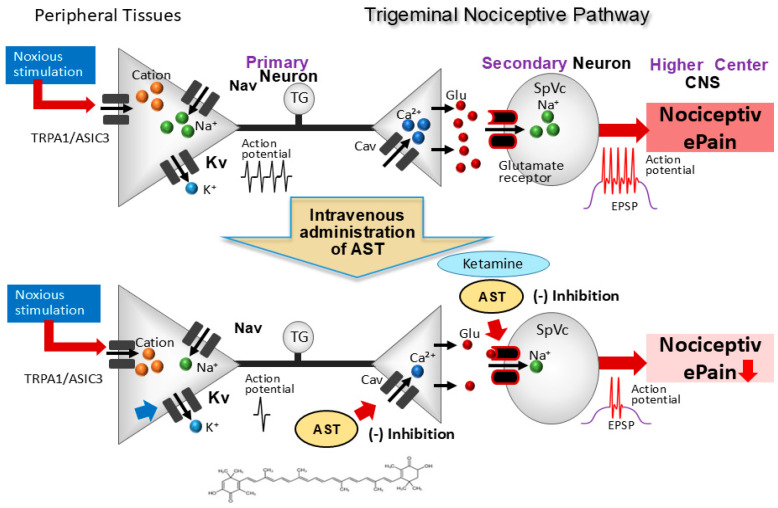 Figure 3
Figure 3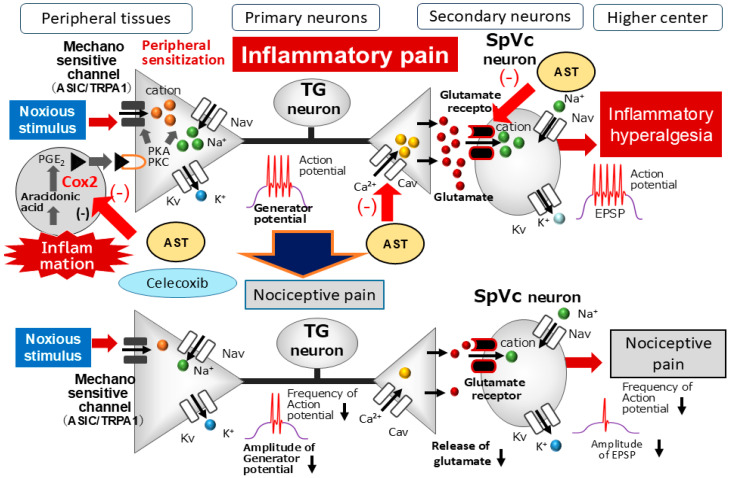 Figure 4
Figure 4- —Grant-in-Aid for Scientific Research (C) from the Japanese Society for the Promotion of Science
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntioxidant Activity and Oxidative Stress · Medicinal Plants and Neuroprotection · Coenzyme Q10 studies and effects
1. Introduction
Conventional Western pharmacological treatments for chronic pain often present limitations for certain patient populations, leading to a rising clinical interest in Complementary and Alternative Medicine (CAM) [1,2,3]. Although CAM—specifically herbal remedies and acupuncture—is sometimes characterized in Western medicine as lacking exhaustive scientific validation, a growing body of evidence highlights the therapeutic potential of natural compounds. These substances have demonstrated substantial biological activities, including antioxidant, anti-inflammatory, and cardioprotective properties, in addition to their antinociceptive effects [4,5,6,7,8].
The clinical utility of traditional analgesics is frequently hindered by severe adverse reactions associated with long-term administration. Consequently, there is a critical need to identify safer natural alternatives that provide therapeutic efficacy with minimal toxicity. Given their diverse pharmacological profiles and favorable safety margins, natural compounds represent a promising foundation for novel analgesic development. While existing local anesthetics and non-steroidal anti-inflammatory drugs (NSAIDs) are effective, they are not without significant risks; for instance, inflammation can compromise the efficacy of local anesthetics in dental contexts [9]. Furthermore, while NSAIDs such as celecoxib effectively inhibit cyclooxygenase-2 (COX-2) and prostaglandin E_2_ (PGE_2_) synthesis, their use is clinically linked to gastrointestinal ulcers and increased cardiovascular risks, such as myocardial infarction [10].
In this context, phytochemicals and fatty acids have emerged as viable CAM strategies for managing pain within inflamed tissues [11]. These compounds, derived from various dietary sources including fruits, vegetables, and marine life, encompass diverse groups such as polyphenols and carotenoids. Among these, Astaxanthin (AST)—a xanthophyll carotenoid prevalent in microalgae, crustaceans, and salmonids—has garnered particular attention [12]. AST exhibits superior antioxidant capacity compared to other carotenoids like lutein and zeaxanthin [13,14,15], and possesses a broad biological spectrum including anti-tumor, anti-diabetic, and immunomodulatory activities [16,17,18].
While the prior literature has reviewed the antinociceptive potential of carotenoids in animal models [10,19,20], a detailed synthesis of their neurophysiological mechanisms remains limited. Specifically, there is a paucity of data regarding how carotenoids influence the excitability of nociceptive neurons under pathological conditions, particularly through in vivo electrophysiological assessments. Nevertheless, recent in vitro evidence suggests that AST modulates neuronal excitability by interacting with key molecular targets, such as voltage-gated Ca^2+^ (Cav) channels, N-methyl-D-aspartate (NMDA) receptors, and COX-2 [21,22,23,24,25]. Recent work by Zhao et al. [26] showed that AST exerts potent inhibitory effects on neuroinflammation and the activation of phosphorylation of extracellular signal-regulated kinase (p-ERK)1/2, phosphorylation of p38 mitogen-activated protein kinase (p38 MAPK), and nuclear factor-kB (NF-κB) p65 nuclear translocation. Consequently, AST holds promise as a new therapeutic tool for treating neuropathic pain. In addition, Zhao et al. [27] demonstrated that AST exhibits anti-inflammatory properties mediated by p38 MAPK and nuclear factor erythroid 2-related 2 (Nrf2) /heme oxygenase-1(HO-1) in inflammatory pain models, highlighting its potential utility as a pharmacological intervention for inflammatory pain relief.
Our laboratory has recently utilized neurophysiological methods to evaluate the analgesic properties of dietary AST. Our published findings demonstrate that both acute intravenous administration and chronic intake of AST significantly attenuate nociceptive and inflammatory pain [23,24,25]. The present review aims to synthesize current knowledge regarding the role of AST in relieving nociceptive and pathological pain, discussing its potential clinical utility and the significance of its chemical structure in mediating analgesic effects, with a particular focus on recent in vivo evidence [23,24,25]. Although this study focuses on the molecular mechanisms in animal models, these findings provide a necessary foundation for future clinical applications and the development of functional foods.
2. Clinical Manifestations of Nociceptive and Pathological Pain
The critical biological necessity of pain is perhaps best illustrated by the rare genetic condition known as congenital insensitivity to pain (congenital analgesia). Individuals with this disorder possess dysfunctional nociceptors [28], rendering them entirely unable to perceive painful stimuli. This sensory deficit leads to catastrophic clinical outcomes, including recurrent fractures, joint deformities, and life-threatening complications such as sepsis or tissue necrosis from undetected burns. These severe consequences underscore the role of pain as an indispensable “biological warning system” that alerts the organism to potential or actual tissue damage, thereby ensuring survival [29]. This protective mechanism is the primary function of physiological (nociceptive) pain.
In contrast, pathological pain represents a maladaptive state in which this “biological warning system” becomes dysfunctional. This condition is defined by neuroplastic alterations in pain-transmission neurons and the persistence of nociceptive signaling, which often continues long after the initial tissue injury has resolved [29,30]. Pathological pain is generally categorized into inflammatory and neuropathic pain. Inflammatory pain results from the peripheral sensitization of nociceptors by biochemical mediators, such as PGE_2_, at the site of injury or inflammation (e.g., thermal burns or rheumatoid joints) [31].
Neuropathic pain, on the other hand, stems from direct damage to the somatosensory nervous system and persists independently of the original wound healing. Common clinical manifestations include diabetic peripheral neuropathy, sciatica, and trigeminal neuralgia. In the context of dentistry, neuropathic pain is frequently encountered as a post-operative complication following invasive procedures such as tooth extractions or dental implantation.
3. Trigeminal Pain Transmission Pathway: Overview
This review focuses on the potential contributions of AST to the alleviation of nociceptive and pathological pain, necessitating an understanding of the general characteristics of trigeminal pain pathways and nociceptive neurons (Figure 1). In the trigeminal nervous system, pain signaling is functionally bifurcated into the lateral and medial systems: the former transmits sensory-discriminative details such as pain intensity and topographical location, while the latter conveys the affective-motivational components, including the emotional perception of “unpleasantness” [32,33,34]. Nociceptive inputs from orofacial regions—including the tongue, dental pulp, periodontal ligament, and temporomandibular joint—are initially carried by trigeminal ganglion (TG) neurons to the trigeminal spinal nucleus caudalis (SpVc) and the upper cervical dorsal horn (C1–C2) [32,35]. At this junction, two distinct neuronal classes process these signals: Wide Dynamic Range (WDR) neurons, which respond to both non-nociceptive and nociceptive stimuli by increasing discharge frequency in correlation with stimulus intensity, and nociceptive-specific (NS) neurons, which react exclusively to noxious stimuli to facilitate spatial localization. Notably, WDR neurons are implicated in the development of hyperalgesia following tissue injury and inflammation [32,35]. From the SpVc/C1-2 complex, sensory-discriminative information is relayed via the ventral posteromedial thalamic nucleus to the primary and secondary somatosensory cortices; simultaneously, affective data are transmitted through the parabrachial and medial thalamic nuclei to the amygdala, insular cortex, and anterior cingulate cortex for comprehensive emotional interpretation [32,35].
4. Mechanisms of Pain Transmission: Electrical and Chemical Signaling
The transmission of nociceptive information relies on primary afferent fibers, specifically the pseudounipolar neurons of the trigeminal ganglion (TG), which are classified into thin, myelinated Aδ fibers and unmyelinated C fibers [32,35]. These fibers serve distinct functional roles: Aδ fibers mediate rapid, sharply localized “fast” pain, while C fibers conduct more diffuse, “slow” aching sensations [32,35]. Structurally, TG neurons extend peripheral axons that terminate as free nerve endings and central axons that establish chemical synapses with second-order neurons in the central nervous system (CNS). As illustrated in Figure 2, this sensory processing sequence encompasses four fundamental stages: (i) transduction of external noxious energy into electrical signals, (ii) generation of action potentials, (iii) conduction along the axon, and (iv) synaptic transmission within the CNS [35,36].
At the peripheral terminals, noxious mechanical, thermal, or chemical stimuli activate specialized mechanosensitive ion channels, such as TRPA1 and ASICs [37,38,39,40]. This activation enhances cation permeability, triggering an inward current that produces a depolarizing generator potential. As a non-propagating analog signal, the amplitude of this generator potential is proportional to the stimulus intensity; once it reaches a critical threshold, it facilitates the transition from an analog signal to “all-or-none” digital action potentials for long-distance propagation.
The initiation of pain signaling begins with the generation of a non-propagating receptor potential at the nociceptor terminal. Once this graded potential reaches a critical threshold in response to noxious stimuli, it triggers an action potential—a rapid electrochemical event characterized by a depolarization phase, mediated by Na^+^ influx through voltage-gated sodium (Nav) channels, followed by a repolarization phase driven by K^+^ efflux via voltage-gated potassium (Kv) channels [35,36]. Nociceptive neurons express a combination of tetrodotoxin-sensitive (TTX-S) and tetrodotoxin-resistant (TTX-R) Nav channels; while Aδ fibers utilize both subtypes, C fibers predominantly express the TTX-R variant [41]. Crucially, the amplitude of the initial generator potential is proportional to the stimulus intensity, which in turn modulates the firing frequency of the subsequent action potentials. These signals propagate actively along the axon to the central terminals, where the arrival of the impulse activates voltage-gated calcium (Cav) channels. The resulting Ca^2+^ influx into the presynaptic terminal facilitates the exocytosis of excitatory neurotransmitters, primarily glutamate, into the synaptic cleft. These neurotransmitters bind to postsynaptic ionotropic glutamate receptors, generating an excitatory postsynaptic potential (EPSP). The amplitude of the EPSP, which is directly determined by the quantity of released neurotransmitter, dictates the firing frequency of the second-order neuron. This frequency-coded information is then transmitted to higher CNS centers for the comprehensive processing and encoding of pain intensity [32,35].
5. Modulation of Nociceptive Transmission by AST
The therapeutic potential of AST in modulating nociceptive transmission is supported by both its biochemical properties and recent electrophysiological evidence. Previous in vitro investigations have demonstrated that AST dose-dependently inhibits glutamate release from rat cortical synaptosomes by suppressing presynaptic N-type Cav channels and the mitogen-activated protein kinase (MAPK) signaling cascade [21]. Furthermore, AST has been shown to ameliorate neuropathic pain by antagonizing N-methyl-D-aspartate (NMDA) glutamate receptors [22]. Given its ability to cross the blood–brain barrier [42], systemic administration of AST likely modulates excitatory glutamatergic transmission within the central nervous system, including the trigeminal nociceptive pathways. Our recent findings further substantiate this hypothesis: (i) AST dose-dependently and reversibly inhibited the firing rates of SpVc WDR neurons in response to both noxious and non-noxious mechanical stimulation, whereas (ii) vehicle administration exerted no significant effect on spontaneous or evoked neuronal activity. Considering the established role of glutamate receptor antagonists in migraine management [43] and the NMDA-antagonistic effects of AST [22], it is plausible that AST may alleviate primary headache syndromes, such as migraine and cluster headache, potentially offering clinical efficacy comparable to the NMDA receptor blocker ketamine.
Collectively, these results provide compelling evidence that acute intravenous AST suppresses trigeminal sensory transmission—even in the absence of pre-existing inflammation or neuropathy—likely through the inhibition of Cav channels and excitatory glutamatergic signaling (Figure 3). Consequently, AST represents a promising CAM candidate for the management of trigeminal nociceptive pain, potentially offering a favorable safety profile without the side effects associated with conventional NMDA antagonists such as ketamine.
6. Modulation of Pathological Pain by AST
6.1. AST as an Intravenous Therapeutic: Targeting Inflammatory Pain Pathways
The efficacy of AST in mitigating inflammatory pain is closely linked to its modulation of ion channel expression and synaptic hyperexcitability. Previous studies have reported a significant up-regulation of N-type Cav channels in dorsal root ganglion and spinal dorsal horn neurons under carrageenan-induced acute inflammation [44], as well as selective up-regulation of NMDA-NR1 receptors in colitis models [45]. These findings suggest that the sensitization of both presynaptic Cav channels and postsynaptic NMDA receptors is a primary driver of inflammatory hyperalgesia.
Recent investigations by Chida and Takeda [25] specifically addressed whether intravenous AST could alleviate the inflammatory hyperexcitability of SpVc WDR neurons. In a Complete Freund’s Adjuvant (CFA)-induced inflammatory model, which was characterized by significantly reduced mechanical escape thresholds and localized edema, AST demonstrated a dose-dependent and reversible inhibition of SpVc WDR neuronal firing. Notably, the maximal suppressive effect on noxious mechanical stimuli occurred within 15 min and persisted for 45 min. A key finding of this study was that AST preferentially inhibited neuronal responses to noxious stimulation over non-noxious inputs, an effect not observed in vehicle-treated controls. This preferential inhibition suggests that AST may specifically target the mechanisms underlying central sensitization, potentially through the dual action of suppressing inflammation-induced up-regulation of Cav and NMDA channels, and potentially enhancing descending inhibitory systems, such as endogenous opioidergic pathways. Collectively, these observations provide robust evidence that intravenous AST effectively suppresses noxious synaptic transmission within the SpVc during inflammation, positioning it as a promising therapeutic candidate for trigeminal inflammatory pain with a superior safety profile compared to conventional treatments.
6.2. AST for Chronic Inflammatory Pain Relief: Modulation of Pro-Inflammatory Cascades
The clinical potential of AST in alleviating chronic inflammatory pain is rooted in its ability to suppress key pro-inflammatory mediators. Studies have demonstrated that AST concentration-dependently inhibits the production of prostaglandin E2 (PGE_2_) and tumor necrosis factor-alpha (TNFα) in both in vivo and in vitro models [46]. Furthermore, AST suppresses the expression of cyclooxygenase-2 (COX-2), the enzyme responsible for PGE_2_ synthesis, in chondrocytes and microglial cells [47,48]. PGE2 is a critical driver of pain hypersensitivity; it sensitizes mechanosensitive channels (e.g., TRPA1 and ASIC) and modulates Nav and Kv channels through protein kinase A (PKA) activation via prostanoid E (EP) receptors at nociceptive terminals [49]. Additionally, PGE_2_ reduces the activity of glycinergic inhibitory interneurons within the spinal dorsal horn, further exacerbating the pain state [50].
In our recent comparative study, the efficacy of systemic AST was evaluated against the selective COX-2 inhibitor celecoxib (CEL) in a CFA-induced orofacial inflammation model [24]. While inflamed rats exhibited marked mechanical hyperalgesia, treatment with AST, CEL, or a combined administration (1/2 CEL + 1/2 AST) successfully restored mechanical thresholds to baseline levels within two days. Electrophysiological recordings from SpVc WDR neurons further corroborated these behavioral results: all three treatment protocols significantly attenuated the heightened spontaneous activity, reduced the evoked firing rates in response to mechanical stimuli, and normalized the expanded receptive field sizes observed in inflamed subjects. These findings are consistent with prior evidence showing that other carotenoids, such as lutein, attenuate inflammatory hyperalgesia by suppressing COX-2 immunoreactivity in peripheral tissues [51]. Collectively, these observations suggest that AST mitigates mechanical hyperalgesia primarily by suppressing SpVc WDR neuronal hyperexcitability through the inhibition of the peripheral COX-2/ PGE_2_ cascade (Figure 4). When combined with its inhibitory effects on Cav channels and glutamate receptors, AST provides a multi-target approach to reducing nociceptive conduction, offering a robust foundation for developing novel analgesic strategies for orofacial pain with a potentially superior safety profile.
7. Functional Significance and Therapeutic Potential of AST
The increasing interest in CAM highlights the urgent need for effective alternatives when conventional Western pharmacotherapy proves inadequate. As detailed in this review, our in vivo neurophysiological research demonstrates that AST exerts potent analgesic and anti-inflammatory effects by targeting multiple molecular pathways. Specifically, AST exhibits: (i) an intravenous anesthetic-like effect on both nociceptive and inflammatory pain, comparable to established analgesics such as ketamine; and (ii) an anti-inflammatory efficacy equivalent to the selective COX-2 inhibitor celecoxib. These findings suggest that AST offers a promising clinical avenue for managing various pain states while potentially circumventing the adverse side effects associated with traditional drugs.
Furthermore, while this review primarily elucidated the sensory-discriminative aspects of pain (the lateral system), the potential impact of AST on the affective-motivational dimensions (the medial system) warrants further investigation. This is particularly relevant given the recent introduction of the term “nociplastic pain” by the International Association for the Study of Pain, describing pain arising from altered nociception despite no clear evidence of tissue or nerve damage [52,53]. Since nociplastic pain is heavily associated with central sensitization and emotional dysfunction, AST’s ability to reduce peripheral sensitization—a key driver of central hypersensitivity—positions it as a candidate for treating such complex conditions. Future research should prioritize the clinical translation of these findings, exploring AST’s role in drug discovery for functional foods and its broader applicability in alleviating the multifaceted pathology of nociplastic pain. Such advancements hold the potential to revolutionize pain management through the development of safer, non-pharmacological, or natural compound-based therapeutic modalities.
It should be noted that the previous studies cited in this review require further investigation into dose–response relationships and bioavailability. Additionally, given the current lack of human data, our conclusions remain speculative until further research provides more robust evidence.
8. Concluding Remark
Recent evidence highlighting the modulation of neuronal excitability—specifically the inhibition of Cav channels, glutamate receptors, and the COX-2 signaling cascade—positions AST as a compelling candidate for Complementary and Alternative Medicine (CAM). Our laboratory’s in vivo neurophysiological findings, as synthesized in this review, confirm that the carotenoid AST possesses a dual therapeutic profile: (i) an intravenous anesthetic-like effect on both nociceptive and inflammatory pain, with efficacy comparable to established analgesics such as ketamine; and (ii) a potent anti-inflammatory action against chronic pain, nearly equivalent to the selective COX-2 inhibitor celecoxib. Collectively, these data substantiate the role of natural compounds in the mitigation of acute and persistent pain, underscoring their significant potential for clinical integration as safe and effective analgesic agents. Future studies should focus on conducting robust clinical trials, refining administration protocols, and evaluating the optimal regulatory framework for AST, ranging from functional foods to pharmaceutical applications.”
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Rao J.K. Mihaliak K. Kroenke K. Bradley J. Tierney W.M. Weinberger M. Use of complementary therapies for arthritis among patients of rheumatologists Ann. Intern. Med.199913140941610.7326/0003-4819-131-6-199909210-0000310498556 · doi ↗ · pubmed ↗
- 2Konvicka J.J. Meyer T.A. Mc David A.J. Roberson C.R. Complementary/alternative medicine use among chronic pain clinic patients J. Perianesth. Nurs.200823172310.1016/j.jopan.2007.05.00318226781 · doi ↗ · pubmed ↗
- 3Rosenberg E.I. Genao I. Chen I. Mechaber A.J. Wood J.A. Faselis C.J. Kurz J. Menon M. O’Rorke J. Panda M. Complementary and alternative medicine use by primary care patients with chronic pain Pain Med.200891065107210.1111/j.1526-4637.2008.00477.x 18564996 · doi ↗ · pubmed ↗
- 4Ernst E. Complementary medicine Curr. Opin. Rheumatol.20031515115510.1097/00002281-200303000-0001212598804 · doi ↗ · pubmed ↗
- 5Fremont L. Biological effects of resveratrol Life Sci.20006666367310.1016/S 0024-3205(99)00410-510680575 · doi ↗ · pubmed ↗
- 6Pervaiz S. Resveratrol: From grapevines to mammalian biology FASEB J.2003171975198510.1096/fj.03-0168 rev 14597667 · doi ↗ · pubmed ↗
- 7Shir Y. Raja S.N. Weissman C.S. Campbell J.N. Seltzer Z. Consumption of soy diet before nerve injury preempts the development of neuropathic pain in rats Anesthesiology 2001951238124410.1097/00000542-200111000-0003111684996 · doi ↗ · pubmed ↗
- 8Tall J.M. Raja S.N. Dietary constituents as novel therapeutics for pain Clin. J. Pain 200420192610.1097/00002508-200401000-0000514668652 · doi ↗ · pubmed ↗