Food as Friend or Foe: A Decadal Narrative Review of Dietary Patterns as Determinants of Gastrointestinal Pathophysiology and Clinical Outcomes (2015–2025)
Lavinia Cristina Moleriu, Raluca Lupusoru, Ruxandra-Cristina Marin, Călin Muntean, Teodora Piroș, Daliborca Cristina Vlad, Andrei Luca Dumitrașcu, Victor Dumitrașcu

TL;DR
This review explores how different dietary patterns affect gastrointestinal health and disease over the past ten years.
Contribution
The paper provides a comprehensive narrative review linking dietary patterns to GI disorders and highlights the role of the gut microbiome and metabolites.
Findings
Western diets are linked to gut dysbiosis and inflammation, while Mediterranean diets are protective.
Dietary strategies like gluten exclusion and low-FODMAP diets are effective for specific GI conditions.
Short-chain fatty acids from the microbiome play a key role in regulating gut health.
Abstract
Diet is a major modifiable determinant of gastrointestinal (GI) health, influencing disease risk and progression through coordinated effects on the gut microbiome, immune regulation, epithelial barrier integrity, oxidative balance, and epigenetic mechanisms. This narrative review synthesizes epidemiological, mechanistic, and clinical evidence from the past decade to examine bidirectional relationships between dietary patterns and seven common GI disorders: celiac disease, irritable bowel syndrome (IBS), Crohn’s disease, ulcerative colitis, Helicobacter pylori-associated gastritis, peptic ulcer disease, and lactose intolerance. Western dietary patterns, characterized by high intake of ultra-processed foods and saturated fats and low fiber consumption, are consistently associated with microbial dysbiosis, impaired barrier function, and enhanced inflammatory signaling. In contrast,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
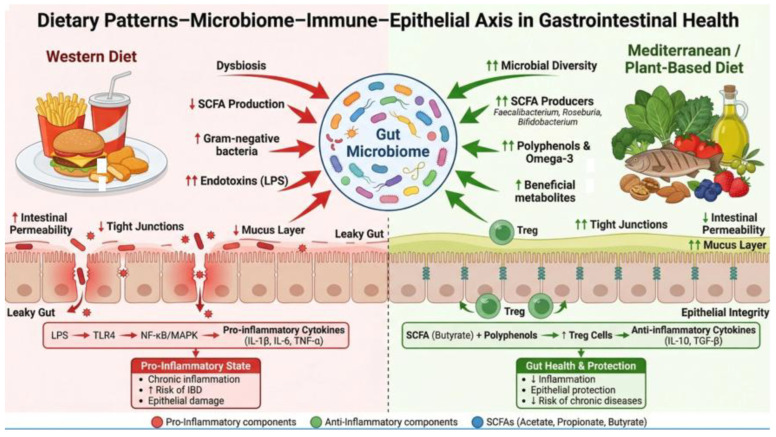 Figure 1
Figure 1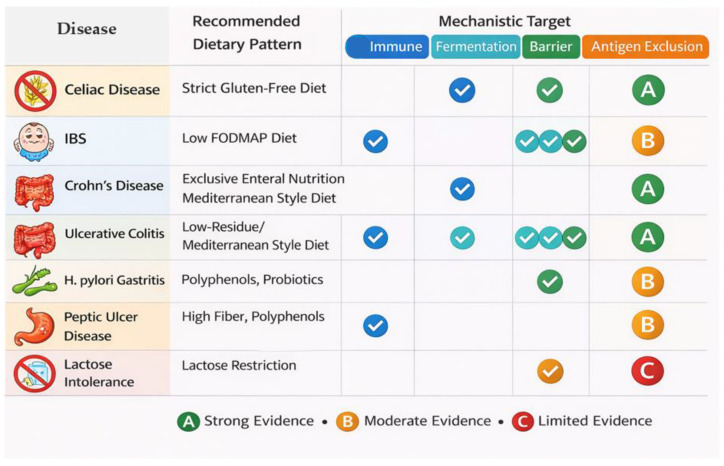 Figure 2
Figure 2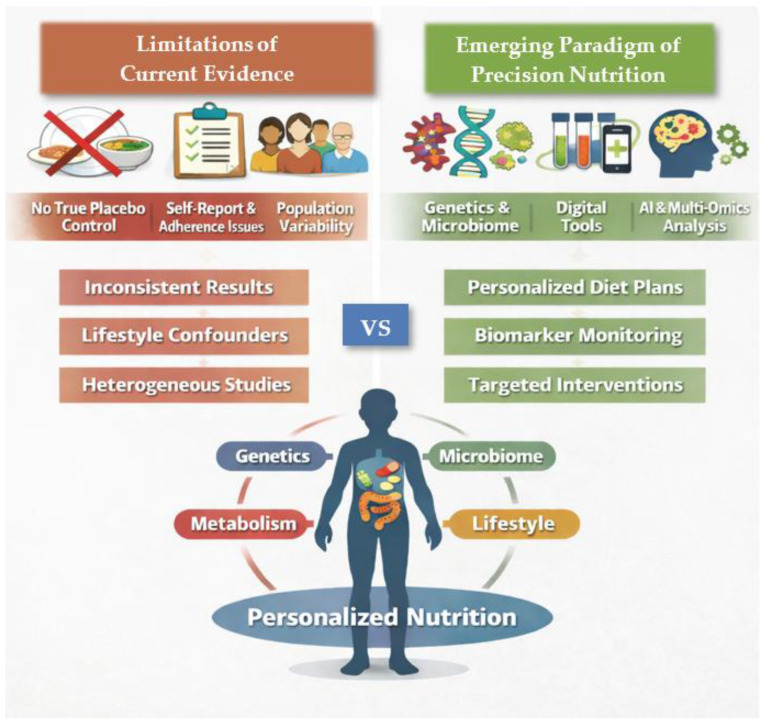 Figure 3
Figure 3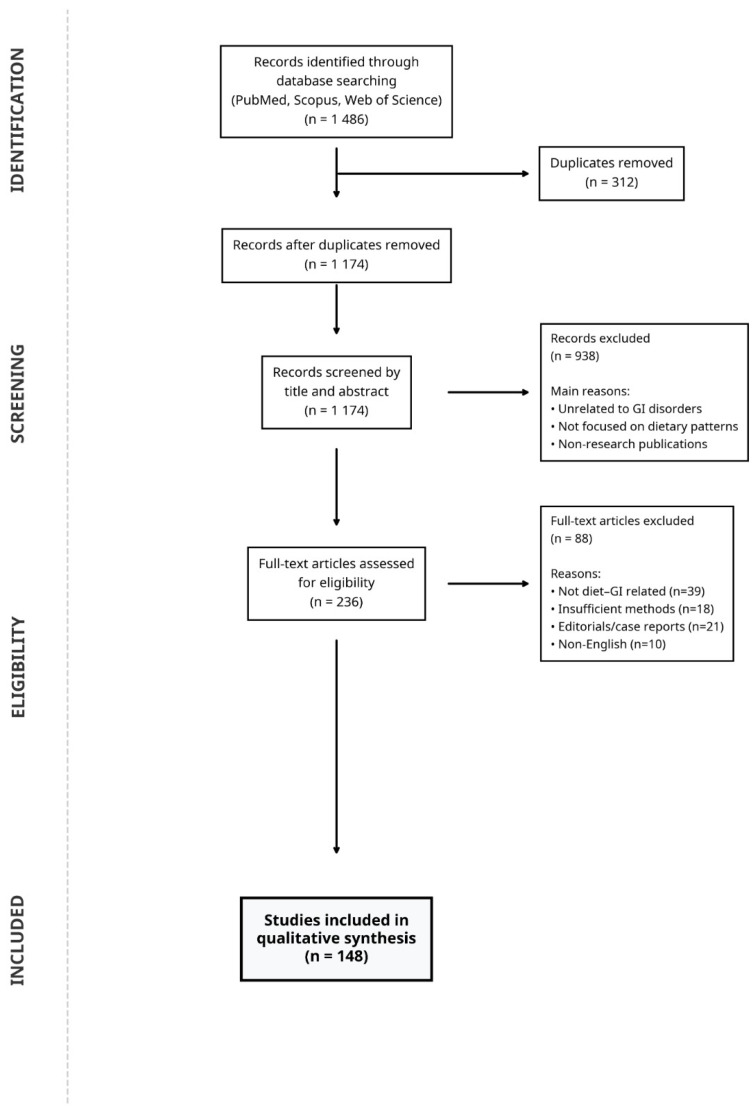 Figure 4
Figure 4- —“Victor Babes” University of Medicine and Pharmacy, Timisoara, Romania
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCeliac Disease Research and Management · Inflammatory Bowel Disease · Gastrointestinal motility and disorders
1. Introduction
Over the last decade, gastroenterology has moved from treating diet as a contextual lifestyle variable to recognizing it as a mechanistically grounded, clinically actionable exposure. This shift was enabled by converging evidence from multi-omics technologies, large prospective cohorts, and mechanistic studies demonstrating that dietary constituents can reshape epithelial signaling, immune regulation, and gut microbial ecology [1,2,3]. In this updated conceptual model, diet functions both as a potential pathogenic driver and as a therapeutic tool—“friend” and “foe”—depending on dietary pattern, host susceptibility, microbiome configuration, and co-exposures.
Global health assessments underscore the clinical relevance of this transition. The Global Burden of Disease Study 2019 identified dietary risks as a leading contributor to disability-adjusted life years (DALYs), with meaningful gastrointestinal (GI) manifestations within this burden [4,5,6]. Inflammatory bowel disease (IBD) has provided one of the most persuasive epidemiologic narratives supporting dietary involvement: while incidence has stabilized in some high-prevalence Western nations, newly industrialized regions (East Asia, South America, Middle East) have shown rapid increases that parallel dietary westernization and expanding ultra-processed food consumption [5,6]. Ecological correlations between indices of ultra-processed food exposure and IBD emergence have been reported, supporting diet as a plausible upstream determinant of risk at the population level [7,8,9,10,11,12,13,14].
Methodologically, nutritional gastroenterology has also matured. Retrospective self-report instruments (FFQs, 24 h recalls) remain widely used but have increasingly been complemented by objective biomarkers of intake (e.g., plasma phospholipid fatty acids, urinary polyphenol metabolites, fecal short-chain fatty acids (SCFA) measurements), mitigating bias and improving exposure specificity [15,16]. Large prospective cohorts have strengthened inference. The United Kingdom (UK) Biobank (over 500,000 participants) enabled robust analyses of dietary patterns and incident disease while adjusting for multiple confounders [17]. In that cohort, higher adherence to the Alternate Mediterranean Diet (AMED) was associated with reduced Crohn’s disease risk (HR 0.58; 95% CI 0.42–0.80), with partial mediation via anti-inflammatory biomarker pathways [18]. Similarly, a 2025 systematic review/meta-analysis (72 cohorts; >2 million participants) reported increased Crohn’s risk with inflammatory dietary patterns and ultra-processed foods, and protection associated with fiber intake and Mediterranean adherence, while ulcerative colitis associations showed greater heterogeneity [19].
These epidemiologic and methodological advances intersect with a major conceptual revolution: the microbiome. Work arising from the Human Microbiome Project and subsequent initiatives reframed diet–host interactions as frequently microbiota-mediated [20,21]. The gut microbiota acts as a metabolically active “virtual organ,” transforming dietary substrates into bioactive molecules that affect barrier function, immune programming, and systemic physiology [22]. SCFAs (acetate, propionate, butyrate) are central mediators: they fuel colonocytes, reinforce barrier integrity via tight junction regulation, shape epithelial proliferation via histone deacetylases (HDAC) inhibition, and exert anti-inflammatory effects through G protein-coupled receptors (GPCR) signaling (GPR41, GPR43, GPR109A) [23,24,25]. The observation that butyrate promotes Treg differentiation through epigenetic modification at the Foxp3 locus provided a mechanistic bridge linking fiber-rich patterns to immune tolerance [26,27]. Dietary pattern shifts can reshape microbial composition rapidly (within 24 h in controlled transition) supporting the feasibility of diet as a lever for microbiome-targeted interventions [28]. Mediterranean-type diets tend to enrich SCFA producers (e.g., Faecalibacterium prausnitzii, Roseburia spp., Bifidobacterium), whereas Western dietary profiles favor proteolytic and sulfate-reducing organisms linked to inflammatory metabolites such as hydrogen sulfide [29,30].
Finally, the decade also introduced precision nutrition as an attempt to move beyond “one-size-fits-all” dietary recommendations [31]. Evidence for strong inter-individual variation in metabolic responses to identical foods (e.g., postprandial glycemia) and the ability of machine learning models incorporating microbiome and lifestyle variables to improve prediction encouraged translation into gastrointestinal contexts [32,33,34]. Although consumer platforms now offer microbiome-informed diet guidance, limitations remain, including training bias toward Western populations, uncertain generalizability, privacy risks, and incomplete validation of clinically meaningful outcomes [35,36]. Nevertheless, early data suggest that baseline microbiome signatures may predict response to diets such as low-fermentable, oligosaccharides, disaccharides, monosaccharides and polyols (FODMAP) in irritable bowel syndrome (IBS), and that individualized fiber prescription strategies calibrated to microbiome capacity could become relevant for maintenance strategies in IBD remission [37,38].
Moreover, the relationship between diet and GI disease is bidirectional rather than unidirectional. GI disorders can themselves reshape dietary exposure by altering food tolerance, digestion, absorption, motility, and host–microbiome interactions. In celiac disease, villous atrophy impairs nutrient absorption and may secondarily modify microbial ecology; in IBS, visceral hypersensitivity and symptom anticipation frequently narrow dietary variety and promote avoidant eating patterns; and in inflammatory bowel disease, active inflammation, bile acid malabsorption, intestinal resections, and pharmacological therapies can alter nutrient handling and tolerance to fiber, fat, and fermentable substrates. Conversely, habitual dietary patterns can influence disease initiation and progression by modulating epithelial barrier integrity, immune activation, oxidative stress, bile acid metabolism, and microbiome-derived metabolites. Diet–microbiome interactions are increasingly recognized as central mediators of these processes, with dietary substrates shaping microbial composition and metabolic output, which in turn influences host immune and epithelial responses [39,40]. At the same time, host physiology and disease states can reciprocally modify microbiome structure and metabolic activity, reinforcing the dynamic and bidirectional nature of diet–host–microbiome interactions in gastrointestinal disease [41,42].
This narrative review synthesizes evidence from 2015–2025 on how five dietary paradigms (Western, Mediterranean, low-FODMAP, plant-based, gluten-free) shape gastrointestinal pathophysiology and outcomes across celiac disease, IBS, Crohn’s disease, gastritis, peptic ulcer disease, and food intolerances (gluten and lactose). Mechanistic emphasis is placed on epithelial signaling, inflammation, oxidative stress, regulated cell death/proliferation, motility, digestion/absorption, vascular patterns, and microbiota modulation, with attention to confounding by medications (proton pomp inhibitors, histamine H2 inhibitors, and biologic therapies).
2. Global Epidemiology, Dietary Risk Factors, and Economic Burden
Digestive diseases account for a substantial proportion of global DALYs, with peptic ulcer disease, IBD, and functional disorders contributing meaningfully [43,44]. Over the past decade, the burden of gastrointestinal disease has shown divergent trajectories: early-industrialized nations are increasingly characterized by stable incidence but rising prevalence of chronic conditions, while newly industrialized regions show accelerating incidence, reflecting the interplay of urbanization, healthcare access, diagnostic capacity, and lifestyle transition.
For IBD specifically, a four-stage model encompassing emergence, acceleration, compounding prevalence, and eventual prevalence equilibrium helps contextualize this divergence [45]. Many Western regions are now in the compounding prevalence stage, incidence stabilizes while prevalence rises because these are chronic, non-fatal diseases, with projections suggesting that more than 1% of populations in some regions may live with IBD within the next decade [46]. In contrast, regions with historically low disease burden (parts of Asia, South America, Middle East) are experiencing rapid incidence increases that align temporally with dietary westernization, including higher consumption of ultra-processed foods, saturated fats, and refined carbohydrates alongside decreased fiber intake [47,48,49]. Global burden of disease study (GBD) analyses document increasing crude prevalence globally (e.g., 1990 to 2019) with decreasing age-standardized prevalence, consistent with demographic effects and improved survival [50]. Strong regional heterogeneity persists, with the highest age-standardized prevalence in North America and Western Europe and lower rates in East Asia and Sub-Saharan Africa, while the steepest incidence increases are observed in transitioning economies [50,51,52,53]. Prospective evidence supports diet-pattern associations more consistently for Crohn’s disease than ulcerative colitis: inflammatory dietary patterns and ultra-processed foods increase Crohn’s risk, whereas fiber and Mediterranean adherence are protective. Ulcerative colitis(UC) associations are more variable [54].
Interpretation of dietary epidemiology requires careful attention to confounding, particularly acid-suppressive medications. Proton pump inhibitors (PPIs) and histamine-2 receptor antagonists (H2 blockers) are widely used and alter gut microbiota composition in ways that can mimic or modify diet-associated microbial signatures, including reduced beneficial taxa and increased susceptibility to enteric infections [55,56,57]. Adjustment for PPIs use is inconsistently performed, and sensitivity analyses (e.g., UK Biobank) suggest modest but meaningful attenuation of diet associations after accounting for acid suppression [58]. This supports a practical recommendation for future observational and interventional designs: systematically adjust/stratify by acid suppression and evaluate effect modification.
Diet is also central to highly prevalent disorders of gut–brain interaction such as IBS, where prevalence varies sharply depending on diagnostic criteria. Rome IV (2016) introduced stricter thresholds than Rome III, lowering estimated prevalence in meta-analyses from approximately 9% (Rome III) to 4% (Rome IV), while heterogeneity remains high across studies and methodologies [59,60,61]. Female sex and psychological comorbidities are consistent correlates [60,62]. A large proportion of IBS patients report food-triggered symptoms, and low-FODMAP dietary interventions show the strongest evidence base among dietary strategies, with clinically meaningful improvement in many adherent patients [63,64,65].
Celiac disease represents a distinct paradigm: a defined immune-mediated enteropathy triggered by gluten in genetically susceptible hosts (human leukocyte antigen (HLA)-DQ2/DQ8). Meta-analyses estimate global seroprevalence 1.4% and biopsy-confirmed prevalence 0.7%, with increased risk among first-degree relatives [66,67]. Incidence appears to be increasing beyond detection effects in some datasets, implicating additional environmental modifiers (microbiota, antibiotics, early-life exposures), while the gluten-free diet remains the cornerstone of management to prevent long-term complications [68,69].
Peptic ulcer disease has declined globally, consistent with improved H. pylori control and changes in nonsteroidal anti-inflammatory drug (NSAID) practice, but substantial geographical disparities persist, particularly in low- and middle-income countries where H. pylori prevalence is higher and diagnostic/therapeutic resources are limited [70,71].
Diet may influence mucosal defense and acid dynamics and may modulate H. pylori virulence and persistence [72]. Chronic gastritis remains highly prevalent worldwide and forms the substrate for progression along the Correa cascade toward gastric malignancy; high salt and preserved foods increase risk, while higher fruit/vegetable intake and Mediterranean patterns appear protective in the observational literature [73,74,75,76].
Food intolerances further contribute to symptom burden and clinical uncertainty. Non-celiac gluten sensitivity (NCGS) is defined by symptom induction after gluten exposure without celiac serology/histology, but true prevalence is likely lower than self-report when evaluated by double-blind placebo-controlled challenge [77,78]. Mechanisms may include innate immune activation, increased permeability, and sensitivity to non-gluten wheat components (fructans, amylase-trypsin inhibitors) [79,80]. Lactose intolerance affects a majority of adults globally with strong ethnic variation; symptoms are dose-dependent and modified by transit time, microbiota, and visceral sensitivity [81,82,83]. It frequently overlaps with IBS and complicates dietary management, especially because lactose restriction is embedded within the broader low-FODMAP approach [64,65,84]. The key epidemiologic characteristics and diet-related associations across major gastrointestinal conditions are summarized in Table 1.
Beyond clinical burden, GI diseases impose major economic costs. In the United States, IBD-related expenditures exceed $25 billion annually, driven largely by biologic therapies, hospitalization, and long-term disease management; similar economic analyses from Europe also demonstrate substantial direct and indirect healthcare costs associated with inflammatory bowel disease [85,86]. IBS, despite being a functional disorder, generates substantial direct costs and disproportionate productivity losses through presenteeism and absenteeism [62,87,88]. Celiac disease is associated with lower direct medical costs but meaningful patient-borne dietary costs because gluten-free products are considerably more expensive than standard alternatives [89]. Emerging health economic analyses suggest that structured dietary interventions (e.g., low-FODMAP guidance for IBS) may be cost-effective compared with standard care, and that preventive dietary strategies could reduce long-term disease burden; however, comprehensive and standardized economic evaluations remain limited [90,91].
3. Epigenetics and Signaling Networks Linking Diet to Gastrointestinal Pathophysiology
Diet modulates GI disease risk not only through exposure patterns but also through epigenetic programming and signaling pathway activation. Epigenetic mechanisms, including deoxyribonucleic acid (DNA) methylation, histone modifications, and non-coding ribonucleic acid (RNAs), provide a molecular interface between diet-derived inputs (and microbiota-derived metabolites) and durable gene expression states [92,93]. Epigenome-Wide Association (EWAS) studies have identified differentially methylated regions in IBD enriched in genes related to immune regulation and barrier integrity, consistent with a role for epigenetic dysregulation in sustaining inflammatory phenotypes [94,95,96]. Importantly, epigenetic marks are potentially reversible, distinguishing nutritional epigenetics from fixed genetic risk and offering a rationale for diet-based modulation.
DNA methylation depends on DNA methyltransferase (DNMT) enzymes and S-adenosyl-L-methionine (SAM) as the methyl donor, linking methylation capacity to one-carbon metabolism and dietary cofactors (folate, B12, B6, B2) [97,98,99]. In IBD, folate deficiency is common due to malabsorption, increased demands, and medication interactions (e.g., methotrexate, sulfasalazine), and has been associated with aberrant methylation patterns that may favor pro-inflammatory gene expression [100,101]. Microbial contributions may further shape methylation capacity, as certain commensals synthesize folate, supporting a microbiome–methylation axis responsive to diet and pre/probiotic strategies [102]. Candidate gene methylation changes have been reported (e.g., Interferon-gamma (IFNG) hypomethylation in UC; methylation shifts in protease-activated receptor (PAR2) and other inflammatory/barrier-related loci), and more recent work has linked methylation signatures to clinically relevant outcomes such as postoperative recurrence risk in Crohn’s disease [103,104,105,106,107].
Histone modifications provide a complementary and highly diet-responsive mechanism. Butyrate is the clearest mechanistic bridge connecting dietary fiber, microbiome activity, and chromatin remodeling: as a potent histone deacetylase (HDAC) inhibitor, it increases histone acetylation at loci governing barrier integrity (tight junction proteins, mucins) and regulatory immune programs, while suppressing inflammatory transcriptional networks including NF-κB-linked outputs [108,109,110,111,112,113]. A key therapeutic implication is the Foxp3/Treg axis: butyrate promotes Foxp3 expression through increased acetylation at promoter/enhancer regions and also acts indirectly through receptors such as G protein-coupled receptors (GPR109A) to support regulatory immune phenotypes (e.g., interleukin (IL)-10) [114,115,116,117]. Reduced colonic butyrate concentrations, depleted butyrate-producing taxa, and impaired regulatory immune function in IBD reinforce the biological plausibility of fiber-rich dietary interventions and butyrate-focused therapeutic strategies [118,119].
At the signaling level, nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and mitogen-activated protein kinase (MAPK) pathways serve as central hubs integrating dietary inputs, microbial sensing, and cytokine cascades. Key diet-responsive signaling pathways involved in gastrointestinal inflammation and their functional outputs are outlined in Table 2.
Western dietary components, especially saturated fatty acids and dietary patterns that promote endotoxemia, can activate toll-like receptor 4/toll-like receptor 2 (TLR4/TLR2) signaling and trigger myeloid differentiation primary response 88 (MyD88)-dependent cascades culminating in nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) nuclear translocation and inflammatory transcription (Tumor Necrosis Factor alpha (TNF-α), interleukin 1 beta (IL-1β), interleukin 6 (IL-6); cyclooxygenase-2 (COX-2); inducible nitric oxide synthase (iNOS)) [120,121,122,123,124,132,133,134]. Mitogen-activated protein kinase (MAPK) pathways (extracellular signal-regulated kinase (ERK), c-jun n-terminal kinase (JNK), p38 mitogen-activated protein kinase (p38)) interact extensively with nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), respond to oxidative stress, and amplify inflammation through transcriptional and post-transcriptional mechanisms including stabilization of cytokine mRNA [125,126,127]. In contrast, peroxisome proliferator-activated receptor gamma (PPAR-γ) functions as a diet-responsive anti-inflammatory nuclear receptor: activation by Mediterranean pattern components (oleic acid, omega-3 polyunsaturated fatty acids (ω-3 PUFA), polyphenols) supports transrepression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and broader immunomodulatory effects; reduced peroxisome proliferator-activated receptor gamma (PPAR-γ) expression in inflammatory bowel disease (IBD) mucosa and therapeutic effects of agonists in ulcerative colitis (UC) provide pathway validation [128,129,135,136,137].
Microbiota-derived metabolites extend epigenetic and immunologic modulation beyond butyrate. Propionate and acetate influence Treg differentiation and immune programming via receptor signaling and HDAC inhibition; polyphenols modulate DNMT activity, histone acetylation states, and miRNA networks (including links to PI3K/AKT/mTOR signaling), though translation to consistent clinical benefit remains variable across compounds and contexts [138,139,140,141,142,143,144]. The principal diet–microbiome–metabolite interactions and their epigenetic and immune consequences relevant to gastrointestinal inflammation are summarized in Table 3.
These layers interact with genetic susceptibility: genome-wide association study (GWAS) have identified >240 IBD loci, yet explained heritability remains limited, consistent with strong environmental contributions. Gene–diet interactions (e.g., nucleotide-binding oligomerization domain-containing protein 2 (NOD2)/autophagy pathways and fiber responsiveness; fucosyltransferase 2 (FUT2) secretor status and microbiota configuration) may contribute to heterogeneity in diet–disease relationships and provide a rationale for precision nutrition approaches [79,145,146,147]. Emerging causal inference strategies (including epigenetic Mendelian randomization) illustrate the potential to identify epigenetic mediators of disease risk, although the cited 2025 example pertains to pollution-related epigenetic marks rather than diet directly [148].
An additional biomolecular dimension through which dietary patterns influence GI disease involves modulation of epithelial junctional integrity. The intestinal barrier is maintained by coordinated interactions among tight junction proteins (including occludin, claudins, and zonula occludens proteins) and adherents junction components such as E-cadherin and β-catenin [149]. Western-style dietary patterns rich in saturated fats, cholesterol, and oxidized cholesterol derivatives (oxysterols) have been shown to impair epithelial barrier function by promoting oxidative stress, altering membrane lipid composition, and activating inflammatory signaling pathways such as NF-κB and MAPK. These processes disrupt tight junction architecture, increase intestinal permeability, and facilitate translocation of luminal microbial products that amplify mucosal inflammation [130,131].
In contrast, bioactive compounds abundant in plant-forward and Mediterranean dietary patterns appear to exert protective effects on epithelial barrier function. Polyphenols, including flavanols such as epicatechin, have been shown to enhance expression and localization of tight junction proteins, reduce oxidative stress, and attenuate pro-inflammatory signaling within intestinal epithelial cells [150,151]. Experimental and translational studies suggest that polyphenol-rich foods can stabilize junctional complexes and improve barrier integrity partly through antioxidant effects and modulation of microbiome-derived metabolites [152,153]. These findings support the concept that dietary composition directly influences epithelial permeability and mucosal resilience, providing a mechanistic link between dietary patterns and susceptibility to inflammatory gastrointestinal disorders.
Dietary patterns influence gastrointestinal health through interconnected effects on the gut microbiome, epithelial barrier integrity, immune signaling, and epigenetic regulation. These mechanisms do not operate in isolation but form an integrated network linking luminal dietary exposures to mucosal inflammation and disease susceptibility. The principal epigenetic and signaling pathways through which dietary patterns modulate gastrointestinal pathophysiology are summarized in Figure 1.
Oxidative stress is a convergent mechanism. Western dietary exposures promote reactive oxygen species (ROS) via mitochondrial dysfunction, nicotinamide adenine dinucleotide phosphate (NADPH) oxidase activation, and advanced glycation end-product formation (AGE) formation, activating redox-sensitive inflammatory pathways (NF-κB, MAPK) and inflammasome signaling [154]. Systemic inflammatory biomarkers that integrate nutritional and inflammatory status, such as composite indices based on C-reactive protein and albumin, have been explored as prognostic indicators in gastrointestinal diseases, highlighting the clinical relevance of systemic inflammatory burden [155]. Saturated fat oxidation can generate higher oxidative burden than unsaturated fats; depleted glutathione pools compromise antioxidant defenses, whereas Mediterranean patterns support redox resilience through antioxidant micronutrients and substrate availability for glutathione synthesis [156,157]. NLRP3 inflammasome activation reflects a key crossroads where metabolic stress can drive pathology (e.g., ceramide and mitochondrial ROS pathways), but epithelial inflammasome signaling can also support protective IL-18 responses under some conditions, emphasizing context-dependent effects [158,159].
4. Characterization of Dietary Patterns: Definitions, Components, and Biological Profiles
Dietary patterns relevant to GI health can be positioned along a continuum ranging from predominantly pro-inflammatory, barrier-disruptive exposures (typified by the Western dietary pattern) to anti-inflammatory and microbiome-supportive patterns (such as Mediterranean and plant-forward diets), alongside targeted therapeutic diets designed to address specific mechanisms, including the low-FODMAP diet for fermentative load reduction and the gluten-free diet for immune-mediated gluten intolerance. These patterns differ not only in macronutrient composition but also in degree of processing, additive exposure, fermentable substrate availability, and bioactive compound density. Together, these features influence microbial metabolism, epithelial signaling pathways, bile acid profiles, and mucosal immune tone. Figure 2 provides a conceptual overview linking major GI diseases with the dietary strategies most commonly applied to modify these biological pathways.
4.1. Western Dietary Pattern (Ultra-Processing, Additive Exposure, and Pro-Inflammatory Signaling)
The Western pattern is characterized by high intake of ultra-processed foods (UPFs), refined carbohydrates, saturated/trans fats, red/processed meats, sodium, and added sugars, with low intake of fiber-rich plant foods [160,161]. UPFs are operationalized via the NOVA classification, with Group 4 defined by industrial formulations containing uncommon ingredients in home cooking and additives designed to enhance palatability and shelf-life [162]. Epidemiologically, UPFs may contribute 50–60% of total energy intake in highly industrialized settings and are increasing globally [163], and recent syntheses connect this exposure to IBD risk via microbiome disruption, permeability, and innate immune activation [164].
Biologically, Western dietary exposure promotes a characteristic pattern of dysbiosis marked by reduced microbial diversity and depletion of barrier-supportive, short-chain fatty acid (SCFA)-producing taxa (e.g., Akkermansia muciniphila, Faecalibacterium prausnitzii), alongside enrichment of pathobionts such as Bilophila wadsworthia, members of the Enterobacteriaceae family, and sulfate-reducing bacteria [165,166,167]. Functionally, these compositional shifts are associated with diminished SCFA generation, compromised mucus layer integrity, and increased epithelial permeability. Postprandial “metabolic endotoxemia” provides a key mechanistic link: high-fat meals can enhance lipopolysaccharide (LPS) translocation and elevate circulating endotoxin levels, activating TLR4–MyD88 signaling and downstream NF-κB-mediated inflammatory cascades, while low dietary fiber limits SCFA-mediated counter-regulatory effects, creating a combined pro-inflammatory milieu [168,169].
In addition to excess saturated fats and refined carbohydrates, Western dietary patterns are also characterized by exposure to cholesterol oxidation products (oxysterols), which can arise during industrial food processing, prolonged storage, and high-temperature cooking of cholesterol-rich foods. These oxidized cholesterol derivatives possess strong pro-oxidant and pro-inflammatory biological activity and have increasingly been implicated in mechanisms linking Western dietary exposures with intestinal inflammation and epithelial barrier dysfunction [170,171,172].
Experimental studies indicate that oxysterols can induce oxidative stress and activate inflammatory signaling pathways such as NF-κB in intestinal epithelial cells, thereby contributing to disruption of epithelial barrier integrity and amplification of mucosal inflammatory responses. In parallel, Western-type diets rich in saturated fats have been shown to impair intestinal barrier function and increase epithelial permeability, further promoting inflammatory signaling in the gastrointestinal mucosa [149,173].
Recent integrative analyses further highlight how Western-style dietary patterns interact with gut barrier function, microbial metabolism, and host immune signaling to sustain chronic low-grade inflammation relevant to gastrointestinal and systemic disease [174].
Additives commonly present in ultra-processed foods are increasingly recognized as biologically active co-exposures rather than inert ingredients. Experimental models suggest that certain emulsifiers (e.g., carboxymethylcellulose, polysorbate-80) can disrupt mucus architecture, promote bacterial encroachment, and trigger low-grade inflammation at exposure levels below traditional toxicity thresholds; although human data remain limited, these findings support a precautionary perspective, particularly in susceptible populations [175].
4.2. Mediterranean Dietary Pattern (Bioactive Density, SCFA Support, and Inflammation Resolution)
The Mediterranean diet (MD) is defined by high intake of fruits/vegetables/legumes/nuts/whole grains, olive oil as the main fat, moderate fish/poultry, low red/processed meat, and (in many definitions) moderate wine with meals [175,176]. The PREDIMED trial established systemic benefits and catalyzed mechanistic work on inflammation and microbial remodeling [177]. A 2024 synthesis reported consistent microbiome benefits with MD adherence, including enrichment of taxa linked to SCFA production and barrier function (e.g., Bifidobacterium, F. prausnitzii, Roseburia, A. muciniphila) [178]. Short interventions (e.g., 8 weeks) have been associated with increased microbiome richness and reductions in systemic inflammatory markers such as C-reactive protein (CRP) in some cohorts [179].
The MD’s anti-inflammatory profile is best understood as a synergy of bioactive exposures rather than a single nutrient effect. Extra-virgin olive oil (EVOO) provides oleic acid (linked to peroxisome proliferator-activated receptor (PPAR)-γ signaling) and polyphenols (oleuropein, hydroxytyrosol, oleocanthal) that exhibit antioxidant effects and NF-κB modulation [104]. Polyphenols from plant foods and wine undergo microbial biotransformation into metabolites (e.g., urolithins, phenolic acids) that can influence immune signaling and epithelial function [180]. High fermentable fiber availability supports SCFA production and butyrate-mediated HDAC inhibition and Treg-related immune regulation. In parallel, ω-3 polyunsaturated fatty acids (eicosapentaenoic acid/docosahexaenoic acid) (PUFAs (EPA/DHA)) support the generation of specialized pro-resolving mediators (resolvins/protectins/maresins), shifting inflammatory biology toward active resolution rather than only suppressing initiation [181].
In addition to its nutrient composition, the Mediterranean dietary pattern provides a diverse array of substrates that interact with the gut microbiome and host metabolic signaling pathways. Microbiome-mediated metabolism of dietary components can generate a broad spectrum of bioactive metabolites capable of influencing host physiology, including pathways linking microbial metabolism with neuroendocrine and immune regulation along the gut–brain axis [182]. Experimental studies further demonstrate that commensal bacteria can modulate host metabolic functions through the production of signaling molecules, including microbiota-derived metabolites that influence intestinal and systemic physiology [183]. Moreover, the gut microbiota contributes to host micronutrient availability through microbial biosynthesis of vitamins and other bioactive compounds, highlighting the integrative relationship between diet composition, microbial metabolism, and host nutritional status [184].
These biological features position the Mediterranean dietary pattern as a microbiome-supportive and anti-inflammatory nutritional framework relevant to gastrointestinal health. The combination of fermentable fiber, polyphenol-rich plant foods, and unsaturated fatty acids promotes microbial diversity, short-chain fatty acid production, and modulation of inflammatory signaling pathways within the intestinal mucosa. Through these mechanisms, Mediterranean-style dietary patterns may contribute to improved epithelial barrier function, immune regulation, and metabolic homeostasis within the gut ecosystem.
4.3. Low-FODMAP Diet (Fermentation/Osmotic Load Reduction with Microbiome Trade-Offs)
The low-FODMAP diet restricts poorly absorbed, rapidly fermented short-chain carbohydrates (fructose in excess, lactose, fructans, galacto-oligosaccharides (GOS), and polyols) and was developed to reduce luminal fermentation and osmotic load within the intestine [185]. Network meta-analysis and meta-analytic syntheses support it as the most evidence-based dietary approach for IBS, with response rates commonly reported between 50–80% in trials, and significant improvements across pain, bloating, flatulence, and bowel habit outcomes [186,187].
Biologically, symptom benefits reflect reduced luminal water (osmotic effect) and reduced fermentation-related gas generation, which is particularly meaningful in the context of visceral hypersensitivity [188]. However, low-FODMAP restriction also reduces prebiotic substrate availability and consistently decreases Bifidobacterium abundance and sometimes overall SCFA output, raising concerns about long-term implementation without reintroduction/personalization [189,190]. Current best practice remains the 3-phase model (elimination → systematic reintroduction → individualized maintenance), optionally supported by targeted low-FODMAP prebiotic strategies to preserve microbiome function.
A key development is the emergence of microbiome predictors of response. IBS microbiome subtypes (e.g., a “pathogenic enrichment” subtype) may experience greater symptom improvement and shifts toward a healthier composition during low-FODMAP implementation, supporting a future precision nutrition pathway, pending larger validation cohorts [191].
4.4. Plant-Based Dietary Patterns (Fiber-Centric Microbiome Support with IBD Practicality Constraints)
Plant-based dietary patterns span vegan to flexitarian approaches but share high fiber intake and high polyphenol/antioxidant exposure, with reduced saturated fat and reduced exposure to heme iron/heterocyclic amines from red/processed meat [192]. Cross-sectional and intervention data associate plant-predominant patterns with increased microbial diversity and enrichment of fiber-degrading/SCFA-producing taxa (e.g., Prevotella, Ruminococcus, Lachnospiraceae members), often within days of dietary change and stabilizing over weeks [193,194].
The biological relevance of plant-based dietary patterns largely reflects their high content of fermentable carbohydrates and resistant starch, which serve as key substrates for microbial metabolism in the colon. Fermentation of these compounds supports the production of short-chain fatty acids, particularly butyrate, acetate, and propionate, metabolites that contribute to epithelial energy supply, mucosal barrier maintenance, and immune regulation within the intestinal environment [195]. Experimental and multi-omics studies further demonstrate that diets enriched in resistant starch can substantially influence microbial gene expression, metabolic pathways, and host–microbiome metabolic interactions, underscoring the capacity of plant-derived carbohydrates to shape the functional activity of the gut microbiota [196]. From a nutritional perspective, however, strict plant-based dietary patterns require careful planning to ensure adequate intake of nutrients that may be less abundant or less bioavailable in plant sources, including vitamin B12, iron, zinc, calcium, and long-chain omega-3 fatty acids. Recent nutritional analyses further highlight that the health effects of plant-forward diets depend not only on macronutrient composition but also on micronutrient adequacy and overall dietary quality, emphasizing the importance of balanced plant-based dietary design in maintaining metabolic and gastrointestinal health.
4.5. Gluten-Free Diet (Essential Therapy for Celiac Disease; Nuanced Role in NCGS/IBS Overlap)
The gluten-free dietary pattern is characterized by the exclusion of gluten-containing cereals, primarily wheat, rye, and barley, and their derivatives. In practice, this pattern relies on naturally gluten-free foods such as rice, maize, legumes, fruits, vegetables, and alternative grains including buckwheat or quinoa, as well as commercially produced gluten-free products. Because gluten-containing cereals represent major sources of dietary fiber and fermentable substrates in many populations, their exclusion can substantially modify macronutrient composition and fermentable carbohydrate availability within the diet [197,198].
From a biological perspective, the relevance of gluten-containing cereals in gastrointestinal physiology relates not only to gluten peptides themselves but also to other wheat components such as fructans (a class of FODMAPs) and amylase–trypsin inhibitors (ATIs), which may influence immune activation and intestinal signaling pathways. These components can interact with epithelial and innate immune mechanisms, contributing to mucosal immune responses and barrier modulation in susceptible individuals [199,200].
The gluten-free dietary pattern can also influence gut microbiome composition through changes in fermentable substrate availability and fiber intake. Reduced consumption of whole-grain cereals may decrease exposure to prebiotic fibers that support beneficial microbial taxa, potentially altering microbial diversity and metabolic output. Conversely, dietary patterns emphasizing naturally gluten-free whole foods may maintain microbial diversity and metabolic activity through alternative fiber sources and plant-derived substrates [201,202].
These distinguishing features, biological signatures, microbiome effects, and clinical applications of the principal dietary patterns discussed are compared in Table 4.
4.6. Integrative Perspective
Across patterns, two pragmatic principles emerge: Mediterranean/plant-predominant patterns function as broadly protective “base diets” for many GI conditions; targeted therapeutic diets (low-FODMAP, GFD) should be applied with defined indications and structured reintroduction/personalization to reduce unnecessary long-term restriction. The following section examines how these dietary patterns influence the pathophysiology and clinical course of specific gastrointestinal disorders.
5. Pathophysiological Impact of Dietary Patterns Across GI Conditions
GI disorders influenced by dietary exposures can be broadly grouped into several pathophysiological categories, including immune-mediated enteropathy (celiac disease), disorders of gut–brain interaction (irritable bowel syndrome), inflammatory bowel diseases (Crohn’s disease and ulcerative colitis), upper gastrointestinal inflammatory conditions (gastritis, Helicobacter pylori infection, and peptic ulcer disease), and carbohydrate malabsorption syndromes such as lactose intolerance. Across these categories, dietary factors influence disease processes through multiple biological mechanisms, including antigen exposure (e.g., gluten), fermentation and osmotic load (FODMAPs and lactose), microbiome–metabolite interactions (such as short-chain fatty acids and bile acids), modulation of epithelial barrier integrity, and activation of inflammatory signaling pathways. At the same time, gastrointestinal disorders themselves can modify dietary tolerance, nutrient bioavailability, digestive physiology, and microbiome composition, reinforcing the bidirectional nature of diet–disease interactions increasingly recognized in microbiome-focused research [39,40].
5.1. Celiac Disease: Gluten Elimination as Causal Therapy (Plus Monitoring and “Next-Generation Adjuncts”)
Celiac disease remains the clearest example of a diet-dependent immune disorder: gluten peptides trigger adaptive HLA-restricted responses and innate IL-15-driven epithelial injury, producing villous atrophy and malabsorption [203,204]. Despite strict dietary therapy, mucosal healing is heterogeneous; pooled analyses report 34% recovery at 2 years and 66% at 5 years, and a substantial minority experience persistent symptoms and/or mucosal inflammation often due to inadvertent exposure [205,206]. Serology can normalize before histology, and gluten immunogenic peptides in stool/urine are emerging adherence biomarkers with very high performance for detecting low-dose exposures that may sustain injury [207]. Nutritionally, the gluten-free diet (GFD) requires planned fiber and micronutrient replacement due to frequent deficits in iron, folate/B12, vitamin D, calcium, and zinc, and microbiome shifts (e.g., reduced Bifidobacterium) that may reflect reduced whole-grain/prebiotic intake unless actively addressed [208]. Adjunct therapies (enzymes, barrier modulators, tissue transglutaminase 2 (TG2) inhibitors such as ZED1227/TAK-227) are promising but not yet standard; thus, strict lifelong GFD remains foundational [209].
5.2. IBS: Symptom-Targeted Dietary Sequencing (Low-FODMAP as Escalation)
IBS reflects a multifactorial disorder involving gut–brain axis dysregulation, visceral hypersensitivity, altered motility, low-grade immune activation, and microbiome perturbations, providing a strong biological rationale for dietary intervention while also explaining the marked interindividual variability in response [210]. Among available strategies, the low-FODMAP diet has the most robust randomized and meta-analytic support; network meta-analyses and umbrella reviews consistently demonstrate clinically meaningful improvements in global symptoms as well as key domains such as abdominal pain and bloating [198,199]. However, because fermentable carbohydrate restriction can reduce microbial diversity and beneficial taxa, its use is recommended within a structured three-phase model (restriction, reintroduction, personalization) to minimize unnecessary long-term exclusion [211,212].
Emerging approaches are exploring microbiome-informed personalization strategies designed to retain dietary diversity while maintaining symptom control, though current evidence remains preliminary and requires cautious clinical translation [213]. Recent work further emphasizes that IBS dietary management should be individualized, integrating symptom phenotyping, microbiome considerations, and patient tolerance to optimize outcomes rather than relying on uniform restriction [214].
In practice, management typically begins with general dietary advice, progresses to a low-FODMAP intervention in non-responders, and then transitions toward personalized modification; wheat or gluten restriction may benefit selected patients, although fructans rather than gluten are often the principal symptom drivers [214,215].
5.3. Crohn’s Disease and UC: Mediterranean/Whole-Food Patterns as Foundation, Induction Diets for Specific Contexts
In Crohn’s disease, diet associations are consistent and mechanistically aligned with dysbiosis and barrier dysfunction. Mediterranean-style diets are increasingly supported as an adjunctive foundation: prospective data in newly diagnosed patients link adherence to improved inflammatory markers and reduced dysbiosis, with multi-omic shifts (e.g., bile acids and metabolites) supporting biological plausibility [216]. Emerging clinical and observational studies further indicate that adherence to Mediterranean-style dietary patterns may influence intestinal inflammation, microbiota composition, and metabolic parameters in patients with inflammatory bowel disease [217]. Dietary intervention in Crohn’s disease dietary inflammatory index for Crohn’s disease (DINE-CD) suggests symptom remission rates comparable to more restrictive protocols, with better long-term feasibility [218]. Pediatric and subclinical inflammation datasets (e.g., fecal calprotectin associations) reinforce that dietary quality may matter even under biologic therapy [219]. For induction, exclusive enteral nutrition remains first-line in pediatric disease due to strong remission and mucosal healing outcomes, while Crohn’s disease exclusion diet (CDED) offers a pragmatic whole-food approach excluding emulsifiers and specific additives, often with partial enteral nutrition support [220,221]. Clinically, fiber restriction should be individualized and reserved for strictures or active flare intolerance rather than applied universally.
In UC, colonic localization increases relevance of luminal metabolites (notably butyrate). Mediterranean-style interventions have demonstrated reductions in inflammation with microbiome shifts in randomized controlled trial (RCT) settings [222,223] with supportive observational signals in pouch-related contexts [224]. Practical UC guidance typically involves fiber modulation by phase: lower insoluble fiber during flares and active symptoms, but restoration during remission to support SCFA production and microbial diversity. Specific components (e.g., limiting red/processed meat and additive-heavy UPFs; supporting ω-3 intake) remain mechanistically plausible but variably supported in clinical trials [225,226].
5.4. Gastritis/H. pylori and Peptic Ulcer Disease: Diet as Adjunct, Not Replacement
For H. pylori-associated gastritis, eradication therapy remains primary, but dietary adjuncts (polyphenols such as extra virgin olive oil (EVOO) components, sulforaphane, cranberry-derived compounds) show in vitro activity and limited/modest human effects; they may be reasonable adjuncts but not substitutes, particularly in high resistance settings or after treatment failure [227,228,229,230]. In gastric health more broadly, salt and preserved foods associate with cancer risk and may exacerbate mucosal injury and H. pylori virulence, whereas fruit/vegetable intake and Mediterranean-like patterns are generally protective [231]. Peptic ulcer disease (PUD) management remains anchored to H. pylori eradication and NSAID strategy; dietary measures are supportive (symptom control, mucosal protection) with limited RCT-grade evidence for specific foods, though fiber and polyphenol-rich patterns are plausible supportive frameworks [232,233,234].
5.5. Lactose Intolerance: Threshold-Based Management with Nutrient Protection
Lactose intolerance is best managed via threshold identification rather than blanket avoidance. Symptoms depend on dose, residual lactase activity, transit time, microbiome fermentation profile, and visceral sensitivity; many tolerate 12–15 g lactose when consumed with meals [235,236]. Practical tools include choosing low-lactose foods (hard cheeses), yogurt with live cultures, lactose-free milk, lactase supplements, and gradual reintroduction for possible colonic adaptation (evidence variable) [237]. Because dairy restriction risks calcium/vitamin D inadequacy, counseling on fortified alternatives and dietary calcium sources is essential [238]. A comparative overview of the dietary strategies, underlying targets, and practical implementation considerations across major gastrointestinal conditions is presented in Table 5.
6. Probiotics, Prebiotics, Synbiotics, and Postbiotics: Biotic Adjuncts to Dietary Intervention
Because dietary patterns reshape microbiome structure and function, biotic strategies can be positioned as adjuncts that support microbial resilience during dietary therapy (e.g., low-FODMAP), target specific outcomes (e.g., UC remission maintenance), or deliver defined microbial functions (e.g., postbiotic butyrate delivery).
6.1. Definitions and Molecular Routes (The International Scientific Association for Probiotics and Prebiotics (ISAPP) Framework)
ISAPP defines probiotics as live microorganisms conferring benefit when administered in adequate amounts; prebiotics as selectively utilized substrates; synbiotics as combinations (synergistic or complementary); and postbiotics as preparations of inanimate microorganisms and/or components that confer benefit [239,240]. Clinically, the most important principle is specificity: probiotic effects are strain-specific, prebiotic effects are substrate- and microbiome-dependent, and postbiotic effects are component- and delivery-dependent.
Across categories, shared mechanistic routes include: barrier reinforcement (tight junction/mucin support), immune modulation (increasing IL-10 and regulatory responses, reducing TNF-α/IL-6), ecological effects (competitive exclusion, bacteriocin production), and functional metabolite support (especially SCFAs such as butyrate) [241,242].
6.2. Probiotics: Evidence Patterns by Condition (IBS, UC, and Crohn’s Disease)
IBS has the broadest probiotic evidence base but also substantial heterogeneity. Meta-analytic syntheses suggest modest benefits for global symptoms and pain, with variable certainty depending on strain; adverse events are generally not increased [90,243,244]. Practically, strain selection should be conservative and evidence-led; the literature most often highlights Bifidobacterium and Lactobacillus strains, with certain named strains used commonly in practice (e.g., B. infantis 35624, L. plantarum 299v, B. coagulans MTCC5260), while recognizing that replication and standardization remain limitations [245]. The most clinically useful positioning is: probiotics as a time-limited trial (e.g., 4–8 weeks) integrated with diet, not as standalone therapy.
In IBD, evidence diverges sharply between UC and Crohn’s disease. Updated syntheses support benefit in UC for remission induction/maintenance in some contexts, with VSL#3/Visbiome and E. coli Nissle 1917 often cited as higher-evidence options, and strong data for pouchitis prevention/treatment [246,247]. In Crohn’s disease, trials are largely negative for induction or maintenance, with only limited signals in select pediatric settings [248]. This UC–CD divergence is clinically important and should be stated explicitly to prevent overgeneralization.
6.3. Prebiotics and Synbiotics: Function Depends on Fermentation “Fit”
Prebiotic substrates, including inulin, fructo-oligosaccharides (FOS), galacto-oligosaccharides (GOS), lactulose, as well as resistant starch and polydextrose, are best conceptualized as modulatory tools aimed at enhancing short-chain fatty acid (SCFA) production and supporting the growth of beneficial microbial taxa. However, their physiological impact is highly context-dependent, varying according to baseline microbiome composition, metabolic functionality, and dose tolerability [192,249,250,251]. Emerging clinical and translational evidence further indicates that even structurally related fructan prebiotics are not interchangeable: differences in polymer chain length and fermentability (e.g., longer-chain inulin versus shorter-chain FOS) can lead to distinct metabolic and functional outcomes, including divergent effects on host metabolic endpoints and microbial biosynthetic pathways such as folate metabolism [192]. These findings reinforce the view that the term “prebiotic” does not represent a single, uniform exposure but rather a heterogeneous class of substrates with differential ecological and metabolic effects that must be matched to host–microbiome characteristics [250].
In GI disorders, symptomatic limits often constrain dosing, and clinical outcome data remain mixed; network analyses do not consistently show prebiotics outperforming probiotics in IBS [252,253].
Synbiotics aim to pair organisms with their preferred substrates and may add value where either component alone is insufficient. Some trials (including combinations incorporating microencapsulated butyrate + probiotics + scFOS) suggest symptom improvement in IBS, but optimization of pairing, ratios, and delivery remains a research frontier [254,255,256].
6.4. Postbiotics: Stability and Safety Advantages, Delivery Challenges
Postbiotics are appealing due to stability and safety (no live translocation risk), but they require effective delivery of the active component. Butyrate is the prototypical postbiotic with HDAC inhibition, GPCR signaling, barrier support, NF-κB modulation, and Treg promotion; oral delivery requires encapsulation/esterification strategies to reach the colon [257,258,259,260]. Cell wall components (MDP, LTA) can modulate innate immunity through NOD2/TLR-related pathways and show experimental anti-inflammatory or metabolic effects, though translation to routine GI practice remains early [261,262]. The principal categories of biotic interventions, their definitions, clinical contexts, and practical considerations are summarized in Table 6.
6.5. Practical Integration with Dietary Therapy and Safety
Biotics should generally be positioned as adjuncts: for example, low-FODMAP diet can reduce Bifidobacterium, supporting a rationale for targeted probiotic co-administration and structured reintroduction of prebiotic foods [100]. In UC, evidence-based probiotic formulations may be combined with 5-ASA in remission maintenance. In celiac disease, microbiome shifts on GFD provide a rationale for emphasizing naturally gluten-free prebiotic-rich foods and considering probiotics selectively, while always prioritizing strict gluten exclusion.
Safety is favorable in immunocompetent individuals, but caution is warranted in severely immunocompromised patients, those with central lines, and other high-risk contexts; postbiotics may offer theoretical advantages where viability-related risk is unacceptable [263].
7. Limitations of Current Evidence and the Emerging Paradigm of Precision Nutrition
Although the preceding sections demonstrate consistent associations between dietary patterns and gastrointestinal health, important methodological and conceptual limitations constrain causal inference and clinical translation. Dietary interventions differ fundamentally from pharmacological trials: diets are complex, multi-component exposures embedded within food matrices, precluding true placebo control and blinding. Adherence is difficult to verify objectively and often relies on self-report, while effect sizes tend to be modest and highly context dependent. These features contribute to substantial heterogeneity across studies and limit the proportion of nutrition recommendations supported by high-level randomized evidence.
In gastroenterology, these challenges are amplified by disease-specific factors. In irritable bowel syndrome, inconsistent stratification by subtype and heterogeneous outcome measures limit comparability across trials. In IBD, dietary effects are difficult to disentangle from relapsing disease courses and concomitant pharmacotherapy. In CD, reliance on self-reported gluten avoidance frequently overestimates true adherence, while objective biomarkers remain underused. Control group selection further complicates interpretation, as habitual diets introduce lifestyle confounding and structured comparator diets may exert unintended biological effects.
Beyond these methodological constraints, the most fundamental limitation of population-based dietary guidance is the marked interindividual variability in response to identical foods. Large-scale metabolic studies demonstrate wide variation in postprandial glycemic and lipemic responses, even among genetically identical individuals, with genetic factors explaining only a modest proportion of this variability. Gut microbiome composition consistently emerges as a key determinant of individual responses, often accounting for more variability than macronutrient composition alone. These findings challenge the conceptual foundation of one-size-fits-all dietary recommendations.
The convergence of these limitations has driven the emergence of precision nutrition, an approach that seeks to individualize dietary advice using integrated biological data rather than population averages. Recent digital-health frameworks further illustrate how individualized nutrition strategies can be operationalized in clinical practice by integrating dietary intake data, metabolic biomarkers, and longitudinal patient monitoring to guide personalized dietary recommendations and follow-up [264]. Contemporary scholarship describes precision nutrition as a paradigm shift enabled by advances in multi-omics profiling, digital health technologies, and machine-learning analytics, which together allow the integration of genetic, metabolic, microbiome, and lifestyle data to generate individualized dietary recommendations and predict heterogeneous responses to foods and interventions [265].
Microbiome-informed algorithms have demonstrated superior predictive performance for postprandial metabolic responses compared with traditional nutrient-based approaches, and early gastroenterology trials suggest that personalized dietary strategies may achieve symptom control comparable to standardized therapeutic diets while preserving microbiome diversity. The interacting sources of methodological bias, biological variability, and emerging precision-nutrition solutions are schematically summarized in Figure 3.
Despite its promise, precision nutrition remains constrained by technical and practical barriers. Current models explain only a limited proportion of response variability, microbiome composition is temporally dynamic, and dietary intake assessment remains imperfect even with digital tools. Implementation requires infrastructure rarely available in routine care, raising concerns regarding cost, equity, data privacy, and algorithmic bias. Consequently, precision nutrition is unlikely to replace conventional dietary guidance in the near term.
Until this paradigm matures, clinical practice must integrate existing evidence pragmatically. Dietary patterns, rather than isolated nutrients, should remain the foundation of recommendations, with Mediterranean and plant-forward approaches offering the most consistent benefit across gastrointestinal conditions. Restrictive therapeutic diets should be implemented with professional supervision, structured reintroduction, and nutritional monitoring. Precision nutrition is likely to enter practice incrementally, particularly for conditions characterized by high response variability, serving as a complementary rather than disruptive evolution of dietary therapy.
8. Materials and Methods
This work was conducted as a narrative review integrating epidemiological, mechanistic, and clinical evidence on the role of dietary patterns in gastrointestinal (GI) health and disease. Although the review was not designed as a formal systematic review, a structured literature search and screening approach was applied to improve transparency and reproducibility. A narrative framework was deliberately chosen to allow conceptual synthesis across heterogeneous study designs, including prospective cohort studies, randomized controlled trials, experimental and translational mechanistic investigations, and microbiome-focused research. Given the complexity of dietary exposures and the multifactorial pathophysiology of gastrointestinal disorders, a purely systematic or meta-analytic framework was considered insufficient to capture the breadth of biological mechanisms, disease-specific nuances, and clinical contexts addressed in this review.
A comprehensive literature search was conducted using PubMed/MEDLINE, Scopus, and the Web of Science Core Collection. The search strategy combined controlled vocabulary terms and free-text keywords related to dietary patterns (including Mediterranean, Western, low-FODMAP, plant-based, and gluten-free diets), gastrointestinal disorders (celiac disease, irritable bowel syndrome, Crohn’s disease, ulcerative colitis, gastritis, peptic ulcer disease, and lactose intolerance), and mechanistic pathways linking diet to gastrointestinal physiology and pathology (gut microbiota, short-chain fatty acids, epithelial barrier function, immune signaling, epigenetic regulation, and precision nutrition). Searches focused primarily on articles published between January 2015 and May 2025, reflecting a period of rapid advances in microbiome science, nutritional epidemiology, and systems-based dietary research. Reference lists of key reviews, meta-analyses, and clinical guidelines were manually screened to identify additional relevant publications.
The literature search initially identified 1486 records across the three electronic databases. After removal of 312 duplicate records, 1174 articles remained for title and abstract screening. During this stage, 938 records were excluded because they did not address dietary patterns in relation to gastrointestinal disorders, focused on unrelated disease areas, or lacked sufficient methodological relevance to the objectives of this review.
The full texts of 236 articles were subsequently assessed for eligibility. Of these, 88 articles were excluded for the following reasons: lack of direct evaluation of diet–gastrointestinal relationships (n = 39), insufficient methodological description or unclear study design (n = 18), publication type such as editorials or case reports (n = 21), or language other than English (n = 10).
Ultimately, 148 publications met the eligibility criteria and were included in the qualitative synthesis. Additional references cited throughout the manuscript were included to provide mechanistic background, clinical context, and supporting epidemiological evidence beyond the studies captured through the structured screening process. The literature selection process is summarized in Figure 4.
Eligible studies included human observational studies, randomized controlled trials, systematic reviews and meta-analyses with clearly described methodologies, as well as experimental and translational studies providing insight into diet–microbiome–immune interactions relevant to gastrointestinal health. Articles were limited to those published in English. Case reports, editorials, and studies lacking sufficient methodological detail were excluded unless they provided unique mechanistic perspectives directly informing the conceptual framework of the review.
Data extraction focused on study design, population characteristics, dietary exposure definitions and assessment methods, gastrointestinal outcomes and diagnostic criteria, and reported molecular endpoints, including microbiome composition, microbial metabolites, inflammatory signaling pathways, and epigenetic markers. Where applicable, effect estimates such as relative risks, odds ratios, hazard ratios, and corresponding confidence intervals were considered. Rather than quantitative pooling, evidence was synthesized using a thematic and biological integration strategy, grouping findings according to dietary pattern, biological pathway, and disease phenotype. Priority was given to consistency across independent cohorts, convergence between mechanistic and clinical data, and relevance to clinical decision-making.
The overall strength of evidence supporting dietary interventions across gastrointestinal conditions was assessed qualitatively, taking into account study design hierarchy, sample size, duration of follow-up, risk of bias, and consistency of findings across populations. The appraisal reflects consideration of the totality of available evidence for each condition rather than reliance on individual studies. Particular attention was paid to potential confounders relevant to gastroenterology, including concomitant pharmacological therapies, disease activity status, and baseline nutritional status.
9. Conclusions and Future Directions
This narrative review integrates epidemiological trends, mechanistic evidence, and clinical studies to clarify the role of dietary patterns in gastrointestinal health. Collectively, the data support the view that diet is a biologically active determinant of gastrointestinal physiology and disease, acting through interconnected mechanisms involving the gut microbiome, immune regulation, epithelial barrier function, oxidative balance, and epigenetic modulation. These pathways link dietary exposures to both short-term functional responses and long-term disease trajectories.
Across GI conditions, dietary patterns, rather than isolated nutrients, emerge as the most clinically meaningful unit of intervention. In this context, the Mediterranean dietary pattern consistently represents a sustainable anti-inflammatory framework, associated with favorable microbiome profiles and improved outcomes across inflammatory bowel disease, functional gastrointestinal disorders, and gastric pathology. Conversely, Western dietary patterns characterized by high intake of ultra-processed foods and low fiber are repeatedly linked to dysbiosis, barrier dysfunction, and heightened inflammatory risk. To translate these concepts into practical guidance, Table 7 summarizes the overall strength of evidence and corresponding clinical recommendations across major GI conditions.
Strict gluten avoidance remains indispensable for celiac disease, while the low-FODMAP diet provides validated symptom control in irritable bowel syndrome when applied with structured reintroduction to preserve microbiome health. In Crohn’s disease, exclusive enteral nutrition and structured exclusion diets are effective for remission induction, whereas long-term management favors whole-food, minimally processed dietary patterns. Adjunctive biotic interventions may enhance dietary strategies when applied selectively and supported by strain-specific evidence, particularly in ulcerative colitis, pouchitis, and irritable bowel syndrome, while postbiotic approaches represent a promising translational extension of mechanistic insights.
From a clinical perspective, these findings underscore the need to integrate dietary assessment and counseling into routine gastroenterological care with rigor comparable to pharmacological management. Nutritional adequacy must be actively monitored whenever dietary restriction is imposed, and collaboration with specialized gastrointestinal dietitians should be considered standard practice for complex dietary interventions. Probiotic use should remain targeted, indication-specific, and time-limited, rather than empirical.
Looking forward, future research should prioritize standardized outcome measures, longer-duration intervention studies, and direct comparisons between dietary patterns. Mechanistic investigations aimed at identifying microbiome-derived mediators of dietary effects will support the development of targeted postbiotic therapies, while advances in nutrigenetics and systems biology may enable more precise patient stratification. Precision nutrition offers a compelling trajectory toward individualized dietary care, particularly for conditions characterized by marked interindividual variability, but its implementation must address issues of feasibility, equity, and interpretability.
In conclusion, diet has evolved from ancillary lifestyle advice to a mechanistically grounded therapeutic component of gastroenterology. Treating dietary intervention with the same scientific rigor as pharmacotherapy, while recognizing its cultural, social, and personal dimensions, represents both an evidence-based obligation and an opportunity to improve long-term gastrointestinal health.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wang Y. Huang Y. Chase R.C. Li T. Ramai D. Li S. Huang X. Antwi S.O. Keaveny A.P. Pang M. Global Burden of Digestive Diseases: A Systematic Analysis of the Global Burden of Diseases Study, 1990 to 2019 Gastroenterology 2023165773783.e 1510.1053/j.gastro.2023.05.05037302558 · doi ↗ · pubmed ↗
- 2Abbafati C. Abbas K.M. Abbasi M. Abbasifard M. Abbasi-Kangevari M. Abbastabar H. Abd-Allah F. Abdelalim A. Abdollahi M. Abdollahpour I. Global Burden of 369 Diseases and Injuries in 204 Countries and Territories, 1990-2019: A Systematic Analysis for the Global Burden of Disease Study 2019 Lancet 20203961204122210.1016/S 0140-6736(20)30925-933069326 PMC 7567026 · doi ↗ · pubmed ↗
- 3Peery A.F. Crockett S.D. Murphy C.C. Jensen E.T. Kim H.P. Egberg M.D. Lund J.L. Moon A.M. Pate V. Barnes E.L. Burden and Cost of Gastrointestinal, Liver, and Pancreatic Diseases in the United States: Update 2021 Gastroenterology 202216262164410.1053/j.gastro.2021.10.01734678215 PMC 10756322 · doi ↗ · pubmed ↗
- 4Alatab S. Sepanlou S.G. Ikuta K. Vahedi H. Bisignano C. Safiri S. Sadeghi A. Nixon M.R. Abdoli A. Abolhassani H. The Global, Regional, and National Burden of Inflammatory Bowel Disease in 195 Countries and Territories, 1990-2017: A Systematic Analysis for the Global Burden of Disease Study 2017 Lancet Gastroenterol. Hepatol.20205173010.1016/S 2468-1253(19)30333-431648971 PMC 7026709 · doi ↗ · pubmed ↗
- 5Wang R. Li Z. Liu S. Zhang D. Global, Regional, and National Burden of 10 Digestive Diseases in 204 Countries and Territories from 1990 to 2019 Front. Public Health 202311106145310.3389/fpubh.2023.106145337056655 PMC 10088561 · doi ↗ · pubmed ↗
- 6Ross F.C. Patangia D. Grimaud G. Lavelle A. Dempsey E.M. Ross R.P. Stanton C. The Interplay between Diet and the Gut Microbiome: Implications for Health and Disease Nat. Rev. Microbiol.20242267168610.1038/s 41579-024-01068-439009882 · doi ↗ · pubmed ↗
- 7Perler B.K. Friedman E.S. Wu G.D. The Role of the Gut Microbiota in the Relationship Between Diet and Human Health Annu. Rev. Physiol.20238544946810.1146/annurev-physiol-031522-09205436375468 · doi ↗ · pubmed ↗
- 8David L.A. Maurice C.F. Carmody R.N. Gootenberg D.B. Button J.E. Wolfe B.E. Ling A.V. Devlin A.S. Varma Y. Fischbach M.A. Diet Rapidly and Reproducibly Alters the Human Gut Microbiome Nature 201450555956310.1038/nature 1282024336217 PMC 3957428 · doi ↗ · pubmed ↗