Origins of Avian Hyperactive Mitochondria, Genome Compaction, and Air-Sac Physiology in Early Theropods During the Carnian Pluvial Episode
Takumi Satoh

TL;DR
The paper suggests that early dinosaurs had bird-like traits, such as efficient mitochondria and compact genomes, which helped them thrive in harsh Late Triassic environments.
Contribution
The study proposes a link between avian-like physiology in early theropods and adaptation to the Carnian Pluvial Episode's harsh climate.
Findings
Early dinosaurs had hyperactive mitochondria and compact genomes, similar to modern birds.
These traits likely enhanced metabolic efficiency and locomotor performance in harsh environments.
The air-sac system and small cell size may have contributed to ecological dominance during the CPE.
Abstract
Extant birds and the earliest dinosaurs may share fundamental metabolic features essential for aerobic exercise, suggesting that the extraordinary physical performance typical of avian species originated when dinosaurs first appeared during the Carnian Pluvial Episode (CPE). This physiological adaptation is complemented by hyperactive mitochondria that exhibit high oxygen consumption and low reactive oxygen species production. Molecular genomics of fossils, the so-called “Jurassic Genome,” indicates that these early dinosaurs possessed compact genomes, 50–60% the size of the human genome, and small cells, implying a highly stringent metabolic regime. We suggest that hyperactive mitochondria, closely associated with compact genomes and small cells, drive theropod adaptation to the hot, dry, and hypoxic environments of the Late Triassic period, ultimately enabling their ecological…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
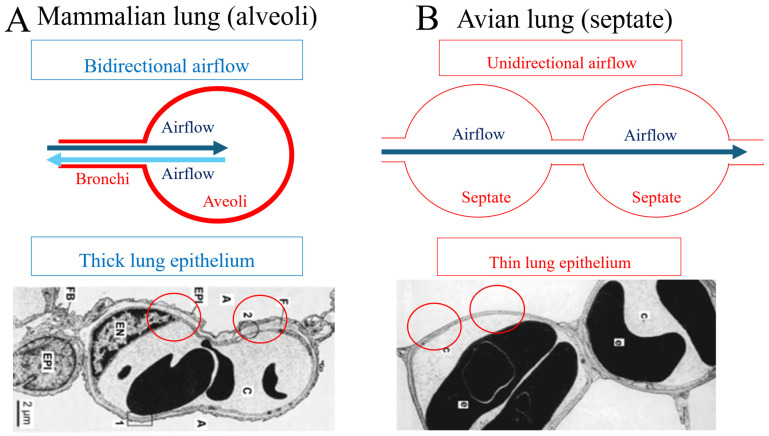 Figure 1
Figure 1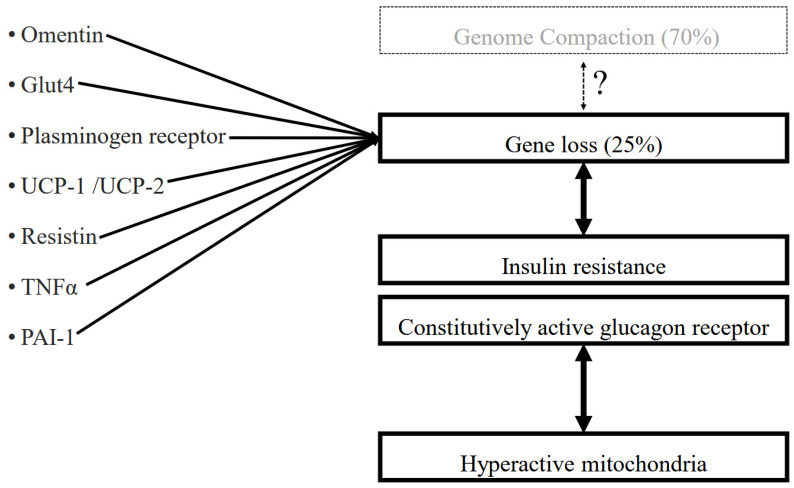 Figure 2
Figure 2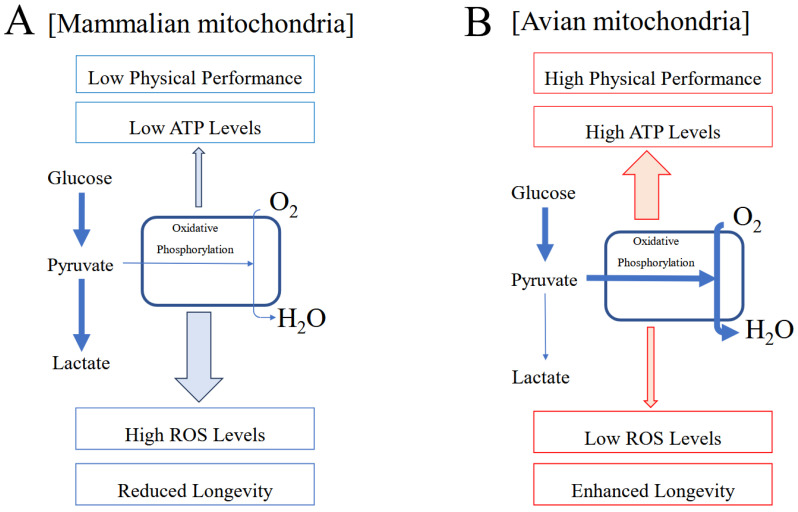 Figure 3
Figure 3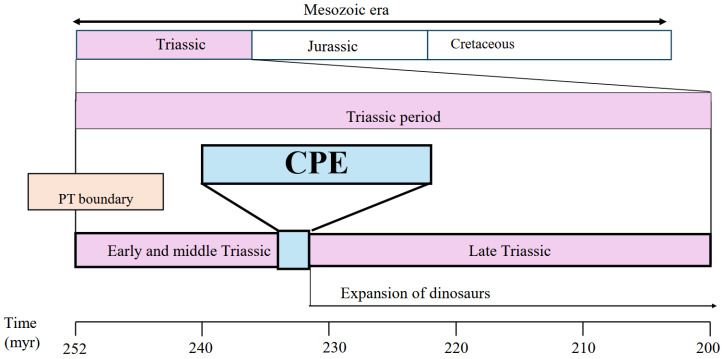 Figure 4
Figure 4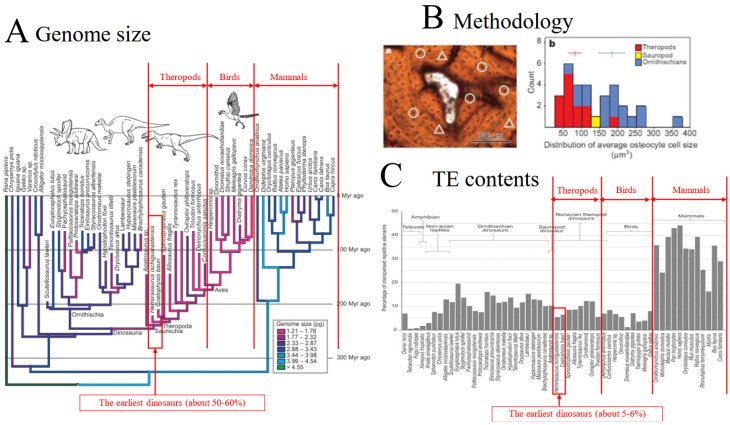 Figure 5
Figure 5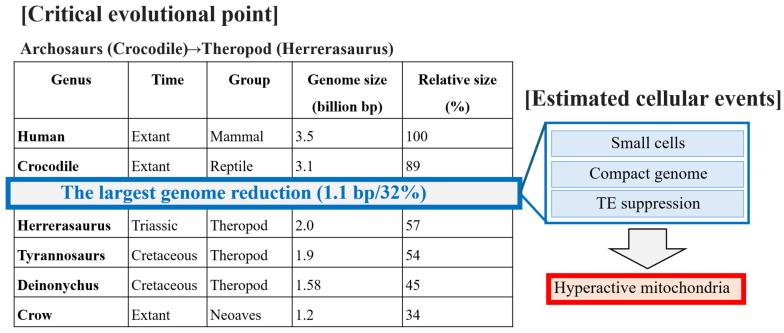 Figure 6
Figure 6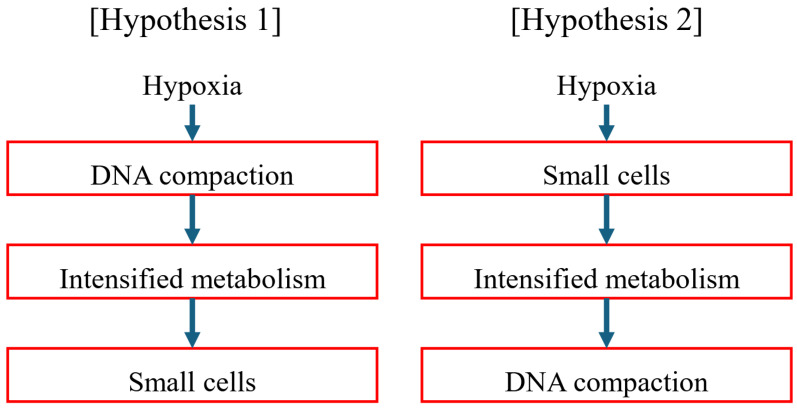 Figure 7
Figure 7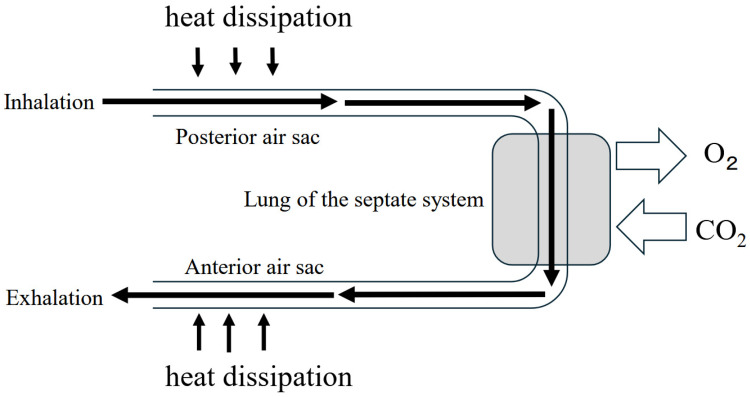 Figure 8
Figure 8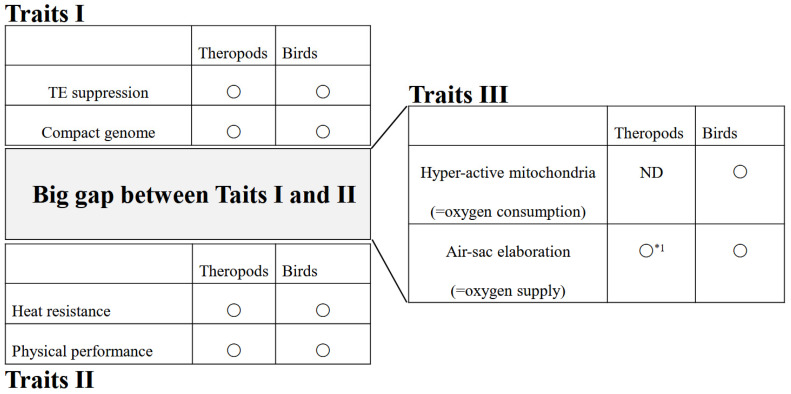 Figure 9
Figure 9Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPaleontology and Evolutionary Biology · Evolution and Paleontology Studies · Paleontology and Stratigraphy of Fossils
1. Introduction
Given that direct evidence of mitochondrial metabolism is not preserved in the fossil record, the timing and mechanisms by which avian-type hyperactive mitochondria and associated high physical performance were established in the theropod–bird lineage remain unclear. This review addresses these questions by proposing a working hypothesis integrating avian physiology, cell biology, and ecology. Birds exhibit this feature because their bodies are uniquely adapted for sustained flight and endurance, with a respiratory system optimized to support exceptionally high aerobic activity, even at extreme altitudes [1,2]. Such mitochondrial activity may be maintained by a unique balance in the insulin–glucagon axis (IGA). Birds possess constitutively active glucagon receptors and exhibit marked insulin resistance, maintaining mitochondria in an intrinsically active state [1,2]. We propose that these traits were inherited from the earliest dinosaurs, such as Herrerasaurus and Coelophysis, during the Triassic Period. Notably, the earliest dinosaurs emerged during the hot and humid conditions of the Carnian Pluvial Episode (CPE). They exhibited athletic bone structures, a primitive air-sac system, and fully developed bipedal locomotion, enabling strenuous activity even under low-oxygen conditions. These physiological and anatomical characteristics are best explained by evolutionary adaptations in aerobic metabolism, including hyperactive mitochondria and an efficient lung–air sac gas exchange system. Under the harsh environment of the CPE, theropod mitochondria may have been exceptionally active [3,4]. We suggest that hyperactive mitochondria, closely associated with compact genomes and small cells, drove theropod adaptation to the hot, dry, and hypoxic environments of the Late Triassic period, ultimately enabling the later evolution of powered flight during the oxygen-enriched conditions of the late Jurassic [3,4].
2. Avian Physical Performance
Among terrestrial animals, birds are considered “extreme athletes,” exhibiting exceptional endurance, speed, and strength. For example, bar-headed geese can fly over the Himalayas at altitudes where oxygen availability is extremely low. This feat is achieved through a highly efficient gas exchange system, enhanced oxygen transport, and remarkable aerobic capacity, which synergistically enable sustained vigorous exercise in hypoxic environments [5,6,7,8,9,10]. Birds possess a flow-through lung system with air sacs that enables continuous oxygen exchange, in contrast to the tidal breathing pattern observed in mammals. This respiratory architecture confers superior gas-exchange efficiency and supports prolonged activity even under oxygen-poor conditions [11,12,13,14,15,16,17].
Additionally, avian muscles and other tissues exhibit enhanced oxygen diffusion capacity, enabling rapid oxygen delivery to working muscles. This adaptation supports both long-distance migration and short bursts of intensely flight [11,12,13]. Birds also maintain elevated circulating levels of glucose and ketone bodies, ensuring a stable and flexible energy supply. Consequently, their mitochondria operate at a high metabolic rate fueled by dual energy substrates, supporting endurance while minimizing oxidative stress. This metabolic strategy ensures the continuous availability of carbohydrates and lipids as fuel sources [14,15,16,17].
3. Avian and Mammalian Lungs
As mentioned earlier, the most fundamental feature of the avian gas exchange system, which constitutes them as “extreme athletes,” is that air flows unidirectionally through the lungs and air sacs, enabling high-efficiency oxygen uptake even under hypoxic conditions [14,15,16]. Electron microscopy reveals that the avian lung epithelium is markedly thinner than that of mammals [17,18,19]. Birds possess septate lungs, a structural specialization permitting extreme thinning of the pulmonary epithelium. This adaptation significantly enhances gas-exchange efficiency by facilitating the rapid diffusion of oxygen and carbon dioxide across the respiratory interface. John B. West identified epithelial thinning as the key mechanism underlying this difference [17,18,19] (Figure 1).
4. Insulin–Glucagon Axis (IGA)
The IGA provides a critical framework for understanding the hormonal regulation of avian physical activity, functioning differently from that in mammals. Whereas mammalian metabolism tightly relies on insulin-mediated regulation of blood glucose and ketone bodies, birds have evolved a glucagon-dominated system that supports a hyperactive lifestyle and enhanced physical performance [20,21,22] (Figure 2). This mechanism favors circulating glucose availability over glycogen storage, ensuring immediate energy supply to meet elevated metabolic demands [23,24,25]. Birds also maintain elevated circulating levels of glucose and ketone bodies, providing a stable and flexible energy supply. Consequently, their mitochondria operate at high metabolic rates fueled by dual energy substrates, ensuring continuous availability of carbohydrates and lipids as energy sources [26,27,28]. Furthermore, birds exhibit markedly elevated circulating glucagon levels compared with mammals [26,27,28] (Table 1).
The striking feature of extant birds are significant insulin resistance [29,30,31]. The avian genome contains approximately 15,000 protein-coding genes, whereas mammalian genomes typically contain >20,000 genes [32,33,34]. Approximately 25% of genes have been lost in extant birds. Several genes essential for insulin sensitivity have been lost in birds [35,36,37,38]. It is suggested that this gene loss contributes to insulin resistance. In birds [1], the insulin signaling pathway appears markedly attenuated in adipose tissue and skeletal muscle, as indicated by reduced insulin receptor phosphorylation, although insulin is retained in the liver. Insulin resistance in adipose tissue and skeletal muscle of chickens may activate mitochondrial metabolism and increase oxygen consumption [39,40,41,42,43,44]. Insulin is essential for embryonic development in both mammals and birds [45,46]. Mammals retain insulin sensitivity throughout life death, whereas birds become insulin-resistant early in life [47].
Given that birds possess a constitutively active glucagon receptor, glucagon signaling remains persistently engaged, thereby driving sustained hepatic glucose release and chronically elevated blood glucose levels. Glucagon promotes energy mobilization by stimulating gluconeogenesis and mitochondrial respiration while suppressing cell growth and proliferation. It also activates the peroxisome proliferator-activated receptor-γ coactivator-1α, a central regulator of mitochondrial biogenesis, thereby enhancing aerobic capacity [23,24,25,26,27,28,29,30,31,32,33,34,35].
5. Avian and Mammalian Mitochondria
Hyperactive avian mitochondria consume large amounts of oxygen while producing minimal levels of reactive oxygen species (ROS). They actively eliminate ROS, thereby limiting oxidative damage, slowing aging-related processes, and supporting longevity. Avian cells are densely populated with mitochondria, enabling large-scale energy production and ROS elimination. During prolonged activity, birds steadily increase oxygen consumption while maintaining extremely low ROS release levels [48,49,50] (Table 2).
Conversely, increased oxygen utilization during intense exercise in mammals enhances ROS production. Therefore, mammalian mitochondria can be characterized primarily as ROS-generating systems. During sustained high-intensity activity, mammalian mitochondria contribute to oxidative stress, thereby accelerating aging [48,49,50] (Figure 3).
6. Theropod–Bird Lineage and the Triassic Environment
The PT boundary: Fluctuations in atmospheric oxygen concentrations were among the fundamental drivers of vertebrate evolution. Particularly, the sharp decline in oxygen levels at the PT boundary approximately 252 million years ago (myr) likely represented a severe physiological challenge faced by vertebrates. These hypoxic conditions exerted strong selective pressure on theropods, favoring smaller genomes to maximize oxygen utilization efficiency. The PT boundary marked a profound shift in the body structure of the theropod–bird lineage, facilitating adaptation to persistently thin air. This reorganization likely included the evolution of more efficient gas exchange systems, enabling survival under hypoxia and permitting diversification and ecological dominance during the Triassic period [54,55,56,57,58,59,60,61,62].
Before CPE: The Earth experienced five major mass extinction events, the largest occurring at the PT boundary and closely associated with a dramatic drop in atmospheric oxygen [54,55,56,57,58,59,60,61,62]. This event led to the extinction of more than 95% of all species, mainly due to hypoxia in the early Triassic period [54,55,56,57,58,59,60,61,62]. The surviving vertebrate lineages were subjected to extreme selection pressure, requiring rapid physiological and anatomical adaptations to hypoxic conditions [63,64,65].
During CPE: The CPE (244–242 myr) represents a remarkable Late Triassic climatic episode characterized by intense global rainfall and elevated humidity that temporarily interrupted the prevailing hot and arid Triassic climate [66,67,68,69,70,71]. During the CPE, dinosaurs’ expansion should have been accompanied by a profound reorganization of body structures, including the establishment of obligate bipedal locomotion, the development of a primitive air-sac system, the compaction of genomes, and the reduction in cell size [1,2,3,4] (Figure 4).
After CPE: In the Late Triassic period, global climate conditions reverted to hot and dry states, resulting in significant ecological restructuring [66,67,68,69,70,71]. Fern-dominated vegetation declined under increasing aridity, whereas drought-resistant conifers persisted and eventually dominated terrestrial ecosystems. Many forests most likely developed open understory spaces beneath tall conifer canopies [72,73,74]. These open habitats favored large, fast-moving theropods, enabling them to capitalize on physiological and locomotor advantages and achieve ecological dominance. At approximately 231 myr, more evolved athletic theropods, such as Herrerasaurus, emerged and became the dominant terrestrial predators because of their physical performance [72,73,74].
7. Compact Genome of the Earliest Dinosaurs
The PT boundary caused a complete change in the basic vertebrate body plan, facilitating adaptation to hypoxia. This novel body plan may have increased survival probability under hypoxic conditions. Genome compaction is considered a highly effective driver of this transition. Under hypoxia, theropods may have totally changed their body plan by reducing genome size. Genome reduction was demonstrated by the Jurassic genome. The genome sizes of extinct dinosaurs can be inferred from fossilized cell dimensions, particularly osteocyte lacunae. Chris Organ demonstrated a progressive decrease in genome size along the avian lineage—from diapsids to small theropods and ultimately to Neoaves—suggesting that genome compaction played a central role in theropod and avian evolution [3,75,76,77,78]. One plausible mechanism underlying genome compaction is the suppression of TEs [79,80,81,82]. Given that TEs significantly contribute to the maintenance and expansion of the vertebrate genome size, reduced TE abundance promotes genome compaction [79,80,81,82]. TEs comprise 30–40% of mammalian genomes, approximately 10% of theropod genomes, and 2–3% of avian genomes, indicating a close association between progressive TE reduction and the evolution of compact genomes [3] (Figure 5).
8. Emergence of Hyperactive Mitochondria
Genome compaction may have begun across the PT boundary when theropod lineages diverged from reptiles (crocodiles) and may have recurred during evolution from theropods (Herrerasaurus, Tyrannosaurus, and Deinonychus) to Neoaves (crow). The positive feedback loop between small cells, compact genomes, and TE suppression driven by hypoxia may have lasted for approximately 20 million years, up to the onset of the CPE [54,55,56,57,58,59,60,61,62]. Notably, the earliest dinosaurs, which appeared just after the CPE [4], possessed genomes approximately 50% the size of the human genome [3], consistent with pronounced genome compaction [83,84,85,86,87]. These traits may have been closely linked to increased metabolic activity and may underline the development of hyperactive mitochondria [83,84,85,86,87]. Particularly, dinosaur expansion during the CPE increased from 5% to 90% [4]. Therefore, the estimated timing of compact genome emergence, TE suppression, and small-cell evolution may be concentrated within the relatively short CPE interval (234–232 myr). Additionally, just after the CPE, the first theropod, Herrerasaurus, which is supposed to possess hyperactive mitochondria, appeared (Figure 6).
Another proxy for mitochondria activation may allow for direct reconstruction of metabolic rates from fossils. In situ Raman and Fourier-transform infrared spectroscopy can quantify the in vivo accumulation of metabolic lipoxidation signals in modern and fossil amniote bones. Inferred ancestral states reveal that theropods had high metabolic rates. Because these signals originated from mitochondria, the suggest that theropods possessed more active mitochondria than other terrestrial animals [88]. However, given that this lipoxidation research focused on Jurassic and Cretaceous theropods, there is no direct evidence on the Triassic period [88]. Furthermore, the Jurassic genome provides evidence of compact genomes only from Herrerasaurus and Coelophysis among Triassic theropods [3]. Further research on the earliest theropods and evidence of hyperactive mitochondria is required.
9. How Did Hypoxia Lead to Avian Hyperactive Mitochondria?
A possible pathway for the emergence of hyperactive mitochondria may have been initiated by a positive feedback loop among compact genome [86,87,88,89], intensified metabolism [48,49,50], and small cells [83,84,85], ultimately leading to the emergence of hyperactive mitochondria [1,2]. How was this positive feedback loop initiated in the theropod lineage? The premise is that hypoxia must have been the most decisive evolutionary driver during the Triassic period. (Figure 7).
Genome DNA may harbor genes that suppress aerobic mitochondrial metabolism, possibly via insulin signaling molecules. In other words, cells harbor genes that regulate mitochondrial activity to prevent dysregulation [35,36,37,38]. If these genes were lost during genome compaction, as shown in Figure 2 [32,33,34], metabolism may have become intensified [1,2]. Increased energy substrates per cell volume would need to be taken up through the introduction of smaller cells [83,84,85], further intensifying metabolism [48,49,50].
Alternatively, in response to hypoxia, cells may increase oxygen and energy substrate uptake per cell volume through cell membrane by reducing cell size [83,84,85]. Increased oxygen uptake may intensify metabolism [1,2]. Cells may allocate available resources toward metabolism by reducing DNA content, ultimately leading to genome compaction [32,33,34].
These causal relationships may become circular and may collectively lead to the emergence of hyperactive mitochondria [1,2]. Hypothesis 2 may be more likely, as increased oxygen supply per volume could occur rapidly. In contrast, hypothesis 1 may require additional time to activate metabolism in response to sudden hypoxia.
10. CPE as the Final Boost for Dinosaurs’ Expansion
The final major expansion of dinosaurs was likely accelerated by the CPE (234–232 myr), a short but intense climatic disruption which occurred during the Triassic period. The earliest known theropod, Herrerasaurus, appeared thereafter (approximately 231 myr). By this interval, theropods had already adapted to hypoxic conditions and undergone substantial genome reduction, which played a pivotal role in reshaping body structure and enhancing physiological performance [64,65,66].
Geological and sedimentological evidence indicates widespread monsoon-like rainfall and regional flooding during the CPE. This event was likely triggered by extensive volcanic activity in the Wrangellia Large Igneous Province, releasing massive quantities of CO_2_ and other gases into the atmosphere. The resulting greenhouse warming intensified evaporation and strengthened global hydrological cycles [67,68,69,70,71].
Early theropods such as Herrerasaurus had emerged, and the number of dinosaurs rapidly increased to constitute more than 90% of terrestrial vertebrate fauna, as suggested by fossil evidence. This expansion was accompanied by a significant reorganization of body structures, including the establishment of obligate bipedal locomotion, the development of a primitive air-sac system, the compaction of genomes, and a reduction in cell size [72,73,74,75].
The Triassic period was characterized by low atmospheric oxygen levels and extreme heat. Sustained locomotion under such conditions requires highly efficient heat-dissipation mechanisms. In theropods and modern birds, air sacs extended into major skeletal elements and internal cavities, forming an extensive internal ventilation network that likely provided a crucial adaptive advantage during the intense Triassic climate [89,90,91,92,93,94].
Faster movement enhances the effectiveness of this internal cooling system. In theropods and birds, air sacs permeate much of the body, enabling direct airflow from the environment. During sustained high-speed locomotion, this system allows uniform body cooling at wind speeds comparable to movement velocity. This adaptation enabled theropods to maintain rapid locomotion under extreme heat without succumbing to heat stress, conferring exceptional thermal resilience [89,90,91].
The evolution of air sacs may therefore be of particular significance [89,90,91]. By extending the pulmonary structures into large, hollow bones, this system greatly enhanced ventilatory capacity and oxygen delivery. Heat stress during the hot and humid CPE conditions may have been more severe than during other Triassic intervals. High humidity substantially amplifies thermal stress in terrestrial animals. In theropods, the air-sac system may have functioned to enhance gas exchange and improve heat tolerance. This physiological adaptation may have contributed substantially to dinosaurs’ dominance over other terrestrial vertebrates during the CPE [92,93,94] (Figure 8).
During prolonged locomotion, air flows rapidly through the lungs, continuously supplying oxygen at elevated levels. Increased velocity may further accelerate internal airflow, thereby improving gas exchange and heat dissipation. These physiological advantages may, in turn, promote skeletal and muscular adaptations supporting high speed locomotion [1]. Such adaptations may have enabled theropods to sustain rapid locomotion under extreme heat without succumbing to heat stress, conferring an exceptional cooling system [92,93,94].
11. New Avenues and Limitations of the Present Study
This study opens two new conceptual avenues. First, it proposes that the hyperactive avian mitochondria originated in the earliest dinosaurs. Second, it advances the hypothesis that the air-sac system evolved to enhance gas exchange and confer heat-stress resistance. Together, these perspectives provide a unified physiological framework for understanding dinosaur and avian success under extreme environmental conditions.
This study is inherently interdisciplinary, integrating evidence from physiology, molecular biology, paleontology, and ecology. Consequently, it is limited by the absence of direct fossil evidence supporting physiological and molecular traits. Parameters such as mitochondrial activity, hormonal regulation, and cellular metabolism are not directly preserved in the fossil record. Therefore, the hypotheses presented here rely only on indirect inference and comparative biological reasoning rather than direct observation (Figure 9).
12. Conclusions
The extraordinary physical performance of birds originates from a unique mitochondrial phenotype, often referred to as “super mitochondria,” characterized by exceptionally high metabolic performance [1]. These mitochondria, which likely played a central role in the evolutionary transition from theropods to birds, are hypothesized to have emerged in organisms adapted to chronical hypoxic environments [1]. This evolutionary transformation was particularly pronounced during the Triassic period, when theropods diverged from their diapsid ancestors under atmospheric oxygen levels lower than those prevailing during most other intervals in Earth’s history. During this interval, theropods underwent extensive genomic reorganization and profound restructuring of their body plans. These changes coincided with the emergence of super mitochondria, enabling the development of exceptional locomotor capacity. During the CPE, the combined effects of hyperactive mitochondria and the air-sac system—functioning in both gas exchange and heat dissipation—likely conferred a decisive adaptive advantage. Together, these innovations contributed to the ecological dominance of dinosaurs and laid the physiological foundation for the later success of birds.
13. Methodological Framework
This study is inherently interdisciplinary, integrating evidence from physiology, molecular biology, paleontology, and ecology. It is limited by the absence of direct fossil evidence supporting physiological and molecular traits, as parameters such as mitochondrial activity, hormonal regulation, and cellular metabolism are not directly preserved in the fossil record. Consequently, the hypotheses presented in this study rely on indirect inference and comparative biological reasoning rather than direct observation. The methodological backbone is therefore implicit and distributed across sections, most clearly in: (i) comparative physiological analyses of avian versus mammalian lungs and mitochondria (Section 1, Section 2 and Section 3 [1,2]); (ii) indirect genomic inference using osteocyte lacuna size as a proxy for genome size (“Jurassic Genome”; Section 5 and Section 6), and (iii) integration of paleoclimate reconstructions of atmospheric oxygen and the Carnian Pluvial Event (Section 7). In Figure 5, genome size and interspersed repetitive element content were inferred using primary histological data, including osteocyte size distribution in extinct dinosaur species, combined with regression models calibrated using extant taxa [3]. In addition, the genomes of theropods and birds exhibit substantially reduced transposable elements (TE) content compared with mammals, consistent with progressive genome compaction during theropod and avian evolution [3]. Data of Figure 5 are derived from [3].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Satoh T. Bird evolution by insulin resistance Trends Endocrinol. Metab.20213280381310.1016/j.tem.2021.07.00734446347 · doi ↗ · pubmed ↗
- 2Hickey A.J. Jüllig M. Aitken J. Loomes K. Hauber M.E. Phillips A.R. Birds and longevity: Does flight driven aerobicity provide an oxidative sink?Ageing Res. Rev.20121124225310.1016/j.arr.2011.12.00222198369 · doi ↗ · pubmed ↗
- 3Organ C.L. Shedlock A.M. Meade A. Pagel M. Edwards S.V. Origin of avian genome size and structure in non-avian dinosaurs Nature 200744618018410.1038/nature 0562117344851 · doi ↗ · pubmed ↗
- 4Bernardi M. Gianolla P. Petti F.M. Mietto P. Benton M.J. Dinosaur diversification linked with the Carnian Pluvial Episode Nat. Commun.20189149910.1038/s 41467-018-03996-129662063 PMC 5902586 · doi ↗ · pubmed ↗
- 5Scott G.R. Elevated performance: The unique physiology of birds that fly at high altitudes J. Exp. Biol.20112142455246210.1242/jeb.05254821753038 · doi ↗ · pubmed ↗
- 6Barve S. Dhondt A.A. Mathur V.B. Cheviron Z.A. Life-history characteristics influence physiological strategies to cope with hypoxia in Himalayan birds Proc. Biol. Sci.20162832016220110.1098/rspb.2016.220127903874 PMC 5136601 · doi ↗ · pubmed ↗
- 7LaguëS.L. High-altitude champions: Birds that live and migrate at altitude J. Appl. Physiol.201712394295010.1152/japplphysiol.00110.201728839002 PMC 5668450 · doi ↗ · pubmed ↗
- 8Faraci F.M. Adaptations to hypoxia in birds: How to fly high Annu Rev Physiol.199153597010.1146/annurev.ph.53.030191.0004232042973 · doi ↗ · pubmed ↗