Uncovering a Medieval Pogrom: Genetic History of a Jewish Community in Catalonia (Spain)
Laura Pallarés-Viña, Daniel R. Cuesta-Aguirre, M. Rosa Campoy-Caballero, Núria Armentano, Anna Colet, Assumpció Malgosa, Cristina Santos

TL;DR
This study uses ancient DNA to uncover the genetic history of a Jewish community in medieval Catalonia, linking them to Jewish populations and confirming they were victims of a 1348 pogrom.
Contribution
The first genomic analysis of medieval Iberian Jews, revealing their genetic affinities and confirming historical attribution to a pogrom.
Findings
Genetic affinities with ancient and modern Jewish populations were identified through PCA and ADMIXTURE analyses.
The Roquetes population's genetic composition is modeled as a mix of Canaan individuals and Iberian non-Jewish medieval people.
Uniparental markers showed high diversity consistent with Jewish community patterns and background endogamy.
Abstract
Background/Objectives. The Black Death pandemic, combined with the antisemitic climate of 14th-century Europe, led to widespread violence against Jewish communities, including numerous pogroms such as the one in 1348 in Tàrrega (Catalonia, Spain). In the Roquetes necropolis of Tàrrega, six communal graves containing at least sixty-nine individuals, with signs of violence, were dated to the mid-14th century. Based on the hypothesis that Iberian medieval Jewish communities preserve genetic similarities to other ancient and modern Jewish communities, our study aims to provide genomic information on medieval Iberian communities, which to date have been unknown. Methods. We analyzed DNA from sixteen individuals from the Roquetes necropolis using Twist ancient DNA enrichment capture. Several paleogenomic analyses based on nuclear DNA and uniparental markers were conducted to determine their…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
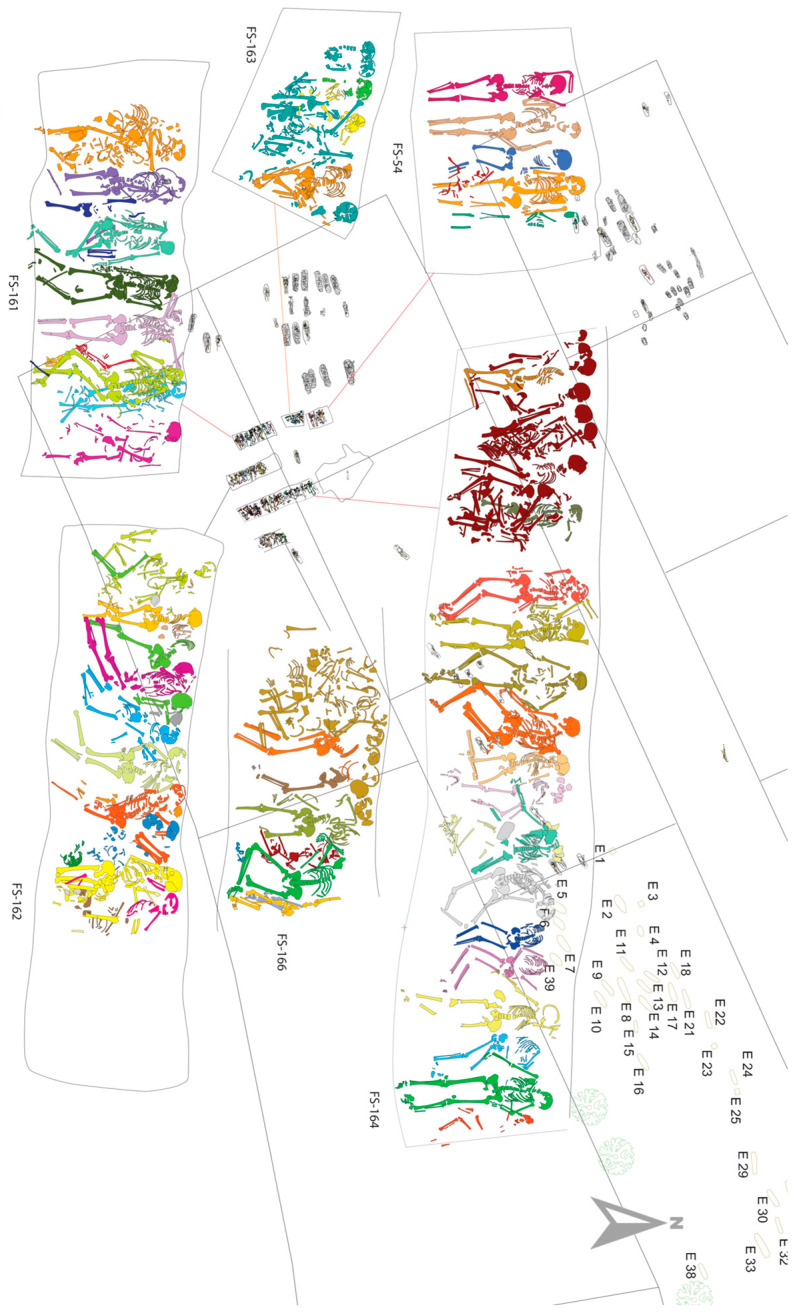 Figure 1
Figure 1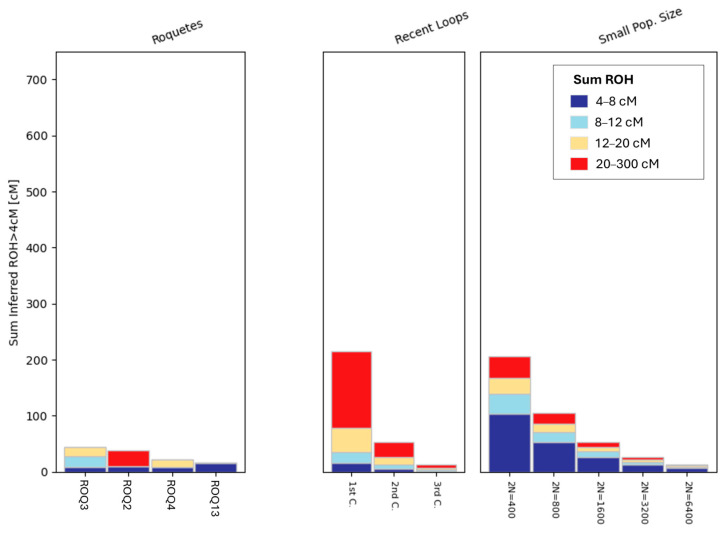 Figure 2
Figure 2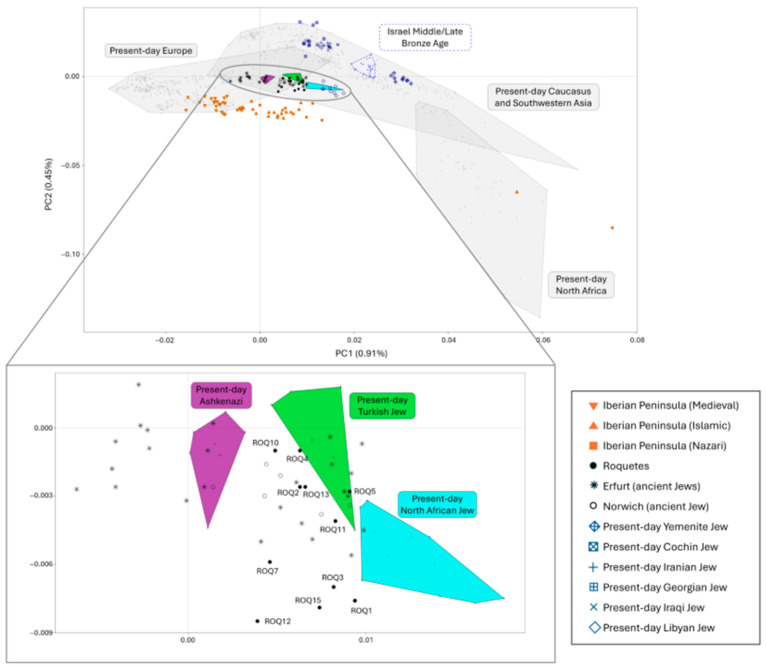 Figure 3
Figure 3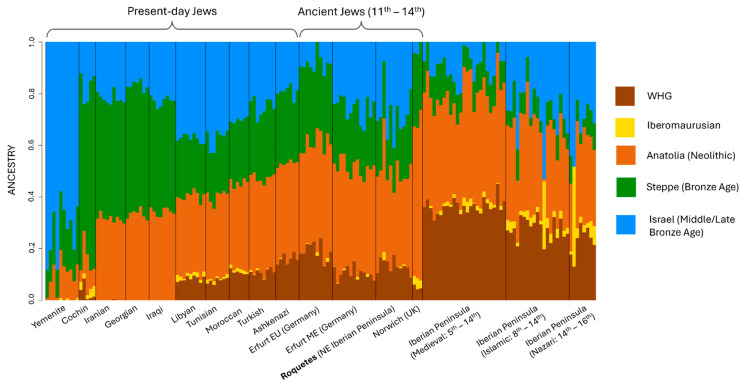 Figure 4
Figure 4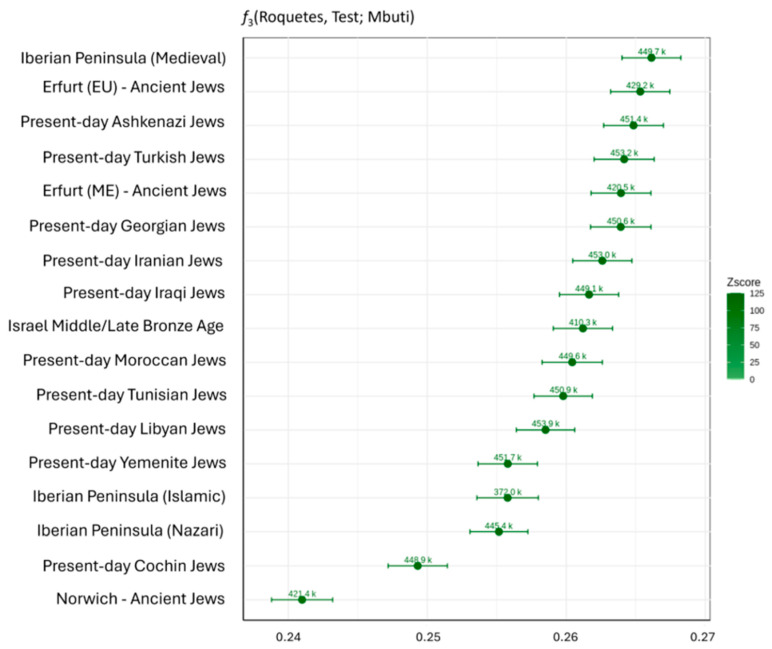 Figure 5
Figure 5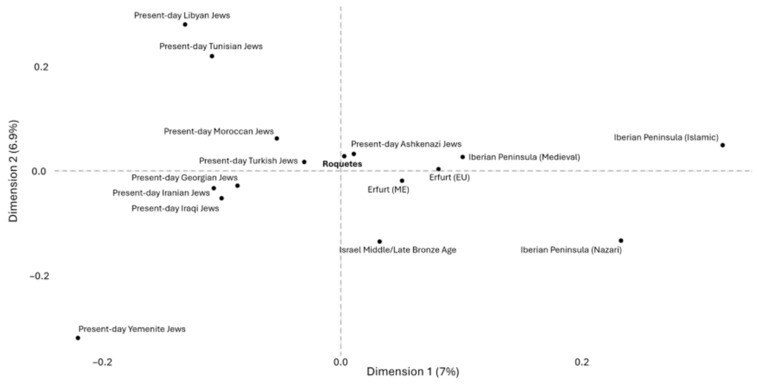 Figure 6
Figure 6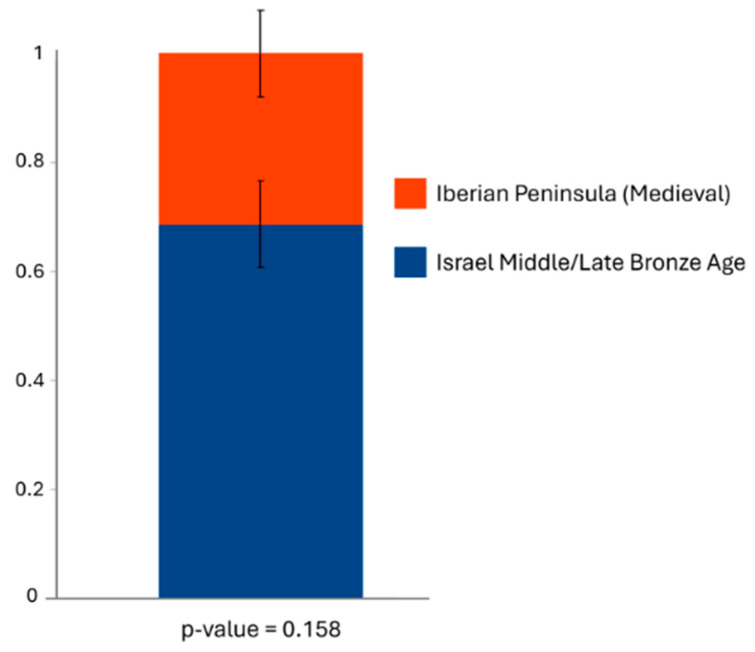 Figure 7
Figure 7- —Department of Culture of the Government of Catalonia
- —Generalitat de Catalunya
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsForensic and Genetic Research · Medieval and Classical Philosophy · Genetic diversity and population structure
1. Introduction
The Jewish people are an ethnoreligious group that traces their origins back to the Late Bronze Age (approximately 1200 BCE) [1] in the Eastern Mediterranean, particularly in the region corresponding to Canaan territory, part of which corresponds to modern-day Israel [1,2,3]. Over the following millennia, successive migratory and exilic movements—both voluntary and forced—gave rise to what is known as the Jewish Diaspora [4]. One of the earliest mass displacements occurred around 720 BCE, with the exile to Mesopotamia after the Neo-Assyrian Empire’s conquest of the northern Kingdom of Israel [5]. Further mass migrations occurred after the destruction of the First Temple by the Neo-Babylonian Empire in 587 BCE [2], and later, after the destruction of the Second Temple by the Roman Empire in 70 CE, when Jewish people started to disperse across the Mediterranean and into Northern Europe [1]. The Diaspora is responsible for the formation of various Jewish communities, two of the most significant being Ashkenazi, corresponding to the Jewish people of Northern Europe, and Sephardic, corresponding to the Jewish people from the Iberian Peninsula [6,7].
Jewish settlement in the Iberian Peninsula is generally dated to the period following the destruction of the Second Temple in 70 CE [1,8]. After the 6th century, with the establishment of Christianity as the main religion, Jewish communities frequently endured social marginalization, legal restrictions, forced conversions, and episodes of expulsion—common phenomena in the European regions where this ethnoreligious group migrated [9,10,11]. During the first period of the Islamic Empire (700–1086 CE), however, the Jewish community experienced a period of relative prosperity, though it —like the Mozarab population (Christians who maintained their religion)—still faced social and institutionalized discrimination. Members of the Jewish community participated in commerce, finance, medicine, and some even partook in governmental structures within the Muslim courts [8,9,12].
This relative stability began to erode with the arrival of the Almoravids in 1086, whose strict orthodoxy curtailed previous tolerance toward dhimmis (non-Islamic population protected under a subordinate legal status within Islamic rule), leading to Jewish emigration to northern Christian kingdoms and forced conversions. The situation worsened under the Almohads in the mid-12th century, who imposed severe restrictions. Many Jews converted under duress while secretly maintaining their faith, and others fled north, effectively ending Jewish cultural prosperity in Al-Andalus [8,9]. In contrast, the Jewish communities of northeast Iberia, the Crown of Aragon, developed their own distinctive trajectory. These communities became centers of intellectual and economic activity, shaping the cultural and religious life of medieval Catalonia [13].
The discrimination against the Jewish community severely worsened in the 14th century [14]. The combined effects of the Black Death (1347–1351), recurrent famines, violent conflicts, and economic crisis generated a climate of insecurity that translated to extended antisemitism across Europe [4,9]. Jewish people were scapegoated for the plague and accused of hoarding wealth, leading to pogroms, violent assaults on Jewish quarters, throughout the continent [4,9,12]. In the Iberian Peninsula, the hostilities culminated in 1492, with the implementation of the Edict of Expulsion, which forced all unconverted Jews to leave the Crowns of Castile and Aragon (present-day Spain) [15]. The exiled Jewish population from the Iberian Peninsula largely fled to the Mediterranean basin, settling in regions such as North Africa and the Ottoman Empire (including present-day Turkey), being referred to as Sephardic Jews [7,9].
The expulsion of the Jewish people from the Iberian Peninsula represented the conclusion of this severely antisemitic climate, characterized by numerous episodes of extreme violence. Within the Crown of Castile, the 1391 Seville pogrom spread across the peninsula, resulting in the death of thousands of Jews and mirroring earlier outbreaks of violence, such as those that occurred in the Crown of Aragon in 1348 following the arrival of the Black Death [9]. In the Crown of Aragon (eastern Iberian Peninsula), the most affected cities by the pogroms associated with the pandemic were Barcelona, Cervera, Lleida, Girona, and Tàrrega [9]. Historical sources chronicle a massacre in Tàrrega in 1348, during which armed attackers entered the Jewish quarter and assaulted houses, destroyed deeds, and caused the death of approximately 300 people [16].
In 2007, six mass graves dating back to the mid-14th century were found in the Roquetes necropolis (Figure 1) in Tàrrega—part of the Crown of Aragon—suggested by historical data to have belonged to the medieval Jewish community. A total of 69 individuals were recovered from the common graves, although it is suspected that the unexcavated areas of the necropolis hold more mass graves [17]. The typology of the funerary ritual indicated the possibility of a synchronic mass death event. Furthermore, the presence of perimortem traumatisms in at least one third of the individuals, mostly affecting the cranial region, indicated an intentional cause of death, strongly supporting the hypothesis that these burials belonged to victims of the medieval Tàrrega pogrom [18].
Recent advances in paleogenomics provide powerful tools to investigate such historical events and the populations affected by them. Due to centuries of relative social isolation, Jewish communities display genetic continuity between ancient and modern populations [19,20,21,22]. Two recent studies have analyzed genome-wide data from medieval Jewish cemeteries: one from Norwich, England [21]—where individuals were victims of an antisemitic massacre—and another from Erfurt, Germany [20]. Both studies revealed populations with mixed Middle Eastern and European ancestry, as well as genetic continuity between the studied medieval Jewish individuals and present-day Ashkenazi Jews.
This study aims to analyze the genetic profiles of individuals recovered in the necropolis of Roquetes. By applying advanced paleogenomic tools, we seek to address the current absence of genomic data on medieval Iberian Jewish populations and to provide the first genomic information for these communities, which remain completely unknown from a genomic perspective.
2. Materials and Methods
2.1. Sample Preparation, DNA Extraction, and Quantification
Bones from sixteen individuals (Table 1), from different osteological sex and age groups, exhumed from all six mass graves at the Roquetes necropolis (Tàrrega, Catalonia, Spain) were selected (Supplementary Material, Table S1). The outer surface of each bone was initially cleaned to remove dirt and then polished with a milling cutter to eliminate the outermost layers. A 1 cm section was selected and pulverized using a hammer for subsequent DNA extraction.
For the petrous bones, 3D photogrammetry models were created prior to pulverization, ensuring the creation of precise digital replicas that capture their detailed morphology.
Around 100–150 mg of bone powder was used for DNA extraction in a 48 h incubation with a solution of Proteinase-K and EDTA, and eluted in 60 µL of EBT as in Vinueza-Espinosa et al. (2020) [23]. Extracted DNA was purified as described by Vinueza-Espinosa et al. (2020) [23], using the High Pure Viral Nucleic Acid Large Volume Kit (Roche), a silica in HE-membrane column-based method. Blank controls were processed with each extraction. A quantitative PCR (qPCR) of a small mitochondrial DNA fragment was performed to confirm the absence of contamination during DNA extraction. Qubit^®^ dsDNA HS Assay Kit (ThermoFisher Scientific, Waltham, MA, USA) was used to quantify each sample, and Bioanalyzer^®^ Agilent High Sensitivity DNA Kit (Agilent Technologies, Barcelona, Spain) was employed to obtain fragmentation profiles.
All sample preparation and processing steps were conducted in the Biological Anthropology Unit’s dedicated clean ancient DNA laboratory at the Universitat Autònoma de Barcelona (UAB).
2.2. Library Preparation, Sequencing, and Enrichment
Twenty µL of extracted DNA was used to perform single-strand libraries with the Santa Cruz Reaction (SCR) protocol [24]. For each sample, 50 µL of the library was recovered and purified using the MinElute PCR Purification Kit (Qiagen, Barcelona, Spain). The indexing PCR cycle number was estimated by qPCR using QuantiNova^TM^ SYBR Green PCR Kit (Qiagen). Overall, 22.5 µL of the library was indexed using the KAPA HiFi HotStart Uracil+ kit (Roche) with the following program: initial denaturalization 3′ at 95 °C; X cycles 20″ at 98 °C, 30″ at 60 °C, 30″ at 72 °C; final incubation 7′ at 72 °C, 4 °C forever. Indexed P7 and indexed P5 primers for the Illumina platform, similar to Kircher et al. (2012 [25]) were used to perform unique dual-index sequencing.
Finally, the amplified libraries were purified using SPRIselect beads (Beckman Coulter, Brea, CA, USA) and quantified with the Qubit^®^ dsDNA HS Assay Kit (ThermoFisher Scientific). Bioanalyzer^®^ Agilent High Sensitivity DNA Kit (Agilent Technologies) was employed to obtain the library size profile. The amplified and indexed libraries were then pooled in equimolar amounts and sequenced with Illumina technology.
Libraries were enriched with the Ancient Human DNA Target Enrichment kit (Twist Biosciences, San Francisco, CA, USA), which includes DNA baits targeting the same ~1.3 million nuclear SNPs from the 1240k panel [26], following the manufacturer’s protocol. After, libraries were pooled in equimolar amounts and sequenced with Illumina technology in a NextSeq 1000 sequencer (Illumina, San Diego, CA, USA).
2.3. aDNA Data Processing and Authentication of Whole Genome Sequencing
After demultiplexing based on a dual-unique index combination developed by the sequence provider, raw sequencing data were processed with the nf-core/eager pipeline v2.4.7 [27,28]. Adapters were removed with AdapterRemoval v2.3.2 [29], and sequences shorter than 30 bp were eliminated. Then, sequences were mapped against the Human Reference Genome GRCh37 (hg19, GenBank accession GCA_000001405.1) with BWA v0.7.17-r1188v [30], aln, and samse/sampe commands. Parameters “-l 1024-n 0.01-o 2” were used, following recommendations from Oliva and colleagues [31,32]. SAMtools v1.12 [33] was used to generate sorted BAM files with all the reads with a mapping quality of over 20. Duplicates were removed with the MarkDuplicates tool from Picard v2.26.0 software [34]. Damage levels were assessed with PMDTtools v0.50 [35]. Finally, seven bases were trimmed at the start/end of the reads with BamUtils v1.1.2 [36] to avoid ancient damage in final genotypes. The quality of FASTQ files was evaluated with FastQC v0.11.9 [37], and the quality of the BAM file was analyzed with QualiMap v2.2.2-dev [37].
2.4. Sex Determination, Contamination, and Genotyping
Genetic sex was determined based on the relative coverage of specific X and Y chromosomes’ SNPs using the tool Sex.DetERRmine.py v1.1.2 [38], implemented in the nf-core/eager pipeline [27,28].
Ancient DNA authenticity was evaluated for all samples using several criteria [39]: A rate of cytosine deamination at the terminal nucleotide above ~10%, since non Uracil-DNA glycosylase treatment was used; a ratio of Y to combined X + Y chromosome sequences below 0.03 or above 0.35, where intermediate values are indicative of the presence of DNA from at least two individuals of different sex; and an upper bound rate for the 95% confidence interval for the rate to the consensus mitochondrial sequence that exceeds 95%, as computed using contamMix-1.0.10 [40]. Additionally, mtDNA was manually reviewed for inconsistencies. We directly discard samples with a rate of cytosine deamination at the terminal nucleotide lower than 5%, or samples with at least two signals of contamination.
Trimmed BAM files were used to call genotypes with pileupCaller, which generates pseudo-haploid genotypes by randomly choosing one allele at every SNP position. The panel with 1240k nuclear SNPs used in ancient population genomics was used [26].
2.5. Mitochondrial and Y Chromosome Haplogroups Assignment
Mitochondrial reads that had already been mapped with the mitochondrial DNA for the hg19 genome were realigned against the revised Cambridge Reference Sequence (rCRS, GenBank accession NC_012920.1) [41] with nf-core/eager pipeline v2.4.6 [27,28]. CircularMapper v1.0 [42] was used to avoid the problems associated with the circularity of the mtDNA molecule. Genotypes were called with freebayes v1.3.6 [43] and vcflib v1.0.3 [44]. Haplogrep 3 v3.2.1 [45] was used to call mitochondrial DNA haplogroups. SNPs were manually double-checked, visualizing alignments with the Integrative Genome Viewer (IGV) v2.11.2 [46]. Results were considered reliable at a coverage of 3X after visual confirmation. In the low-coverage regions, the damage, mutational, and heteroplasmic profiles [47,48,49,50] were examined to improve credibility.
For males, Yleaf v3.2.1 [51] with parameters-r 2 (minimum number of reads for each base), -q 30 (minimum quality for each read), -b 60 (minimum of a base result for acceptance), -dh (draw the haplogroups in the haplogroup tree), and -p (search for private mutations), was used to determine the Y-chromosome haplogroup. The trimmed BAM files were used.
2.6. Genetic Relatedness
Autosomal SNPs were used to define first- and second-degree relatedness among pairs of individuals based on their mismatch rate (PMR), using the tool BREADR v1.0.2 [52]. Specifically, the pairs of individuals were categorized as “Same/Twins”, “First-degree”, “Second-degree”, or “Unrelated” (in the first or second degree), and only pairs with at least 500 shared SNPs were trusted.
2.7. Principal Components Analysis (PCA)
Newly reported samples were merged with previously published datasets of ancient and modern individuals from the Allen Ancient DNA Resource (AADR) [53]. Only one of the individuals was considered when there was a first- or second-degree of relatedness. Samples from Human Origins [54] were also included for principal component analysis (PCA). Ancient European Jewish samples from Norwich, UK [21] and Erfurt, Germany [20] were compiled and added. Samples were labeled for each analysis as indicated in the Supplementary Material (Table S2).
The PCA was performed using the smartpca software from EIGENSOFT v8.0.096,97 with the “lsqproject” and “shrinkmode” options activated (YES) and a list of modern populations from Europe, North Africa, Caucasus, and Southwestern Asia (Supplementary Material, Table S2). All ancient individuals, including those from the medieval period of the Iberian Peninsula, were projected onto PC1 and PC2.
2.8. f-Statistics
The f-statistics were performed with ADMIXTOOLS v5.1 (https://github.com/DReichLab/AdmixTools, accessed on 28 October 2025) and plotted with R v4.4.2 [55] using the ggplot2 v3.5.1 package [56].
Outgroup f3-statistics, computed using qp3Pop with the ‘inbreed’ parameter activated, measured the shared genetic drift between two populations (A and B) relative to an outgroup (C), providing a similarity measure: higher values indicate greater genetic similarity between A and B. In this study, we calculated outgroup f3-statistics in the form f3 (Roquetes, Test; Mbuti), where the test populations were present-day and ancient Jewish populations and non-Jewish Medieval populations from the Iberian Peninsula, with Mbuti, a present-day Sub-Saharan hunter-gatherer population from the Democratic Republic of Congo, being used as the outgroup.
Moreover, for each present and ancient Jewish population, and for the non-Jewish Medieval populations from the Iberian Peninsula, their genetic similarity was quantified using the outgroup f3-statistic of the form f3 (Pop1, Pop2; Mbuti), calculated with qp3Pop from ADMIXTOOLS v5.1 (https://github.com/DReichLab/AdmixTools, accessed on 28 October 2025). The f3 results were converted into a pairwise distance matrix where each cell represented 1-f3 (Pop1, Pop2; Mbuti), such that smaller values corresponded to higher shared genetic drift. The resulting matrix was symmetric, and diagonal values were set to zero to preserve matrix completeness. Multidimensional scaling (MDS) was then performed using the classical metric scaling function “cdmscale” in R v4.4.2 [55] (base stats package), with k = 2 dimensions and otherwise default parameters. The eigenvalues returned by “cdmscale” were used to calculate the proportion of variance explained by each of the first two MDS dimensions. The MDS coordinates were visualized using ggplot2 v3.5.1 [56].
2.9. Admixture Modeling
Related individuals were excluded from admixture modeling analyses. ADMIXTURE v1.3.0 was used to define the main genetic cluster profiles [54]. Five populations, presenting differential relatedness between them and the Roquetes population, were selected as fixed reference groups of ancestry (k = 5) to implement a supervised ADMIXTURE model [54]:
Western Hunter-Gatherers: a group of Mesolithic hunter-gatherers who scattered over western, southern, and central Europe.
Iberomaurusian: a Late Pleistocene population from Morocco, accounts for potential North African-related ancestry to improve differentiation between southern European, Levantine, and North African genetic components.
Anatolia_Neolithic: population of farmers that scattered from Anatolia to West Eurasia around 7000BC.
Russia_Samara_Early Bronze Age_Yamnaya: a population, associated with the Yamnaya culture, originated in the Samara Bend of present-day Russia, dating from 5000 to 4000 BP. They present both an Eastern hunter-gatherer component and a Caucasus hunter-gatherer component.
Israel_Middle Late Bronze Age: population from Canaan, territory of present-day Israel, dating from the Middle Late Bronze Age (MLBA), the period of time associated with the emergence of Judaism.
The model was replicated twenty times, and the results were merged with Pong v1.5 [57].
We modeled ancestry using qpAdm from the ADMIXTOOLS v5.1 package (https://github.com/DReichLab, accessed on 5 November 2025), enabling “allsnps: YES” to maximize the number of SNPs used (and thus the power to reject implausible models). We follow the terminology and framework of Harney et al. (2021) [58] in which qpAdm evaluates whether a target population can be represented as a mixture of one or more Source populations, using a panel of reference populations to differentiate shared genetic drift among the source population groups.
In qpAdm, the reference populations serve as statistical outgroups: they are selected to be differentially related to the source populations to provide leverage to separate their ancestry contributions. Although they are not phylogenetic outgroups in the strict sense, they function as outgroups within the qpAdm framework by anchoring deep ancestry components and breaking symmetries among the source groups [58].
Following Harney et al. (2021) [58], we interpret qpAdm p-values as model diagnostics: models with p > 0.05 are termed plausible, whereas models with p ≤ 0.05 are implausible and are therefore rejected. We describe models as plausible/rejected under the qpAdm framework.
We first evaluated one-way models to test whether the target could be adequately modeled as deriving entirely from a single source. Conceptually, a plausible one-way model indicates that the target is indistinguishable from forming a clade with that source relative to the reference panel; it does not imply admixture, but rather identity or near identity of ancestry profiles in the qpAdm sense. We then tested two-way models to evaluate whether the Roquetes population could be explained as a mixture of Canaan (labeled as Israel MLBA) individuals and non-Jewish medieval Iberian populations.
For all the models applied here, we used a set of 4 outgroups (reference populations) [58]:
Mbuti, a present-day Sub-Saharan hunter-gatherer population from the Democratic Republic of Congo, serves as a deep non-West Eurasian outgroup that helps distinguish West Eurasian ancestry from older African lineages without introducing the long-branch biases associated with chimpanzee outgroups.
Ust’-Ishim, an Upper Paleolithic individual from western Siberia (~45 ka), represents an early non-African lineage and provides resolution among West Eurasian populations before their subsequent diversifications.
Iberomaurusian, a population previously presented.
Villabruna, a Mesolithic hunter-gatherer sample from northern Italy, represents the Western European hunter-gatherer (WHG) ancestry component and provides sensitivity to differential relatedness among European-derived populations.
2.10. Runs of Homozygosity (ROH)
To assess demographic and inbreeding patterns in the population, we examined runs of homozygosity (ROH), defined as stretches of the genome that lack genetic variation and thus reflect the coinheritance of identical haplotypes from related parents. Software hapROH v0.64 [59] was used with default settings to identify runs of homozygosity within the genome of individuals with more than 400,000 autosomal SNPs covered from the 1240k panel [26]. Following Ringbauer et al.’s (2021) [59] indications, ROH were categorized in the following longitude intervals (in centimorgans-cM): 4–8 cM, 8–12 cM, 12–20 cM, and 20–300 cM.
3. Results
3.1. Quality of the Samples
Overall, it was possible to sequence samples of most of the individuals, and only one sample presented signs of being contaminated. Twelve out of the sixteen individuals available presented enough short fragments (50–220 bp) in the extracted DNA for generating single-strand libraries (Supplementary Material, Table S1). The library corresponding to ROQ 8 was discarded for being considered contaminated, as it presented 4.30% of C-to-T changes at the terminal nucleotide (Supplementary Material, Table S3). The remaining eleven libraries presented no signs of contamination: a rate of cytosine deamination at the terminal nucleotide above ~10%, indicative of post-mortem damage expected in ancient DNA; a ratio of Y to combined X + Y chromosome sequences consistent with the presence of one single individual; and a confidence rate for the contamMix-1.0.10 [40] (an upper bound rate for the 95% confidence interval for the rate to the consensus mitochondrial sequence that exceeds 95%) (Supplementary Material, Table S3). Most of these samples (ROQ1, ROQ5, ROQ7, ROQ10, ROQ11, ROQ12, and ROQ13) presented levels of endogenous DNA below 0.5%. Three samples (ROQ2, ROQ3, and ROQ4) presented endogenous DNA percentages between 5 and 20% (Supplementary Material, Table S3), and one (ROQ15) had a percentage of endogenous DNA of 63.77% (Supplementary Material, Table S3), yielding a considerable number of SNPs (121,312 SNPs) from the 1240k panel [26].
For the ten samples with a percentage of endogenous DNA lower than 20% (Supplementary Material, Table S3), DNA capture procedures were applied, substantially increasing the proportion of endogenous DNA (Supplementary Material, Table S4). A minimum of approximately 50,000 SNPs was recovered per captured sample, with several achieving much higher values—for instance, ROQ2 and ROQ4 yielded over 990,000 and 1,000,000 SNPs, respectively (Supplementary Material, Table S4).
3.2. Genetic Sex, Genetic Relatedness, and Endogamy
The genetic sex was estimated for the eleven individuals, obtaining four males and seven females. BREADR genetic relatedness analyses revealed no first- or second-degree relationships between the analyzed individuals from Roquetes (Supplementary Material, Table S5).
Results of the analysis of runs of homozygosity (ROH) revealed the presence of some medium and long homozygous regions (Figure 2 and Figures S1–S4), and put the focus on individual ROQ2, which exhibited long ROHs (20–300 cM), indicating background endogamy.
3.3. Uniparental Markers
High mitochondrial diversity was identified, with ten different haplogroups in twelve unrelated individuals (Table 2 and Supplementary Material, Table S6). The two repeated haplogroups (L2a1c + 16129 and H1bo) belonged to individuals without evidence of close genetic relatedness (Supplementary Material, Table S5), despite sharing their mitochondrial haplotypes (Supplementary Material, Table S6).
The mtDNA haplogroups identified in the Roquetes individuals (Table 2) were contextualized by assessing their presence in ancient populations using the Allen Ancient DNA Resource (AADR v62.0) [53] and by evaluating their occurrence in present-day Jewish populations based on published data [60]. Several haplogroups detected in Roquetes have been previously reported in ancient Jewish contexts, including K1a1b1a and U5a2b, both identified among medieval Jews from Erfurt, as well as U5a2b2a in modern Ashkenazi populations (Table 2). Other haplogroups, such as M1a1b1c, J1c1, R0a4, and H1bo, have not been directly observed in the AADR database but are present in modern Jewish populations, particularly among Ashkenazi and Sephardic groups (Table 2). In some cases, closely related parent haplogroups (e.g., M1a1b1 or H20a) have been identified in ancient individuals from the Iberian Peninsula, the eastern Mediterranean, or the Near East, suggesting broader Mediterranean or Near Eastern maternal lineages (Table 2).
The Y-chromosome haplogroups identified in the male individuals from Roquetes (Table 3 and Supplementary Material, Table S10 and Figures S5–S8) were contextualized using the Allen Ancient DNA Resource (AADR v62.0) [53]. The detected lineages (J, E, and G) show diverse geographic and temporal distributions in the ancient record. Haplogroup J2a2a (ROQ1) and E1b1b (ROQ2) have broad distributions in the Mediterranean and Near East, with E1b1b also reported in medieval Jewish populations from Norwich and Erfurt (Table 3). The G1a1a lineage identified in ROQ4 has been documented in Chalcolithic and Bronze Age Iran and in historical-period Turkey, suggesting an eastern origin (Table 3). Haplogroup E2a (ROQ7) is rare in ancient DNA datasets and has been reported in a limited number of ancient and present-day individuals from eastern Africa (Table 3).
3.4. Exploratory Data Analysis: PCA and ADMIXTURE
To investigate the genetic profile of the Roquetes population, we merged our dataset with a collection of present-day populations and medieval Jewish individuals from Erfurt and Norwich (see Supplementary Material, Table S2 for labels). These individuals were projected onto the first principal components (PC1 and PC2) of a Principal Component Analysis (PCA) based on present-day populations from Europe, North Africa, the Caucasus, and Southwestern Asia (Figure 3). After filtering, 553811 autosomal SNPs from the 1240k panel [26] remained for analysis. The Roquetes individuals overlapped with present-day Turkish Jews (Sephardic), as well as with the ancient Jewish individuals from Erfurt and Norwich (Figure 3). Conversely, Roquetes individuals were genetically distant from the non-Jewish Iberian medieval population (Figure 3).
ADMIXTURE analyses suggested the Roquetes population’s ancestry could be modeled as a mixture of Western hunter-gatherer (WHG), Steppe Bronze Age, and Anatolian Neolithic components, together with ancestry related to populations from Canaan (territory of present-day Israel in the MLBA), and a minor Iberomaurusian contribution for some individuals (Figure 4). This ancestral profile closely resembles that observed in the Jewish individuals from medieval Erfurt.
3.5. Formal Tests of Genetic Affinity and Admixture
In order to identify the closest genetic matches for the medieval Jews from the Roquetes and further assess the PCA and ADMIXTURE-based observations, we first performed outgroup f3-statistics in the form f3 (Roquetes, Test; Mbuti) using individuals with more than 50,000 autosomal SNPs (Figure 5 and Supplementary Material, Table S13). Our results indicated that Roquetes individuals shared higher genetic drift with non-Jewish non-Islamic Medieval population from the Iberian Peninsula and Medieval Ashkenazi Jews from Erfurt (Germany), as well as with present-day Ashkenazi and Sephardic (Turkey) Jewish populations (Figure 5 and Supplementary Material, Table S13). In contrast, lower genetic affinity was observed with the Islamic and the Nazari (the last Islamic dynasty of Al-Andalus [9]) individuals from the Iberian Peninsula and Medieval Norwich (UK) Jews, as well as with other present-day Asian Jews, such as Cochin (India) or Yemenite (Yemen) Jews (Figure 5 and Supplementary Material, Table S13).
The MDS based on pairwise 1–f3 distances largely confirms the affinities observed in the PCA and outgroup f3 analyses (Figure 6 and Supplementary Material, Table S14 and Figure S9). Roquetes individuals clustered closely with present-day Ashkenazi Jews and Sephardic Jews from Turkey, as well as with the medieval Ashkenazi individuals from Erfurt (Germany), indicating elevated levels of shared genetic drift among these groups. Non-Jewish non-Islamic Medieval population from the Iberian Peninsula occupied a nearby position in the MDS space, consistent with the high f3 values observed in pairwise comparisons.
To formally test whether the genetic affinities inferred from PCA, ADMIXTURE, outgroup f3-statistics, and MDS could be explained by specific ancestry models, we performed qpAdm analyses using one- and two-way admixture scenarios, always using the same group of outgroups (reference populations). For the one-way models, we tested whether the Roquetes population could be plausibly modeled from a single source population, including non-Jewish non-Islamic medieval individuals from the Iberian Peninsula, Islamic and Nazari medieval Iberian populations, medieval Jewish individuals from Erfurt —separated into individuals of European (EU) and Middle Eastern (ME) genetic origin—, medieval Jewish individuals from Norwich, Canaan (territory of present-day Israel in the MLBA) Age individuals, and present-day Jewish populations from Turkey (Sephardic), Ashkenazi, and Morocco (Supplementary Material, Table S15). Among these models, only the Erfurt medieval Jewish population with Middle Eastern genetic affinity yielded a plausible fit (p = 0.098), whereas the other one-way models were rejected since the p-value was lower than 0.05 (Supplementary Material, Table S15).
We tested whether the Roquetes populations could be modeled as a mixture of individuals from Canaan (labeled as Israel Middle/Late Bronze Age) and medieval populations from the Iberian Peninsula, including non-Jewish non-Islamic, Islamic, and Nazari groups, using two-way models (Supplementary Material, Table S15). Among the tested models, only the combination of Canaan individuals (labeled as Israel Middle/Late Bronze Age) and the non-Jewish non-Islamic medieval population from the Iberian Peninsula provided a plausible fit (p-value = 0.158), with estimated ancestry proportions of 0.686 and 0.314, respectively (Figure 7 and Supplementary Material, Table S15).
These results indicate strong genetic affinity between the Roquetes population and the Erfurt medieval Jewish population with Middle Eastern genetic affinity, and provide a plausible model of ancestry by combining Canaan individuals and the non-Jewish non-Islamic medieval population from the Iberian Peninsula.
4. Discussion
We analyzed genome-wide and uniparental data from 16 individuals exhumed from the Roquetes necropolis (Tàrrega, Catalonia, Spain), including the six mass graves identified previously, securely dated to the mid-14th century, and archeologically interpreted as a single catastrophic episode [17,18]. While the sample covers only a subset of the excavated individuals, multiple lines of evidence converge on a non-familial burial event: we find no first- or second-degree genetic kinship among analyzed individuals, and a significant fraction of the broader burial assemblage exhibits perimortem cranial trauma [18]. Genomic data cannot identify the cause of death; however, it revealed affinity with ancient and present-day Jewish communities that, when integrated with the archeological context, dating, and historical sources for 1348, reinforce the attribution to the Tàrrega pogrom.
Following ancient Jewish law, religious affiliation is maternally assigned [70]; consequently, mtDNA variation can offer complementary insights into Jewish community histories and origins. The Roquetes individuals show high mitochondrial diversity (haplogroups H, J, K, L, M, R, U), contrasting with the narrow mtDNA spectra documented in medieval Ashkenazi communities at Erfurt (Germany), where strong founder effects and drift operated [20], and at Norwich, where elevated levels of consanguinity could also have influenced the genetic drift [21]. The pattern observed in Roquetes aligns more closely with Sephardic-descended communities [71,72], where modern Jewish populations that received exiles from Iberia—Bulgaria, Turkey, and Morocco—display substantial haplogroup diversity, in contrast to other Jewish groups (e.g., Azerbaijan, Georgia, Libya, Mumbai (India), and Belmonte (Portugal)) that exhibit bottleneck signatures [19,71,73,74].
Notably, we observe haplogroups with strong Jewish associations or prior medieval occurrences—e.g., K1a1b1a (common in Ashkenazi; present in Erfurt), U5a2b, and M1a1b1c (present in Erfurt), with broader presences of H1 lineages, R0a, J1 subclades in Jewish and non-Jewish populations [20,21,60,75]. Their occurrence in Roquetes does not imply exclusivity to Jewish groups but, in combination with autosomal affinity patterns, highlights that Iberian Jewish communities maintained higher mtDNA heterogeneity than contemporaneous Central/Northern European groups [19,74,76].
Among the four males, we detect J2 (J2a2a1~*), E (E-CTS9507; E-Y231455), and G (G-PH1944/FT19393) haplogroups. These lineages have deep Neolithic–Bronze Age roots in the Eastern Mediterranean/Levant and adjacent regions and are frequent in Jewish diasporic contexts [19,66,77,78]. The recurrence of E1b1b-lineages in medieval Jewish cemeteries (Erfurt, Norwich) and the presence of J and G among present-day Jewish groups further underscore continuity from Levantine–Mediterranean ancestry sources [20,21,78,79,80]. Given the constraints of coverage and marker density in ancient Y-chromosome inference, subclade resolution should be interpreted cautiously; nonetheless, the observed diversity argues against a tight, highly drifted paternal pool at Roquetes.
PCA places Roquetes in proximity to medieval Jewish samples from Erfurt/Norwich and to modern Sephardic Jews (e.g., Turkey, North Africa), while distinct from non-Jewish medieval Iberians. Supervised ADMIXTURE (k = 5) highlights a combination of Anatolian Neolithic, Steppe Bronze Age, and Canaan (territory of present-day Israel in the MLBA)-related components, with minor Iberomaurusian signals in some individuals, an ancestry palette recurrent in Jewish diaspora datasets [20,22]. Importantly, ADMIXTURE components are not population-specific and can reflect shared deep ancestries across the Mediterranean (e.g., Levantine-related contributions also present in North Africa via Phoenician and later historical contacts). We therefore rely on formal tests for specificity. In this line of evidence, outgroup f3 statistics in the form of f3 (Roquetes, Test; Mbuti) and MDS on 1–f3 distances confirm elevated shared drift with medieval Erfurt Jews and with modern Jewish groups (Ashkenazi; Sephardic from Turkey), and comparatively reduced affinity to Islamic/Nazarí medieval Iberians and to Asian Jewish groups (e.g., Cochin, Yemenite) [20,21]. The apparent genetic distance to Norwich is compatible with its strong drift/consanguinity, which can accentuate separation in low-dimensional summaries [21].
Crucially, qpAdm identifies two-way models that fit the Roquetes population as a mixture of Canaan (labeled as Israel Middle/Late Bronze Age) and non-Jewish non-Islamic medieval Iberian populations (p = 0.158), with point estimates around ~0.69 for Levantine ancestry and ~0.31 for Iberian medieval ancestry. Among one-way models, only the Erfurt subgroup with Middle Eastern affinity is marginally plausible (p = 0.098), consistent with a Levantine-centered core plus Iberian admixture. Together, these results indicate that the Roquetes community preserved a distinctively Jewish genetic signature while incorporating local Iberian ancestry.
Genetic evidence for limited but not zero admixture at Roquetes correlates with historical records: conversions and interfaith relationships occurred across medieval Iberia despite religious proscriptions [81,82,83]. On broader scales, present-day Jewish groups—including Sephardic—harbor substantial European admixture (~30–60%) [22,84,85,86,87,88], and Sephardic ancestry persists detectably in modern Iberians [79,89]. The mtDNA diversity we document aligns with the Sephardic pattern [19] and contrasts with communities shaped by founder effects [19,21].
As mentioned before, kinship analyses show no first- or second-degree relationships, indicating that the mass burials do not reflect family-based interments. Moreover, ROH profiles reveal background endogamy in one case (ROQ2), which exhibits long ROH segments that could be compatible with mating patterns in relatively small or socially insular communities [59]. This mirrors patterns seen in other medieval Jewish contexts yet remains milder than the strong consanguinity reported in Norwich [21].
5. Conclusions
This is the first study ever to report genomic data for medieval Iberian Jews. Across uniparental and autosomal lines, the Roquetes individuals exhibit (i) Eastern Mediterranean/Levantine ancestry consistent with Jewish origins, (ii) mtDNA diversity characteristic of Jewish communities, (iii) paternal lineages common in Jewish diasporas (J2, e.g., (iv) formal models requiring Canaan plus Iberian medieval sources, and (v) endogamy without close kinship. When integrated with the 14th-century context and osteological evidence of lethal violence, these data support the previous attribution of the mass graves to victims of the 1348 Tàrrega pogrom. Beyond the event attribution, the study broadens the genetic portrait of medieval Iberian Jewish communities, underscoring their heterogeneity and regional interactions within Iberia.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Goodman M. A History of Judaism Princeton University Press Princeton, NJ, USA 2018
- 2Lederhendler Grabbe L.L. A History of the Jews and Judaism in the Second Temple Period T & T Clark International London, UK 2004
- 3Na’aman N. The Canaanites and Their Land, Ugarit-Forschungen 26; Internationales Jahrbuch für die Alertumskunde Syrien-Palästinas Ugarit Verlag Münster, Germany 1994397418
- 4Ben-Sasson H.H. A History of the Jewish People Ben-Sasson H.H. Harvard University Press Cambridge, MA, USA 1976
- 5Na’aman N. Ancient Israel and Its Neighbors Interaction and Counteraction: Collected Essays Eisenbrauns Ann Arbor, MI, USA 2005 Volume 110.1515/9781575065656 · doi ↗
- 6Frishman A. The Early Ashkenazi Jews: Since Their Settlement in North-West Europe to the First Crusade Hakibbutz Hameuchad Tel-Aviv, Israel 2008
- 7Benbassa E. Rodrigue A. Sephardi Jewry: A History of the Judeo-Spanish Community, 14th–20th Centuries 1st ed.University of California Press Berkeley, CA, USA 2000
- 8Baer Y. A History of the Jews in Christian Spain Jewish Publication Society of America Philadelphia, PA, USA 1961